Kaarten: Moleculaire biologie H24: De celcyclus en mitose | Quizlet
1/54
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
55 Terms
24.1. Overzicht van de celcyclus
Uit welke fasen bestaat de eukaryotische celcyclus
Celcyclus = M-fase + interfase
Interfase = G1 + S + G2
G0 = stoppen tijdelijk met delen (bv. terminale differentiatie of wachten voor groei-stimulus)
M-fase = mitose + cytokinese
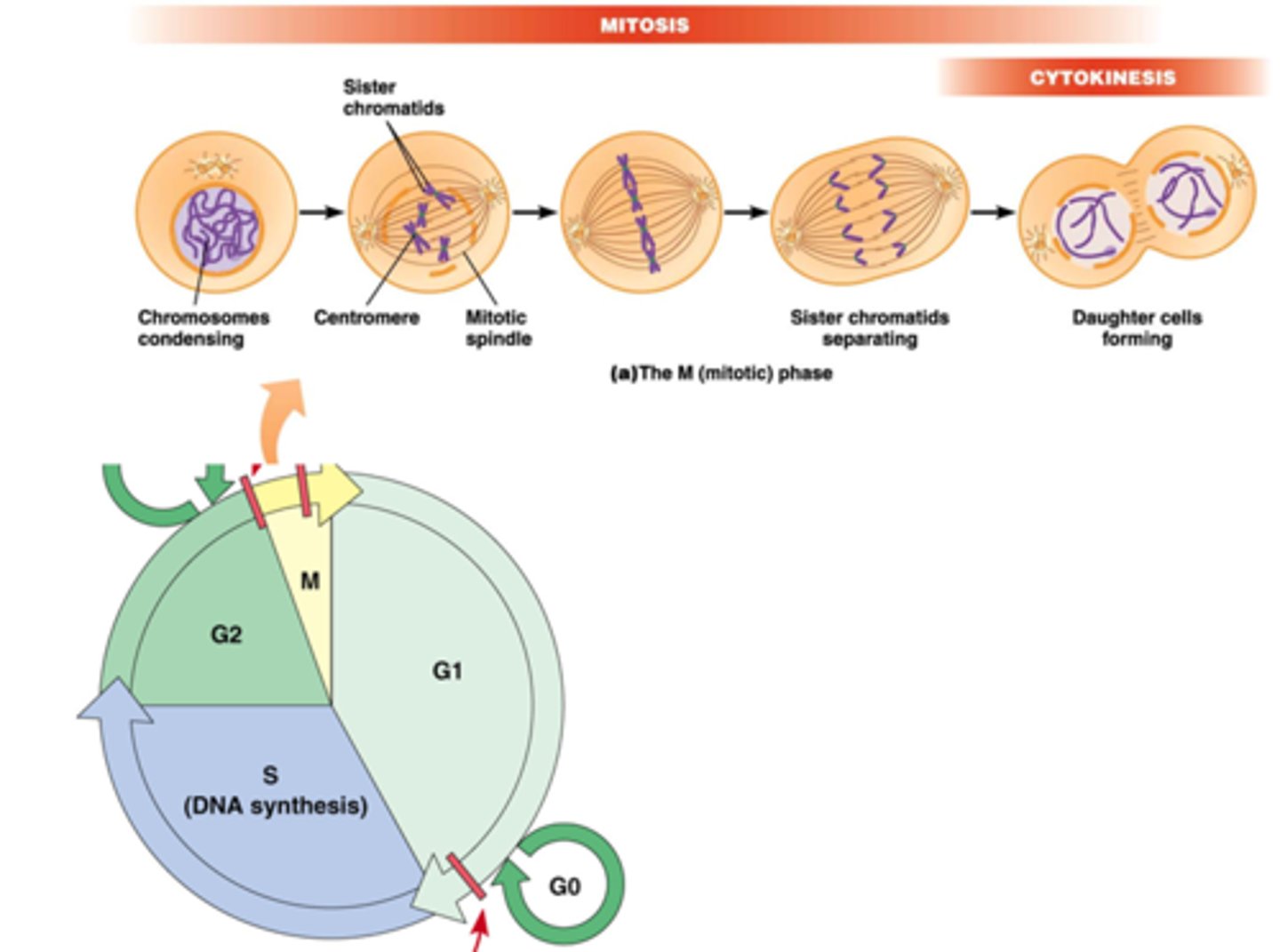
Hoelang duurt elke fase?
•Generatietijd: 18-24h voor sneldelende cellen in cultuur.
•M-fase = mitose + cytokinese (30-45 min in sneldelende cellen). Mitotische index = 3-5% voor cellen in cultuur.
•S-fase: replicatie (6-8h), gebruik van 3H-deoxythymidine voor meten van lengte van S-fase
•G1 (variabele duur)
•G2
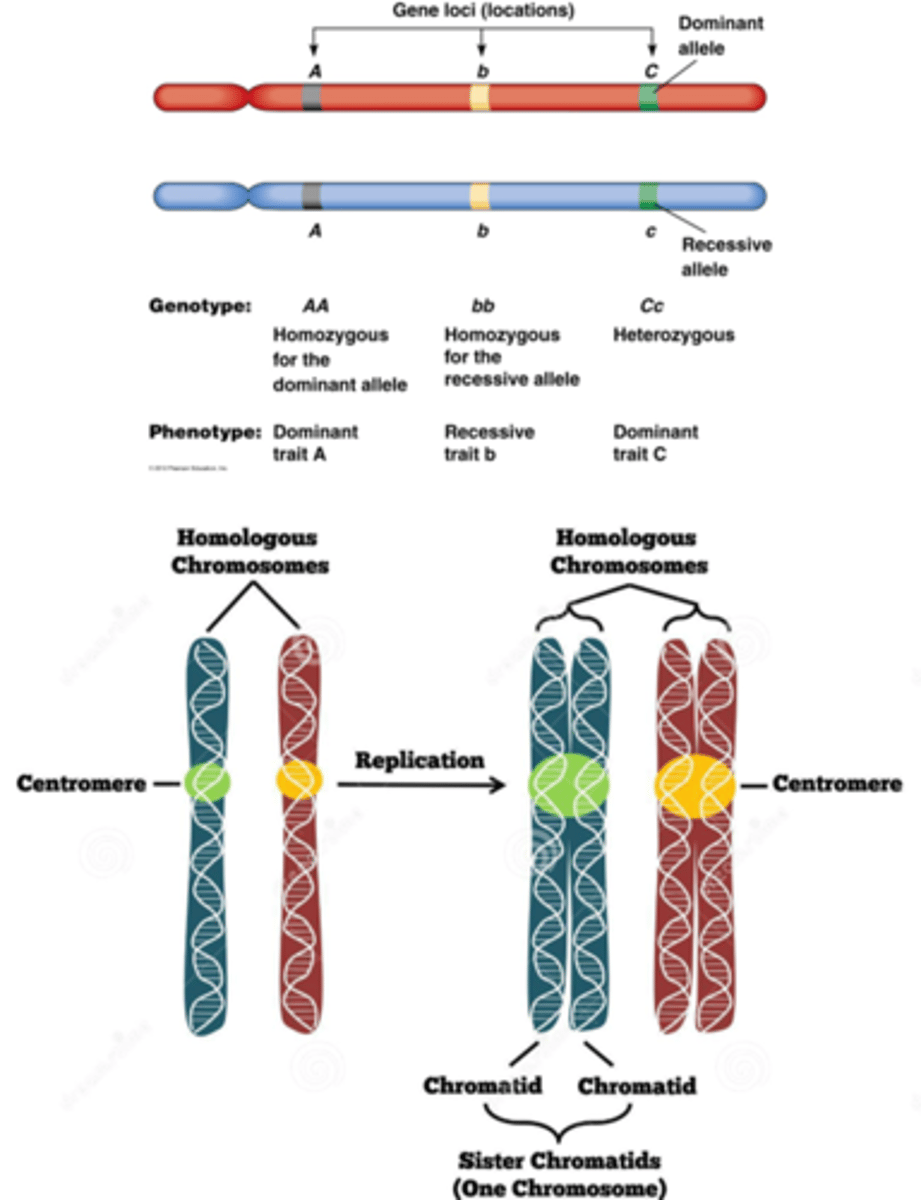
herhaling over chromosomen, chromatiden en allellen
Allel= gen variant
Als het allel anders is: heterozygoot
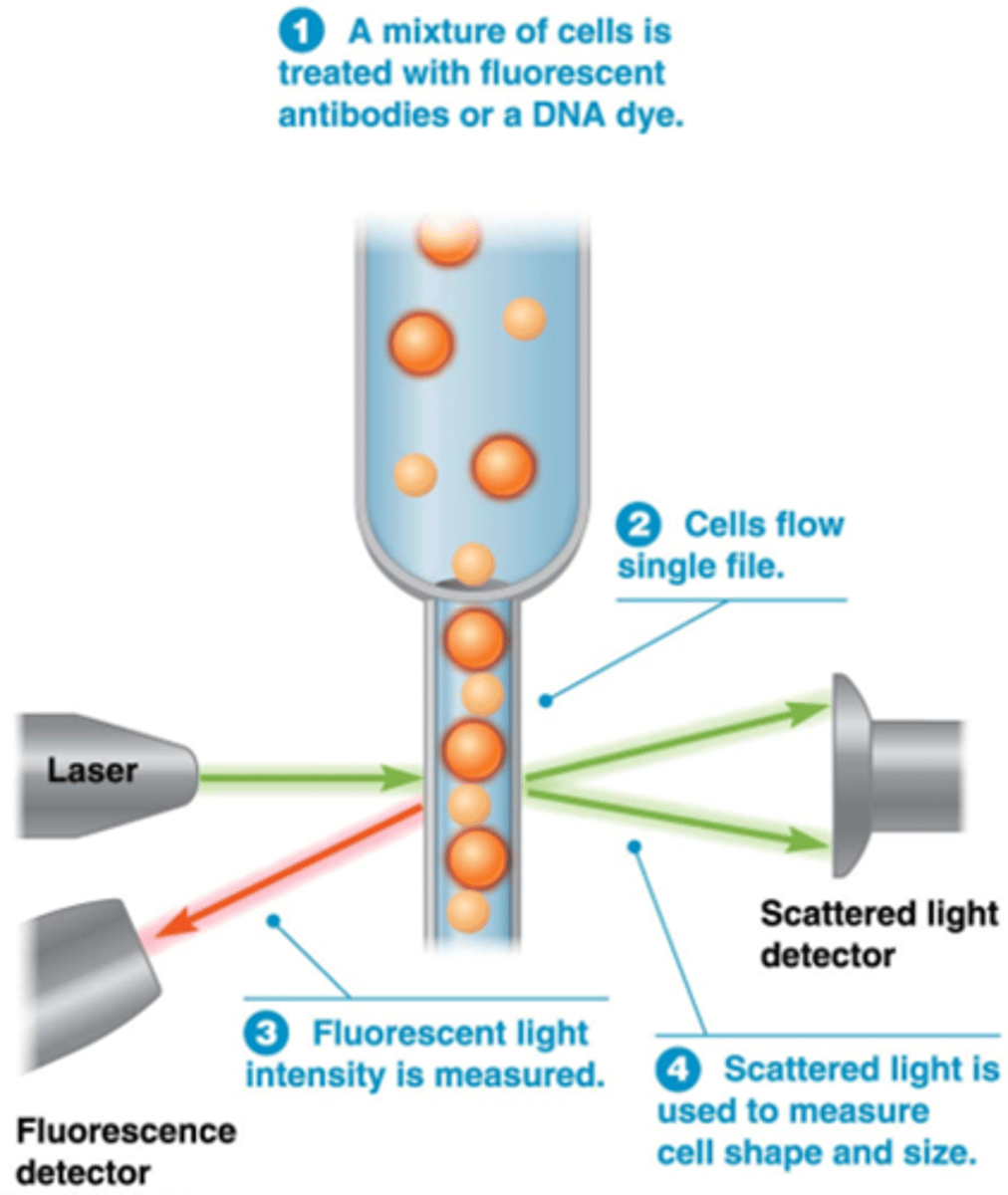
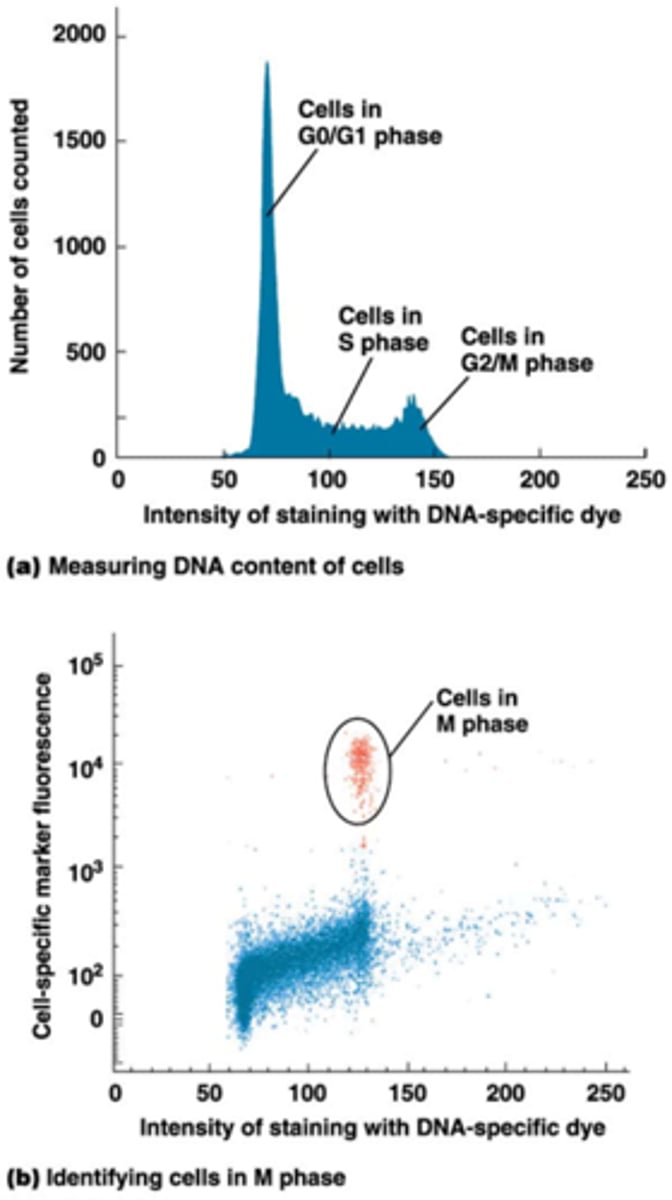
Wat is flow cytometrie?
•mengsel van verschillende celtype's (bv bloedstaal)
•deze gaan door een dun buisje waar slechts 1 cel door kan
•Flow cytometrie laat meting toe van optische (celvorm en -grootte) en fluorescentiekarakteristieken van individuele cellen in suspensie.
•Fluorescente kleurstoffen voor nucleïnezuren (Bv. propidium Jodide voor DNA) en eiwitten (Bv. fluorescent gemerkt antilichaam tegen oppervlaktereceptor).
