Looks like no one added any tags here yet for you.
What does KM refer to?
Concentration of substrate needed to reach ½ of the max
or the affinity of the enzyme for the substrate
what kind of relationship does Km have with enzyme affinity
An inverse relationship
The lower the Km value, the higher the affinity
What does a Vmax refer to?
The maximum velocity of the enzyme catalyzing conversion when its saturated; when all active sites are saturated
What Kcat refer to?
Turnover rate of an enzyme; number of substrate molecules being converted into a product by a single enzyme molecule per unit time when the enzyme is fully saturated
TLDR: catalytic efficiency of enzyme
kCat = (vmax/[e])
Higher kcat = more efficient at catalysis
What is the difference between the active site and allosteric site?
Active site - region of the enzyme where the substrate (or competitive/irreversible inhibitor) binds to
Allosteric site - separate site on the enzyme where non-competitive inhibitor can bind to in order to change the conformation of the active site to prevent the substrate binding or being released
How does [E] affect Vmax?
More [E] available = more active sites allows Vmax rate reaction to be higher; vice versa
What is an irreversible inhibition?
Inhibitor is attached to the enzyme’s active site for long periods of time due to covalent bonds
What is the function of penicillin
Penicillin is an antibiotic that is an irreversible inhibitor, it mimics the substrate of glycopeptide transpeptidase (GPT)
GPT catalyzes the final step in the synthesis of the bacterial peptide can cell walls
Function: stops bacteria from growing and eventually dies by targeting the cell wall
Mechanism: penicillin binds to the active site and covalent bonds with a key Ser residue to the bacteria’s active site, resulting in the enzyme being deactivated
What does KM refer to?
when the inhibitor binds to an enzymes active site using weak ionic interactions
How do you calculate Km and Vmax using the lineweaver burk plot?
graph 1/V0 vs 1/s → from this we can calculate:
y intercept: 1/vmax → go to the line that crosses the y-not, read the number and take the inverse
X intercept: -1/km → read where the line crossed the x-int, take the minus inverse of that number
What is competitive inhibition
A type of reversible inhibition that occurs when the inhibitor has the same shape as the substrate, competing with the substrate to bind at the active site
result: only one can bind at the active site at a time, thus catalysis is slowed as there is less ES
What is the effect on competitive inhibition of the Km, Kcat, and Vmax
vmax: No change (because adding enough substrate can outcompete the inhibitor).
Kcat: stays the same because catalytic activity is not impacted, only substrate binding
Km: Increases because more substrate is needed to reach half-maximal velocity (higher KM, the lower the affinity; inverse relationship)
What is the effect on irreversible inhibition of the Km, Kcat, and Vmax
vmax: lowers [E], thus decreases
Kcat: changes functional [E], while [E] stays the same thus it will decrease
Km: does not change; will always have the same affinity
What is non-competitive inhibition
A type of reversible inhibition that binds at the alosteric site (not active site) and inhibits the enzyme
substrate can still bind but no product will be formed
Adding more substrate will not outcompete the inhibitor
What is the effect of non-competitive inhibition on a Vmax, Kcat, and Km
Vmax: decrease because number of active sites available Decreases
Kcat: if vmax decreases, and [E] stays the same but active site decreases, k at will decrease
Km: stays the same because enzyme affinity doesn’t change since it is binding at the aloesteric site
What is a protease
A protease breaks down proteins by cleaving peptide bonds by hydrolysis
What is chymotrypsin
A serine protease that is found in the small intestines.
function: cleaves peptide bonds at the C-terminal after large hydrophobic a.a residues (Tyr, Phe, Trp, Met)
What is the structure of chymotrypsin
Composed of 3 chains (heterotrimer) formed by the cleavage of a single polypeptide
Why is Serine 195 so reactive?
Ser 195 can H-bond to the midsole ring on His57
result: this positions Ser195 and polarizes the hydroxyl group (has more alkoxide character), turning it into a base catalyst (proton acceptor)
It pulls protons from Ser195, making it a better nucleophile
Stabilization by Asp 102
Asp102 helps to position His57 and stabilize the + charge (that develops during proton extraction), this stabilization makes it easier for His57 to deprotonate Ser195
Asp 102 makes His57 more basic, easier to deprotonate Ser195
What amino acids are in the catalytic train
Asp 102
His 57
Ser 195
Ser smith has Asperger’s
What is the first step of the chymotrypsin mechanism?
Substrate binds to chymotrypsin’s active site
the hydrophobic amino acid residue is recognized by the hydrophobic pocket (S1 pocket)
What is the second step of the chymotrypsin mechanism?
Ser 195 does a nucleophilic attack on the carbonyl chain of the substrate
result: proton from Ser 195 is transformed to His 57
How does cooperative binding Works in hemoglobin (full tetramer):
When one oxygen molecule binds to one of hemoglobin’s four subunits, it slightly shifts the structure. This weakens the interactions that stabilize the T-state and makes it easier for the other subunits to bind oxygen. With each additional oxygen molecule binding, hemoglobin shifts further toward the R-state, increasing oxygen affinity for the remaining unbound subunits.
What is the difference and one similarity between chymotrypsin and trypsin
chymotrypsin cuts after large hydropobic groups (Tyr, Trp, Phe, Met)
Trypsin cuts after Arg or Lys
Both are proteases that have a catalytic triad mechanism for cleaving peptide bonds
How is chymotrypsin/trypsin specific?
