Cours 18-19 - Épigénétique
1/38
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
39 Terms
Génétique
Partie de la biologie qui étudie les gènes et les lois de l'hérédité
Matériel génétique
Que est que on hérite?
Lois de l’ hérédité
Comment on hérite le matériel génétique ?
Épigénétique
Décrit des facteurs au-delà du code génétique
L'étude des mécanismes qui conduisent à des changements dans l'expression des gènes qui peuvent être transmis d'une cellule à l'autre et qui sont réversibles, mais qui n'impliquent pas de changement dans la séquence de l'ADN.
Régule les phénotypes cellulaires
Dr. Ernest Everett Just
Le père oublié de l’ épigénétique
Années 1930
→ théorie de l'interaction nucléaire-cytoplasmique pour expliquer comment les cellules de l'embryon à un stade précoce participent au processus de développement.
Théorie de la restriction génétique
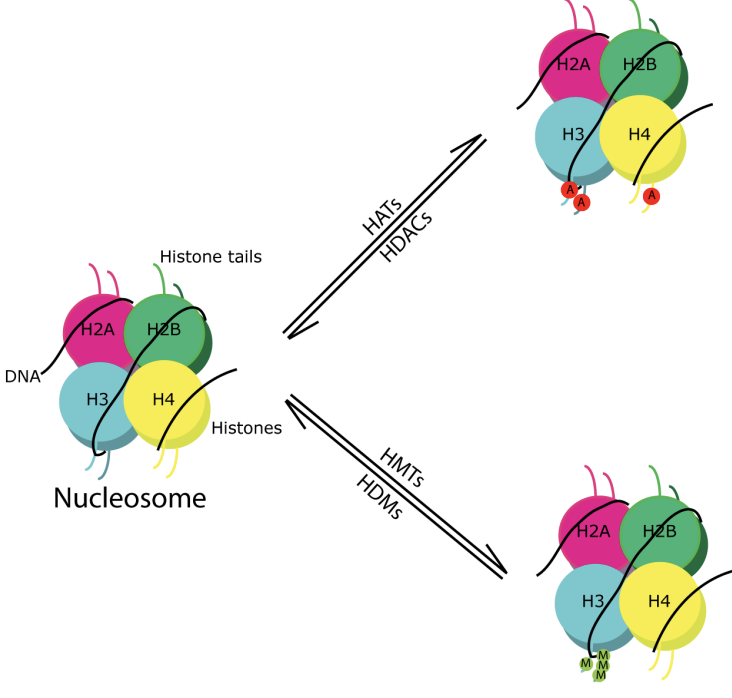
Nucléosome
147 pb
Octamères d’histone
2x H2A-H2B
2x H3-H4
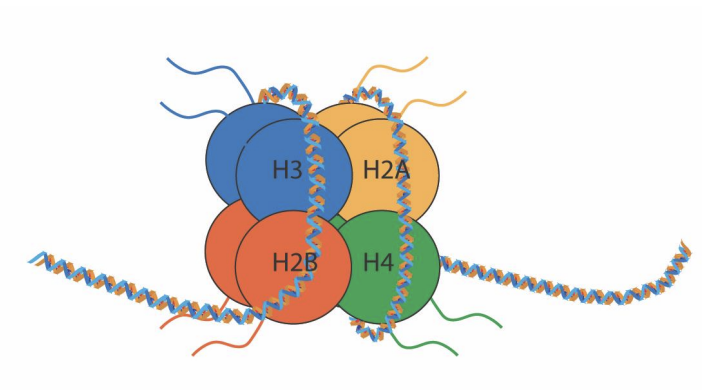
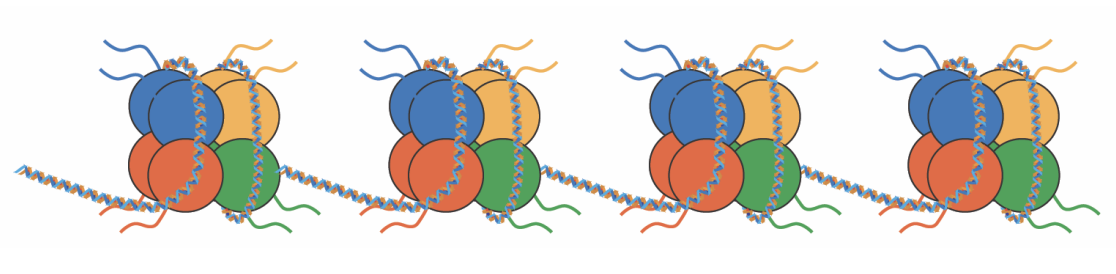
Chromatine
La combinaison de l’ADN et de ses protéines associées
Hétérochromatine
Serrée et condensée
Généralement les gènes sont mal ou pas exprimés
Constitutif ou facultatif
Hétérochromatine Constitutif
Régions toujours serrées entre différentes cellules (télomères, centromères ou chromosomes entiers
