3. Inmunidad Innata
1/28
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
29 Terms
Cuál es el primer mecanismo de la inmunidad humoral?
El sistema complemento
Sistema del complemento
Cuando el complemento se activa, siempre hace tres cosas:
Lisa los patógenos (mata las bacterias)
Induce inflamación
Opsoniza patógenos
Una vez se activa el complemento, sin importar quien lo active, únicamente hará estas tres cosas
Es un sistema que no tiene control sobre sí mismo
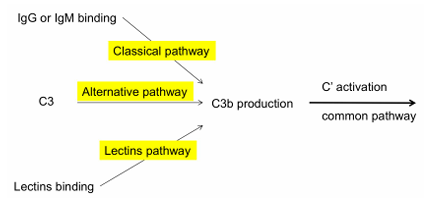
Activación del complemento
Tenemos 3 rutas:
Ruta clásica
Requiere anticuerpos (IgM o IgG) unidos a un antígeno.
El complejo Ag–Ac activa C1q, que inicia la cascada.
Por eso se llama “clásica”: fue la primera descubierta.
Aunque el complemento es innato, esta vía necesita anticuerpos, que son adaptativos. Por eso se dice que es una vía “mixta”
Ruta alternativa
se puede activar por sí mismo sin necesidad de anticuerpos
Es la vía que más rápido responde ante un patógeno nuevo
Ruta de las Lectinas (MBL)
Involucra unas proteínas llamadas Lectinas
Estas son proteínas que reconocen carbohidratos en la superficie de microbios.
Son solubles
Cuando MBL se une a manosa, activa MASP-1 y MASP-2, que inician la cascada
Ruta clásica
No se activa por primera vez, por qué?
Si una bacteria entra por primera vez, tu cuerpo no tiene anticuerpos contra ella.
Por tanto:
❌ Ruta clásica no se activa
✔ Ruta alternativa
✔ Ruta de las lectinas
La segunda vez que entra la bacteria, ya tienes anticuerpos → sí se activa la clásica
Es innata o adaptativa la ruta clásica?
Es una vía del complemento (innata) que puede depender de la inmunidad adaptativa.
No encaja perfectamente en la división clásica “innata vs adaptativa”.
Es un ejemplo de cómo ambos sistemas cooperan
Convergencia de las vías
Aunque empiecen distinto, todas convergen en C3:
C3 → C3a (inflamación) + C3b (opsonización)
Luego activan C5 → MAC → lisis
A partir de C3, todas las vías son idénticas
Se activan con la misma potencia?
Al inicio, la alternativa suele ser la más rápida.
Cuando ya hay anticuerpos, la clásica se vuelve muy potente.
La de lectinas es intermedia.
Pero una vez convergen en C3, la potencia final es la misma
Ruta clásica: interacción antígeno-anticuerpo
La ruta clásica del sistema del complemento comienza cuando los anticuerpos IgG o IgM se unen a la superficie de un patógeno, como una bacteria.
→ interacción Ag-Ac
Unión del anticuerpo al antígeno → cambio conformacional
Cuando un anticuerpo se fija a un antígeno:
Cambia su forma tridimensional (cambio conformacional).
Ese cambio expone una región del anticuerpo llamada dominio Fc, que antes estaba “oculta”.
Este cambio es crucial porque permite que el complemento lo reconozca. (proteína C1 en forma de dedos)
Activación del complejo C1
C1 tiene forma de “dedos” que detectan anticuerpos unidos al patógeno.
C1 solo se activa si se une a al menos dos anticuerpos pegados a la superficie.
Si C1 está libre en el plasma, no se activa.