•Meting van fluorescentie: verschillende kleuren gelijktijdig.
•Lichtverstrooiing = functie van celvorm en celgrootte.
Wat zijn toepassingen van Flow cytometrie?
Toepassingen:
•Meting van aantallen witte bloedcellen (=leukocyten) bv. macrofagen, lymphocyten
•Meting van de fase van de celcyclus: meting van hoeveelheid DNA door DNA fluorescent te kleuren (bovenste foto)
•Flow cytometrie met twee kleuren: hoeveelheid DNA + merker van M-fase (antilichaam dat een mitotische modificatie van histonen herkent) (onderste foto)
->techniek evolueert heel sterk, nu kunnen we 6 verschillende markers gebruiken en tegelijkertijd onderzoeken
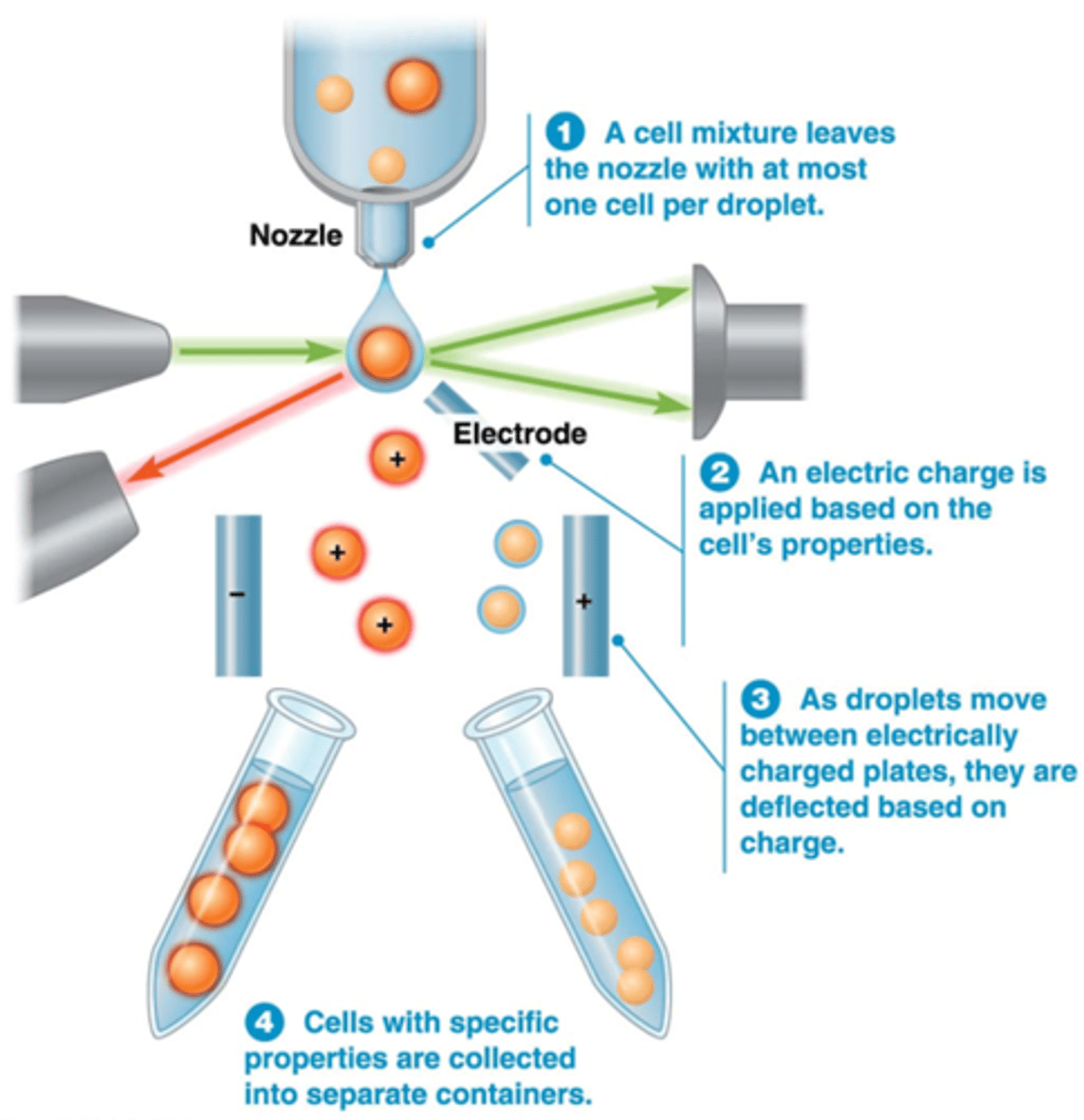
Wat is Fluorescence-activated cell sorting (FACS)
•Flow cytometrie + selectie/sortering van cellen op basis van elektrische lading
•Geselecteerd celtype kan vervolgens apart worden geanalyseerd (Bijv stamcellen), je kan ze ook apart laten groeien in een petrischaaltje
24.2. Kern en celdeling
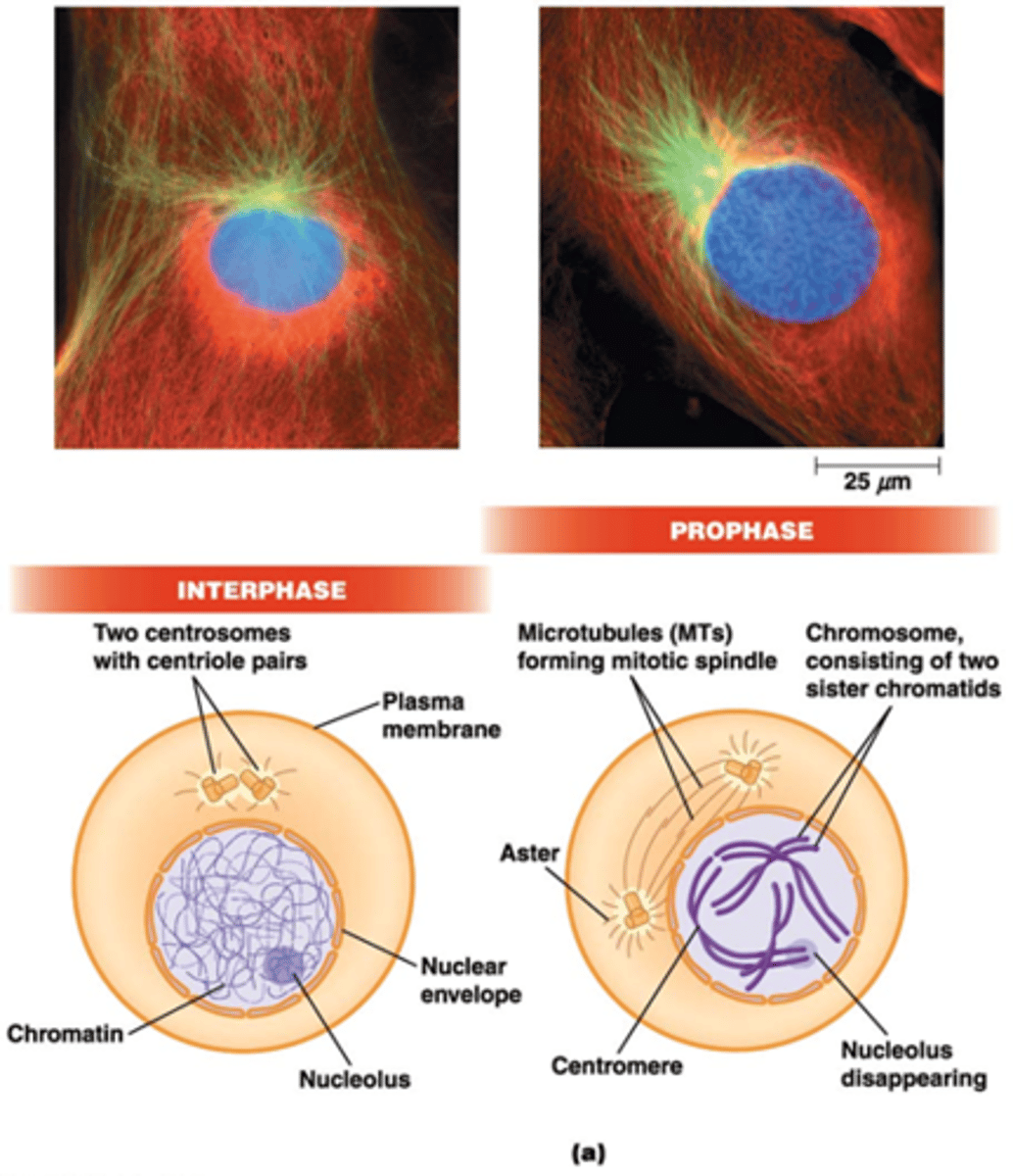
Wat gebeurt er in de profase?
•De chromosomen worden zichtbaar (zusterchromatiden)
•Verdwijnen van nucleoli
•Twee centrosomen* – microtubule-organizing centers (MTOC): nucleatie + verankering van microtubuli → mitotische kernspoel + aster
*Elk centrosoom bestaat uit 2 orthogonale centriolen (opgebouwd uit 9 tripletten van MT) (zie celbio)
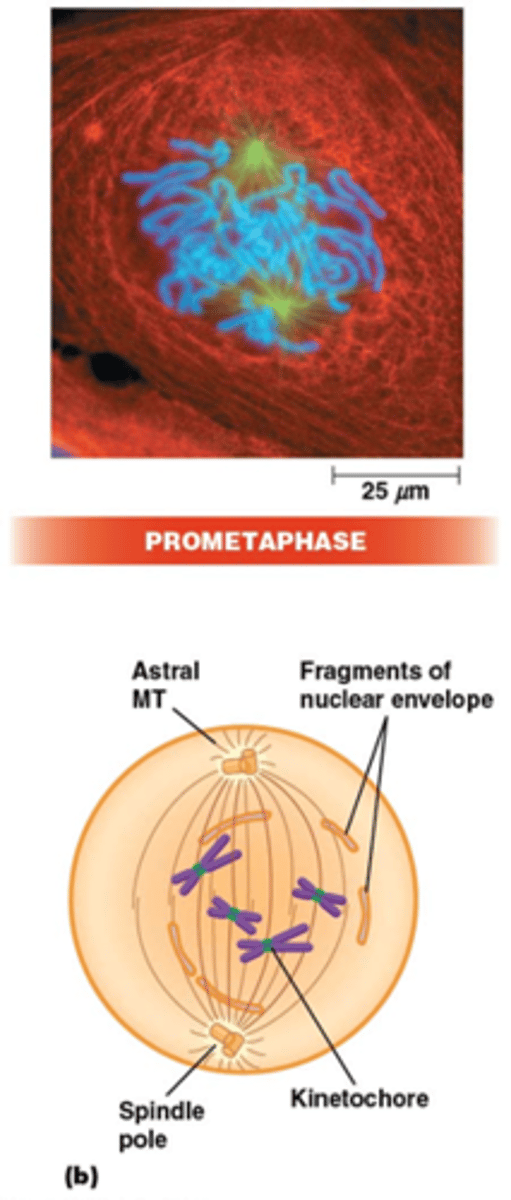
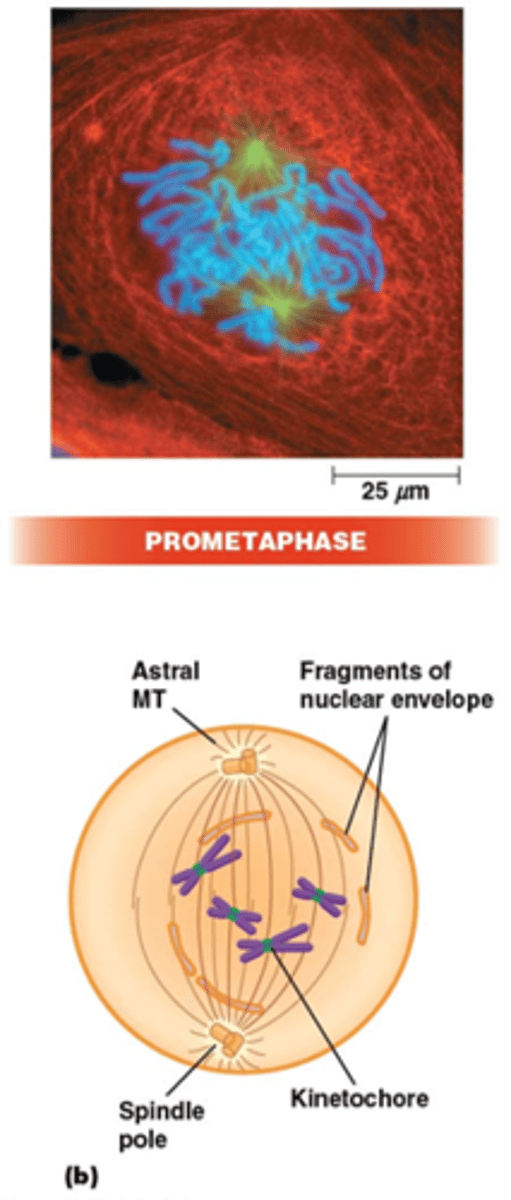
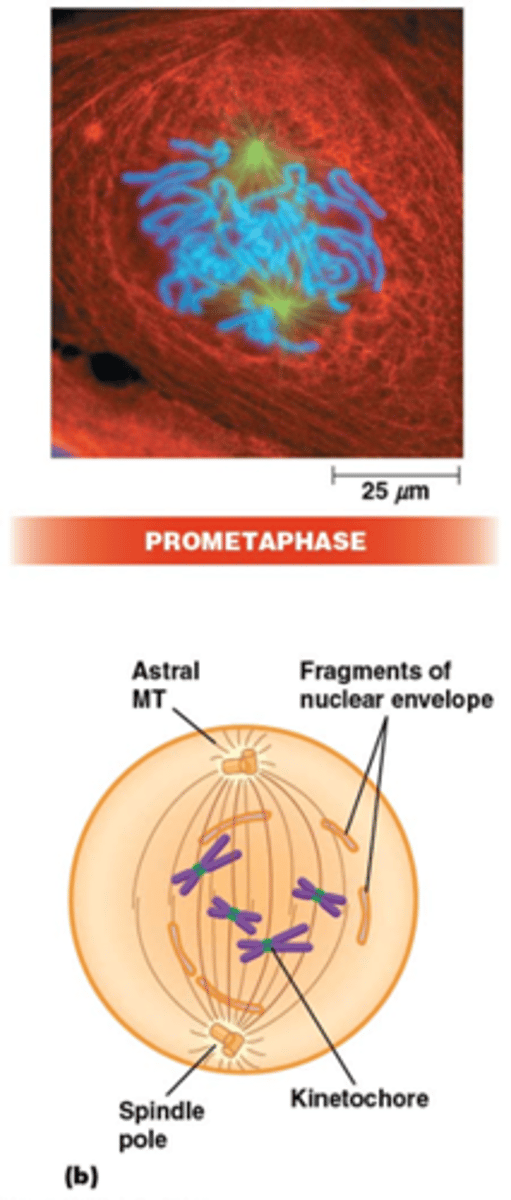
Wat gebeurt er in de prometafase?