Hétérochromatine Facultatif
La région emballée dans certaines cellules mais pas dans d’autres, peut devenir accessible
Euchromatine
Légèrement emballée, riche en gènes et généralement sous transcription active
Mécanismes épigénétiques de régulation de la transcription
Desserrement ou resserrement de la chromatine pour réguler la barrière physique de transcription
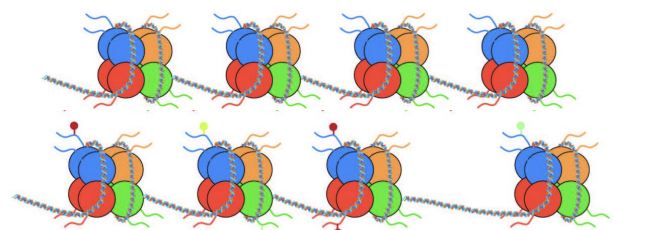
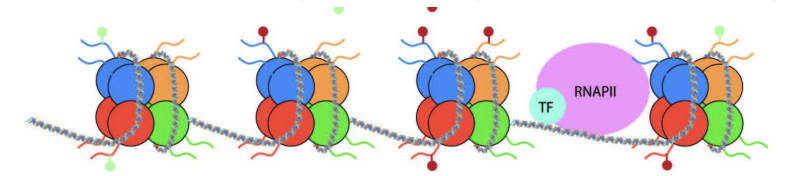
Recrutement de TF et d’autres protéines pour initier ou réprimer la transcription
Modifications post-traductionnelles des histones (PTM)
Des combinaisons de différents PTM sur des nucléosomes spécifiques peuvent activer ou réprimer la transcription (code des histones)
Les histones contiennent une extrémité N intrinsèquement désordonnée et flexible de 19 à 39 résidus s’étendant à partir de la structure globulaire du nucléosome appelée queues d’histones
Les chaînes latérales de résidus d’histones et d’acides aminés sont sujettes à cette modification
Écrivains
Enzymes qui ajoutent des PTM aux histones
Acétyltransférase (HAT)
Méthyltransférase (HMT)
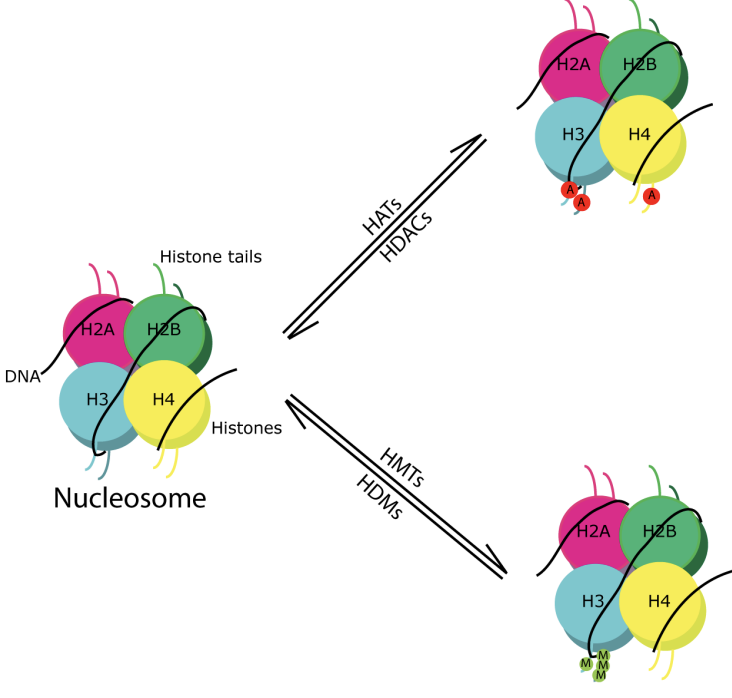
Gommes
Enzymes qui éliminent des PTM spécifiques des histones
Désacétylase (HDAC)
Déméthylase (HDM)
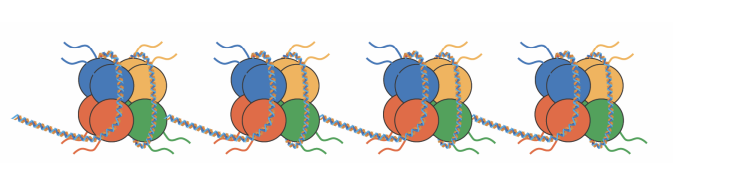