The S1 pocket determines specificity
What is the S1 pocket
A deep hydrophobic pocket located in chymotrypsin’s active site where the hydrophobic group of a substrate can bind to
Describe the S1 pocket in the serines proteases
chymotrypsin: large hydrophobic group
Trypsin: has a negatively charged Asp at the bottom
Elastase: has 2 Val in the side chain of the S1 pocket, narrowing it (only small hydrophobic residues can fit)
what are the 4 protease groups
serine protease family
ex. thrombin (blood clotting enzyme)
cysteine proteases
use cysteine side chain as a nucleophile
aspartyl proteases
use Asp to make H2O a strong nucleophile
ex. HIV protease
metallaprotease
use a metal ion and H2O as a nucleophile
cut off at hemoglobin
what is metabolism
a highly integrated network of chemical pathways that extract energy from the environment to make energy
what are the 2 types of metabolism
catabolism - reactions that use building block molecules into useful energy
cat = all of benji’s small acts makes me love him more
anabolism - reactions that use energy to break down complex structures into simple ones
what must happen for a pathway to proceed to its final product
the individual reactions must be specific for one set of products
all reactions must be thermodynamically favored (negative delta G)
what are the conditions at the standard free energy change
T = 298 K (25 degrees)
pressure = 1 atm
pH = 7
all [ ] = 1M
what is the most common way to drive cellular endergonic reactions?
couple them to adenosine triphosphate (ATP) hydrolysis
this makes it so the equilibrium shifts to favor the products
what is the structure of adenosine and ATP
adenosine = adenine + ribose (nitrogenous base)
adenosine triphosphate (ATP)
adenine (nitrogenous base)
ribose: a 5-carbon sugar that connects to the phosphate group using phosphodiester bonds
3 phosphate groups: linked by high-energy phosphoanhydride bonds
-
where are the phosphodiester bonds in ATP
These bonds connect the phosphate group to the ribose sugar in ATP (specifically between the alpha phosphate and the 5' carbon of ribose).
These bonds are not considered high-energy like the phosphoanhydride bonds.
why is there so much energy in the phosphodiester bonds of ATP
relief of charge repulsion: the 3 phosphate groups are negatively charged and repel each other. Holding these charges so close together takes a lot of energy, so breaking the bonds relieves this repulsion.
resonance stabilization of pi: the phosphate group is more stable by itself than when it is attached to ATP
ionization of ADP: once ATP is hydrolyzed the products (ADP and Pi) can be more easily stabilized by water molecules through hydration
increase in entropy: the hydrolysis of ATP increases entropy, which is thermodynamically favored
what is phosphoryl transfer
the movement of a phosphate group from one molecule to another molecule
what is phosphoryl transfer potential
the likelihood a molecule can transfer a phosphate based on their ΔG° of hydrolysis of the phosphate
what does a more negative ΔG° mean on the PTP
the more negative the ΔG° = the higher the PTP = really wants to give away the phosphate group
ex. the higher the PTP, the easier it is to phosphorylate something with a lower PTP
what is the ΔG° of ATP synthesis
+30.5
what is the reaction mechanism for the creation of ATP using creatine phosphate
creatine phosphate + ADP ←→ ATP + creatine
what is creatine phosphate
it is used to generate ATP in the muscles during the first few seconds of heavy muscle contraction when [ATP] is low
it bridges the gap until the catabolic pathway kicks in
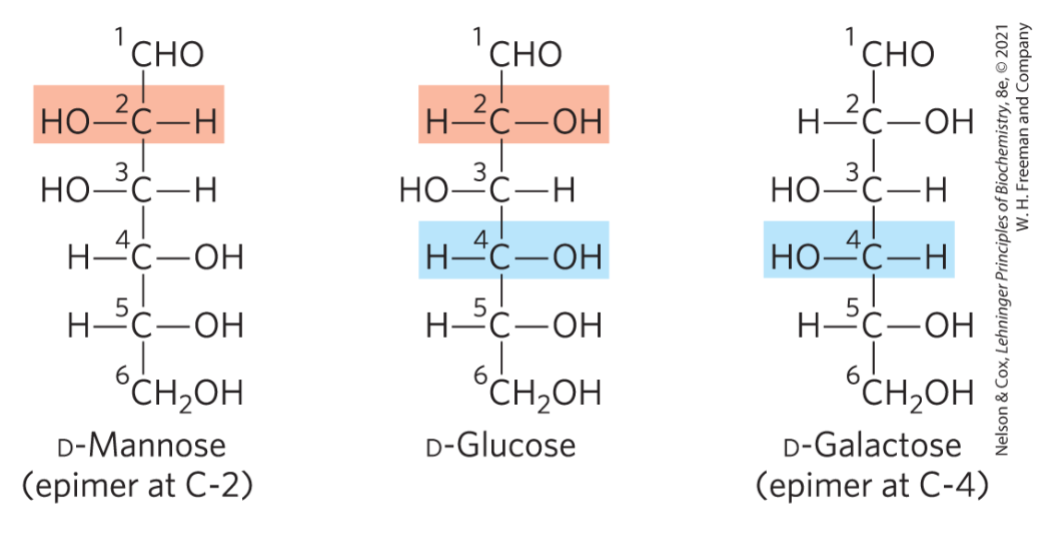
what is the difference between an L or D sugar
D sugar = OH is on the right
L sugar = OH is on the left
how can you switch from a D to L sugar
you need to flip everything
what is an epimer
type of diastereomer that differ only at one chiral carbon
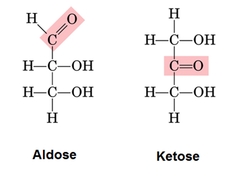
what is the difference between a ketose and aldose
ketose = contains a ketone group
aldose = contains an aldehyde group
what is this?
an epimer
D-glucose & D-galactose
D-glucose & D-mannose
what is an anomer?