Una vez activado, C1 inicia una cascada de cortes proteolíticos
Corte de C4
C1 corta la proteína C4:
C4a → Se libera al medio
C4b → Se pega a la superficie del patógeno
Corte de C2
C1 corta C2, y uno de sus fragmentos se une a C4b
Esto forma:
El complejo C4b2a (o C4b2b)
Este complejo es la C3 convertasa de la vía clásica
Formación de C3b y C3a
La C3 convertasa corta C3
C3b → se pega masivamente a la superficie del patógeno
C3a → se libera y participa en inflamación
Punto de convergencia
La C3 convertasa es el punto donde las tres vías del complemento (clásica, alternativa y lectinas) se vuelven comunes.
A partir de aquí, todas generan:
Opsonización
Lisis
Inflamación

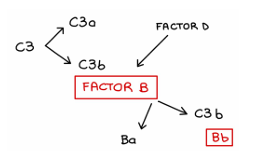
Ruta alternativa
Recordemos que se caracteriza porque no necesita anticuerpos para activarse, a diferencia de la ruta clásica
—
Activación espontánea de C3
Aquí la C3 se puede activar espontáneamente
→ ocurre de forma continua y a bajo nivel en el organismo.
Cuando hay superficies bacterianas, este proceso espontáneo:
Se estabiliza
Se vuelve efectivo
Permite que la vía avance
→ C3 se rompe en:
C3a, que se libera al entorno
C3b, que se adhiere a la superficie del patógeno
Unión de C3b a la proteína B
Una vez C3b está unido al patógeno:
se une a la proteína B
esta combinación sufre una autocleavage (autoproteólisis)
Ese proceso genera la C3 convertasa específica de la ruta alternativa
Amplificación
Esta C3 convertasa corta grandes cantidades de C3, produciendo más C3b.
Así se amplifica la respuesta inmunitaria.
Convergencia
La ruta alternativa llega al mismo punto central que las otras vías:
la generación masiva de C3b sobre la superficie del patógeno.
Característica principal
Lo más destacable es que no necesita anticuerpos, por lo que es una respuesta:
innata
inespecífica
capaz de actuar contra cualquier microorganismo invasor directamente
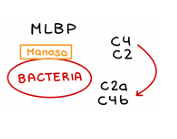
Ruta de las lectinas
Las lectinas son proteínas que reconocen y se unen a azúcares específicos, como la manosa, presentes en la superficie de muchas bacterias.
Cuando una lectina como MBL (lectina de unión a manosa) detecta estos azúcares:
• se une a la superficie bacteriana
ACTIVACIÓN DE MASP
Una vez unidas, las lectinas activan unas enzimas llamadas MASP que:
Imitan la función de las C1
Comienzan la cascada cortando C4
Luego cortan C3
Esto forma el complejo C4b2a, que es la C3 convertasa
Continuación de la vía:
Desde aquí, la ruta de las lectinas sigue los mismos pasos que la clásica:
genera C3b
amplifica la respuesta inmunitaria
Importancia
Esta vía permite que el sistema inmune:
reconozca y ataque bacterias sin haberlas visto antes
actúe de forma innata e inespecífica
se base en estructuras comunes de muchos microorganismos
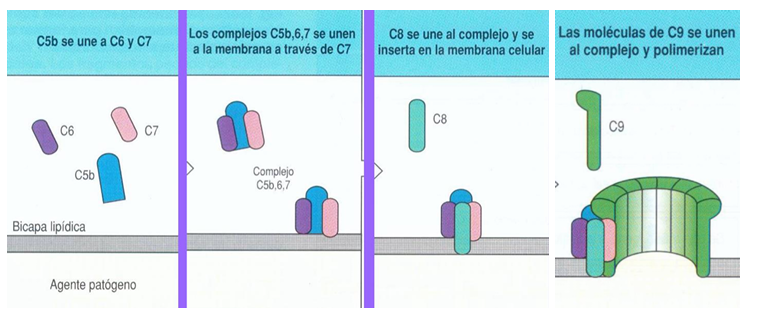
Rama de salida del complemento