Transcription inactive
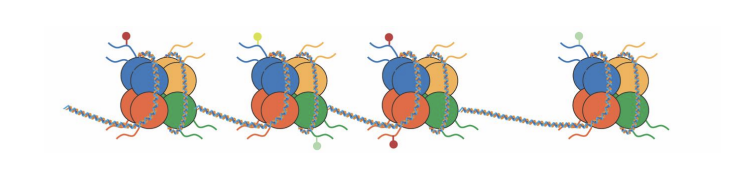
Transcription active
Méthylation de l’ADN
Ajout d’un groupe méthyle au résidu 5 d’une cytosine suivie d’une guanine (CpG)
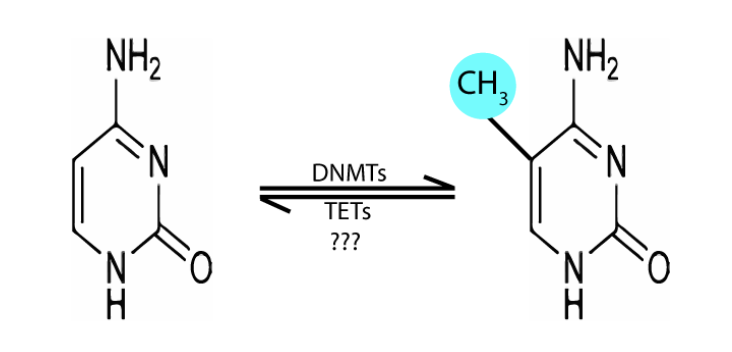
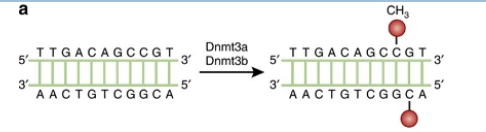
ADN méthyltransférase (DNMT) de novo
Écrivain
DNMT3a et DNMT3b
Méthylent le C dans une région où il n’y a pas de 5-méthyl C dans l’un ou l’autre des brins d’ADN.
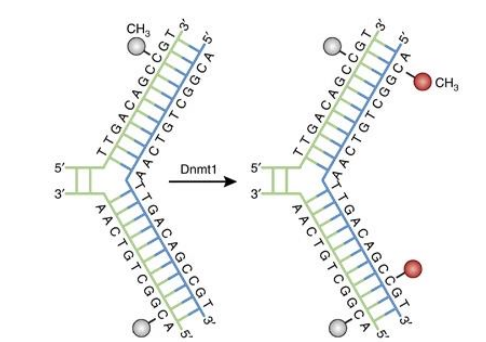
ADN méthyltransférase (DNMT) de maintenance
Écrivain
DNMT1
Actif pendant le cycle cellulaire pour méthyler le nouveau brin d’ADN
Lecteurs
Protéines MBD
Protéines UHRF
Protéines à doigts de zinc
Protéines MBD
Contiennent un domaine de liaison au méthyl-CpG (MBD) conservé qui confère une affinité plus élevée pour les sites CpG méthylés uniques
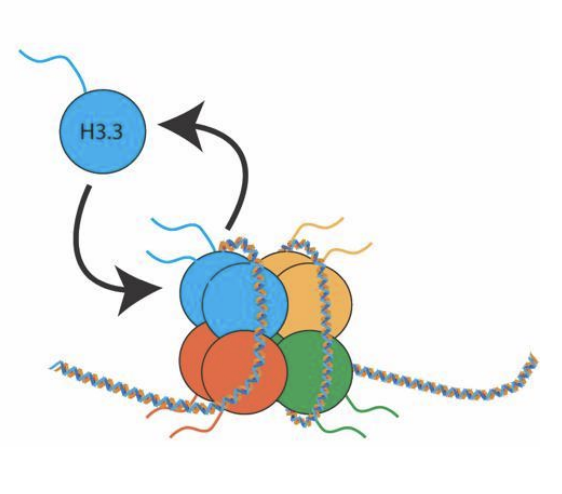
Échange
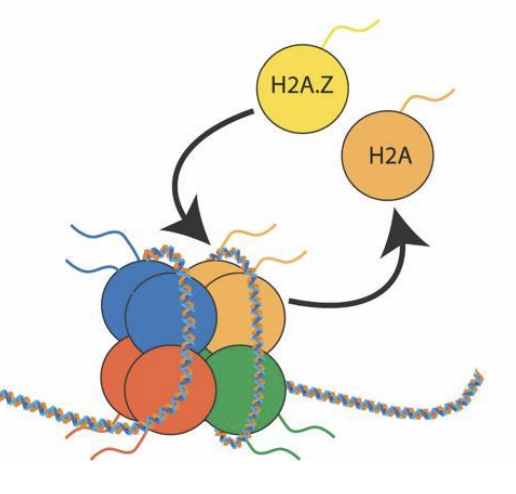
Renouvellement
Reprogrammation
Pour que les gamètes forment un organisme entier, leur épigénome doit être effacé par un processus appelé…
Vrai ou Faux. Presque toutes les méthylations des parents sont effacées, d’abord au début de l’embryogenèse. Une fois le blastocyste formé, la méthylation peut commencer, et une vague de méthylation a alors lieu jusqu’à la phase d’implantation de l’embryon.