when a carbon cyclizes, the new chiral carbon formed is the anomeric carbon
anomer = chirality = has 2 forms
what is the difference between a beta and an alpha anomer
alpha anomer = when the OH is below
alphas are trans
beta anomer = when the OH is above the ring
what is mutarotation
the conversion between alpha and beta anomers of a carbohydrate in aqueous solution, resulting in a change in the optical rotation of the solution.
it is a spontaneous process in glucose and fructose
why does mutarotation occur
Mutarotation occurs because the α and β forms are not stable alone and constantly interconvert through the open-chain form.
what is glycosylation
the process of attaching carbohydrate groups (glycans) to proteins, lipids, or other organic molecules.
what is a glycosidic bond
when a cylic sugar reacts with a glucose/fructose’s hydroxyl or amine group to form a glycosidic bond
A glycosidic bond is a covalent bond that connects a carbohydrate (sugar molecule) to another molecule,
how do you draw glucose
draw a 6-membered ring
an O replaces the top right C
alternate between H and OH
at the end draw the CH2OH
what are the monomers that make up sucrose
glucose (alpha → beta) fructose
what are the monomers that make up lactose
galactose (B1 → C4) glucose
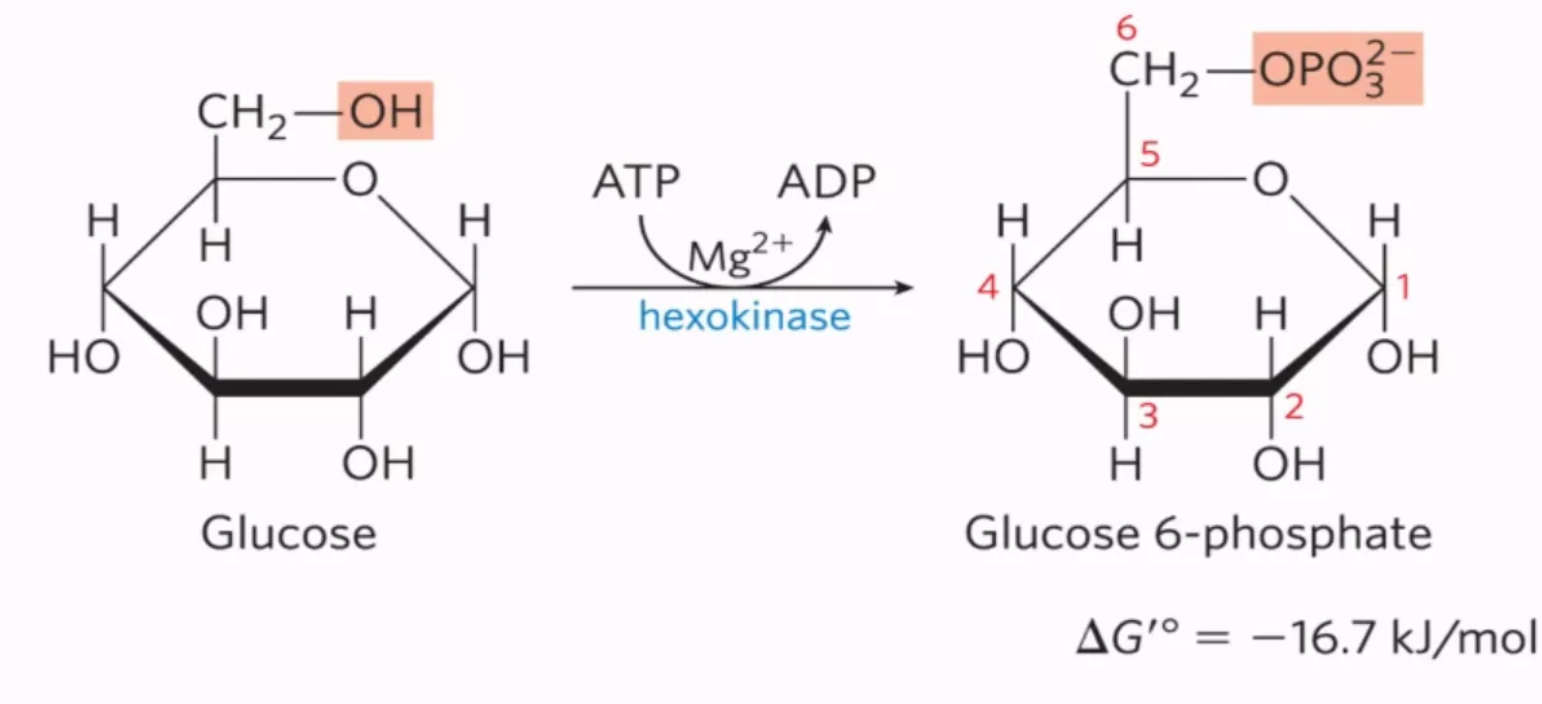
What is the first step of glycolysis, what enzymes mediate it, and why do we do it?
mechanism: glucose is phosphorylated using ATP by hexokinase on C6, turning into glucose-6-phosphate
enzyme: hexokinase
why do we do it? phosphorylation of glucose traps the glucose in the cell (negatively charged, cannot cross the membrane)
what step is this?