FORMACIÓN DE LA C5 CONVERTASA
Una vez que cualquiera de las 3 rutas ha generado C3 convertasa, esta enzima puede evolucionar y convertirse en C5 convertasa
Su función principal es cortar la proteína C5 en 2 fragmentos (a y b)
INICIO DEL COMPLEJO DE ATAQUE A LA MEMBRANA (MAC)
El fragmento C5b inicia una nueva fase del complemento
Secuencia de unión:
C5b se une a C6
Luego se une a C7
→ Este complejo se fija a la superficie de la bacteria
Después recluta C8
Finalmente se unen múltiples moléculas de C9
YA TENEMOS MAC FORMADO
Las moléculas de C9 pueden polimerizar, es decir, formar una estructura repetitiva que atraviesa la membrana
Resultado:
C9 forma un poro en la membrana bacteriana, creando una abertura que compromete su integridad
Función del MAC
El MAC es el primer mecanismo del complemento con acción letal directa:
perfora la membrana de la bacteria
La bacteria pierde contenido
Finalmente muere
FUnciones adicionales del complemento
Además de matar bacterias, el complemento participa en otros procesos importantes:
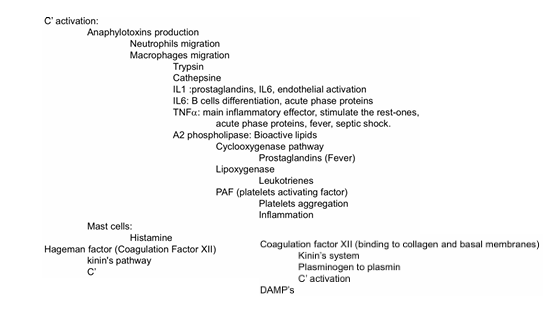
ANAFILOTOXINAS
Los fragmentos solubles C3a y C5a, liberados durante la activación:
Actúan como anafilotoxinas
Producen inflamación
Atraen células del sistema inmune al sitio de infección
Así, el complemento funciona también como señalizador de peligro para el resto del sistema inmune
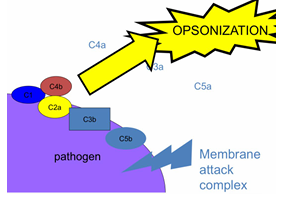
OPSONIZACIÓN
Otra función esencial es la opsonización.
La bacteria queda “tapizada” o “rebozada” con proteínas, especialmente C3b
Estas proteínas actúan como etiquetas de marcado
Facilitan que células fagocíticas (como macrófagos) reconozcan y engullan la bacteria
Esto permite eliminarla sin necesidad de matarla por lisis
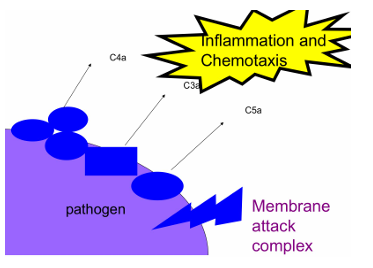
Resumen
En resumen, el complemento tiene tres funciones clave: matar bacterias mediante la formación del poro (MAC), generar inflamación atrayendo otras células inmunes, y opsonizar bacterias para facilitar su fagocitosis.
Por tanto, entender cómo se activa (a través de las tres rutas) y qué efectos genera (la rama de salida) es esencial para comprender su papel en la inmunidad innata.
Finalmente, también se menciona la existencia de una posible forma más primitiva y simple del complemento, conocida como la ruta de las cininas
Ruta de las quininas
Es una vía más sencilla que las del complemento, pero sigue una lógica similar: funciona mediante una cascada de activación proteolítica. Aunque es menos compleja, cumple funciones esenciales en la respuesta inflamatoria
Inicio de la vía
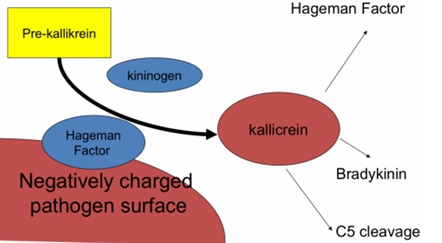
ACTIVACIÓN DEL FACTOR XII (factor de Hageman)
La ruta comienza cuando:
Una bacteria o,
Una superficie con carga negativa
entra en contacto con el factor de Hageman (factor XII), una proteína del sistema de coagulación.
