Kap. 14
1/40
Earn XP
Description and Tags
Biochemistry II
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
41 Terms
Preparatory phase
1-5: Glucose → 2 Glyceraldehyde 3-phosphate (G3P)
Requires 2 ATP
Payoff phase
6-10: 2 Glycosealdehyde 3-phosphate (G3P) → 2 pyruvate
Gives off 2 NADH & 4 ATP
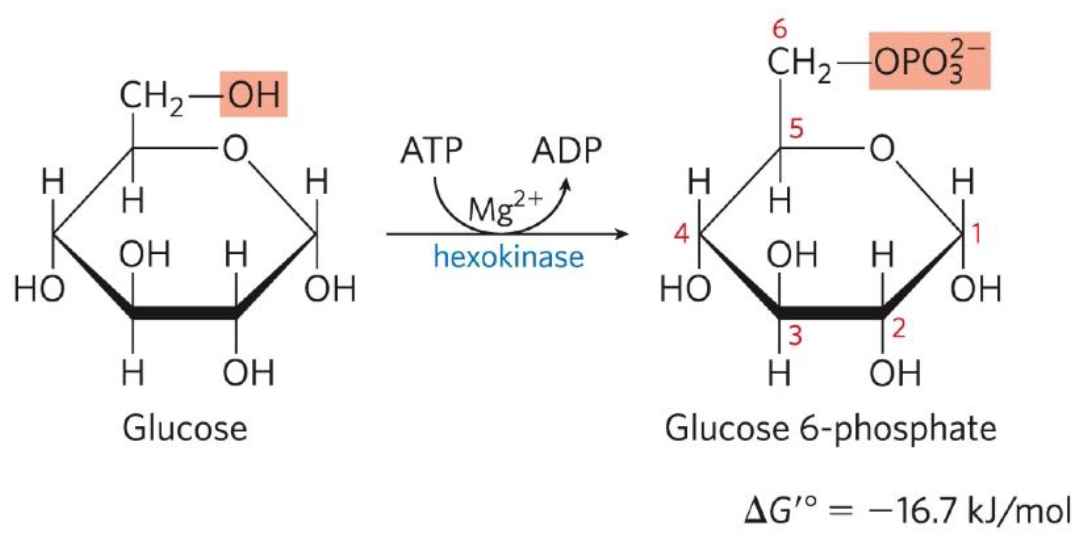
Phosphorylation of Glucose
First reaction: Glucose + ATP → Glucose 6-phosphate (G6P) + ADP
Catalysed by hexokinase, by phosphorylation with ATP as a phosphorylation donor.
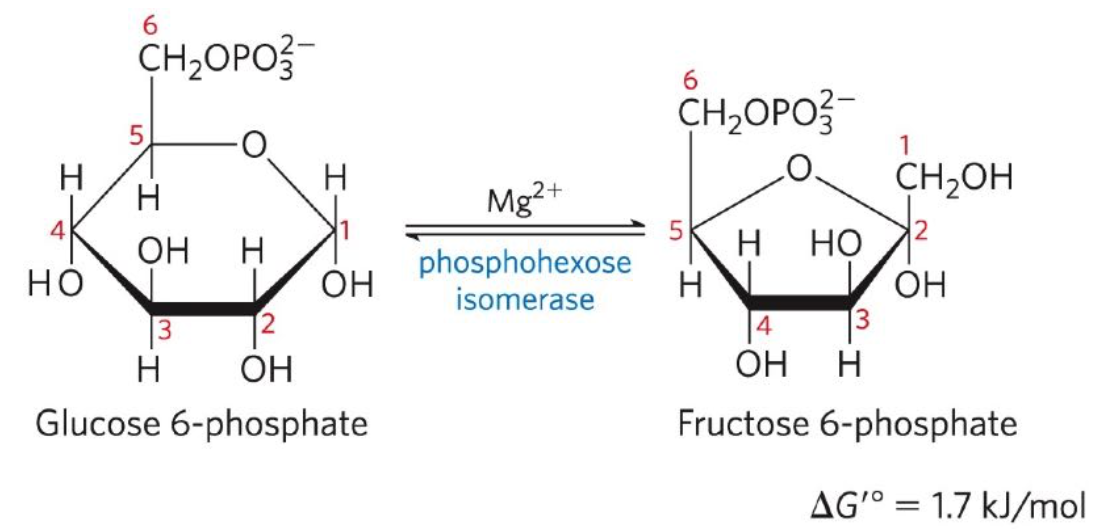
Conversion of G6P to F6P
Second reaction: Glucose 6-phosphate (G6P) ←→ Fructose 6-phosphate (F6P)