#1: phosphorylation of glucose to glucose-6-phosphate by hexokinase
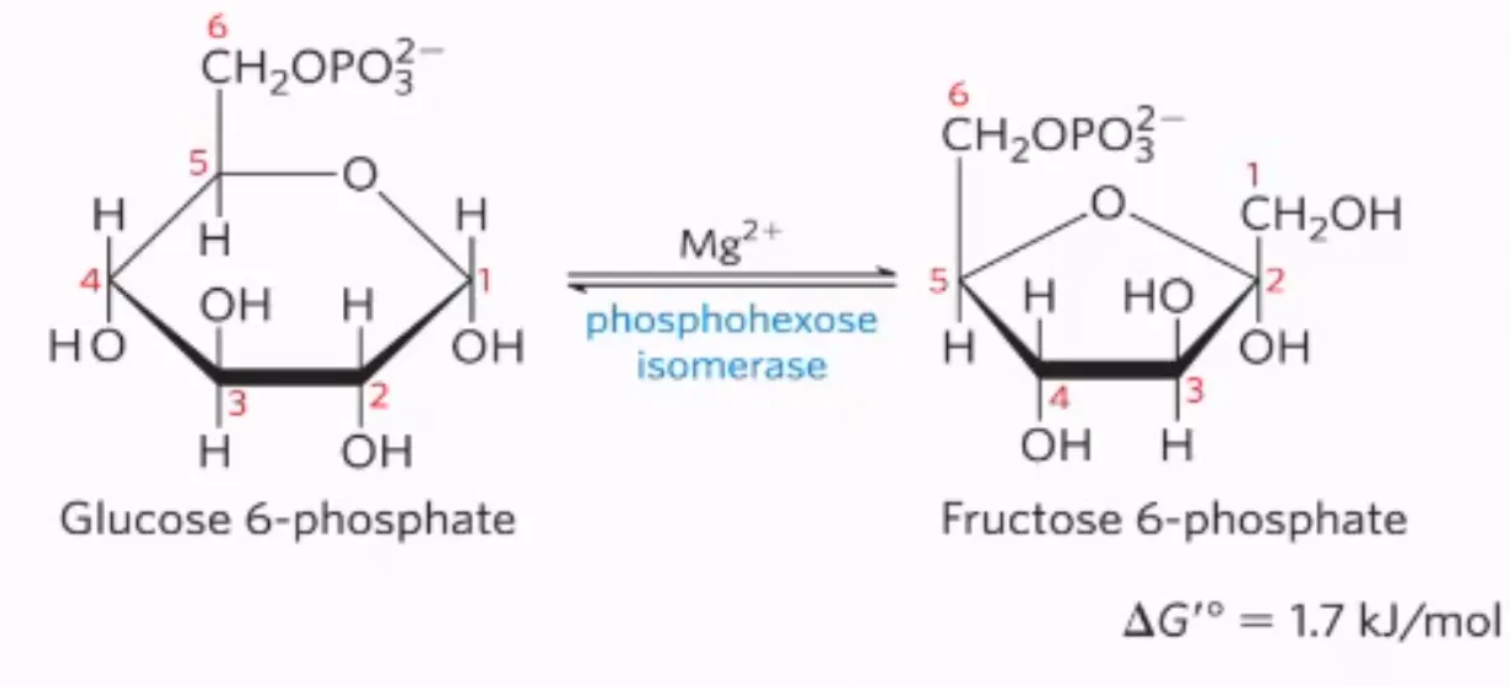
What is the second step of glycolysis, what enzymes mediate it, and why do we do it?
mechanism: glucose-6-phosphate is converted into fructose-6-phosphate by phosphoglucoseisomerase
converts glucose-6-phosphate into an oen chain
repositions the double bond, forming fructose-6-phosphate (open chain)
cycalize to form furanose
enzyme: phosphoglucoseisomerase
why do we do it? fructose is more symmetrical than glucose
what step is this?
#2: isomerization of glucose-6-phosphate into fructose-6-phosphate
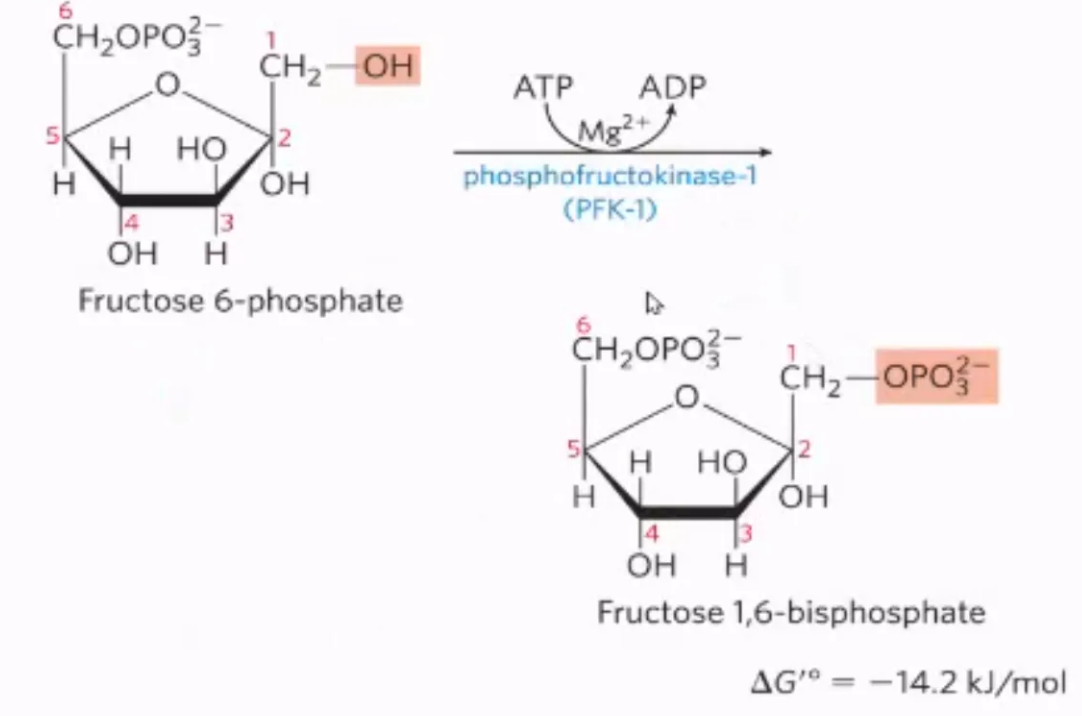
What is the third step of glycolysis, what enzymes mediate it, and why do we do it?
mechanism: transfer of phosphate from onto onto fructose-6-phosphate, converting it into fructose-1,6-bisphosphate by phosphofructokinase
enzyme: phosphofructokinase
why do we do it? To make fructose even more symmetrical since there is only one phosphate (on one side)
what step is this?
