Michaelmas ecology and evolution
1/30
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
31 Terms
what were the two schools of thought around darwinism and what did they believe?
biometricians supported darwinism
they analysed data from natural populations, of continuous variation
this made them believe natural selection acted to cause small variations
mendelians opposed darwinism
they analysed data from lab experiments + crosses, of discontinuous variation
this made them believe that natural selection wouldn’t cause appreciable changes that accounted for very different forms of species in a viable time frame
they believed in ‘mutations’ or big leaps causing the widely different forms
what are the assumptions in the hardy-weinberg theorem and what does it state?
assumptions:
there is an infinite population size
mating is random
the genes of interest have no impact on fitness/do not have differences in viability
the hardy-weinberg theorem states that allele and genotype frequencies in a population will remain constant from generation to generation in the absence of other evolutionary influences eg. mutations, selection, or genetic drift
P² + 2PQ + Q² = 1
P + Q = 1
how can the hardy-weinberg theorem be adapted to account for selection?
fitness (W) can be calculated- the relative reproductive rate of an individual with a given genotype (how likely the allele is to be inherited)
this is dependent on:
the selection coefficient (s)- whether the allele is more beneficial (positive coefficient) or more deleterious (negative coefficient) than Q
the dominance (h)- dominant = 1, recessive = 0
fitness is measured relative to Q, so Q = 1:
the fitness of P in P homozygous individuals (P²) is 1+s
the fitness of P in heterozygous individuals (2PQ) is 1+hs (accounts for whether it is dominant or recessive)
the fitness of P in Q homozygous individuals (Q²) is 0
if you add the fitnesses for the P allele and the Q allele (1), you get the average fitness (W) = P²(1+s) + 2PQ (1+hs) + Q²(1)
the frequency of the P allele in the next generation is P²(1+s)/W + PQ(1+hs)/W
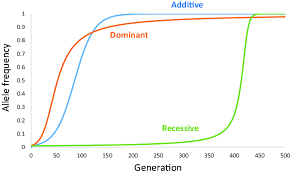
what is the graph of a beneficial allele frequency when it is introduced to a new population?
recessive beneficial alleles take longer to fix in a population because they can only show a beneficial effect when homozygous, which is rare at first