Phosphohexose isomerase catalyzes the reversible isomerization of G6P (aldose) to F6P (ketose)
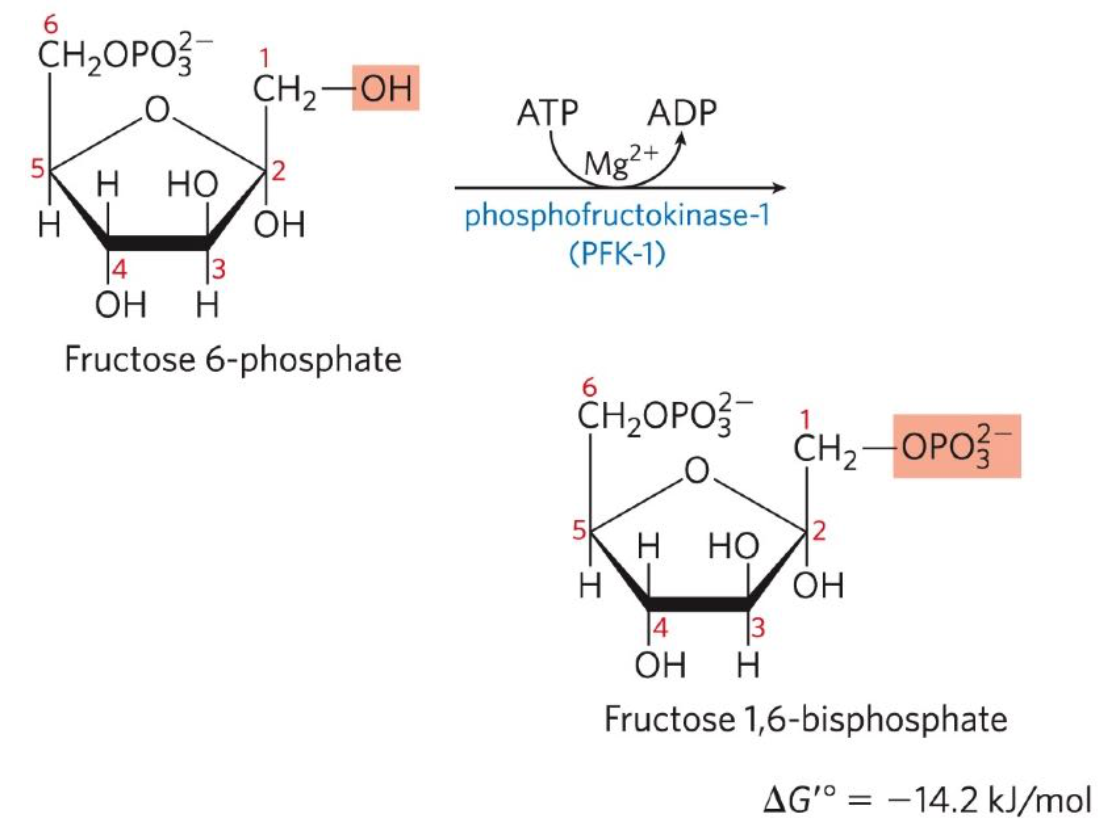
Phosphorylation of F6P to F-1,6-BP
Third reaction: Fructose 6-phosphate + ATP → Fructose-1,6-Bisphosphate + ADP
Phosphofructokinase-1 (PFK-1) catalyses the transfer of F6P to yield F-1,6-BP
Cleavage of F-1,6-BP
Fourth reaction: Fructose-1,6-Bisphosphate ←→ Dihydroxyacetone phosphate + Glyceraldehyde 3-phosphate (G3P)
F-1,6-BP aldolase (aldolase) catalyses a reverse aldol condensation
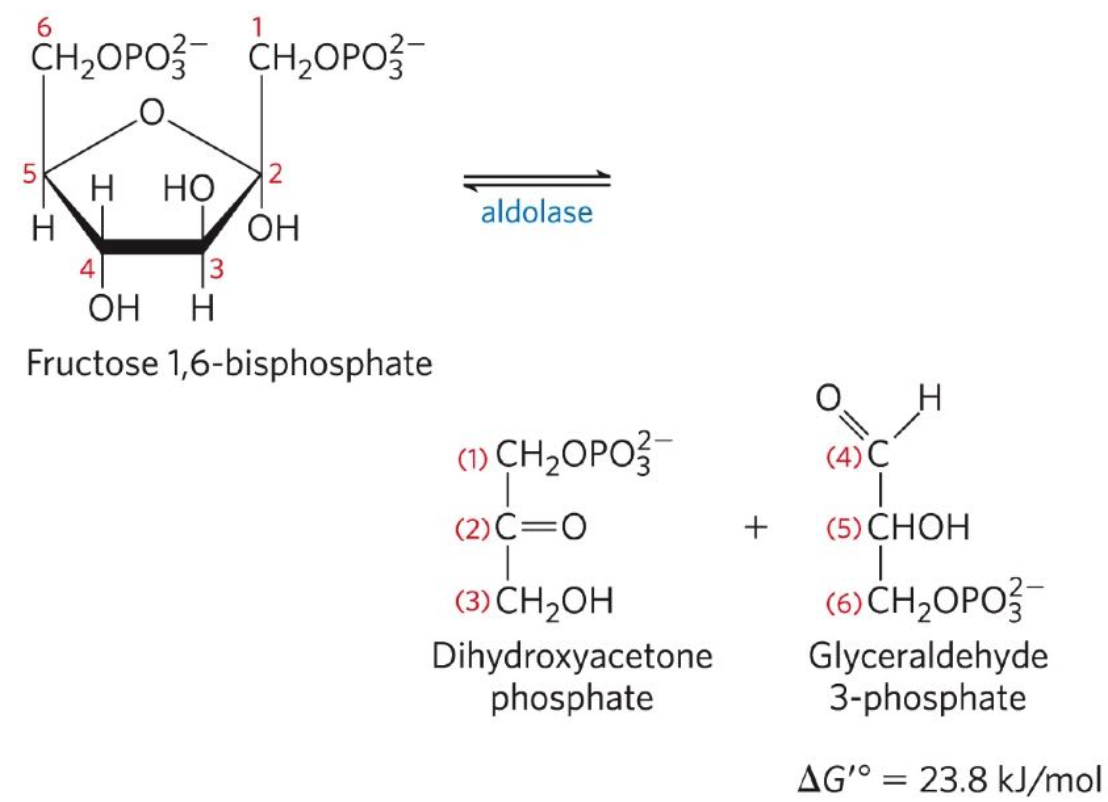
Interconversion of the Triose phosphate
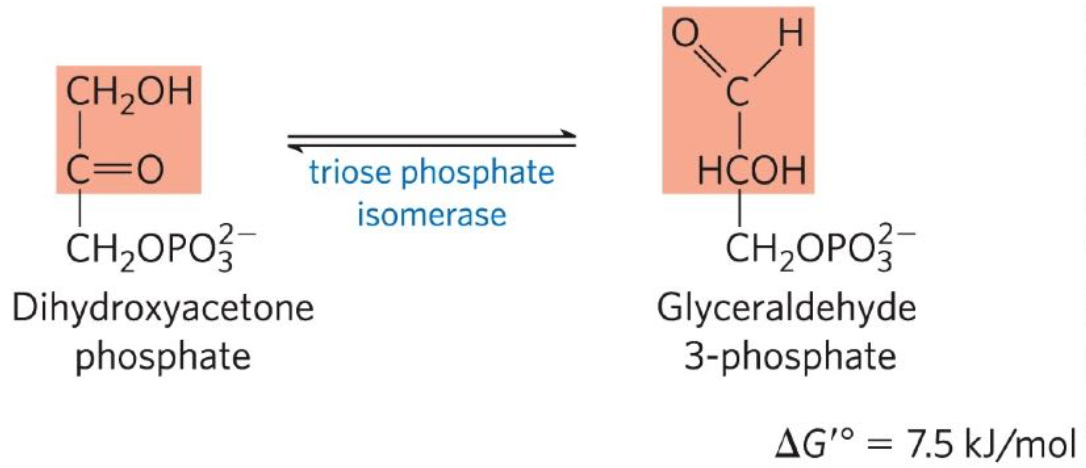