Cuando el factor XII se activa —por ejemplo:
al unirse a bacterias
o al colágeno expuesto tras una lesión vascular—
no solo inicia la coagulación intrínseca, sino que también activa la ruta de las quininas.
Esto conecta coagulación e inmunidad
QUE OTRA PROTEÍNA INTERVIENE?
El kininógeno
Colabora para activar kallicreína
→ enzima capaz de provocar inflamación
y Atraer células del sistema inmunitario al sitio de infección o lesión
Este mecanismo explica fenómenos cotidianos:
Cuando te tuerces un tobillo, parte de la inflamación proviene no solo del daño físico, sino también de la activación inmunitaria a través de esta vía compartida con la coagulación
Papel dentro de la inmunidad innata
La ruta de las quininas, junto con el complemento, forma parte de la respuesta humoral innata, es decir, mecanismos no específicos que actúan como primera línea de defensa.
Mientras tanto, el complemento cumple un papel fundamental al marcar patógenos:
C3b “tapiza” a las bacterias
funciona como una etiqueta molecular
guía a células inmunitarias como los macrófagos hacia el objetivo
Los macrófagos siguen:
el “rastro” de moléculas liberadas por la bacteria
y las señales del complemento
hasta llegar al sitio de infección, donde pueden fagocitar al patógeno
Macrófagos
Son un tipo de fagocitos
células especializadas en englobar y destruir microorganismos, pero no los utilizan como fuente de nutrición
Función principal:
Eliminar patógenos una vez que los han localizado
Qué necesitan los macrófagos para poder fagocitar?
Receptores específicos que les permitan reconocer qué hay que fagocitar
Mecanismos
OPSONIZACIÓN Y RECONOCIMIENTO DE C3B
Un mecanismo clave es la opsonización:
Las bacterias son “aliñadas” con proteínas como C3b del complemento.
C3b actúa como una etiqueta molecular.
Los macrófagos tienen receptores para C3b, lo que les permite reconocer con alta afinidad a las bacterias marcadas y fagocitarlas eficazmente
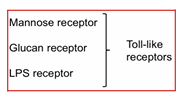
RECEPTORES TOLL-LIKE (TLR)
Qué detectan?
PAMPs (patrones moleculares comunes en patógenos) (Patrón Molecular Asociado a Patógenos)
El nombre “Toll-Like” viene de:
Toll, un receptor de Drosophila melanogaster usado en desarrollo embrionario
Los TLR se parecen estructuralmente a ese receptor, aunque no tengan relación funcional
Qué reconocen los TLR?
Reconocen, por ejemplo:
Manosa en paredes bacterianas
ARN de doble cadena, típico de ciertos virus
Cuántas familias tenemos?
Tenemos en total 7 - dónde se encuentran?
Algunos en la superficie de los macrófagos, otros dentro (como TLR4, que reconoce ARN de doble cadena)
Para que un macrófago detecte ARN de doble cadena:
Debe estar infectado por un virus, porque normalmente no hay ARN de doble cadena en el citoplasma
Por lo que reconoce el patrón molecular de algo que no va bien
Esto permite que actúen rápida e inespecíficamente, sin contacto previo con el patógeno
Otros receptores importantes
Los macrófagos también reconocen:
Fragmentos de C3b
Opsonización
Uniones Ag-Ac
Los mismos que activan la ruta clásica
Scavenger receptors
Receptores “carroñeros” que limpian restos celulares, actuando como los barrenderos del cuerpo
Funciones
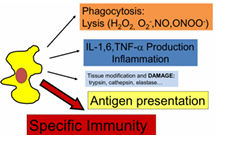
FAGOCITOSIS
Es la primera función esencial
El macrófago engloba la bacteria → fagosoma
El fagosoma se fusiona con un lisosoma → fagolisosoma
El fagolisosoma contiene:
Enzimas digestivas
Radicales libres (muy tóxicos)
Los radicales libres oxidan proteínas, las vuelven no funcionales y causan la muerte de la bacteria
PAPEL EN LA INFLAMACIÓN
Los macrófagos también participan en la inflamación:
Liberan interleucinas proinflamatorias
Liberan TNF, que puede inducir fiebre actuando sobre el hipotálamo
Esta inflamación suele estar precedida por el complemento, que atrae a los macrófagos.