the frequency slowly increases at first, until some organisms are homozygous, where it causes a ‘selective sweep’ because that allele will then be greatly selected for
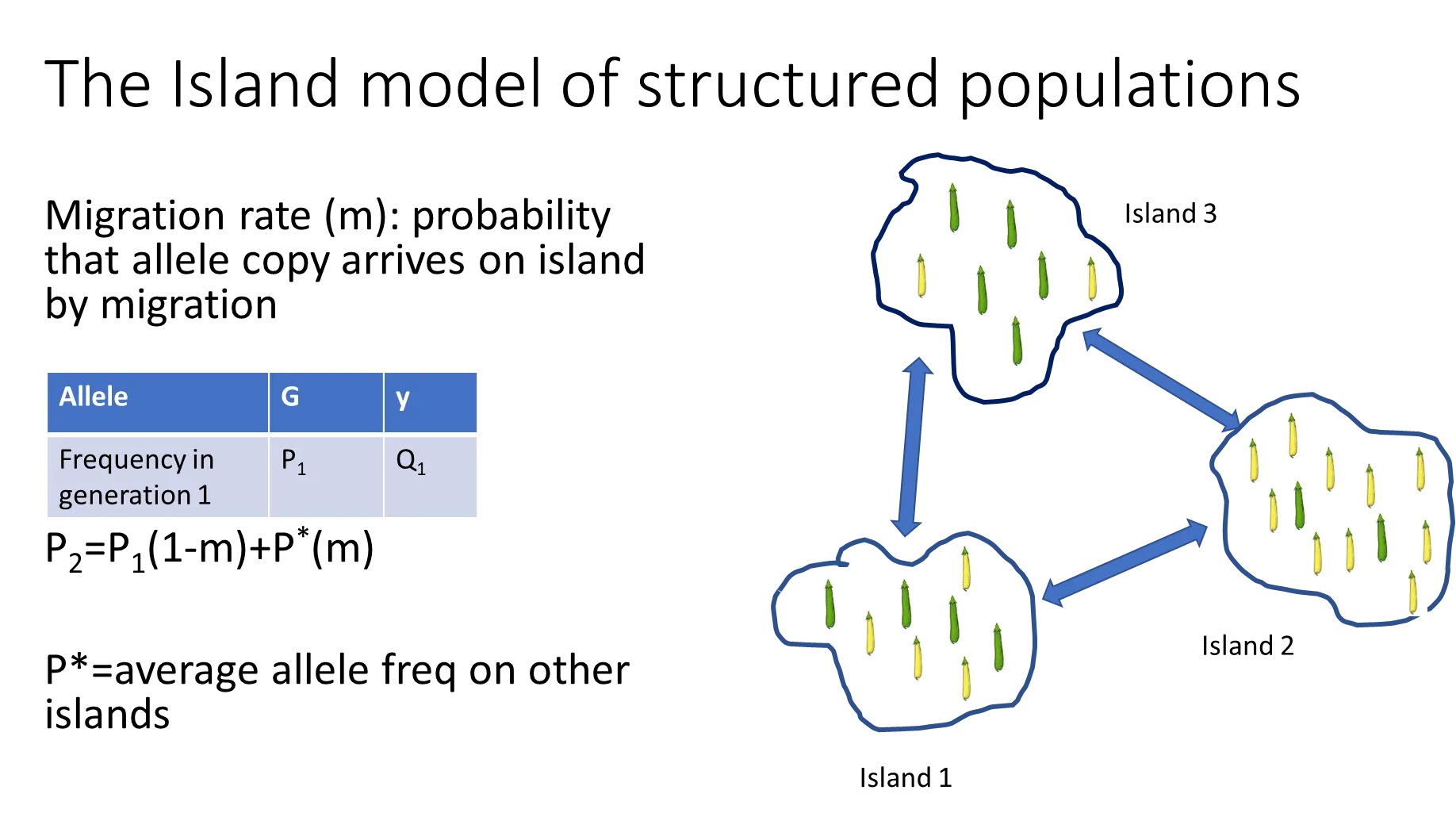
how can the hardy-weinberg theorem be adapted for an island model of structured populations?
the allele frequency in the next generation (on one island) is dependent on the migration rate (m)- the probability that the allele arrives on the island by migration
the frequency in the next generation (P2) = P1(1-m) + P* (m)
where P* = the average allele frequency on other islands
(the 1-m is because if there is migration of the allele onto the island, there is also migration of the allele off the island)
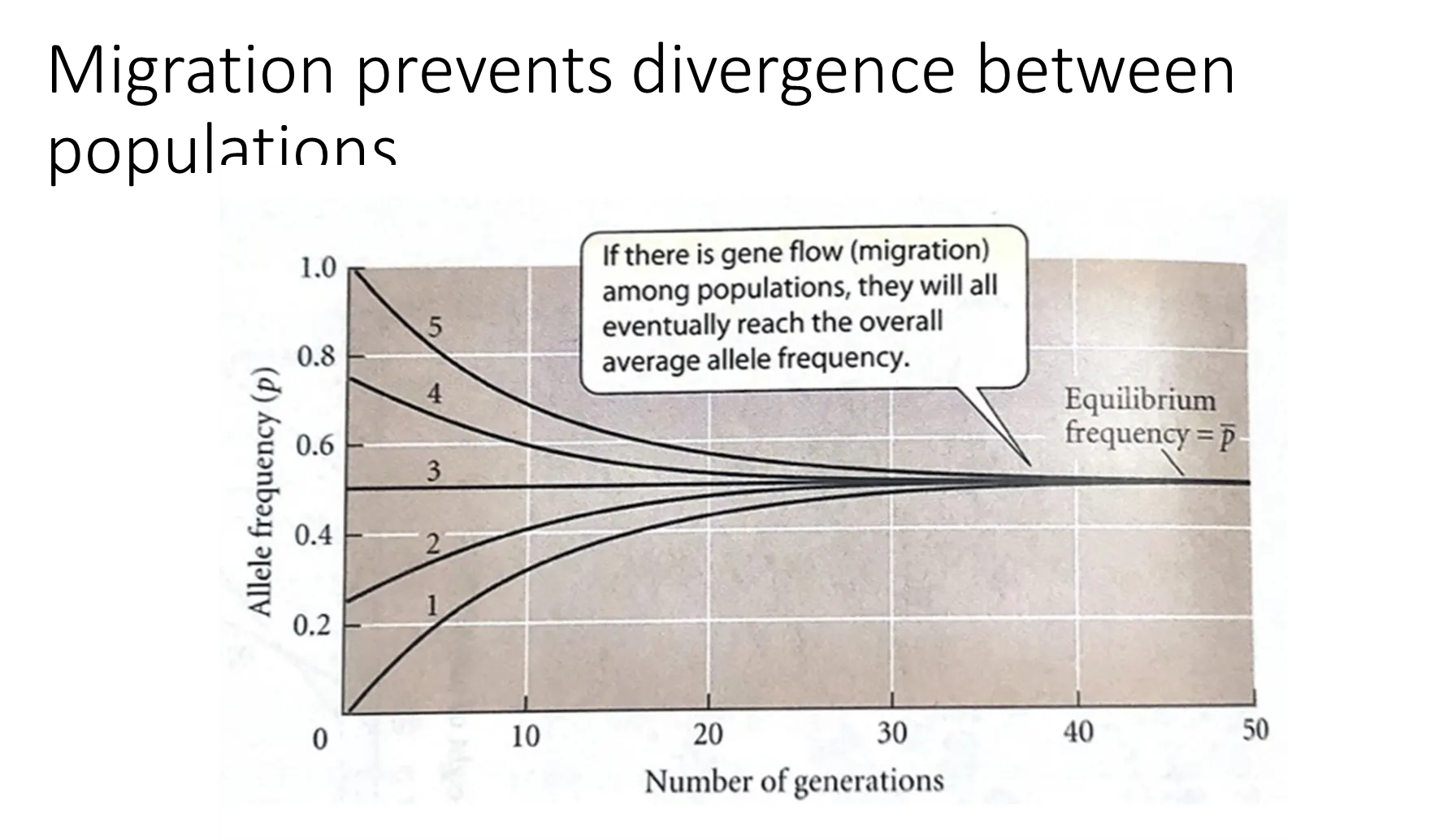
how does migration affect allele frequencies?
if there is gene flow between populations, they will all eventually reach an average allele frequency
the number of generations that this takes to occur is dependent on the rate of migration
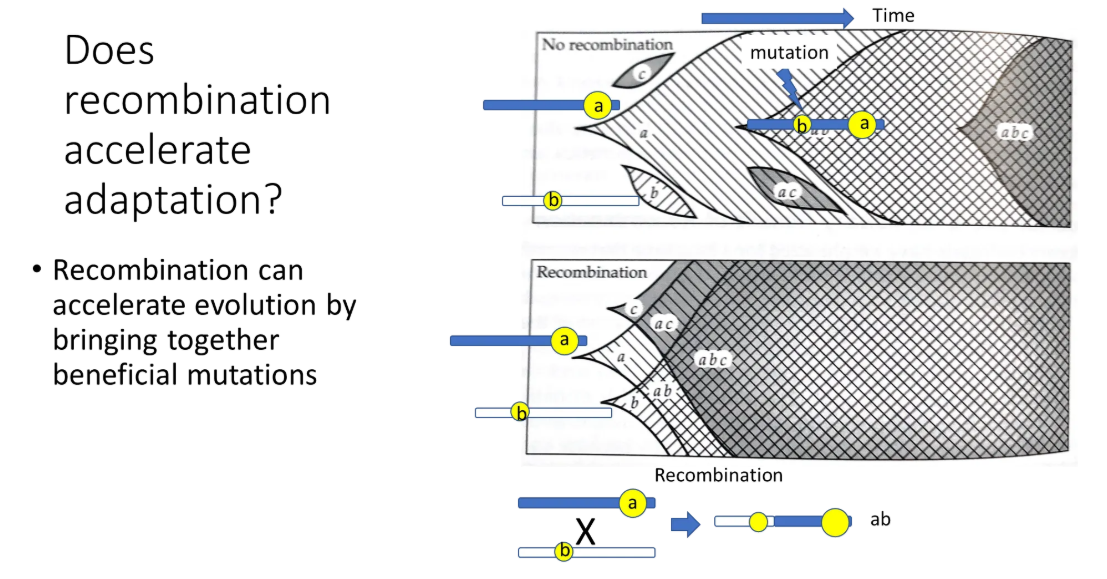
how does recombination affect adaptation?