Fifth reaction: Dihydroxyacetone phosphate ←→ Glyceraldehyde 3-phophate (G3P)
Triosphosphate phosphate isomerase reversible convert 1 triode phosphate to G3P
Oxidation of G3P to 1,3-Bisphosphoglycerate
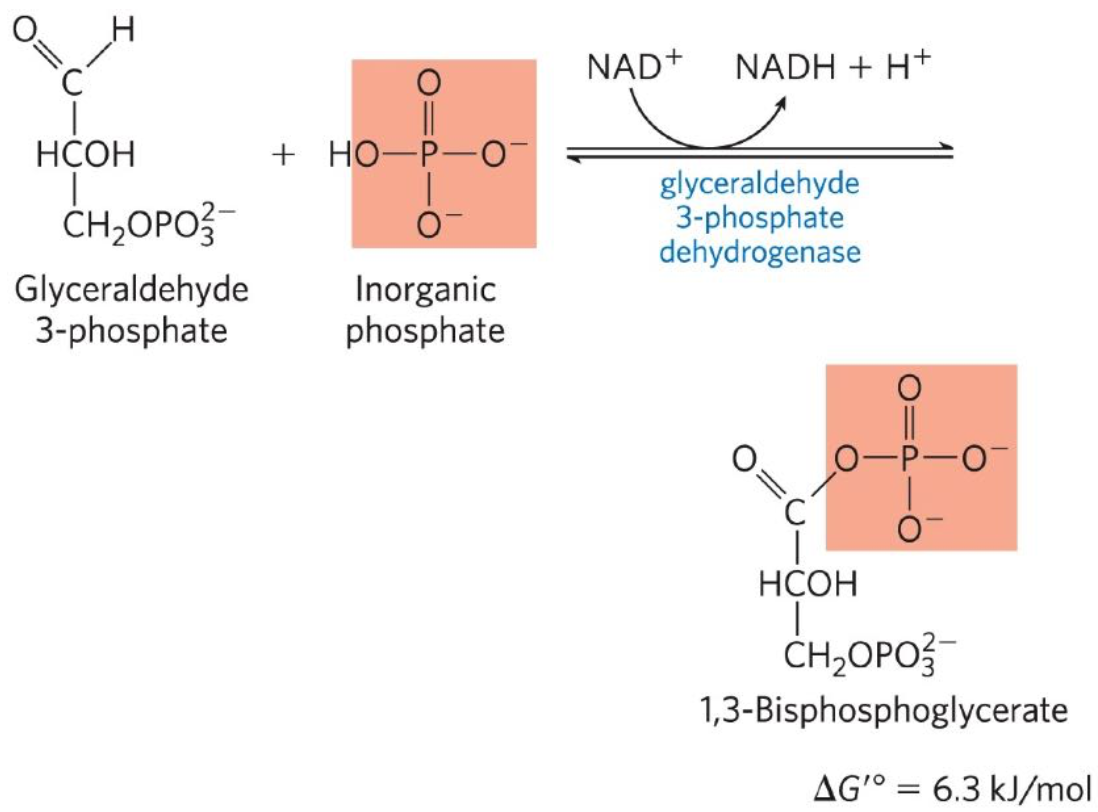
Sixth reaction: Glyceraldehyde 3-phosphate (G3P) + Inorganic phosphate + NAD+ ←→ 1,3-Biphosphoglycerate (1,3-BPG) + NADH + H+
Oxidation of G3P catalyzed by gluceraldehyde 3-phosphate dehydrogenase
Phosphoryl transfer from 1,3-BPG to ADP
seventh reaction: 1,3-Bisphospoglucerate + ADP ←→ 3-phosphoglucerate + ATP
Phosphoglycerate kinase transfers the high-energy Phosphoryl group to ADP.