Vrai
Zygote
Dans le… l’ADN des spermatozoïdes e des œufs est activement déméthylée
Protamine
Les histones sont remplacées par de la … dans les spermatozoïdes
RCE (régions de contrôle de l'empreinte)
Éléments de séquence qui sont méthylés et modifient d’une façon différente entre les allèles parentaux.
Sont protégées par la reprogrammation pendant la gamétogenèse et embryogenèse
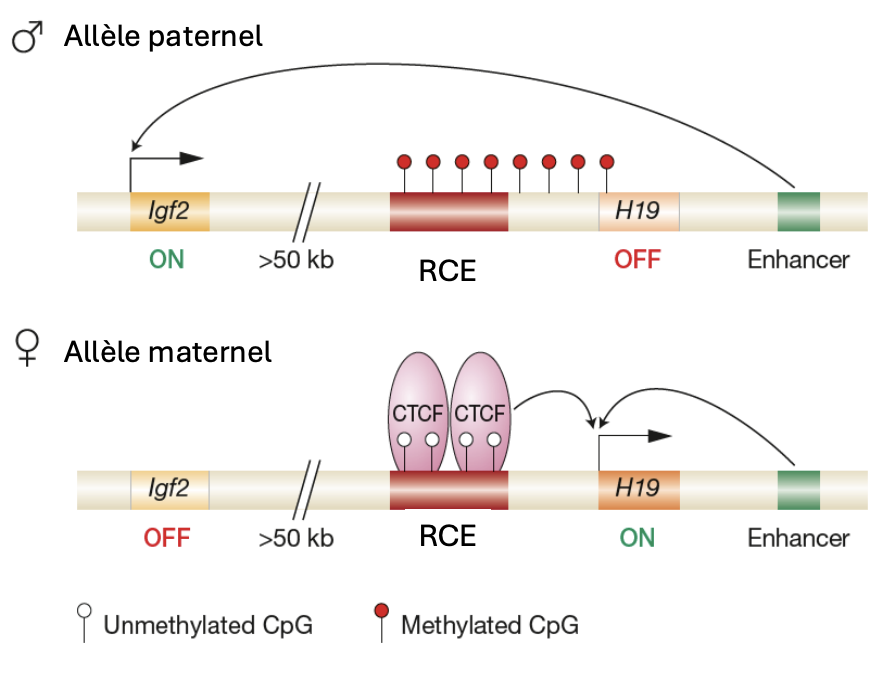
Igf2
Seulement l'allèle paternel est exprimé
Empreinte maternelle non modifiée
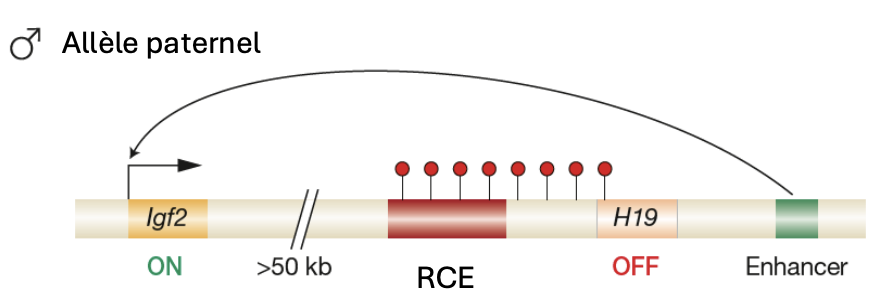
H19
Seulement l’ allèle maternel est exprimé
Empreinte paternelle non modifiée
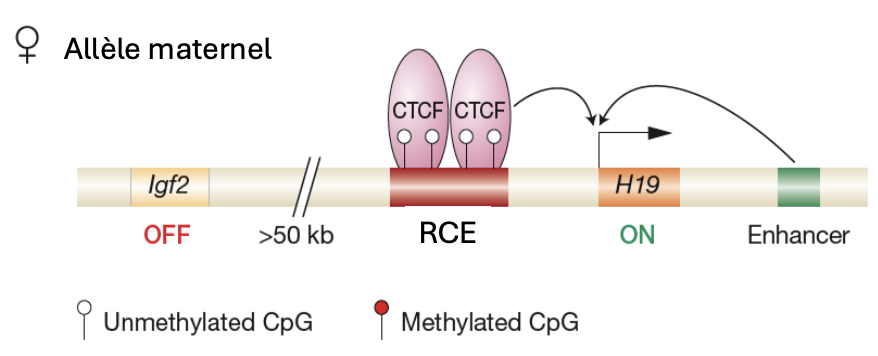
RCE
Est méthylé dans les gamètes males mais non dans les gamètes femelles.
Cette méthylation réprime H19 dans les gamètes males
ICR est lié à CTCF qui bloque la transcription de Igf2 et renforce la transcription de H19