#3: phosphorylation of fructose-6-phosphate into fructose-1,6-bisphosphate by phosphofructokinase
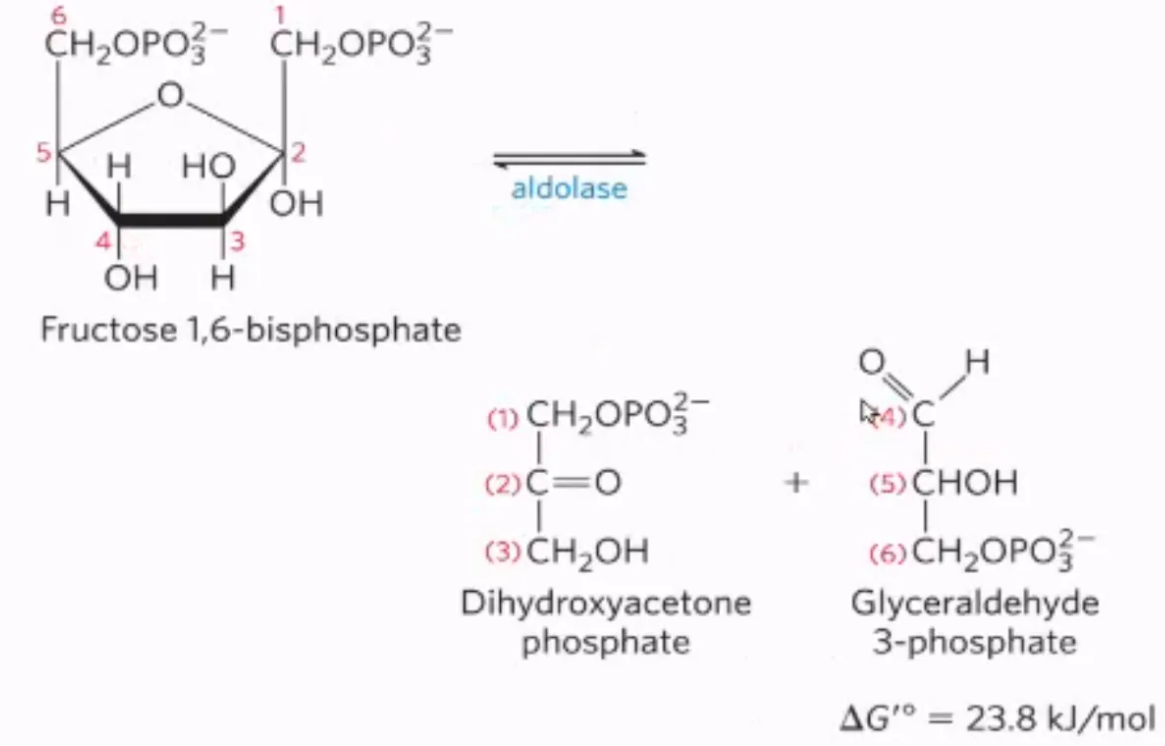
What is the fourth step of glycolysis, what enzymes mediate it, and why do we do it?
mechanism: fructose-1,6-bisphosphate is cleaved into dihydroxyacetonephosphate (DHAP) and glyceraldehyde-3-phosphate (GAP) by aldose
enzyme: aldose
why do we do it? Splitting glucose makes glycolysis more efficient at capturing energy
what carbons is DHAP and G3P composed of?
from the original fructose-1,6-bisphosphate…
DHAP = carbons 1-3
G3P = carbons 4-6
what is this step?