Substrate level phosphorylation
Conversion of 3-phosphoglycerate to 2-phosphoglycerate
Eight reaction: 3-phosphoglycerate ←→ 2-phosphoglycerate
Phosphoglycerate mutase catalyzes the reversible shift of phosphorylation group
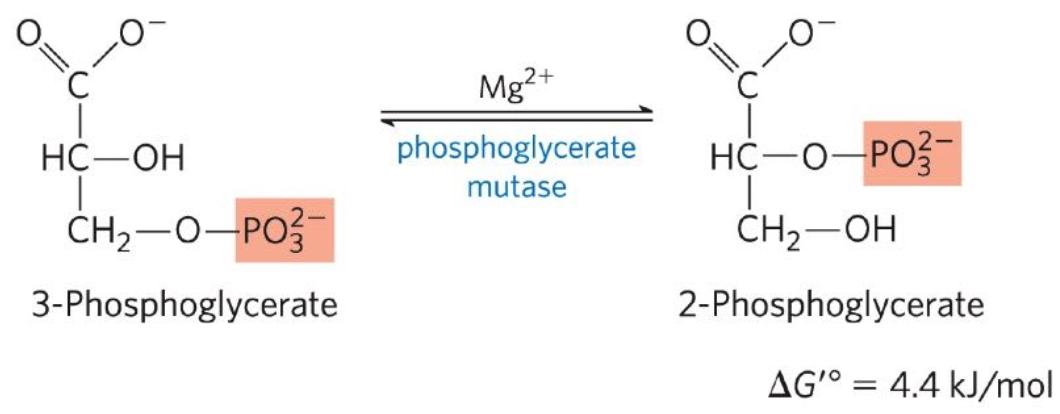
Dehydration of 2-phosphoglycerate to PEP
Ninth reaction: 2-phosphoglycerate ←→ phosphenolpyruvate (PEP)
Enolase promotes reversible removal of H2O
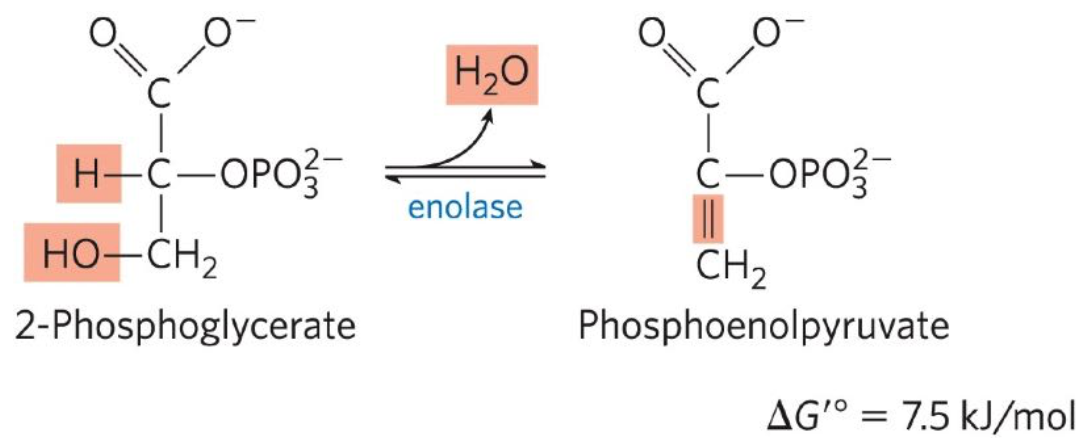
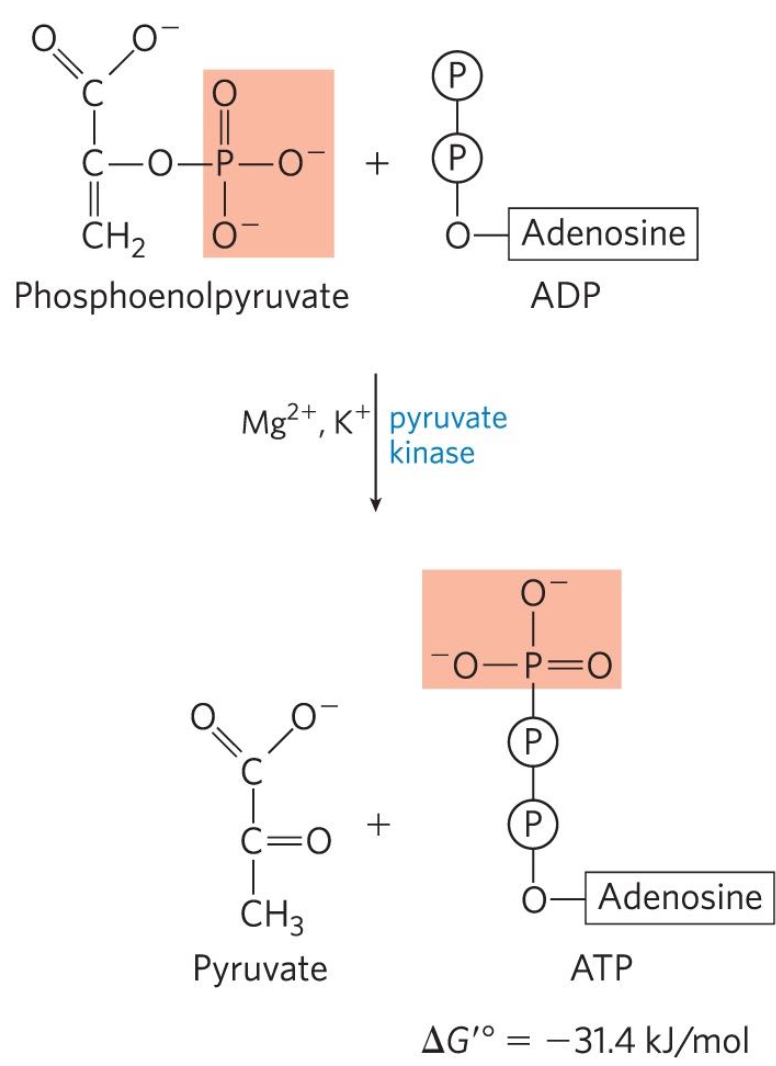
Transfer of phosphoryl group from PEP to ADP
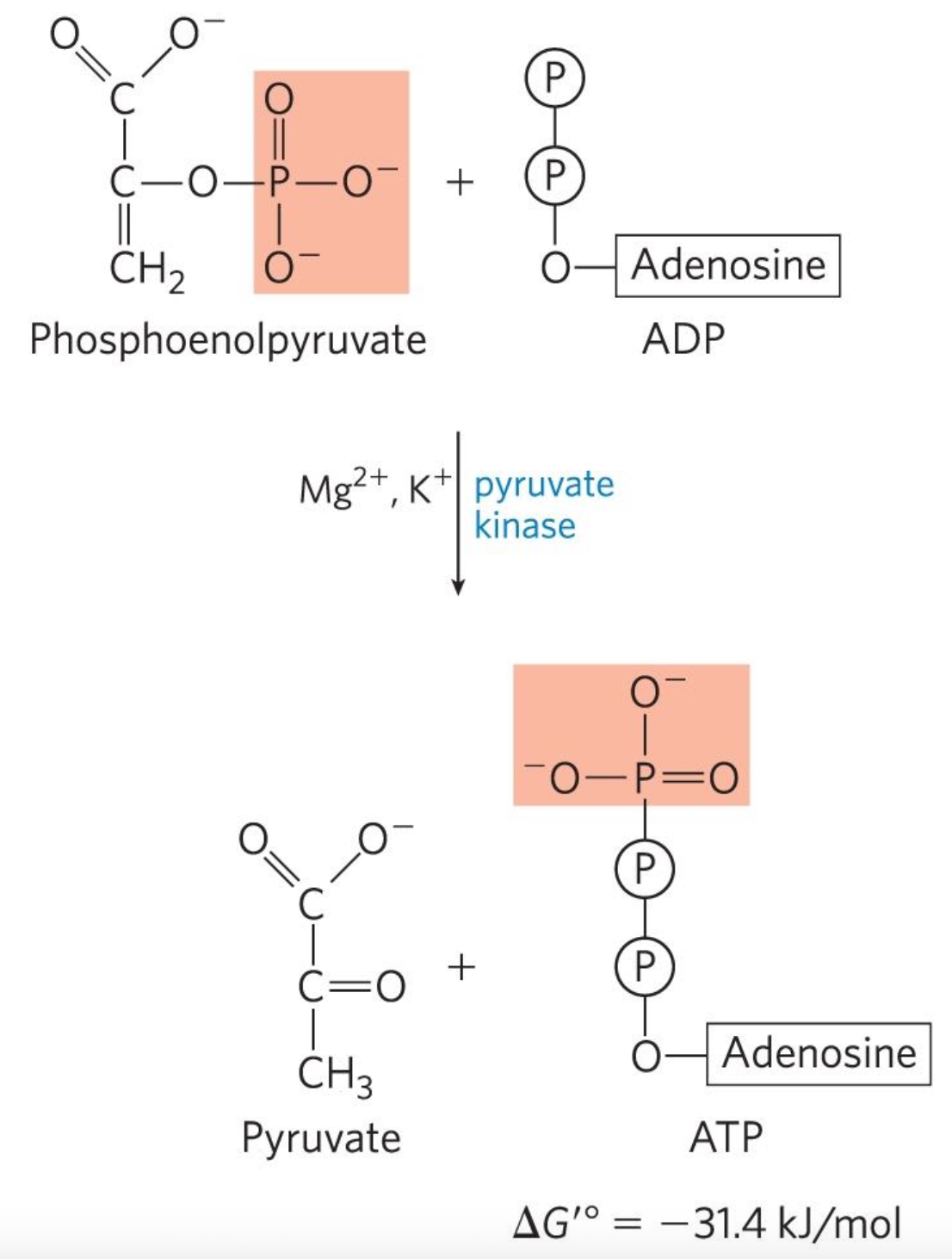
Tenth reaction: Phosphenolpyruvate (PEP) + ADP → Pyruvate + ATP
Pyruvate kinase catalyzes the transfer
Substrate level phosphorylation
Hexokinase
First glycolysis reaction enzyme:
Catalyzes the phosphorylation of hexoses - glucose, mannose & fructose
Requires Mg2+ as it shields the negative charges of P-groups in ATP making the terminal P an easy target for nucleophilic attack by -OH from a hexose.
Hexokinase undergoes shape change: 2 domains get closer when ATP binding, bringing ATP closer to a hexose also bound by the enzyme & blocks H2O which could disrupt the charges.
Phosphofructokinase - 1 (PFK-1)
Third glycolysis reaction enzyme:
PFK-1 is under complex allosteric regulation, activity increasing ac ATP is low or when its products (ADP & AMP) is accumulated.
Is inhibited when ATP levels are high (negative feedback)
Substrate-level phosphorylation
A metabolic process that directly forms ATP (or GTP) by transferring a phosphate group from a high-energy phosphorylated metabolic intermediate (the substrate) to ADP (or GDP).