recombination accelerates adaptation and evolution by bringing together beneficial mutations
without recombination (in a haploid population), two beneficial alleles would have to compete with each other, as they could never be inherited together
in the top diagram, without recombination, b and c are outcompeted by a, and ac is outcompeted by ab
in the bottom diagram, with recombination, evolution happens much faster because the beneficial mutations can combine
sexual reproduction creates much greater genetic diversity through recombination
what is hamilton’s rule of cooperation and how can we use it to classify social behaviours depending on the values involved?
an action will be favoured by natural selection when rb-c>0, where:
r is the fraction of relatedness between the actor and the recipient (proportion of own genes that will be passed on via the recipient’s offspring)
b is the benefit to the recipient (how many more offspring the recipient will be able to produce)
c is the cost to the actor (how many fewer offspring the actor will be able to produce)- if c is negative, the action also benefits the actor

what are direct, indirect and inclusive fitness?
direct fitness- the offspring the actor has
indirect fitness- the offspring the recipient has X the relatedness between the actor and recipient (how many of the actor’s genes will be passed down via the recipient’s offspring)
inclusive fitness- the sum of the direct and indirect fitness

what are fraternal and egalitarian cooperation and what are some examples?
fraternal cooperation is between cooperators that are closely related, eg:
the cells within a multicellular organism cooperate
the individuals within a population can cooperate (or may not, depending on degree of relatedness)
egalitarian cooperation is between cooperators of that are unrelated or distantly related, eg:
different genes contribute to a genome
different genomes (nuclear, mitochondrial…) contribute to a complex cell
populations of different species can cooperate
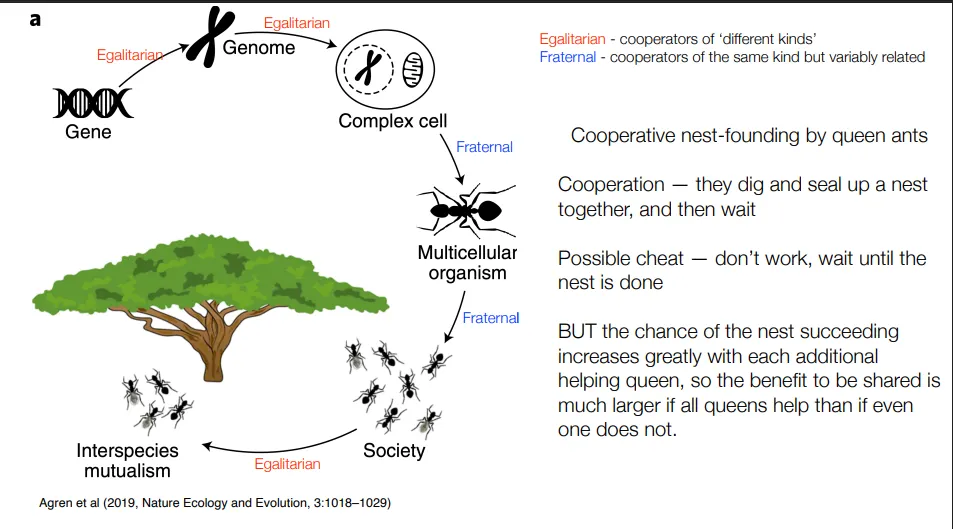
what is unenforced direct benefit cooperation? give an example
a kind of direct cooperation- no real reason/benefit to cheat
it’s just very beneficial to cooperate, so no enforcement is needed
eg. cooperative nest founding by queen ants:
burrowing through soil to form a nest is difficult, so it helps to have multiple queens doing so (they then fight to the death)
possible cheat- to not work and wait until the nest is done
but the chance of a successful nest increases greatly with each additional queen, so not cooperating isn’t beneficial
what is reciprocity cooperation? give an example
a kind of direct, enforced cooperation- two partners do repeated actions that help eachother with consequences for cheating
each action looks altruistic, but the exchange becomes direct benefit
eg. cleaner fish:
cleaner gets to feed on ectoparasites and don’t get eaten, big fish ‘client’ gets cleaned
possible cheat- client eats the cleaner, cleaner takes a bit of the client
but the client would then not find another cleaner, and the cleaner would lose clients, so there are consequences for breaking reciprocity
‘prisoners dilemma’
what is policing cooperation? give an example
a kind of direct, enforced cooperation- cheating is prevented by systematic action
eg. worker laying of male eggs in eusocial insects