#4: cleavage of fructose-1,6-bisphosphate into GAP and DHAP
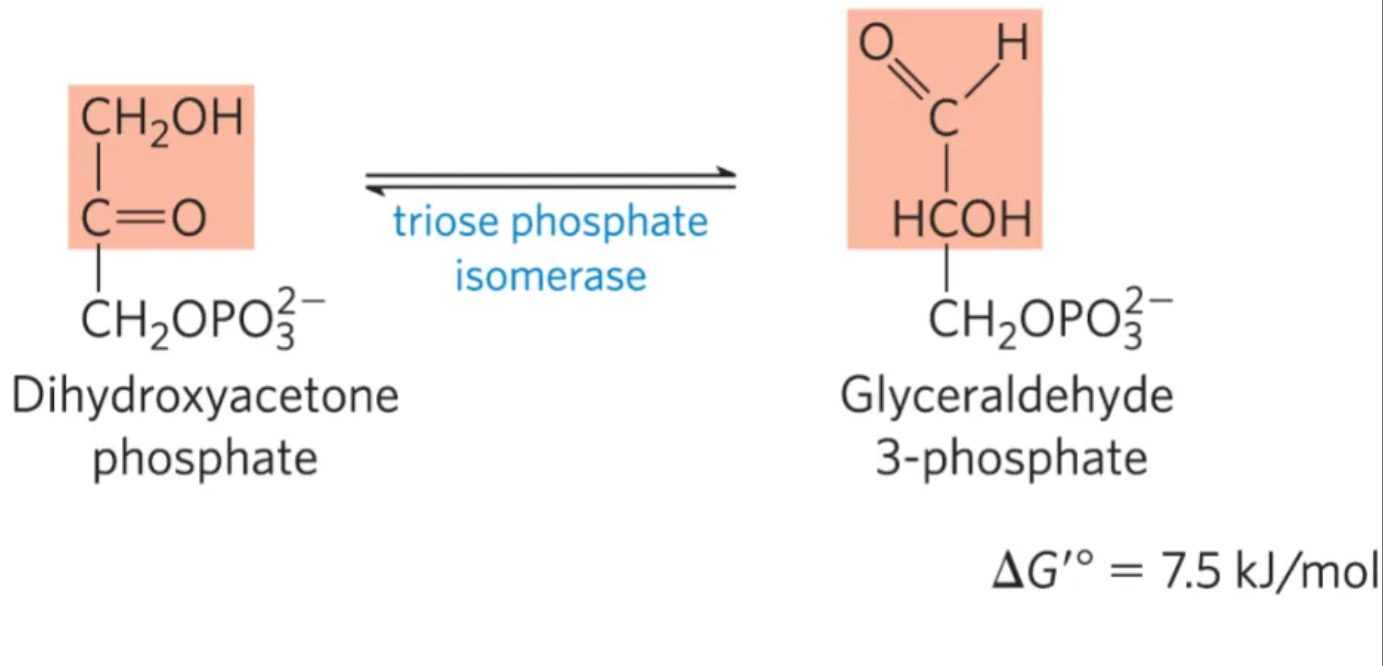
What is the fifth step of glycolysis, what enzymes mediate it, and why do we do it?
mechanism: isomerization of dihydroxyacetonephosphate (DHAP) to glyceraldehyde-3-phosphate (GAP) by triose phosphate isomerase
enzyme: triose phosphate isomerase
why do we do it? GAP is more favorable than DHAP
what is this step?
#5: isomerization of dihydroxyacetoe-phosphate into glyceraldehyde-3-phosphate by triose phosphate isomerase
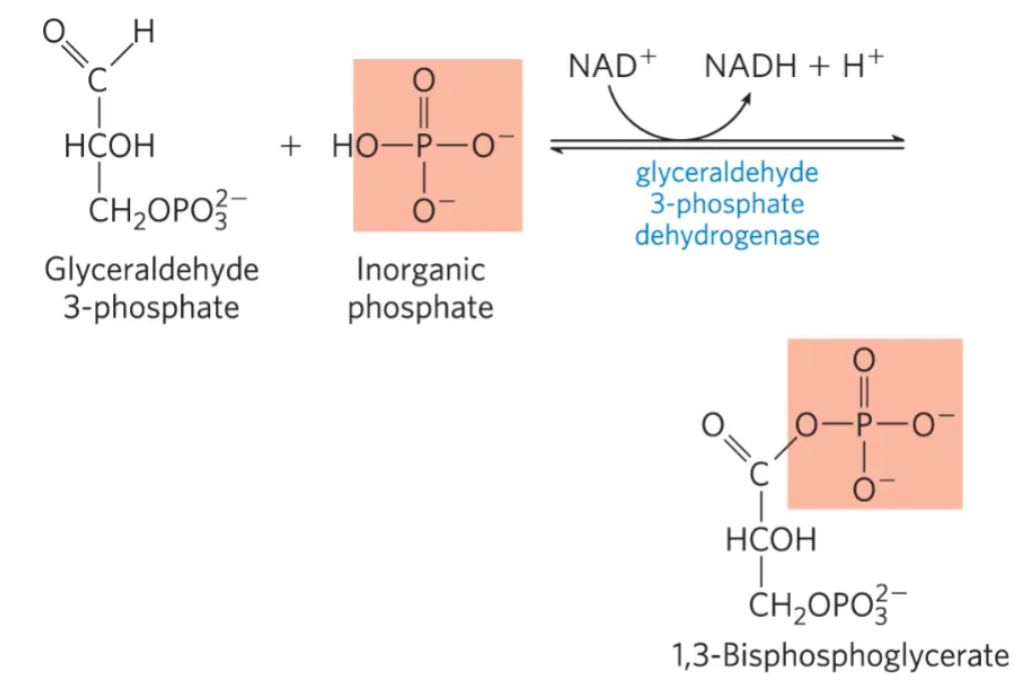
What is the 6th step of glycolysis, what enzymes mediate it, and why do we do it?
mechanism: phosphorylation and oxidation of GAP to 1,3-Bisphosphoglycerate by glyceraldehyde-3-phosphate dehydrogenase
Formation of a Hemithioacetal Intermediate:
GAP (glyceraldehyde 3-phosphate) reacts with a cysteine residue in the active site of GAPDH (enzyme)
The thiol group (–SH) of cysteine nucleophilically attacks the carbonyl carbon of GAP, forming a hemithioacetal intermediate.
Oxidation of the Hemithioacetal to a Thioester:
NAD+, also bound in the active site, accepts 2 electrons and 1 proton from the hemithioacetal intermediate.
This oxidation converts the hemithioacetal to a thioester intermediate, while NAD+ is reduced to NADH.
The thioester bond is high-energy, facilitating the next step.
Release of NADH and Replacement with NAD+:
The newly formed NADH leaves the active site and is replaced by a fresh NAD+.
This exchange is crucial because the presence of NAD+ activates the enzyme for the next catalytic cycle.