Respiration-linked phosphorylation
= Oxidative phosphorylation
Cells harness the energy from oxidized nutrients to generate Adenosine Triphosphate (ATP) by driving electrons through an electron transport chain and utilizing a subsequent proton gradient.
Endogenous Glycogen and Starch are degraded by phosphorolysis
Stored glycogen is mobilized by phosphorolytic reactions:
Glycogen (or starch) → Glucose 1-phosphate
Catalyzed by glycogen phosphorylase
Glucose 1-phosphate ←→ Glucose 6-phosphate
Converted by phosphoglucomutase
G6P can continue through glycolysis or the pentose phosphate pathway
G6P → fructose 6-phosphate…
Dietary polysaccharides and disaccharides undergo hydrolysis to monosaccarides
Dietary glycogen (or starch) + H2O → Glucose
Digestion starts in the mouth where salivary Alpha-amylase hydrolyses the internal glycosidic linkages, producing di- and trisaccharides (hydrolysis reactions)
Salivary alpha-amylase is inactivated in the stomach (low pH), but an alpha-amylase from the pancreas is released into the small intestine. Pancreatic alpa-amylase yields limit dextrins (di- & trisaccharides of glucose/oligosaccharides), which are fragments of amylopectin, removed by limit dextrinases.
Only monosaccharides are taken up from the intestine to be catabolized by glycolysis.
Lactose → glucose and galactose
Converted by the enzyme lactase
Galactose + UTP → Glucose 1-phosphate
Galactose passes from the blood to the liver where it is first phosphorylated at the expense of ATP by galactokinase to glucose 1-phosphate.