•Fragmentatie van de kernmembraan (fosforylering van laminen)
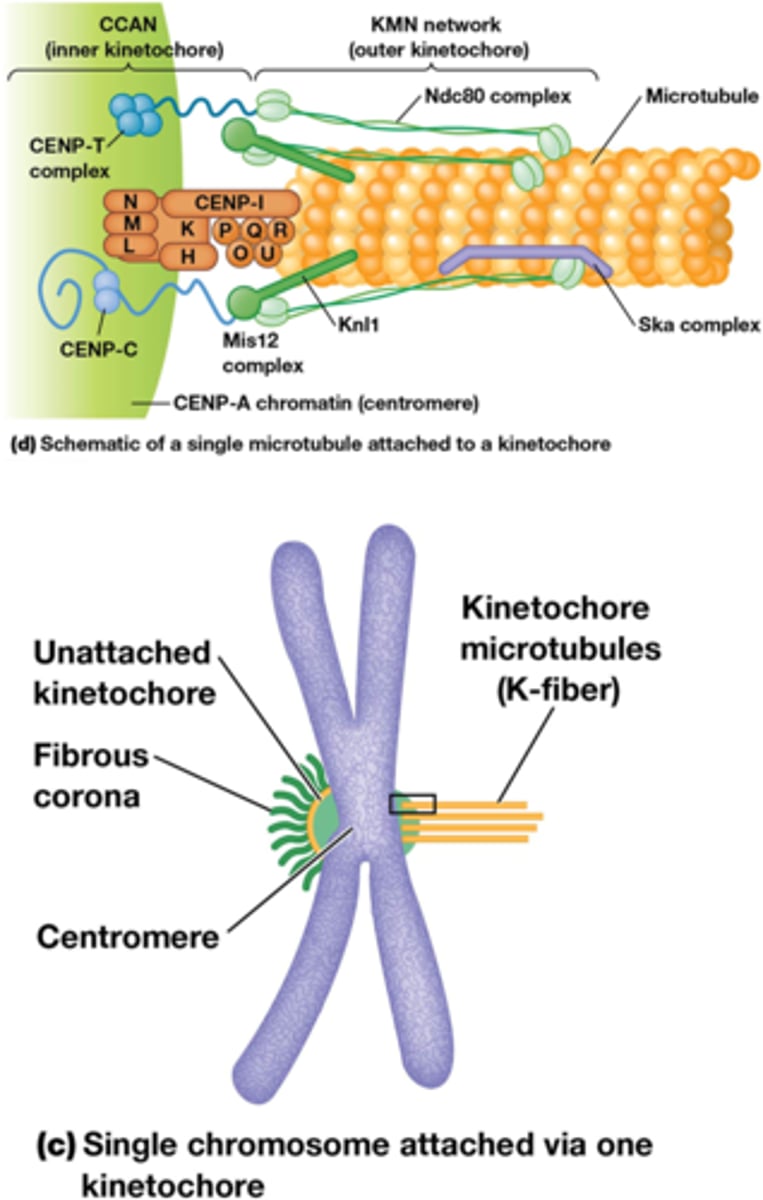
•MT-kinetochoor-interactie. Centromeren: repetitief DNA (CEN sequentie), histon H3 variant (CENP-A) recruteert kinetochoor complex waaraan MTs binden via + uiteinde
•Vorming binnenste (na S-fase) en buitenste kinetochoor (tijdens mitose)
•Fibreuze corona: enkel zichtbaar bij niet-gebonden kinetochoren
•Elk zusterchromatide heeft een eigen kintochoor, de 2 kinetochoren zijn verschillend georiënteerd, elk naar een andere pool. De orientatie is een willekeurig proces
Alle namen van de eiwitten zeker niet kennen want er zijn er nog veeel meer
Wat kan je nog zeggen over kinetochoor microtubili?
Kinetochoor microtubule:
-MT binden aan kinetochoor
- Alle MT die gebonden zijn op een kinetochoor zijn afkomstig vandezelfde pool.
- De MT die gebonden zijn op de kinetochoor van de andere zusterchromatide zijn afkomstig van de tegengestelde pool
- Welke kinetochoor verbonden is met welke pool is willekeurig!
Hoe kunnen chromosomen verplaatsen naar het evenaarsvlak?
- ‘Congression’(uitlijning) = het proces waarbij de chromosomen zich bewegen/verplaatsen naar midden van de cel (evenaarsvlak)
MT zijn dynamisch instabiel, ze polymeriseren (groeien) en depolymeriseren (krimpen) -> trekken en duwen de chromosomen heen en weer. Bij ‘congression’ zijn ook verschillende en kinesins en dyneins bij betrokken
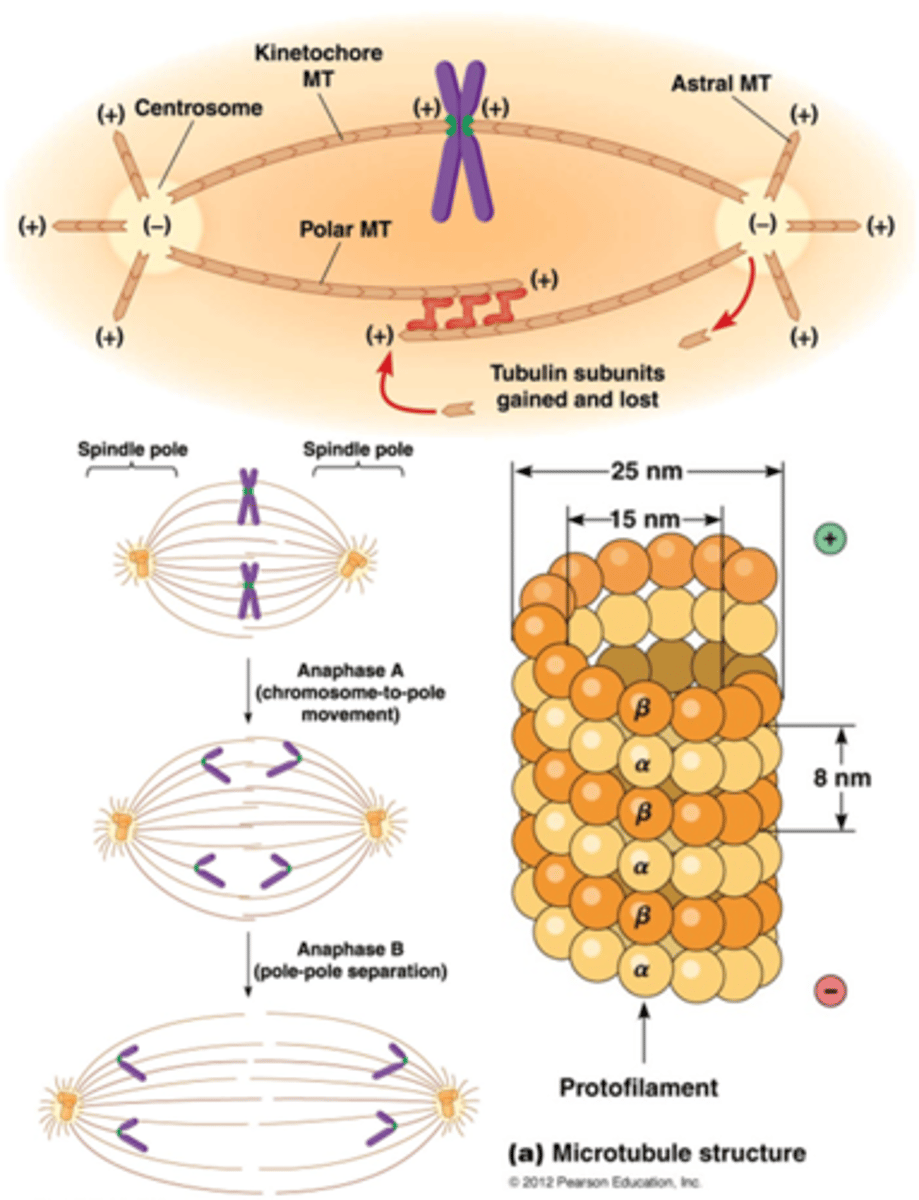
Welke 2 andere soorten microtubili hebben we gezien en wat zijn hun eigenschappen?
•Polaire microtubuli (van pool tot pool) (welke pool is willekeurig want de orientatie is willekeurig)
•Astrale microtubuli (contact met eiwitten van de celcortex (actine microfilamenten geassocieerd met het plasmamembraan))
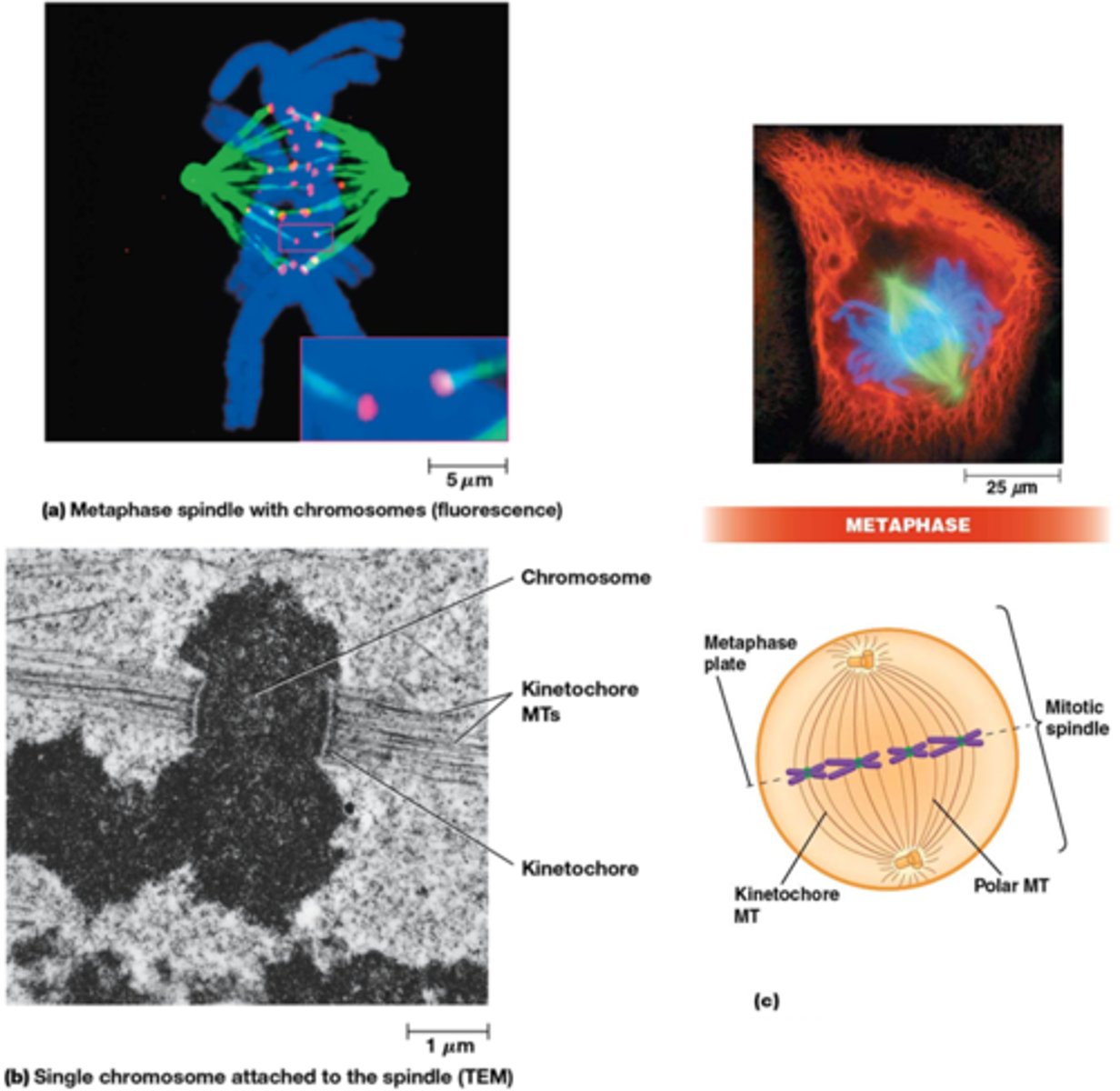
Wat gebeurt er in de metafase?
•Chromosomen gealigneerd in equatorvlak ter vorming van metafaseplaat
•De twee zusterchromatiden zijn elk verbonden via kinetochoor MT verbonden met een tegengestelde pool
•Microtubuli heel dynamisch, ook in metafase
Wat gebeurt er in de anafase?
•Breking van binding tussen zusterchromatiden: afbraak van cohesine
•Anafase A: kinetochoor MTs worden korter en trekken chromatiden naar tegenovergestelde pool.