Orthophosphate Attack and Product Release:
Inorganic phosphate (Pi) attacks the thioester carbonyl, forming 1,3-bisphosphoglycerate (1,3-BPG).
The thioester bond is cleaved, releasing the product and regenerating the free cysteine residue in the enzyme’s active site.
enzyme: glyceraldehyde-3-phosphate dehydrogenase
why do we do it?
Produces NADH, which carries electrons to the electron transport chain (ETC).
Forms 1,3-BPG, a high-energy intermediate that will participate in substrate-level phosphorylation in Step 7 to produce ATP.
This reaction is exergonic (releases energy), which helps drive the subsequent ATP formation.
what is this step?
#6: oxidation and phosphorylation of glyceraldehyde-3-phosphate to 1,3-biphosphoglycerate by glyceraldehyde-3-phosphate dehydrogenase
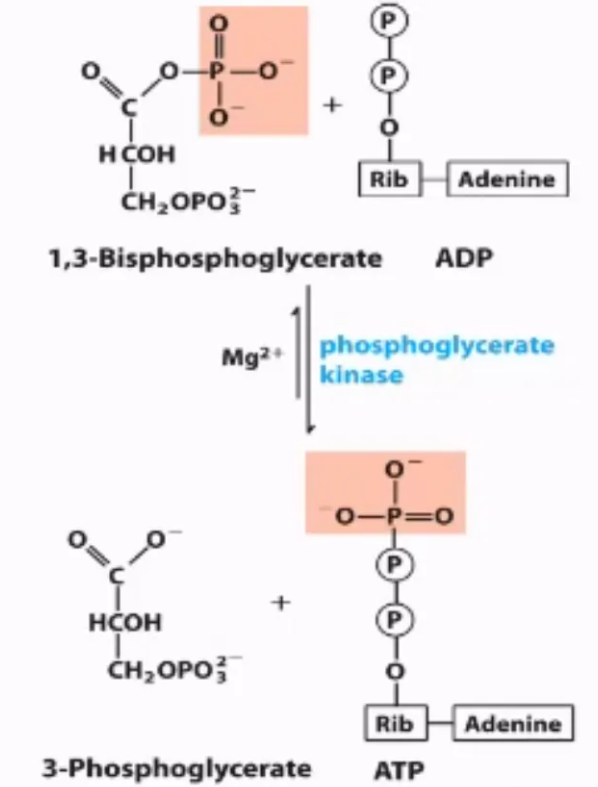
What is the 7th step of glycolysis, what enzymes mediate it, and why do we do it?
mechanism: transfer of the phosphate from 1,3-BPG onto ADP to make ATP and 3-phosphoglycerate
enzyme: phosphoglycerate kinase
why do we do this step?
first time we form ATP in glycolysis
1,3-BPG is a high-energy intermediate with a high phosphoryl transfer potential.
Converting it to 3-phosphoglycerate (3PG) while generating ATP ensures that the energy stored in the high-energy bond is not wasted.
what step is this?
#7: ATP generation via substrate-level phosphorylaton from 1,3-biphosphoglycerate to ADP by phosphoglycerate kinase
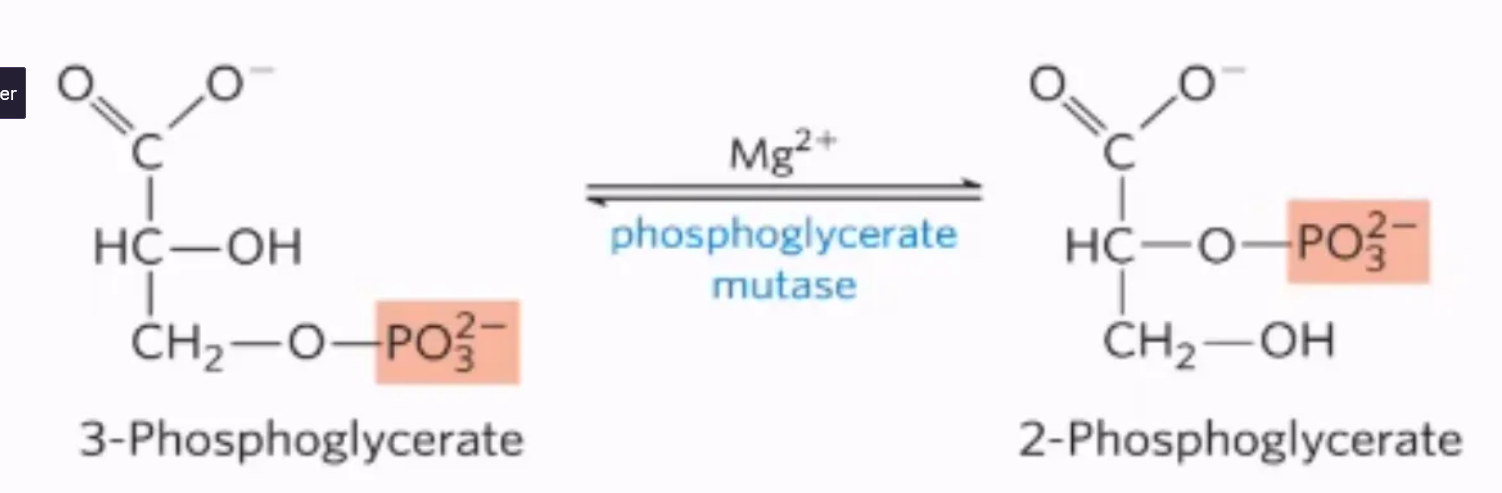
What is the 8th step of glycolysis, what enzymes mediate it, and why do we do it?
mechanism:
Phosphate Group Shuffle:
The enzyme phosphoglycerate mutase shifts the phosphate group 3-phosphoglycerate to 2-phosphoglycerate (2PG)
This involves a temporary phosphorylation of the enzyme itself by a histidine residue, forming a phospho-enzyme intermediate.