queen mates with many males
this produces female workers which all collaborate to raise subsequent offspring as relatives
possible cheat- a laying worker could lay its own eggs (to produce males that can mate with the queen)
but worker-laid eggs get routinely removed by workers very effectively (because they have a greater regression relatedness to queen-laid eggs than worker-laid eggs in a multi-father population), so cheating is unsuccessful
the best route to passing on your genes is by raising queen-laid eggs, so slacking is also not beneficial
what is imposed incentive cooperation? give an example
a kind of direct,enforced cooperation- cooperation is encouraged by one individual to others via rewards, punishments and sanctions
eg. soybeans and Rhizobium
soybean plants supply Rhizobium bacteria with oxygen and carbohydrates in return for fixed nitrogen
possible cheat- Rhizobia could stop fixing nitrogen
but the soybean plants withhold oxygen from Rhizobium that do not fix enough nitrogen- this is an imposed incentive
what is limited dispersal cooperation? include the calculations
also called population viscosity
a theoretical kind of indirect cooperation- offspring don’t move far from where they were born, so they are surrounded by close relatives and can all help eachother
however, this has been shown to not be a valid theory, as the degree of relatives helping eachother is cancelled by the local resource competition between them
applied to hamilton’s rule:
an altruistic action is favoured by selection if rb-c > 0
however the gain to the recipient must be balanced by a cost (of b - c) to another group of individuals (which the actor may also be related to)
relatedness to this ‘density-dependent group’ is called r_e
so hamiltons rule becomes rb - c - r_e(b - c) > 0
if the cost is spread across the whole population (isn’t density dependent), r_e = 0, so hamilton’s rule stays the same
if the actor is closely related to the density-dependent group (in the case of limited dispersal), r = r_e, so the expanded rule cancels down to rc - c > 0
in this case, the benefit b to the recipient becomes irrelevant, so there is no reason to be altruistic (theory isn’t valid)
overall altruistic actions are favoured with low comp/high r (and vice versa):
high relatedness to the recipient, where the cost comes to a wider, less related population (global competition)
what is kin discrimination cooperation? give an example
a kind of indirect cooperation- individuals are able to recognise their relatives, so they can be altruistic to just their kin and pass on their genes indirectly
eg. long-tailed tits (and other birds)
long-tailed tits try to mate on their own, but if they fail, they try to help at another nest- most of the time they select a relative to help
the more helpful the individual is, the more likely they are to be discriminant in helping only relatives
what is greenbeard cooperation? give an example
a semi-theoretical kind of indirect cooperation- a gene that can cause the individual to have a distinctive trait (a green beard), recognise it in other individuals, and behave altruistically to other bearers
eg. invasive fire ants (potential example?)
workers carrying the b allele kill young queens which don’t have it (BB), relying on a chemical odour
what are the four species concepts?