•Anafase B: polaire MTs worden langer en verwijderen de polen verder van elkaar
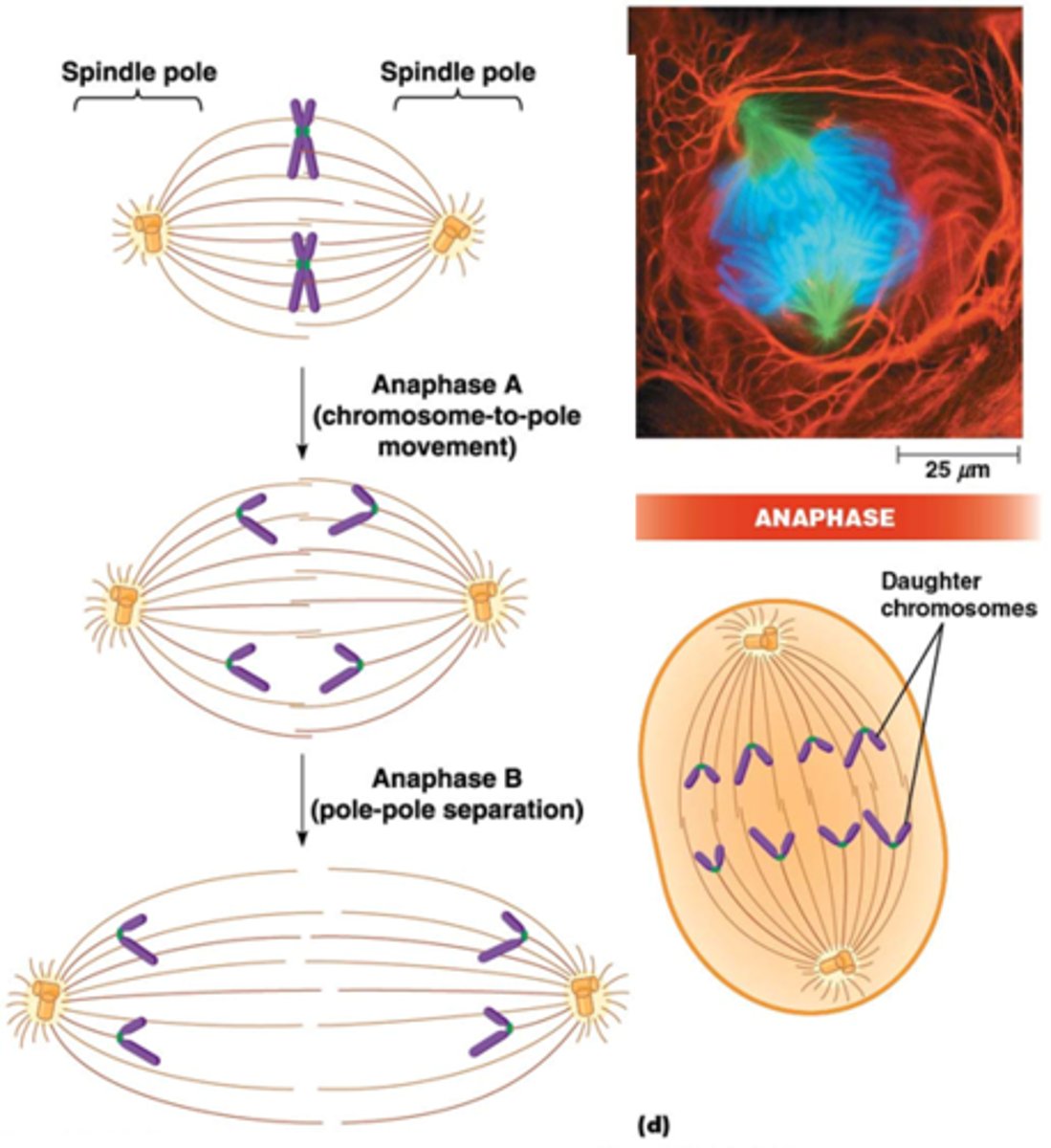
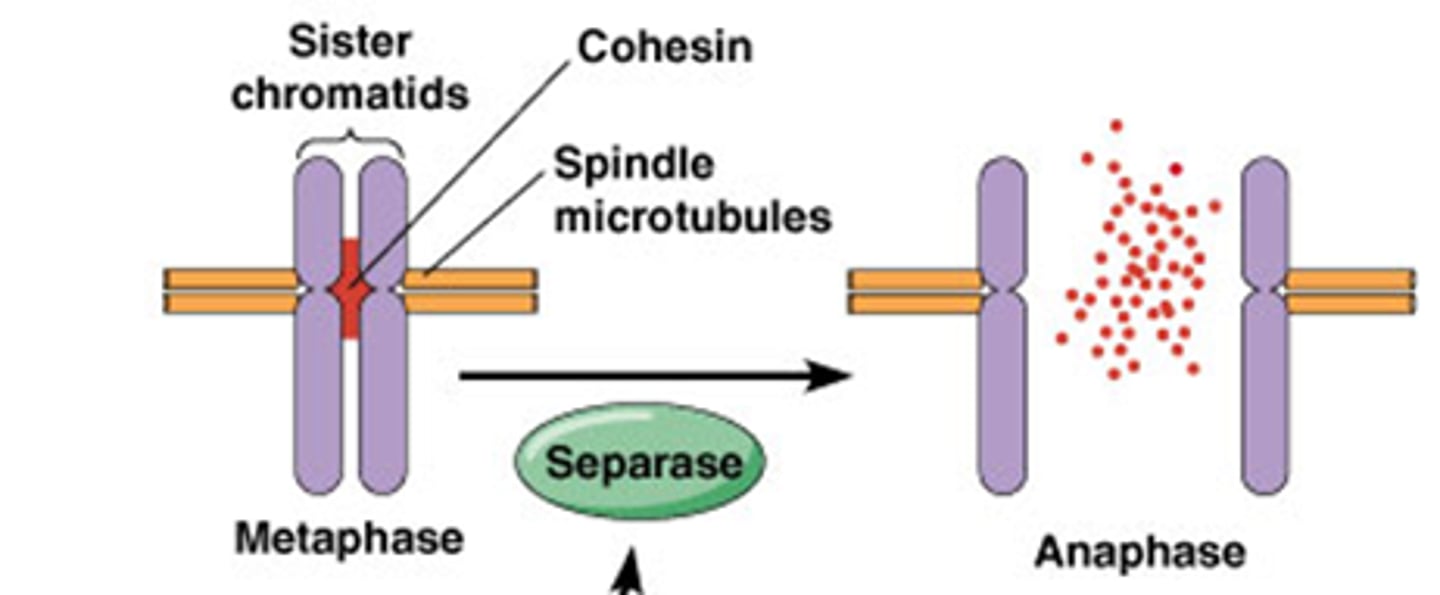
Hoe wordt het centromeer afgebroken?
hier worden de identieke dochtercellen gemaakt, hierdoor moet het centromeer (cohesine is hiervoor belangrijk) afgebroken worden separase
Wat is de rol van de kernspoel?
De kernspoel scheidt chromosomen tijdens celdeling. Microtubuli, met een polair - uiteinde bij het centrosoom en een + uiteinde bij het kinetochoor, trekken zusterchromatiden naar tegenovergestelde polen van de cel.
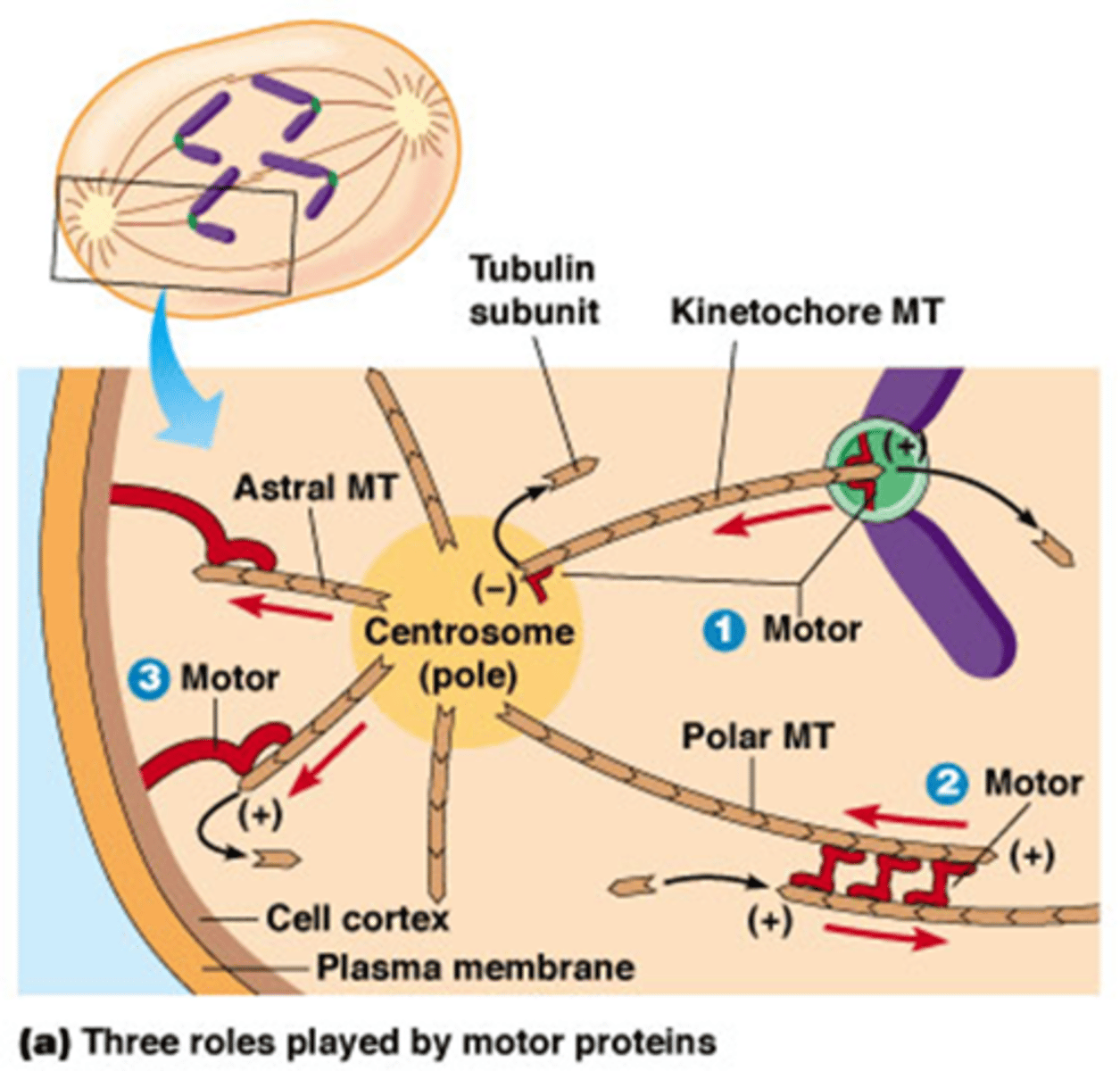
Welke 3 motoreiwitten spelen een rol in de anafase? volgende kaartjes gaan in detail
1. Kinesine gebonden aan kinetochoor-MTs plus-uiteinde:
2. Bipolaire kinesine motor-eiwitten
3. Cytoplasmatische dyneinen
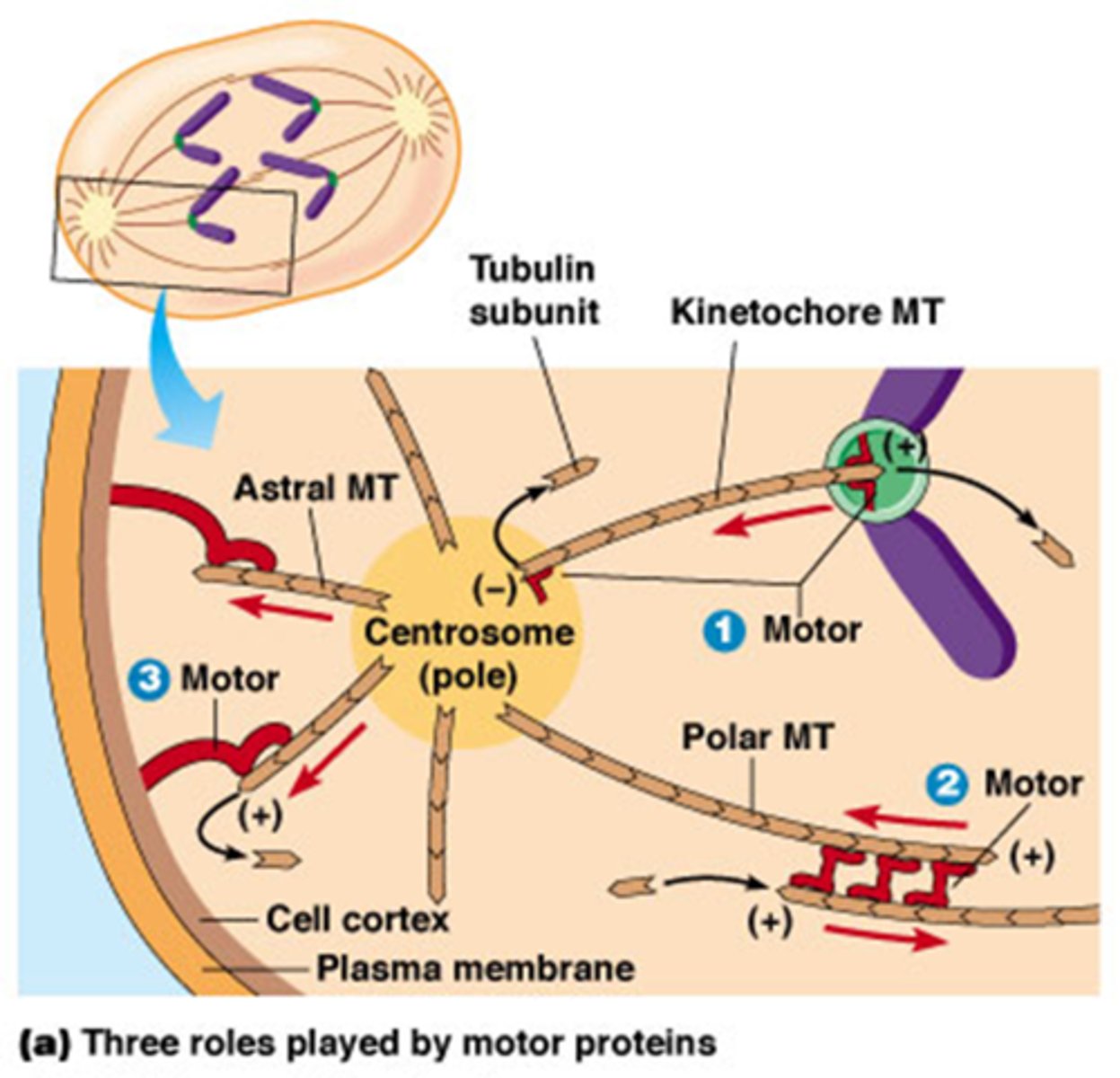
Wat is de rol van kinesines aan de kinetochoor-MT + uiteinde?
beweging van de chromosomen naar polen van de kernspoel tijdens anafase A, gekoppeld aan MT depolymerisatie.