Glucose 1-phosphate is then converted to glucose 6-phosphate by phosphoglucomutase (as before).
Glycolysis of fructose & mannose
Fructose + ATP → Fructose 6-phosphate + ADP
Phosphorylated by hexokinase
Fructose + ATP → fructose 1-phosphate + ADP
Catalyzed phosphorylation by fructokinase
F1P is then cleaved to glyceraldehyde and dihydroxyacetone phosphate by F1P aldolase.
Mannose + ATP → mannose 6-phosphate + ADP
Phosphorylated by hexokinase
Mannose 6-phosphate is then converted to F6P of glycolysis, by phosphohexose isomerase
Lactose digestion & intolerance
Mammary glands produce lactose for the nourishment of infants.
Lactase converts lactose to glucose and galactose, both of which are taken up in the small intestine and metabolized by glycolysis.
Lactase persistance phenotypes can be seen in adults
They continue to produce lactase and are therefore able to digest milk into adulthood.
Lactose intolerance
Seen as lactase activity decreases with age, without intestinal lactase, lactose cannot be completely absorbed and digested in the small intestine.
It will pass to the large intestine, where bacteria convert it to toxic products that cause abdominal cramps/diarrhea.
Aerobic fate of pyruvate
The pyruvate formed in the last step of glycolysis is oxidized to acetate (acetyl-CoA) by PDH complex, which enters the citric acid cycle and is oxidized completely to CO2 & H2O
Anaerobic fate of pyruvate
Oxygen is needed to deoxidize NADH to NAD+ for glycolysis to continue, in these conditions glucose is degraded by: fermentation.
Lactic acid fermentation:
Pyruvate accepts electrons from NADH and is reduced to lactate while generating NAD+ necessary for glycolysis.
Ethanol (alcohol) fermentation:
Pyruvate is further catabolized to ethanol
Lactic acid fermentation without mitochondria
NAD+ is regenerated from NADH by the reduction of pyruvate to lactate
Some tissues/cell types (have no mitochondria and thus cannot oxidize pyruvate to CO2), produces lactate from glucose even under aerobic conditions.
Pyruvate catalyzed by lactate dehydrogenase.
Overall reaction eq. favors locate formation.
The Pasteur effect
Both the rate and the total amount of glucose consumption under anaerobic conditions is much higher than in aerobic
The Warburg effect
Cancer cells preferably produce energy by consuming glucose and breaking it down to lactate, even in aerobic conditions.
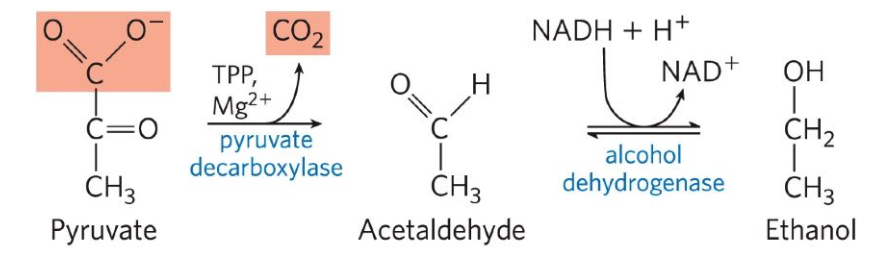
Ethanol fermentation
Glucose is metabolized to pyruvate, which is converted to ethanol and CO2 in a two-step process:
Pyruvate is decarboxylated to form acetaldehyde in an irreversible reaction catalyzed by pyruvate decarboxylase
Acetaldehyde is reduced to ethanol through the action of alcohol dehydrogenase, with the reducing power furnished by NADH derived from dehydrogenation of G3P.
Gluconeogenesis
Synthesis of glucose from noncarbohydrate precursors (pyruvate, lactate & glycerol, as well as certain a.a.)
Regulated differently in different tissue, following will be in mammal liver
7 of the 10 glycolysis reaction in reverse
Not: 1, 3 & 10 (they have large negative free-energy change, the others ~0)
The irreversible steps are bypassed by a separate set of enzymes
Energetically expensive, but essential.
Gluconeogenesis: bypass reactions
Conversion of pyruvate to phosphoenolpyruvate (PEP) via 2 exergonic reactions
Pyruvate is first transported from the cytosol into mitochondria (or generated in mitochondria by alanine transamination), then pyruvate carboxylase (which requires the coenzyme biotin) converts pyruvate to oxaloacetate.
Pyruvate carboxylase requires acetyl-CoA as a positive effector, acetyl-CoA is produced by fatty acid oxidation.
This reaction can replenish intermediates in the citric acid cycle
Mitochondria have no transporter for oxaloacetate, therefore it needs to be reduced to malate by malate dehydrogenase at the expense of NADH. In the cytosol the malate is reoxidized to oxaloacetate
Oxaloacetate is then converted to PEP by phosphoenolpyruvate carboxykinase
Dephosphorylation of fructose 1,6-bisphosphatase to fructose 6-phophatase
Catalyzed by Mg2+ - dependent fructose 1,6-bisphosphatase (FBPase-1)
Fructose 1,6-Bisphosphate + H2O → fructose 6-phosphate + Pi
Dephosphorylation of glucose 6-phosphate to glucose
Catalyzed by glucose 6-phosphatase does not require ATP (which reversal of hexokinase would), as it is a simple hydrolysis of a phosphate ester
Glucose 6-phosphate + H2O → glucose + Pi
Isozyme
Different forms of the same enzyme that catalyze the same reaction but is encoded by different genes, ex. hexokinase isozymes.