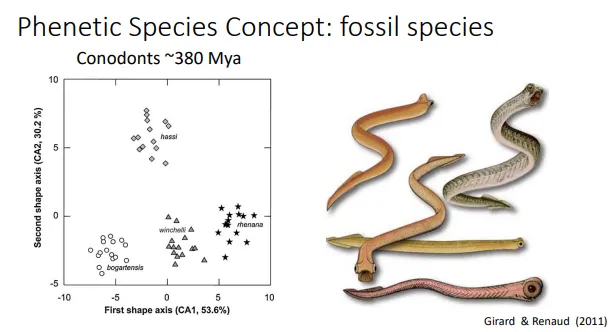
when/why is the phenetic species concept useful and what are its limitations?
defining species by phenotypic similarities is particularly useful for fossils eg. conodonts, as we can’t study the other species concepts properly
this is also used for some extant species eg. afrian gerbils- found in museum samples but hard to access in the wild
plotting measurements for two phenotypic traits (eg. length and width) against each other on a graph will show groupings for species- overlap may be present, but other traits can be applied to define the species
however:
this process may fail when you apply different traits or increase the sample, as they don’t cluster properly, and an be subjective beause it relies on human interpretation
this concept also doesn’t include an explanation for why the species are distinct
polymorphism- discontinuous variation can exist within species due to different forms- these polymorphs can have interbreeding and gene flow and can still be highly genetically similar eg. black bellied seedcracker bill size
plasticity- in some species, individuals may be able to change their phenotype (based on conditions at birth, not based of genetic differences) eg. deep sea vent tubeworms
cryptic species- multiple species can share one phenotype that aren’t immediately recognised as being distinct eg. neotropical skipper
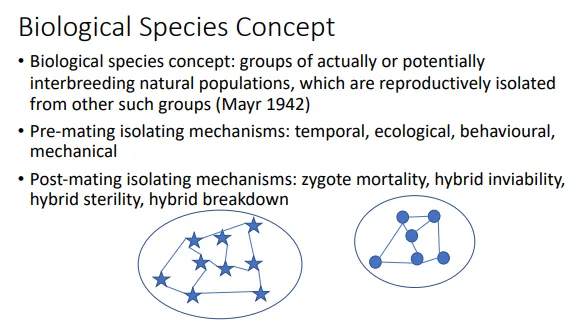
when/why is the biological species concept useful and what are its limitations?
defining species biologically is possible when two groups are reproductively isolated from each other, either by pre-mating (temporal, ecological, mechanical or behavioural eg. different mating calls) or post mating (hybrid mortality/sterility/inviability eg. mules) isolating mechanisms
this is helpful because it provides a mechanism for how/why the species are distinct
however:
it is difficult to know whether it would be possible for two species to mate, especially if their geographical ranges don’t overlap (allopatric), because we can’t necessarily force the two to mate- hybridisation could be possible even between what we thought were very distinct species
ring species- there is gene flow between adjacent populations around a geographical barrier (eg. a mountain range), but no gene flow between the species where the ring meets, which evolved in different directions around the barrier
this concept can’t be applied to fossils or to exclusively or alternately asexual species because reproduction must be observed
when/why is the phylogenetic species concept useful and what are its limitations?
this concept is useful because it focuses on the evolutionary relationships between distinct species
this can also be applied to fossils and asexual organisms
however:
a very high resolution of the phylogenetic tree is needed
taxonomic inflation- there is a tendency for the number of distinct species to continually increase with each new paper published, because it is hard to draw the lines, and each species could keep being divided up further and further (subjective)

when/why is the ecological species concept useful and what are its limitations?