Kinesines die zitten aan de + uiteindes zorgen voor depolymerisatie van MT -> ze worden korter langs beide kanten
Kinesine aan de – uiteinde zal ook depolarymerisatie induceren
->het binnenhalen van de chromsomen
(rood hoekje van 90°)
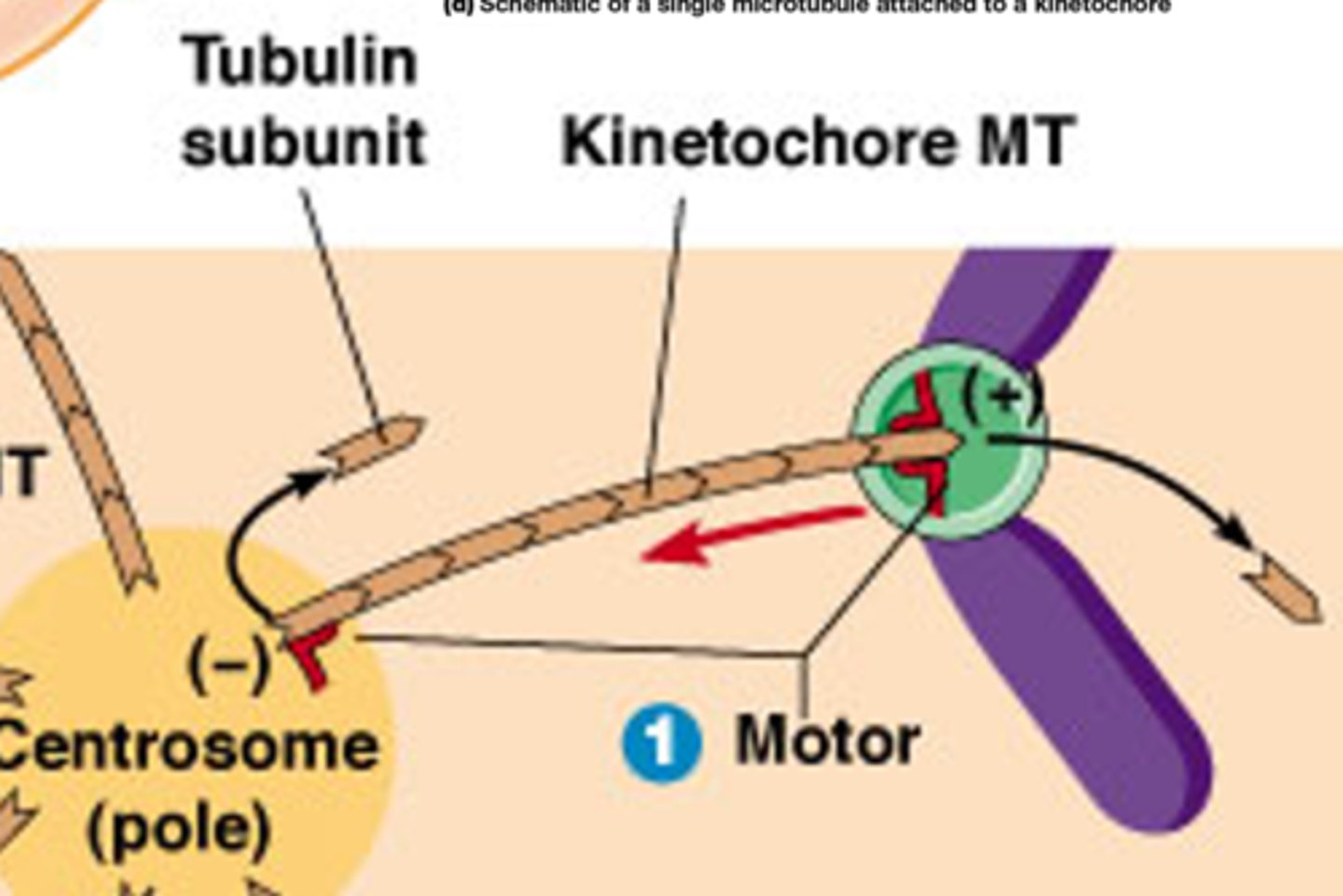
Wat is de rol van Bipolaire kinesine motor-eiwitten
Bipolaire kinesine motor-eiwitten (tetrameer) binden aan overlappende polaire MTs en duwen de polen verder uiteen tijdens anafase B
4 verschillende kopjes, ze bewegen beide naar de plus uiteinde maar deze zijn naar de andere richting, hierdoor gaan de microtubili uit elkaar schuiven
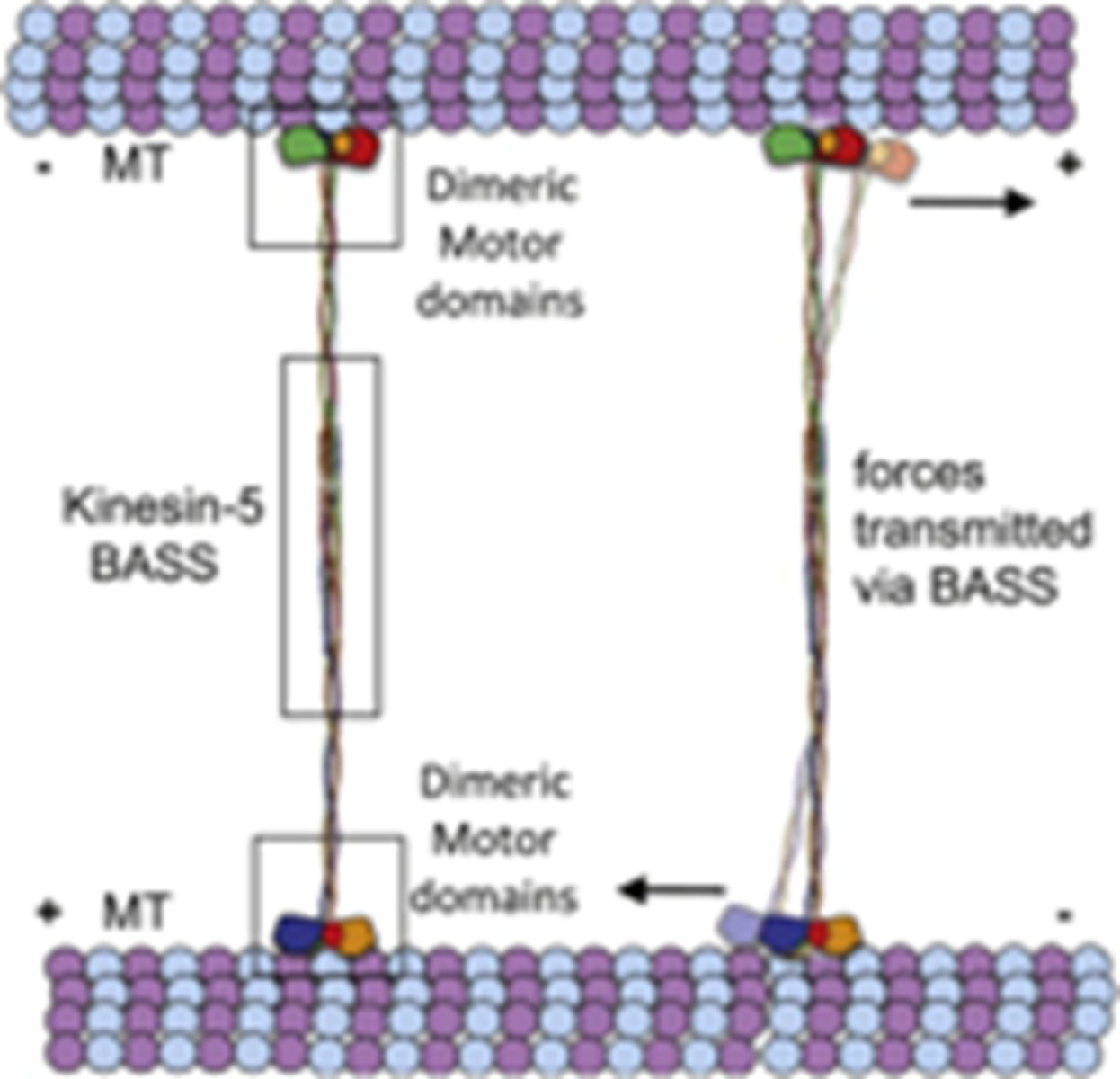
Wat is de rol van Cytoplasmatische dyneinen
ze zijn gebonden aan celcortex en binden aan astrale MTs, bewegen naar min uiteinde van MT en trekken de polen van de kernspoel richting cortex.
3 op de foto
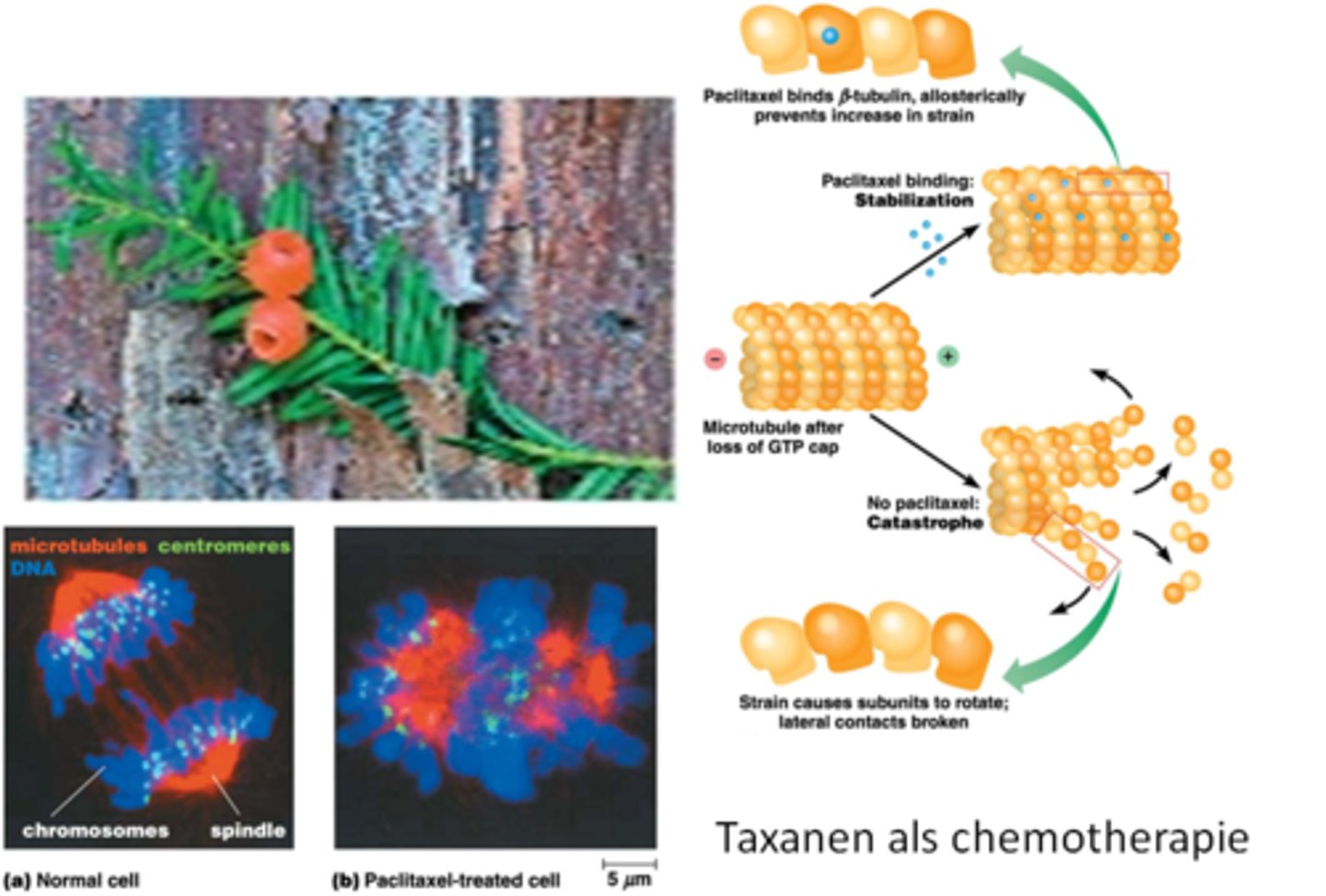
Wat is de link tussen ethnobotany en kanker?
Ethnobotany: planten in de geneeskunde
-stof extraheren (een taxane) van de taxushaag
-taxanes kunnen de mitose stoppen
-hier is de stof paclitaxel, deze stof bindt op de beta tubiline
-binding leidt tot stabilisering van beta tubline, je gaat niet meer GTP hydrolase kunnen doen
-als paclitaxel bindt krijg je geen GTP cap en ga je depolymerisatie hebben van het MT uiteinde*
->chemotherapie maar veel neveneffecten omdat de goede cellen ook aangetast worden
herhaling celbio: groei/catastrofe gebeurt op + uiteinde, groei gebeurt als beta GTP een groot genoeg concentratie heeft (alpha heeft altijd GTP gebonden): GTP cap
Wat gebeurt er tijdens de telofase?
•Chromosoom decondensatie, nucleoli verschijnen + kernmembraan wordt gevormd
•Tijdens telofase begint ook al cytokinese
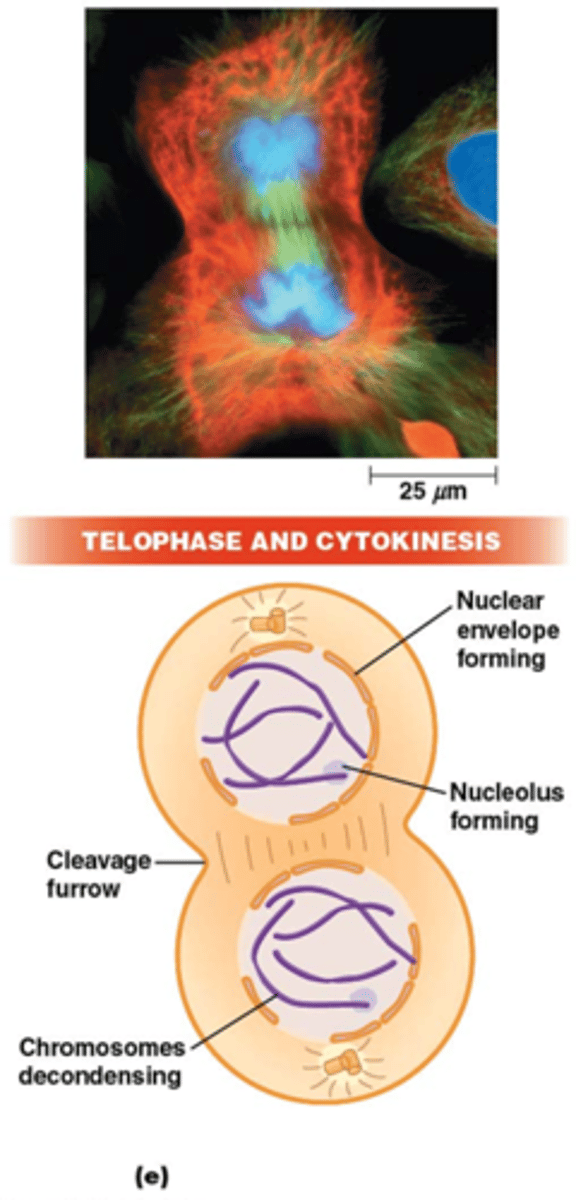
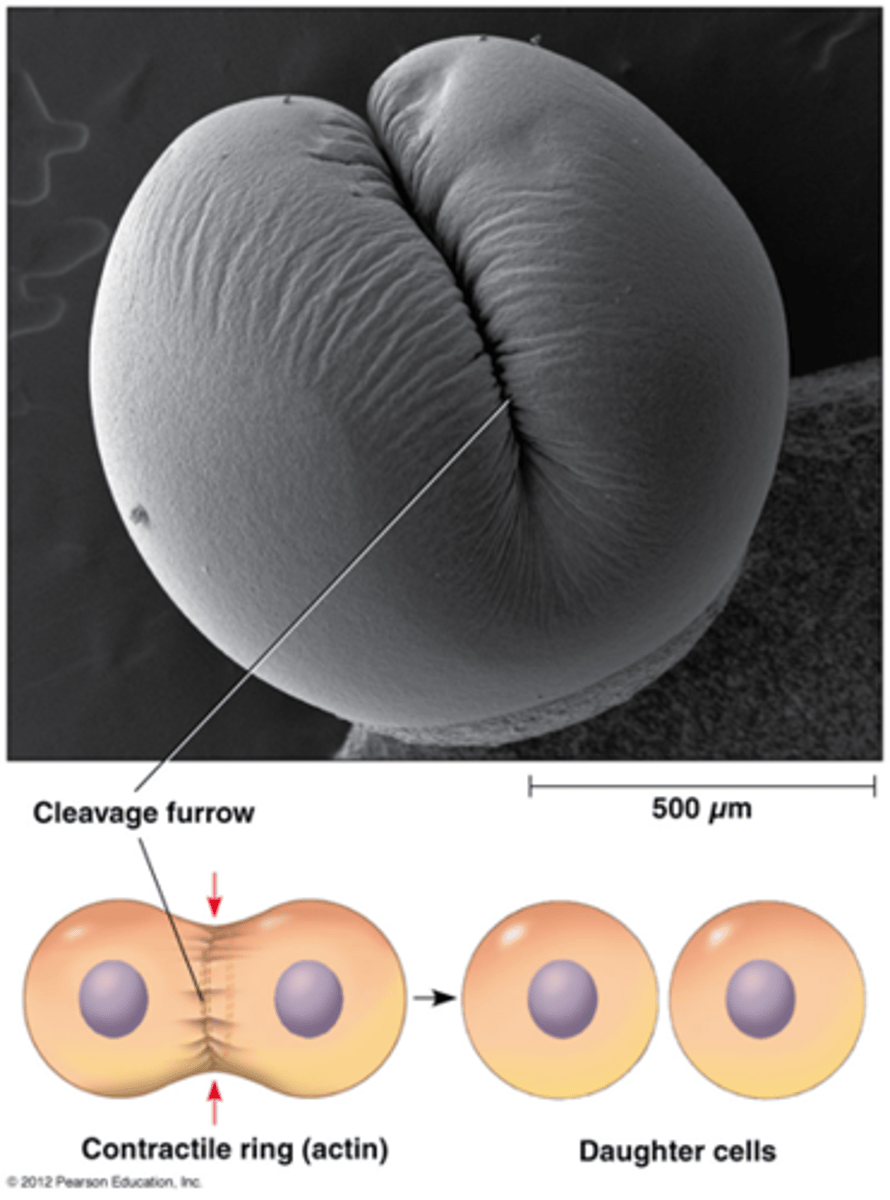
Wat gebeurt er tijdens de cytokinese (in dierlijke cellen)
•Insnoering van plasmamembraan in vlak dat door de equator gaat.