Hexokinase isozyme regulation
Mucle hexokinase I & II are allosterically inhibited by their product (G6P)
When G6P increases → isozymes temporarily inhibited, bringing balance to G6P concentration
Liver hexokinase, predominantly hexokinase IV (glucokinase), differs from the other hexokinase isozymes:
Glucose conc. for hexokinase half saturation is higher than the rest. Pga glucose transporter in hepatocytes (GLUT2) rapidly eq. the glucose conc. in cytosol & blood
Glucose is high = Excess glucose is transported into hepatocytes where hexokinase IV converts it to G6P
Low glucose = low conc. in hepatocytes relative to the Km of hexokinase IV, and the glucose generated by gluconeogenesis leaves the cell before being trapped
Hexokinase IV is not inhibited by G6P, can therefore continue to operate when the accumulation of G6P inhibits the other hexokinases
Hexokinase IV is subject to inhibition by reversible binding of a regulatory protein specific to liver
Binding is much tighter in the presence of the allosteric effector F6P, glucose competes with F6P for binding & causes dissociation of the regulatory protein from the hexokinase releving the inhibition.
High glucose → enters hepatocyte via GLUT2 and activates hexokinase IV by this mechanism
Low glucose (fast) → F6P triggers the inhibition of hexokinase IV by the regulatory protein, so the liver doese’t compete with other organs for scarce glucose.
The protein anchors hexokinase IV inside the nucleus, where it is segregated from the other enzymes of glycolysis in the cytosol.
Glucose increases in the cytosol, it equilibrates with glucose in the nucleus by transport through nuclear pores
Glucose cause dissociation of the regulatory protein, and hexokinase IV enters the cytosol and begins to phosphorylate glucose.
Hexokinase IV is also regulated by protein synthesis:
Circumstances that call for greater energy production (exercise) or higher glucose consumption cause increased transcription of hexokinase IV gene
G6P, the gluconeogenic enzyme that bypass the hexokinase step of glycolysis, is transcriptionally regulated by factors that call for increased production of glucose
Phosphofructokinase-1 & Fructose 1,6-Bisphosphate reciprocal regulation
Glucose 6-phosphate can flow either into glycolysis or other pathways (ex. glycogen synthesis or pentose phosphate pathway.
PFK-1 catalyses the reaction that commits glucose to glycolysis
Has several regulatory sites (allosteric activators/inhibitors)
ATP: a substrate for PFK-1 & end product of the glycolytic pathway
High ATP → inhibits PFK-1 by binding to an allosteric site and lowering the affinity of the enzyme for F6P.
Low ATP → high AMP/ADP, act allosterically to relieve ATP inhibition → high PFK-1 activity.
Citrate (key intermediate in aerobic oxidation of pyruvate/FA/a.a.) is also a allosteric regulator of PFK-1:
High citrate → Increases ATP inhibitory effect, further reducing the flow of glucose through glycolysis.
Citrate serves as an intracellular signal that the cell is meeting its current needs for energy-yielding metabolism by the oxidation of fats & proteins.
The corresponding step in gluconeogenesis is the conversion of F1,6BP → F6P
FBPase-1 catalyzes the reaction & is strongly inhibited (allosterically) by AMP, when ATP is low
Opposing steps in the glycolytic pathway & gluconeogenesis are catalyzed by PFK-1 & FBPase-1. AND regulated in a coordinated & reciprocal manner.b
Fructose 2,6-Bisphosphate is a potent allosteric regulator of PFK-1 & FBPase-1
Rapid hormonal regulation of glycolysis & gluconeogenesis is mediated by fructose 2,6-bisphosphate = allosteric effector for PFK-1 & FBPase-1.
F2,6-BP binds to PFK-1 allosteric site → increases the enzymes affinity for F6P (its substrate) & reduces its affinity for the allosteric inhibitors ATP/citrate
PFK-1 is virtually inactive in the absence of F2,6-BP
F2,6-BP has the opposite effect on FBPase-1: it reduces its affinity for substrate, thereby slowing gluconeogenesis.
Coordination of glucose levels in the liver
Blood glucose decreases → hormone glucagon signals the liver to produce & release more glucose + stop glucose consumption.
Glucose sources:
Glucogen = a source of glucose stored in the liver
Gluconeogenesis = using pyruvate, lactate, glycerol & certain a.a.
High blood glucose → insulin signals the liver to use glucose as a fuel and as a precursor for the synthesis & storage of glycogen
Fructose 2,6-Bisphosphate regulation
The cellular conc. of F2,6-BP is decided by the relative rates of its formation & breakdown
Formation: By phosphorylation of F6P, catalyzed by phosphofructokinase-2 (PFK-2)
Broken down: By fructose 2,6-Bisphosphatase (FBPase-2)
PFK-2 & FBPase-2 are two separate enzymatic activities of a single, bifunctional protein
The balance between the two in the liver is set by glucagon & insulin.
Fructose 2,6.Bisphosphate regulated by glucagon & insulin
Glucagon:
Stimulates adenylyl cyclase of liver → producing cAMP from ATP
cAMP activates cAMP-dependent protein kinase, which transfers a phosphoryl group from ATP to the bifunctional protein PFK-2/FBPase-2
Phosphorylation of this protein enhances FBPase-2 activity & inhibits PFK-2
Glucagon thereby: lowers the cellular level of fructose 2,6-Bisphosphate → inhibiting glycolysis & stimulating gluconeogenesis.
Insulin: Opposite effect
Stimulates the activity of a phosphoprotein phosphatase that catalyzes removal of the phosphoryl group from the bifunctional protein PFK-2/FBPase-2,
Activating its PFK-2 activity → increasing fructose 2,6-bisphosphate levels, stimulating glycolysis, & inhibiting gluconeogenesis.
Fructose 2,6-Bisphosphate regulation by Xylulose 5-phosphate (FA metabolism)
Xylulose 5-phosphate = product of PPP
Mediates the increase in glycolysis that follows ingestion of a high-carbohydrate meal.