Una vez llegan, los macrófagos aumentan la inflamación mediante interleucinas.
El aumento de temperatura no es perjudicial: puede limitar el crecimiento bacteriano.
Sin embargo, esta activación también puede causar daño tisular, porque los macrófagos liberan proteasas que transforman el tejido en tejido fibroso, más difícil de atravesar por la infección
PRESENTACIÓN ANTIGÉNICA
Finalmente, los macrófagos también realizan presentación antigénica:
Tras fagocitar un patógeno, procesan sus antígenos
Los exponen en su superficie
Otras células del sistema inmune pueden reconocerlos
Esto convierte a los macrófagos en un puente entre inmunidad innata y adaptativa:
Son parte de la respuesta inespecífica
Pero activan a células como los linfocitos T, esenciales en la inmunidad adaptativa
Inflamación
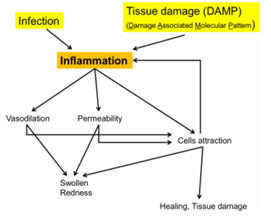
La inflamación es una respuesta compleja ante infección o daño tisular.
Aunque participan muchas moléculas, puede entenderse por sus efectos principales
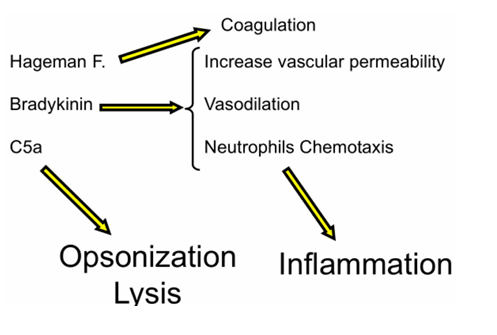
Papel del complemento
Uno de los principales desencadenantes es el complemento:
Sus moléculas solubles (anafilotoxinas) tienen un fuerte efecto vasoactivo
Se unen al endotelio y provocan vasodilatación
Aumenta el flujo sanguíneo
Aparece el enrojecimiento
También aumenta la permeabilidad vascular, permitiendo:
Salida de líquido → edema
Salida de células inmunes, como monocitos, que en el tejido se conviertes en macrófagos
Orígenes de la inflamación
La inflamación puede ser:
Infecciosa → respuesta frente a patógenos
Traumática → torceduras, golpes
Aquí interviene el factor de Hageman (factor XII)
La inflamación ocurre sin infección real
El sistema inmune puede causar daño innecesario
Por eso se aplica frío en lesiones traumáticas:
→ reduce la inflamación
→ evita daño tisular por una respuesta inmunitaria excesiva
Respuesta innata
Durante una infección, lo primero que se activa es la respuesta innata, y en concreto el sistema del complemento, ya que existen muchas más moléculas que células inmunitarias circulantes.
Cómo se activa el complemento?
A través de distintas rutas de entrada según el contexto
Por ejemplo, si se trata de una bacteria desconocida, se activará principalmente la ruta alternativa, que no necesita anticuerpos para funcionar.
Una vez activado, el complemento realiza 3 funciones principales:
Lisis de bacterias
Opsonización (marcar patógenos para facilitar su destrucción)
Inflamación, que incluye:
atracción de macrófagos
vasodilatación del tejido infectado
Después, entra en fuego la fagocitosis, donde participan receptores como los TLR.