•contractiele actine-microfilamenten
•Mitose zonder cytokinese leidt tot multinucleaire cellen
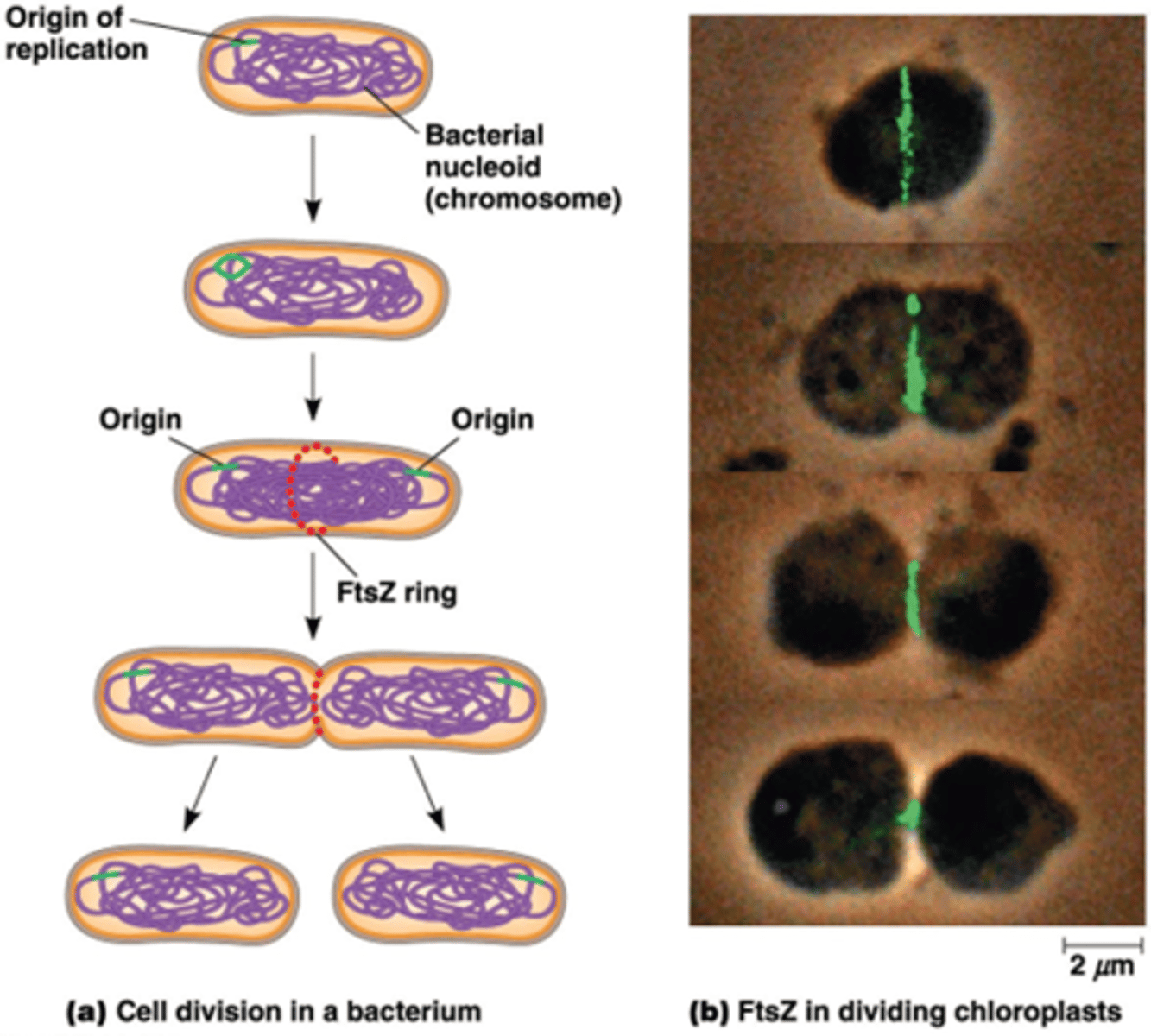
Hoe verloopt de cytokinese van bacterien en eukaryotische organellen?
Cytoskeletaal eiwit (FtsZ) dat een ringstructuur vormt
Mitochondriën en chloroplasten delen volgens gelijkaardige mechanismen
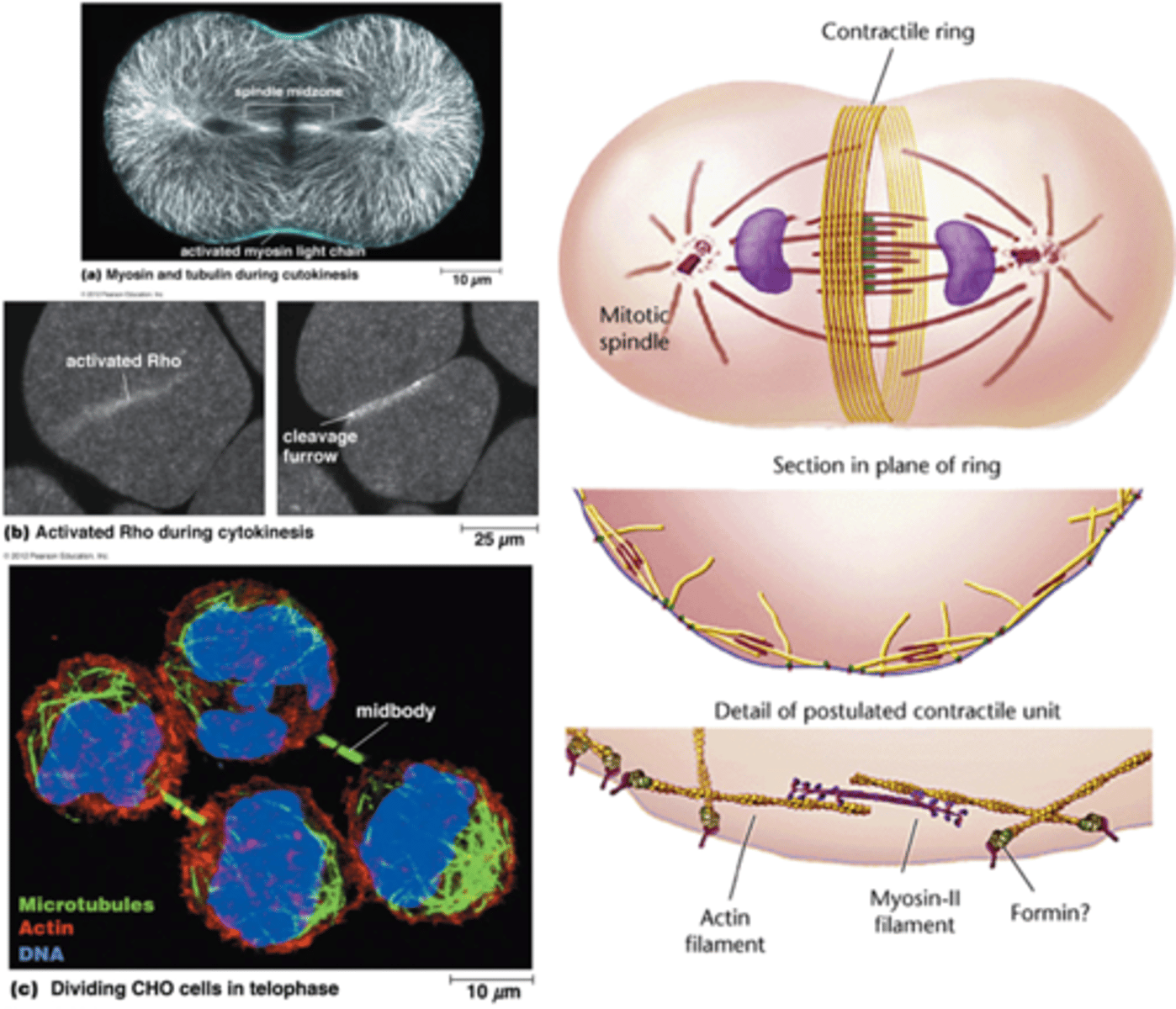
Wat is de contractiele ring en hoe werkt het?
De contractiele ring is een beetje gelijkaardig met spiercontractie!
•Interactie actine – myosine (motor-eiwit)
•Rho A-GTP stimuleert actine polymerisatie en activeert myosine (Rho A-GTP stimuleert rho-kinase, rho kinase activeert actine en myosine)
-Na insnoering vorming van midbody= bundel van microfllamenten omgeven door PM->doorknippen van midbody= abcission
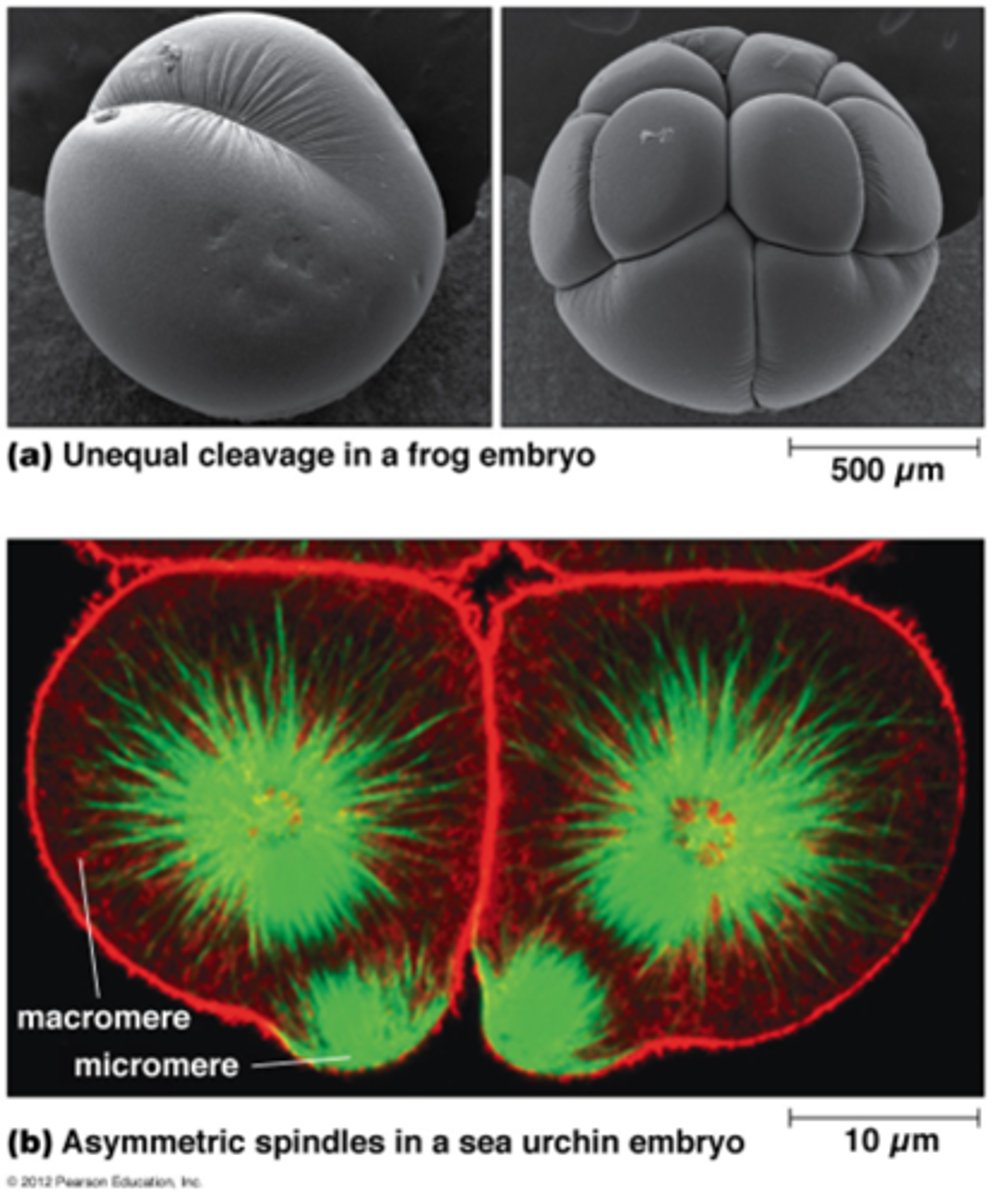
Waarom is de celdeling soms assymetrisch?
•Assymmetrische deling: twee dochtercellen verschillen in grootte en/of in 'fate'
•Van belang bij embryonale ontwikkeling en bij stamcellen: Bij deling één dochtercel = stamcel, andere dochtercel = meer gedifferentieerd
•Kernspoel asymmetrisch (macromeer en micromeer)
•Asymmetrische deling kan ook t.g.v. asymmetrische verdeling van cytoplasmatische componenten.
24.3. Regulatie van de celcyclus
Wat is het verschil tussen celtypes op vlak van celdeling?
•Sneldelende (embryonale cellen, hepatocyten) versus traagdelende cellen (bv zenuwcellen).
Soms enkel deling na stimulatie (lymfocyten). G0 fase = verlengde G1.
In G1 fase veel genexpressie (eiwitsynthese) omdat de cel na de mitose qua volume kleiner is dus die moet wel groeien (groeien= eiwitten aanmaken)
Cellen zijn in of buiten de celcyclus: induceerbaar door groeifactoren
Wat doet eiwitkinase TOR?