Xylulose 5-phosphate increase as glucose enters the liver & is converted to G6P, entering both glycolysis & PPP
Xylulose 5-phosphate activates phosphoprotein phosphatase 2A, which dephosphorylates the bifunctional PFK-2/FBPase-2 enzyme
Dephosphorylation activates PFK-2 and inhibits FBPase-2, and the resulting rise in F2,6-BP conc. stimulates glycolysis and inhibits gluconeogenesis
Increases acetyl-CoA & NADPH
Pyruvate kinase
Is allosterically inhibited by ATP
High conc. of ATP, acetyl-CoA & long fatty-chain fatty acids → allosteric inhibition of all pyruvate kinase isozymes .
Liver isozyme (L form) is subject to further regulation by phosphorylation, not muscle isozyme (M form).
Low blood glucose → glucagon release,
cAMP-dependent protein kinase phosphorylates the L isozyme inactivating it → slowing glucose use, sparing it for brain
Muscle isozyme:
In response to epinephrine → cAMP activates glycogen breakdown and glycolysis, providing fuel needed for fight or flight.
Pyruvate → phosphophenolpyruvate (PEP)
= Conversion stimulated when FA are available:
FA readily available → breakdown to acetyl-CoA in mitochondria → acetyl-CoA signals that further oxidation of glucose.
Acetyl-CoA = positive allosteric modulator of pyruvate carboxylase & negative modulator of pyruvate dehrydogenase
Energy needs are met => oxidative phosphorylation slows, NADH conc. rises & inhibits citric acid cycle & acetyl-CoA accumulates.
Increased acetyl-CoA → inhibits pyruvate dehydrogenase complex → slowing acetyl-CoA formation & stimulates gluconeogenesis by activating pyruvate carboxylase → forming oxaloacetate
Oxaloacetate → converted to PEP, by PEP carboxkinase
Fasting/high glucagon → increase the transcription rate & stabilizes mRNA, acting through cAMP
Insulin/high glucose → opposite effect
ChREBP
= Carbohydrate response element binding protein
TF important to carbohydrate metabolism
Primarly expressed in the liver/adipose tissue/kidney
Coordinates the synthesis of enzymes needed for ex. FA metabolism
Is inactive in its phosphorylated form
Xylulose 5-phosphate regulates the protein by controlling the activity of PP2A (that activates/ phosphorylates ChREBP)
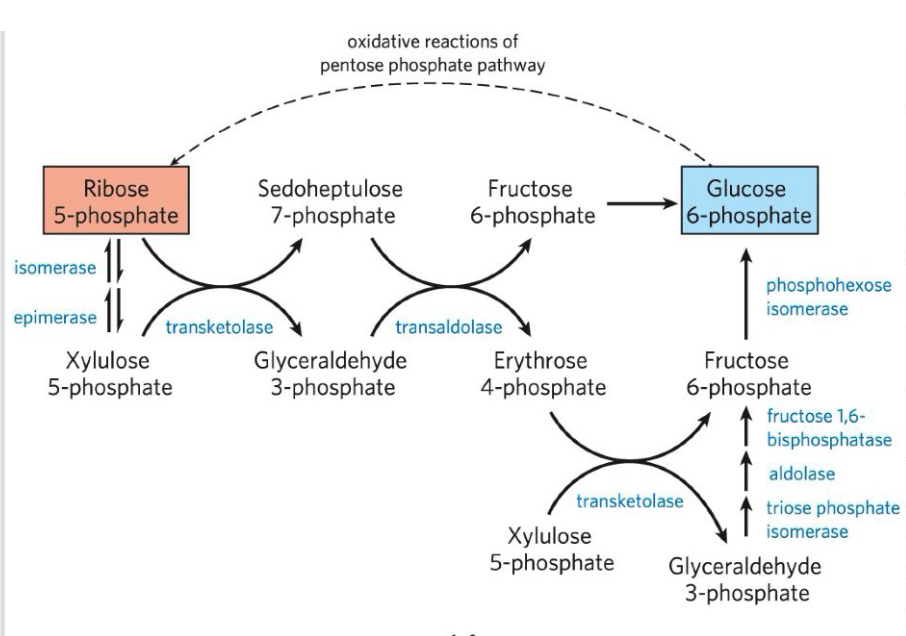
Pentose phosphate pathway (PPP) - overview
= Oxidative pathway, yielding ribose 5 phosphate & NADPH needed in other pathways.
G6P → ribose 5-phosphate
Has an non-oxidative phase: ribulose 5-phosphate → G6P
Oxidative phase of PPP
Oxidation of glucose 6-phosphate (G6P) + NADP+ → 6-phosphoglucono-gamma-lactone + NADPH + H+
By glucose 6-phosphate dehydrogenase (G6PD) & NAD+ as electron acceptor
Hydrolysation of 6-phosphoglucono-gamma-lactone + H2O → 6-phosphogluconate
By Lactonase
Oxidation & decarboxylation of 6-phosphogluconate + NADP+ → ribulose 5-phosphate + NADPH + H+ + CO2
By 6-phosphogluconate dehydrogenase
Conversion of ribulose 5-phosphate → ribose 5-phosphate
By phosphopentose isomerase
Nonoxidative phase of PPP
= Recycles pentose phosphates to G6P
Epimerization of ribulose 5-phosphate → xylulosze 5-phosphate
By ribulose 5-phosphate epimerase
Then in a series of rearrangements of the carbon skeletons, six five-carbons sugar phosphates are converted to five six-carbon sugar phosphates, completing the cycle.
Continued recycling leads ultimately to the conversion go G6P to 6 CO2.
2 unique enzymes act in these interconversions:
Transketolase - catalyzes the transfer of a two-carbon fragment from a ketose donor to an aldose acceptor
Requires the cofactor thiamine pyrophosphate (TPP)
Transaldolase - catalyzes the reaction similar to the aldolase reaction in glycolysis.
Uses Lys side chain to form Schiff base with the carbonyl group of its substrate.
Oxidative PPP 1st & 3rd step are irreversible.
Nonoxidative reactions are readily reversible