Si los macrófagos encuentran bacterias opsonizadas con C3b, su capacidad de fagocitar aumenta muchísimo:
se convierten en “súper fagocitos”.
Cuando estos fagocitos se activan, generan una respuesta sistémica que incluye:
fiebre
producción de proteínas de fase aguda
daño tisular
presentación antigénica (clave para activar la inmunidad adaptativa)
Hay competencia entre rutas del complemento?
Puede parecer que una ruta podría bloquear a otra (por ejemplo, si hay mucho C3b, impedir el acceso de anticuerpos o lectinas).
Pero no existe competencia real.
Esto se debe a que:
actúan en escalas distintas (molecular vs celular)
no interfieren de forma significativa
pueden activarse simultáneamente
👉 El sistema inmunitario está diseñado para ser redundante y eficaz, atacando desde varios frentes sin que una vía bloquee a las demás
Proteínas de fase aguda
Las proteínas de fase aguda son proteínas producidas principalmente en el hígado como parte de la respuesta inflamatoria sistémica inducida por la activación del sistema inmunitario.
Cuando se activa la respuesta inmune (complemento + macrófagos), aparece inflamación.
La inflamación es necesaria, pero también causa daño tisular, por lo que debe ser regulada.
El organismo necesita mecanismos de inactivación para recuperar el equilibrio.
Esta inactivación es humoral, basada en proteínas solubles en sangre.
Curiosamente:
👉 el mismo sistema que genera inflamación activa también los mecanismos que la frenan.
Los macrófagos activados liberan señales que inducen al hígado a producir estas proteínas reguladoras.
Las proteínas de fase aguda:
modulan la inflamación
facilitan la reparación tisular
refuerzan la defensa frente a patógenos
Su concentración aumenta según la intensidad de la inflamación.
La más conocida es la Proteína C Reactiva (PCR), usada clínicamente como marcador de inflamación
Proteína C reactiva
La PCR es una proteína de fase aguda fundamental y un marcador clínico de inflamación.
Su aumento en sangre indica un proceso inflamatorio activo, aunque no sea visible.
Es útil para detectar infecciones, inflamaciones ocultas o monitorizar enfermedades crónicas.
Sus funciones parecen contradictorias, pero en realidad muestran el equilibrio entre activación y regulación:
🔸 Función detoxificadora
Se une a moléculas tóxicas liberadas por células dañadas.
Las neutraliza y limita la expansión del daño
🔸 Inducción de citoquinas antiinflamatorias
Puede activar macrófagos para producir citoquinas antiinflamatorias, frenando la inflamación cuando el patógeno ya está controlado.
🔸 Función proinflamatoria y opsonina
En fases iniciales, actúa como opsonina, recubriendo microorganismos.
Facilita su reconocimiento por fagocitos.
Incluso puede activar la ruta clásica del complemento.
Estas funciones se complementan:
👉 activa el sistema inmune cuando es necesario,
👉 lo apaga cuando la amenaza ha pasado
Este equilibrio evita daño excesivo al organismo
Otras proteínas de fase aguda
Proteína Sérica Amiloide A (SAA)
Elimina restos celulares: lípidos, colesterol, células muertas
Ayuda a resolver la inflamación inhibiendo la fiebre
Puede elevarse en enfermedades inflamatorias crónicas o autoinmunes
2. Proteína α‑1‑antitripsina
Función antiinflamatoria: inhibe enzimas proteolíticas que dañan tejidos
Controla la fibrosis: ayuda a revertir el endurecimiento fibroso generado durante la inflamación
Favorece la producción de citoquinas antiinflamatorias y la regeneración tisular
⭐ Conclusión
Estas proteínas:
limpian,
regulan,
reparan el daño causado durante la inflamación.
Son esenciales para que la respuesta inmunitaria no se prolongue más de lo necesario, evitando inflamación crónica o autoinmunidad.