this concept is very systematic, as adaptive zones (ecological niches) are defined apriori- before looking at the population itself
this can be applied to asexual organisms
it also allows for some gene flow/HGT between groups
however:
this assumes that ecologically discrete zones occur in nature with gaps in between, not as a continuous range
niche construction- certain species are able to change their environment to suit themselves, so they alter the adaptive zone they’re in
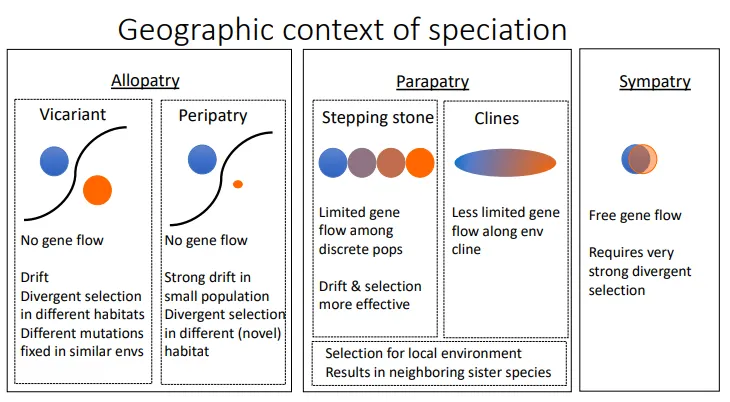
what are the different kinds of speciation?
allopatry- speciation when organisms are in two different places (geographically isolated) so there is no gene flow at all, and the populations experience divergent selection (selection against the median phenotype) in their separate habitats
vicariant- the populations are of similar sizes, a few methods may affect it
peripatry- one of the populations is much smaller, and so is strongly affected by drift
parapatry- gene flow between adjacent populations along a long scale may occur, resulting in neighbouring sister species
stepping stone- more discrete populations with limited instances of gene flow
clines- more gene flow along a spectrum
sympatry- free gene flow within a population in same geographical area, relies on very strong divergent selection, against intermediate phenotypes and towards fitness peaks (more common than previously thought)
what evidence is there for vicariant speciation?
diptera mating experiments- separated fly populations that are combined after multiple generations under different degrees of selection/drift exhibit behavioural isolation and some incomplete reproductive isolation
snapping shrimp- strong reproductive isolation and genetic/morphological differences observed between sister species that became separated by the emergence of the isthmus of panama, which created different habitats

what evidence is there for parapatric speciation?
evidence for parapatric speciation is quite weak, because it is difficult to rule out allopatric divergence in the past
ring species- gene flow between adjacent populations, but not between the populations at either end
little greenbul- there is a forest-savanna ecotone (cline), across which speciation is starting to occur eg. birdsong frequencies shifting through the different habitats (selection- the sound travels differently in open vs sheltered habitats)
M. guttatus- copper waste from a mine created patchily contaminated soils and copper pollution resistance evolved only within these patches- there was some gene flow between patches but very strong selection, now hybrids don’t perfom well in either habitat

what evidence is there for sympatric speciation?
there are few good examples of sympatric speciation, but it is theoretically possible
lord howe island palms- defined by the analysis of molecular-based phylogenies and genetic data
apple maggot fly- underwent a host shift from hawthorn to apple by assortative mating (adults tend to mate on the fruit they fed on as larvae) and incomplete temporal isolation (apples fruit earlier)
this may not be a good example due to recent evidence of a geographically separate southern species introgressing with one of the sympatric forms and causing genetic divergence

what are some other methods/examples of speciation?
speciation by hybridisation:
the ‘big bird’ lineage of G. fortis
an immigrant bred with a resident, resulting in offspring with an extreme bill phenotype that caused reproductive isolation within 3 generations, as the hybrid relatives only bred together (and inbreeding depression didn’t occur)
speciations due to polyploidy:
polyploidy individuals carry more than 2 sets of chromosomes
a change in the number of chromosome pairs that an individual carries can produce almost immediate reproductive isolation (common in plants)
what are two examples of selfish drivers that have little effect on the phenotype?