•Coördinatie celgroei en celdeling: eiwitkinase TOR: regelt eiwitsynthese maar ook overgang naar S-fase.
belangrijk in de G1 fase want daar is veel eiwitsynthese
Wat gebeurt er alles er iets fout is: bv DNA schade?
Celcyclus kan tijdelijk worden stopgezet (DNA schade, replicatie niet beëindigd, kinetochoor-MT interactie niet matuur).
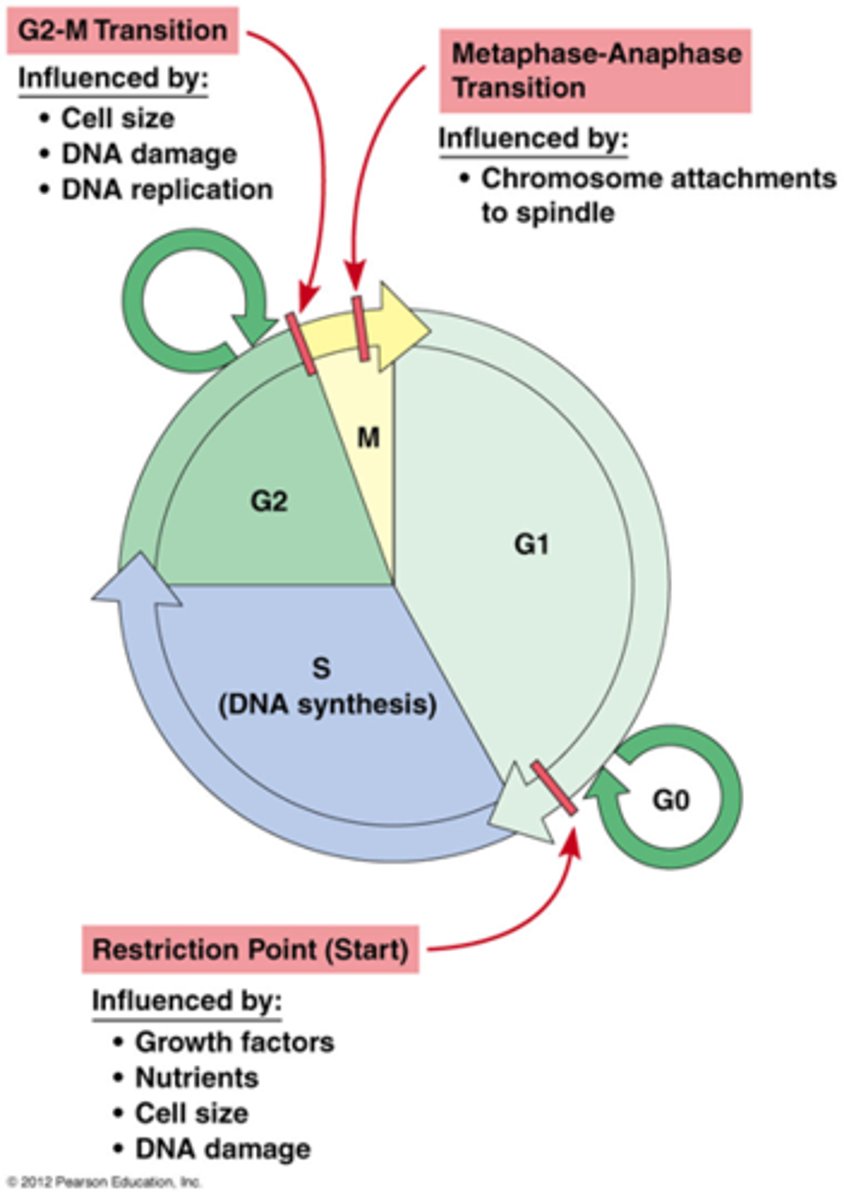
Wat zijn de transitiepunten?
Transitiepunten: er moeten bepaalde voorwaarden voldaan worden voor die in die transitie kan gaan
Overgang tussen de fasen zijn scherp
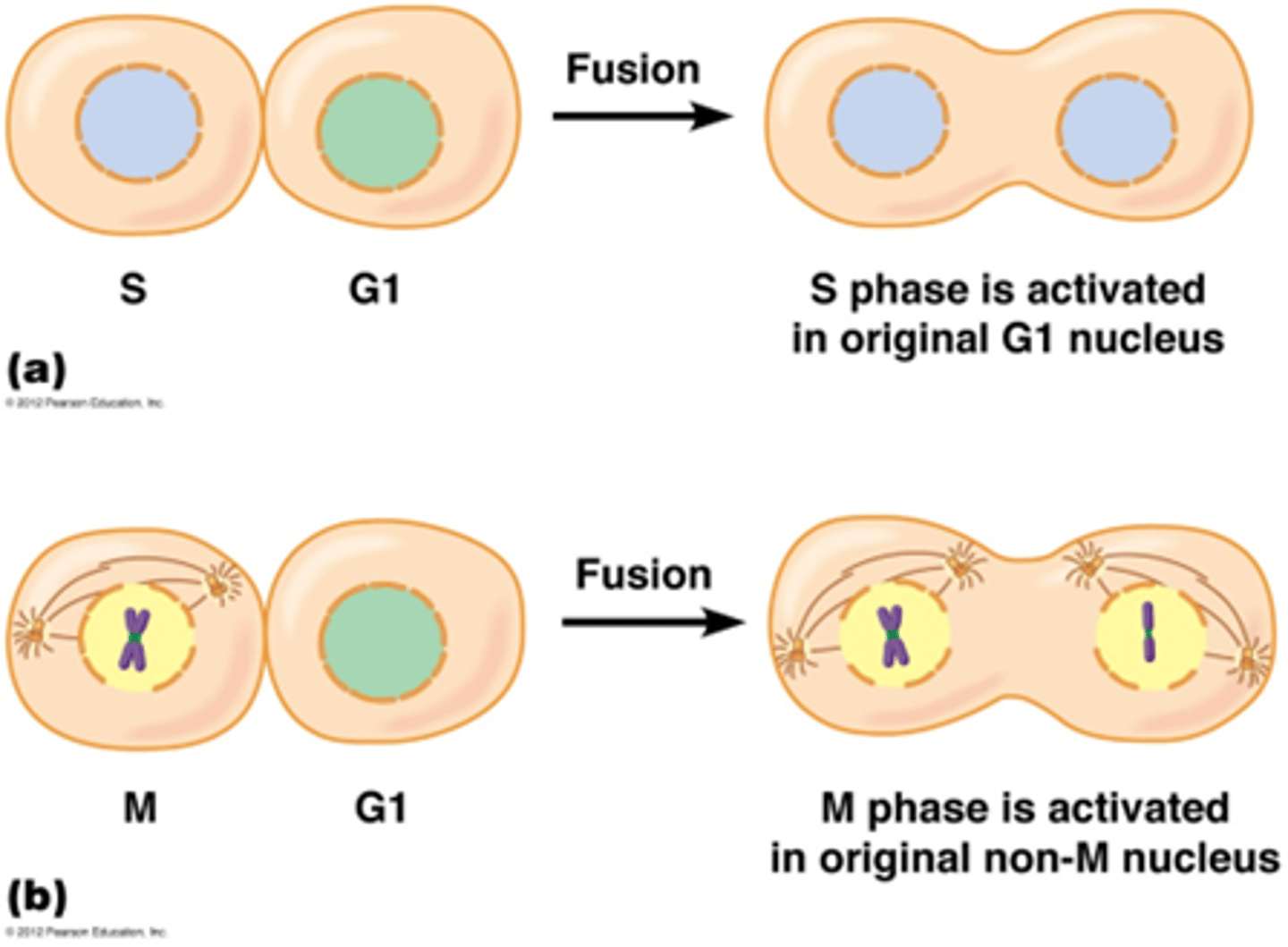
Wat waren de celfusie experimenten? (microscopie)
- heterokaryon S-fase + G1 fase → S-phase inductie
-heterokaryon M-fase + G1→ M-fase inductie
Door een S en G1 te combineren zie jedat de cel insant in de S fase komt, er moet dus iets in die linker cel zitten dat de S fase induceert
Door een M en G1 fase te combineren zie je dat de mitose instant start -> er is een overdraagbaar element dat de M fase induceert
genetica:
Hoe hebben ze de cell-division-cycle (Cdc) genen in gist ontdekt?
temperatuur-afhankelijke mutanten: ondermeer cycline-afhankelijke kinasen (Bv. Cdc2 induceert mitose, Cdc2 = Cdk1)
zo hebben ze de Cdc genen ontdekt, deze zijn belangrijk voor de celcyclus
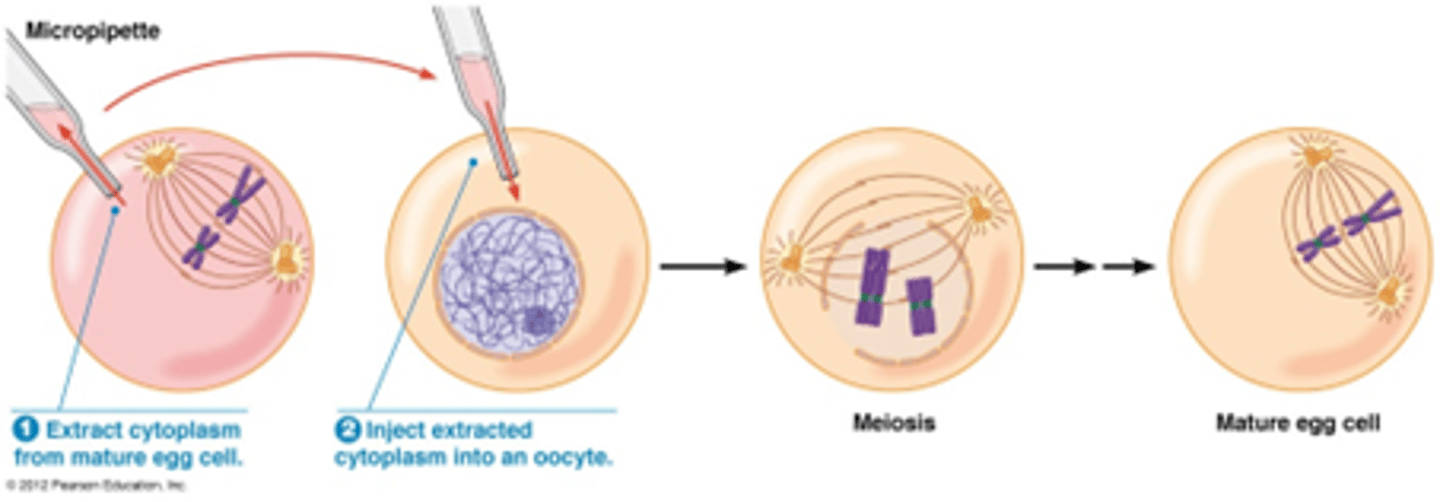
biochemie: injectie cytoplasma van rijpe oocyt in oocyt geblokkeerd in meiose
injectie cytoplasma van rijpe oocyt in oocyt geblokkeerd in meiose induceert vervollediging van meiose (Mitosis Promoting Factor = MPF = Cdc2 + mitotische cycline)
(zie hoofdstuk 25)
Wat zijn Cdk's? belangrijk
Cycline-afhankelijke eiwitkinasen (Cdk)
Bestaat uit een dimeer dat gaat zorgen voor kinase activiteit:
-kinase eiwit
-cycline eiwit
je hebt verschillende Cdk's en elke Cdk complex heeft zijn eigen cycline:
Cyclinen voor G1, S, G2, G2-M: activatoren voor specifieke Cdks. Bv. Cycline B in MPF