Removal of phosphate
the phosphate group comes off the intermediate (2,3-BPG) and re-phosphorylates the enzyme
enzyme: phosphoglycerate mutase
why do we do it?
The rearrangement creates 2-phosphoglycerate, which is a better substrate for dehydration in the next step.
what step is this?
#8: intramolecular shift of 3-phosphoglycerate into 2-phosphoglycerate by phosphoglycerate mutase
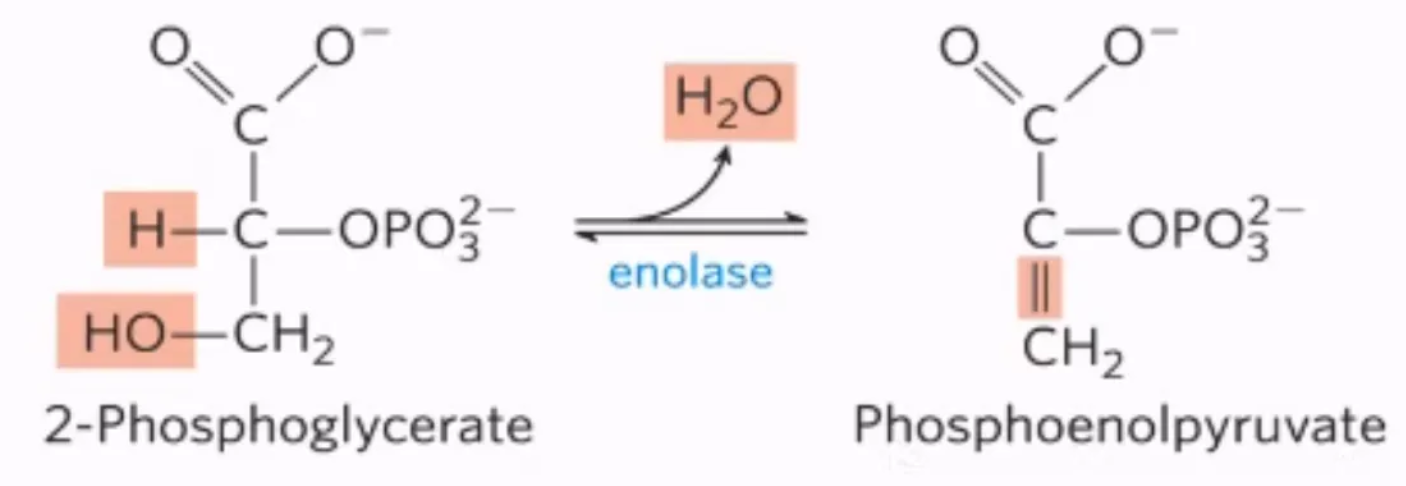
What is the 9th step of glycolysis, what enzymes mediate it, and why do we do it?
mechanism: enolase removes water from 2PG by enolase, forming phosphoenolpyruvate
enzyme: enolase
why do we do it?
enol is more unstable than keto, meaning it has a higher PTP, which will help us generate ATP in the next step
what step is this?
#9: dehydration of 2-phosphoglycerate, forming phosphoenolpyruvate by enolase
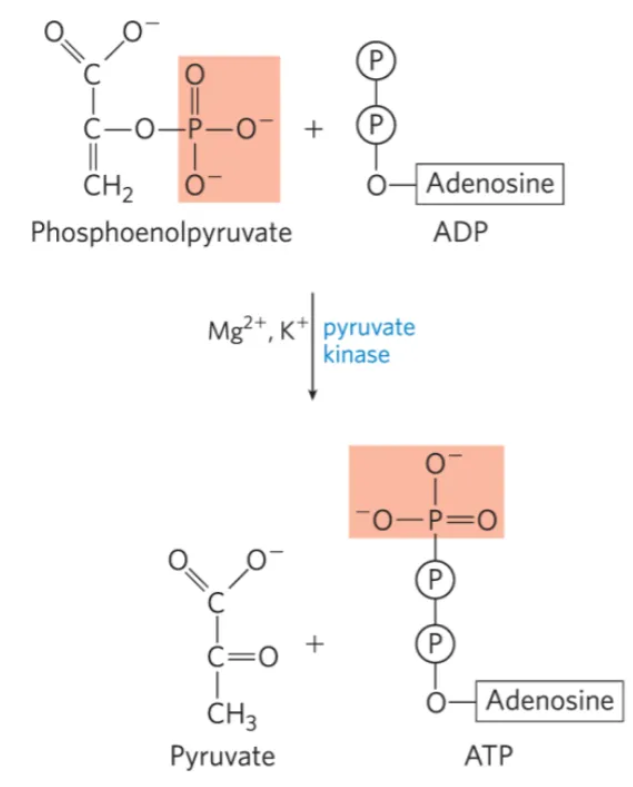
What is the 10th step of glycolysis, what enzymes mediate it, and why do we do it?
mechanism: pyruvate kinase transfers a phosphate group from phosphoenolpyruvate to ADP, forming ATP and pyruvate
phosphoenolpyruvate immediately goes to its enol form (pyruvate) once the phosphate comes off
enzyme: pyruvate kinase
why do we do it?
forms pyruvate which is needed in the citric acid cycle
what step is this?
#10: generation of ATP and pyruvate via substrate level phosphorylation from phosphoenolpyruvate by pyruvate kinase
what is decarboxylase and carboxylase
decarboxylase = removes carboxyl
carboxylase = adds carboxyl group