Syndrome de Prader-Willi/ Angelman
Groupe des gènes imprimés sur le chromosome 15
La délétion d’une partie du chromosome 15 donnera un syndrome ou l’ autre en dépendant si on perd les gènes maternels ou paternels
Peuvent aussi être causées quand on hérite de deux chromosome 15 de la mère ou du père
Majorités des gènes est exprimés seulement par le chromosome paternel
Mère: méthylation
Père: pas de méthylation
Syndrome de Prader-Willi
Délétion d’une partie du chromosome 15 du père (75%)
15q11q13
Deux chromosome 15 sont hérites par la mère (25%)
Certains gènes sur le chromosome 15 sont exprimées seulement du chromosome du père
Syndrome de Angelman
Délétion d’une partie du chromosome 15 de la mère (75%)
15q11q13
Deux chromosome 15 sont hérites par le père (25%)
Gene UBE3a (Chr 15) a une empreinte paternelle (seulement l’ allèle de la mère est exprimée)
Hyperméthylation
Prader-Willi, les gènes maternels sont méthylés
Hypométhylation
Angelmann, les gènes paternels ne sont pas méthylés
Inactivation du chromosome X
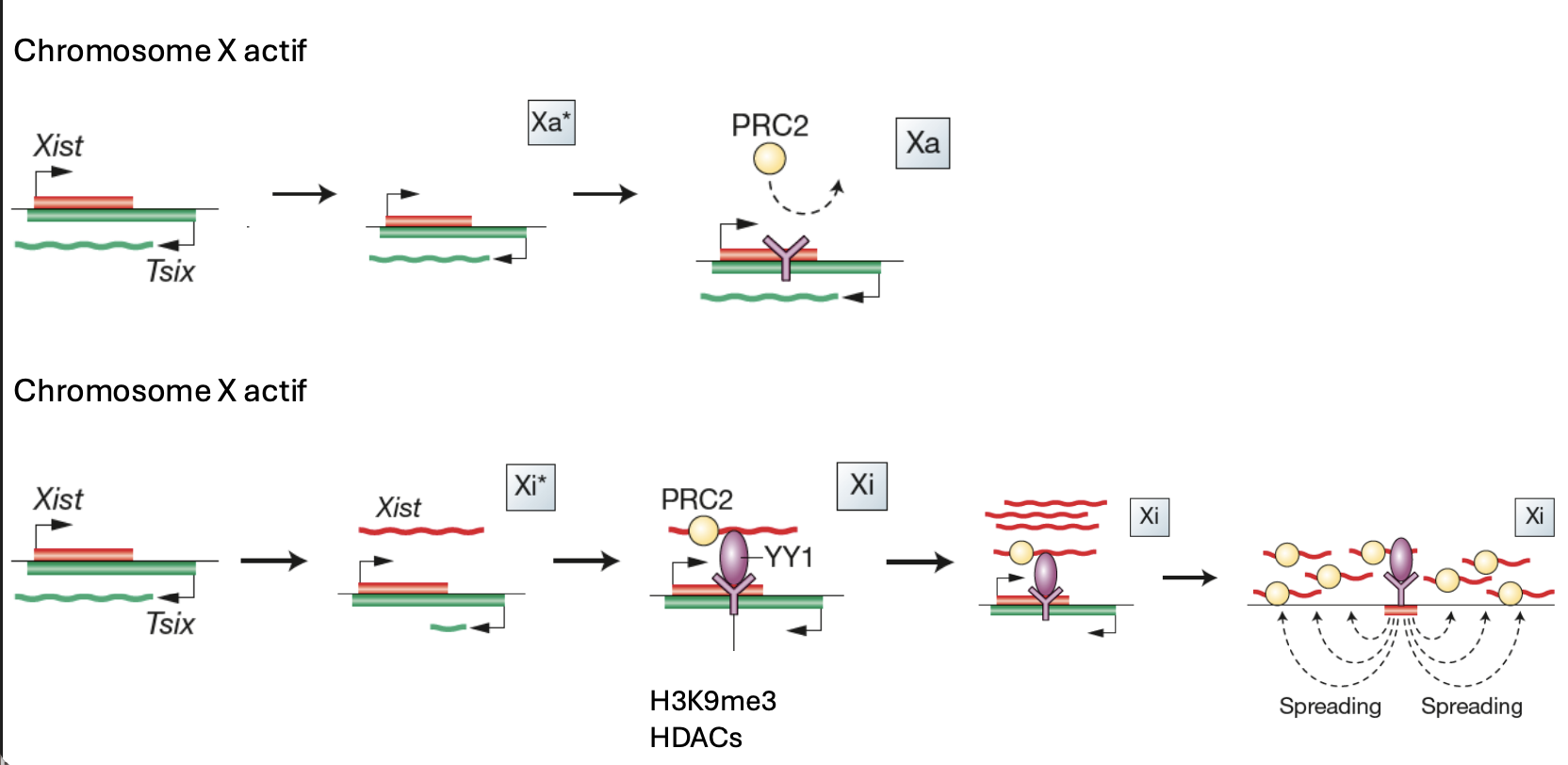
Long ARN non-codant → Xist (X inactive specific transcript)
Transcrit anti-sense → Tsix (Xist au contraire)
Exprimé par le même locus
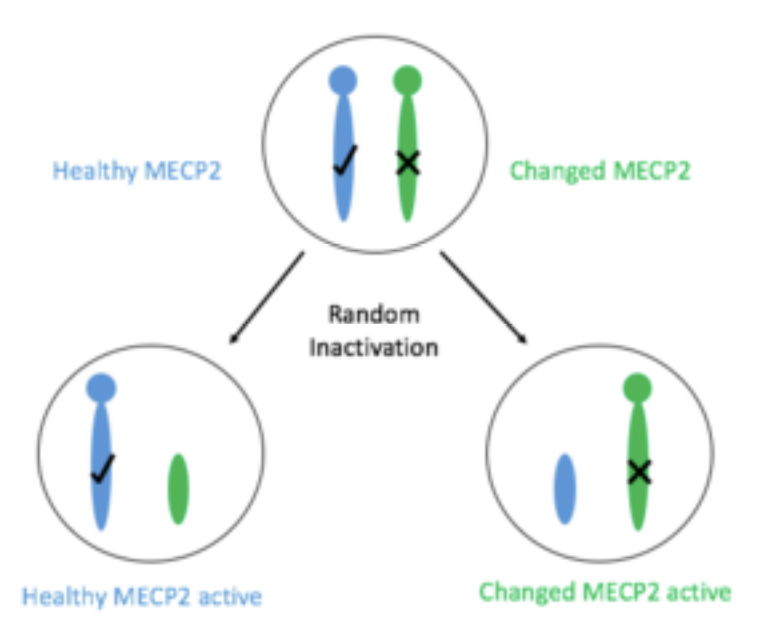
Syndrome de Rett
Mutation de MeCP2 sur le chromosome X
Hérité d’ une façon liée a l’X dominant
Létale pour les males
Des femmes ont la mutation mais elles sont complètement saines
Si beaucoup des cellules inactivent le chromosome mutée la maladie va être très subtile ou pas symptomatique
Ces femmes peuvent transmettre la maladie a la progéniture
Si beaucoup des cellules inactivent le chromosome de type sauvage on va avoir le développement du syndrome de Rett