the t haplotype in mice:
this is a group of 300 genes on chromosome 17
it contains multiple inversions that suppress recombination, so they are inherited as a unit (a haplotype)
the offspring of a male that is heterozygous for the t haplotype will inherit the t haplotype 90% of the time (rather than 50%)
this is because the t haplotype sperm kill other sperm
homing endonuclease genes (HEG) in yeast:
in individuals that are heterozygous for HEG, the gene produces endonuclease enzymes that cut the non-HEG chromosome at a recognition site
this dna gets repaired using the HEG chromosome as a template, so that the organism is made homozygous for HEG
HEG frequencies don’t change in inbred homozygous HEG or non-HEG populations, because there are no heterozygous individuals
if the two populations outbreed together, the HEG gene frequency will increase rapidly because heterozygous offspring are produced, so HEG can self-promote and spread more
what is an example of a selfish driver that affects the phenotype?
sex ratio (proportion of males) drivers in Diptera:
the X chromosome sperm want all of the offspring to carry two X chromosomes, and the Y sperm want all of the offspring to carry an X and a Y chromosome
there are X drivers found in Diptera, which kill Y-bearing sperm- any female that mates with an X driver male will produce more female offspring (and vice versa with Y drivers)
X drivers lead to a female-biased sex ratio- they are selfish because they do this at a cost to the rest of the genome
in a female-biased population, males are more valuable, and so the average male has more chances to pass on their genes than the average female- males are more fit than females
an X driver-bearing male will produce more females, which are less fit, so the rest of the father’s genome is hindered
why are sex ratio drivers rare?
sex ratio drivers are rare, because:
when females mate with multiple males, X driver-bearing males have a lower sperm success, because non-driver males will produce more viable sperm and have more mating success
the Y chromosome often evolves suppression for the driver
the autosomes also evolve suppression for the driver, because the X driver comes at a cost to the rest of the genome (female bias makes males more successful), so suppression would be selected for- ‘the parliament of the genes’ suppress selfish genes
real populations don’t really have sex ratio biases, not because the drivers aren’t present, but because the suppressors are
conflict is often hidden, but can be detected by crossing between species/populations, which separates the drivers from their suppressors
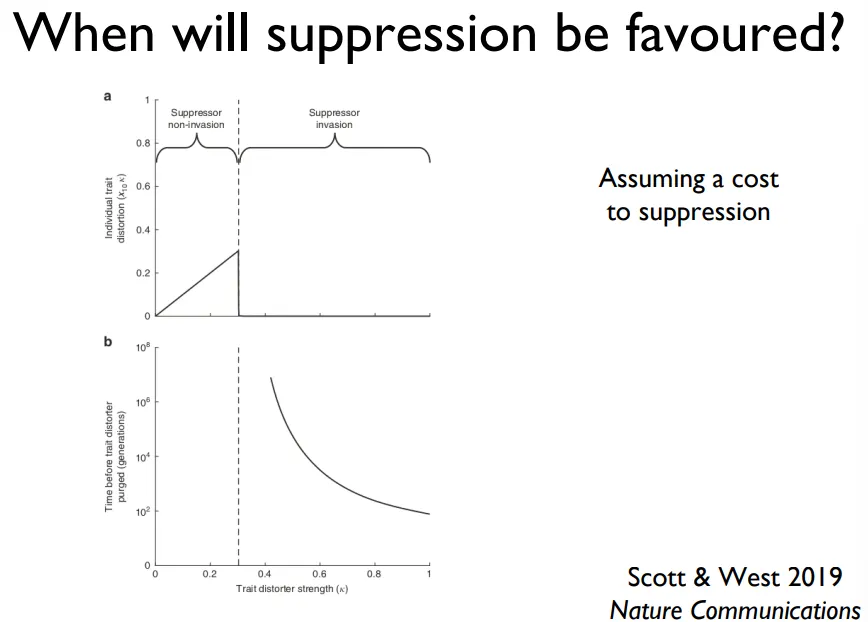
why are sex drivers not always suppressed?
suppression comes at a cost, so they are only present if the driver causes a significant distortion of the sex ratio
as the drivers distortion strength increases, distortion increases up until a point where it drops to 0, when suppressors suddenly become worth the cost
selfish genes either produce small effects that aren’t suppressed or large effects that are suppressed
selfish genes evolve, but are over-ambitious- they will drive themselves into situations where they then get suppressed