•Cdks = meester-regulatoren van de celcyclus
grafiek van de mitotische cycline:
-zwarte lijn is activiteit (er is pas activiteit na een bepaalde concentratie) (heel stijl: zie volgend kaartje over de pos feedback)
-blauwe lijn is concentratie van cycline (Cdk is pas actief als cycline gebonden is)
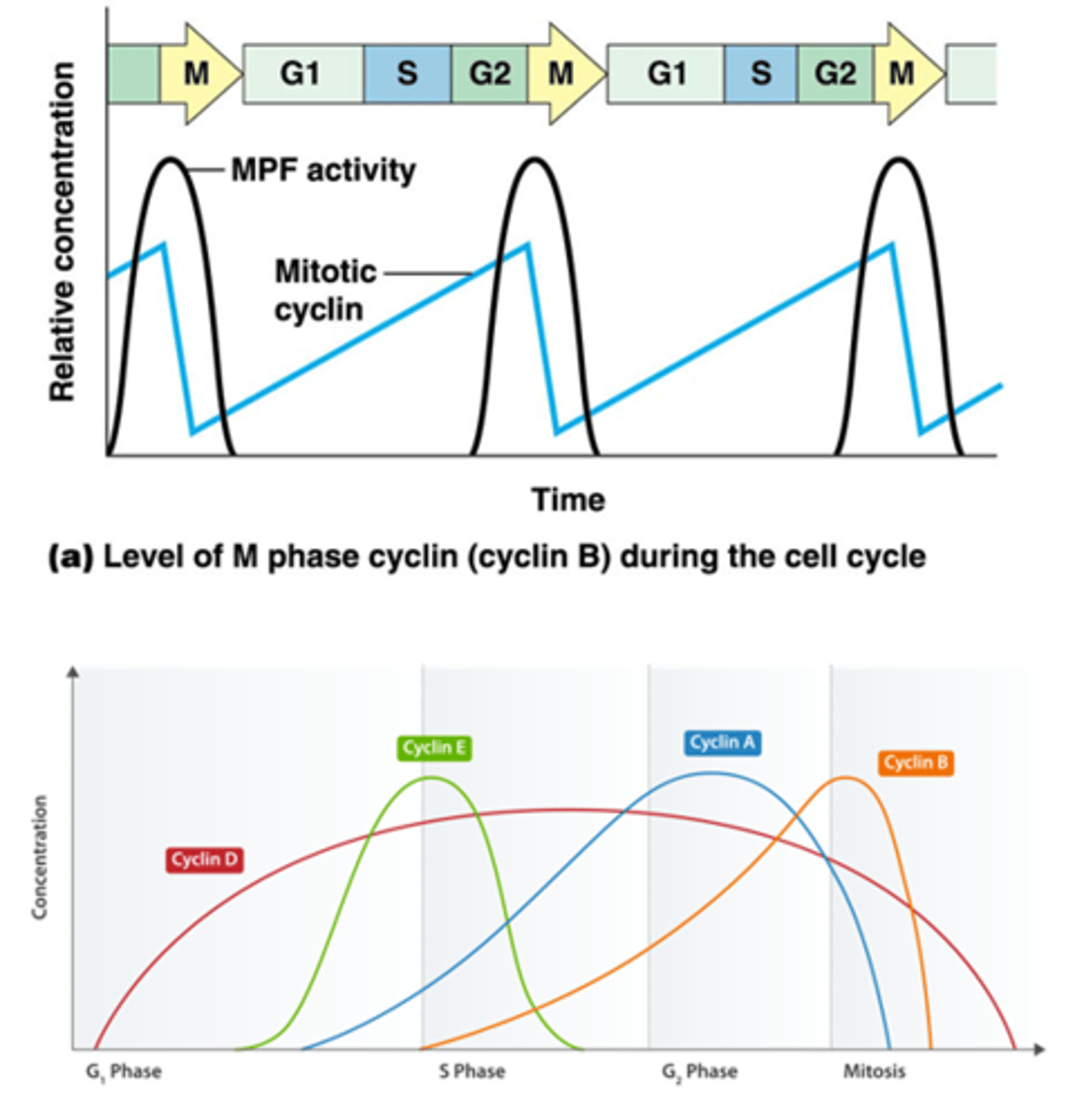
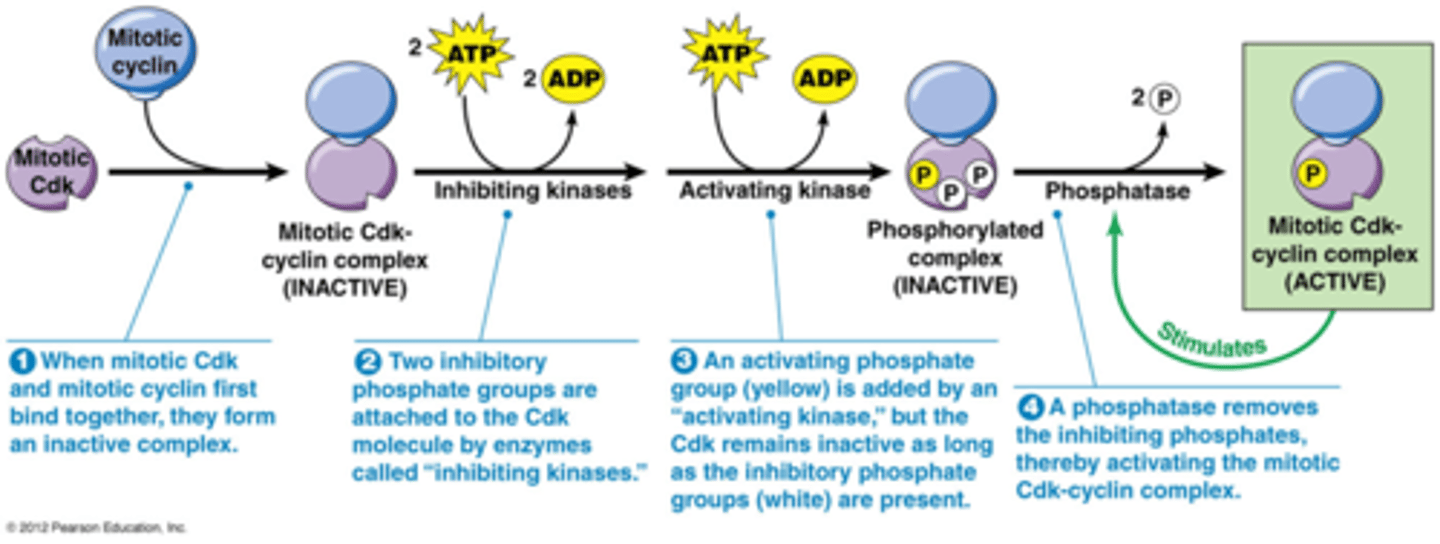
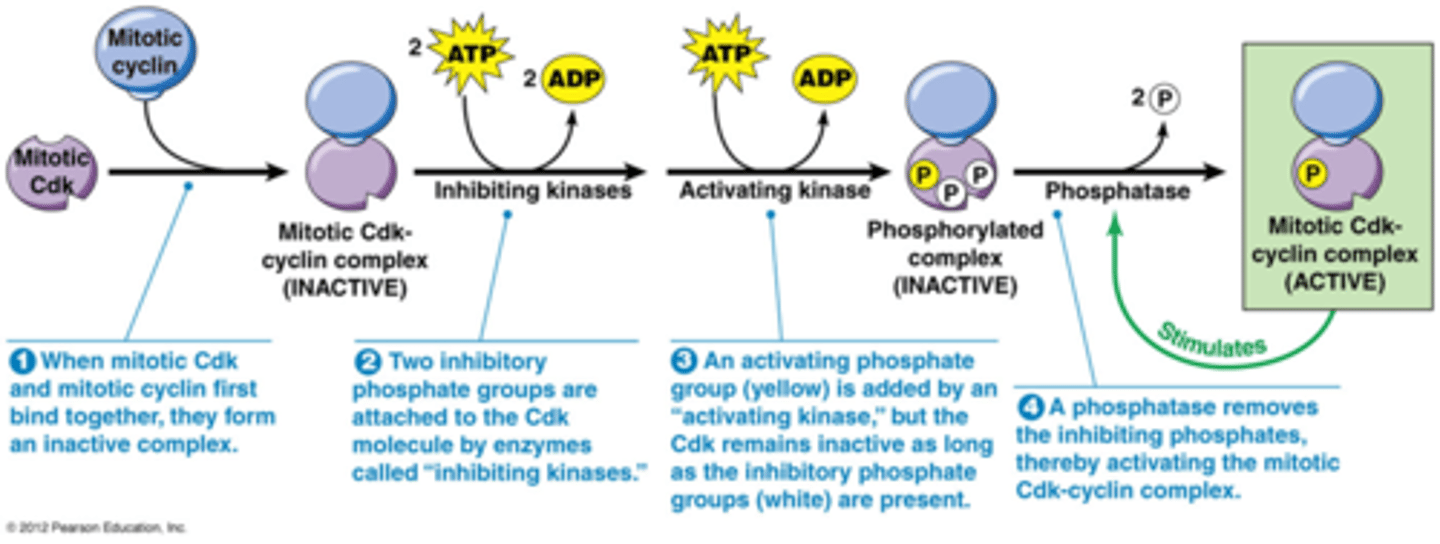
Wanneer zijn de Cdk's actief? Hoe werkt de regulatie?
1. Cdk's moeten een cycline gebonden hebben om actief te worden
2. bijkomende regelering van Cdks door fosforylering en defosforylering.
3. Het wordt geregeld door p21: een Cdk inhibitor (zie kaartje 43)
Je krijgt 3 fosforyleringen
2 door een kinase dat inhibitie geeft
1 door een activerend kinase
->effect van die 2 soorten in verschillend
Om een actief kinase te hebben moeten de gele fosfaten gedefosforyleerd zijn door een fosfatase
Het actief molecule heeft een positieve feedbackloop, het zorgt voor meer defosforylatie
->plotse stijging in Cdk activiteit
Bv. MPF (mitotisch cycline-Cdk1)
•Enkele actieve moleculen cycline B –Cdk1 kunnen alle cycline B-Cdk1 complexen activeren
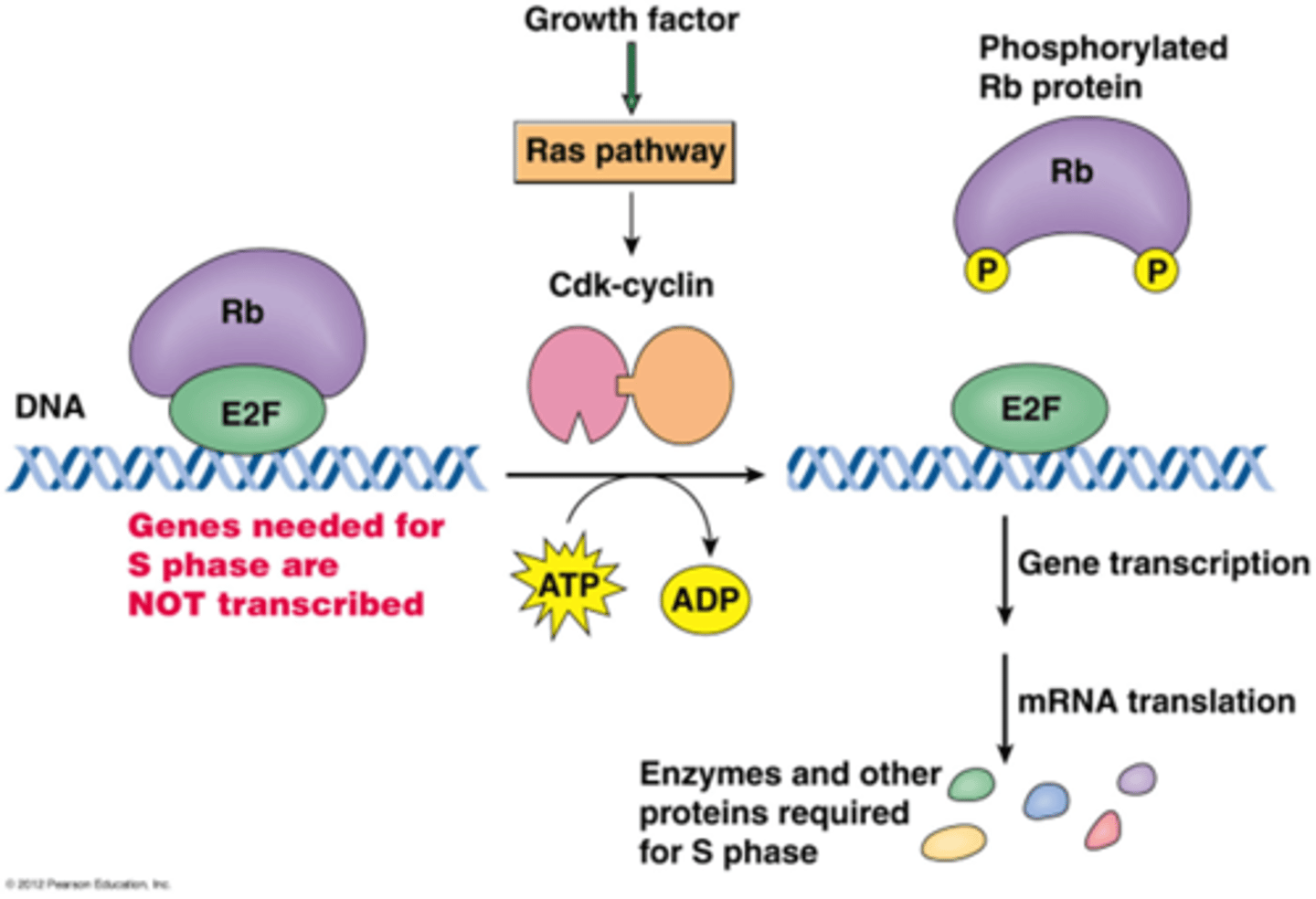
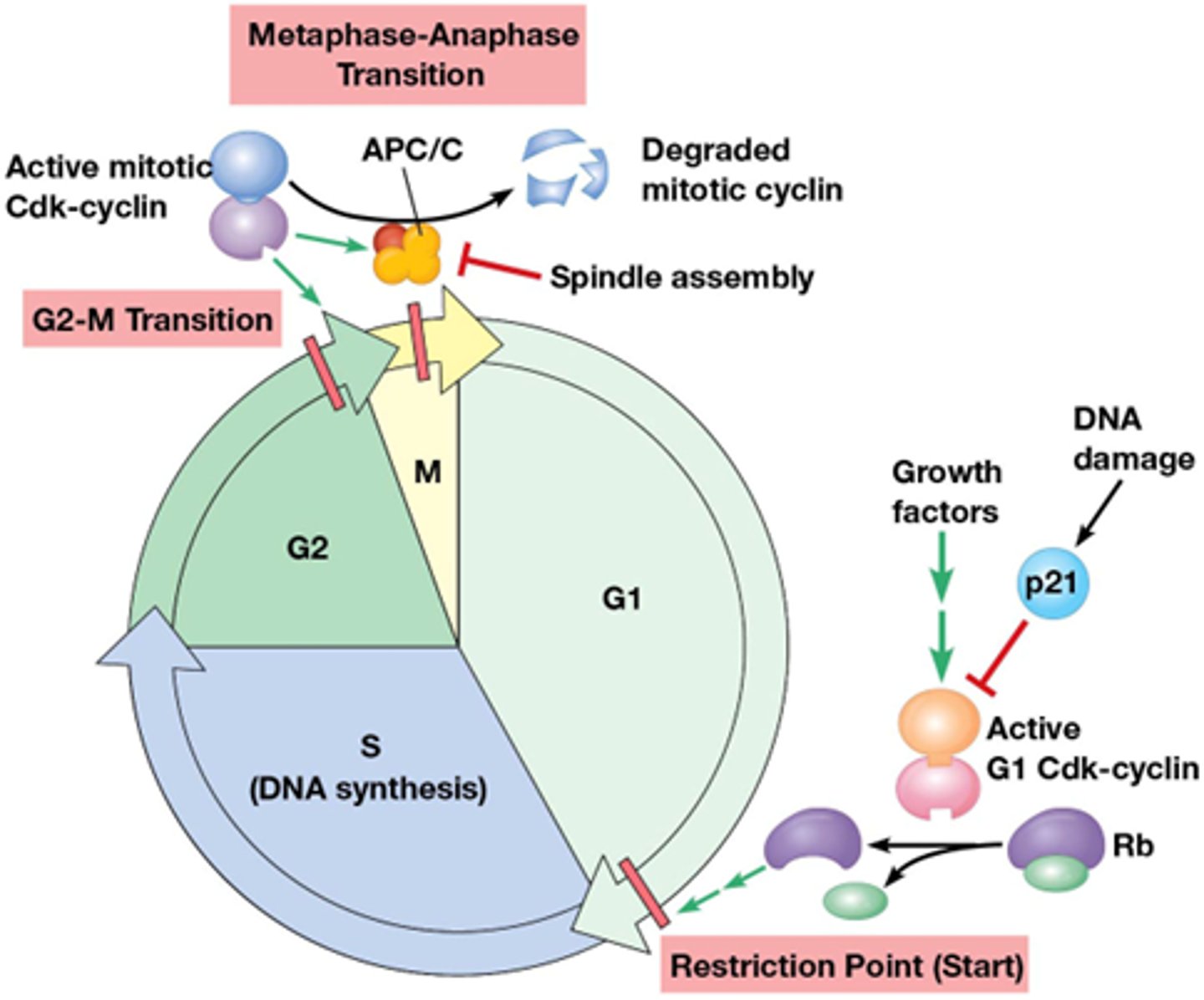
Wat gebeurt er in de G1/S transitiepunt?
Eens het in de S fase gaat ga je eigenlijk de mitose ook krijgen, dit moet dus sterk geregeld worden door de Cdk's (G1 Cdk en G1 cycline)
-een groeifactor gaat de Ras pathway activeren -> activatie van Cdk Cycline complex
•G1 Cdk fosforyleert retinoblastoma eiwit (Rb): een eiwit op E2F ( een transcriptiefactor) waardoor E2F wordt vrijgesteld.
->E2F induceert transcriptie van replicatiefactoren
->translatie en synthese van eiwitten die nodig zijn voor de S fase (eiwitten voor de replicatie)
Alle eiwitten en hun functie kennen
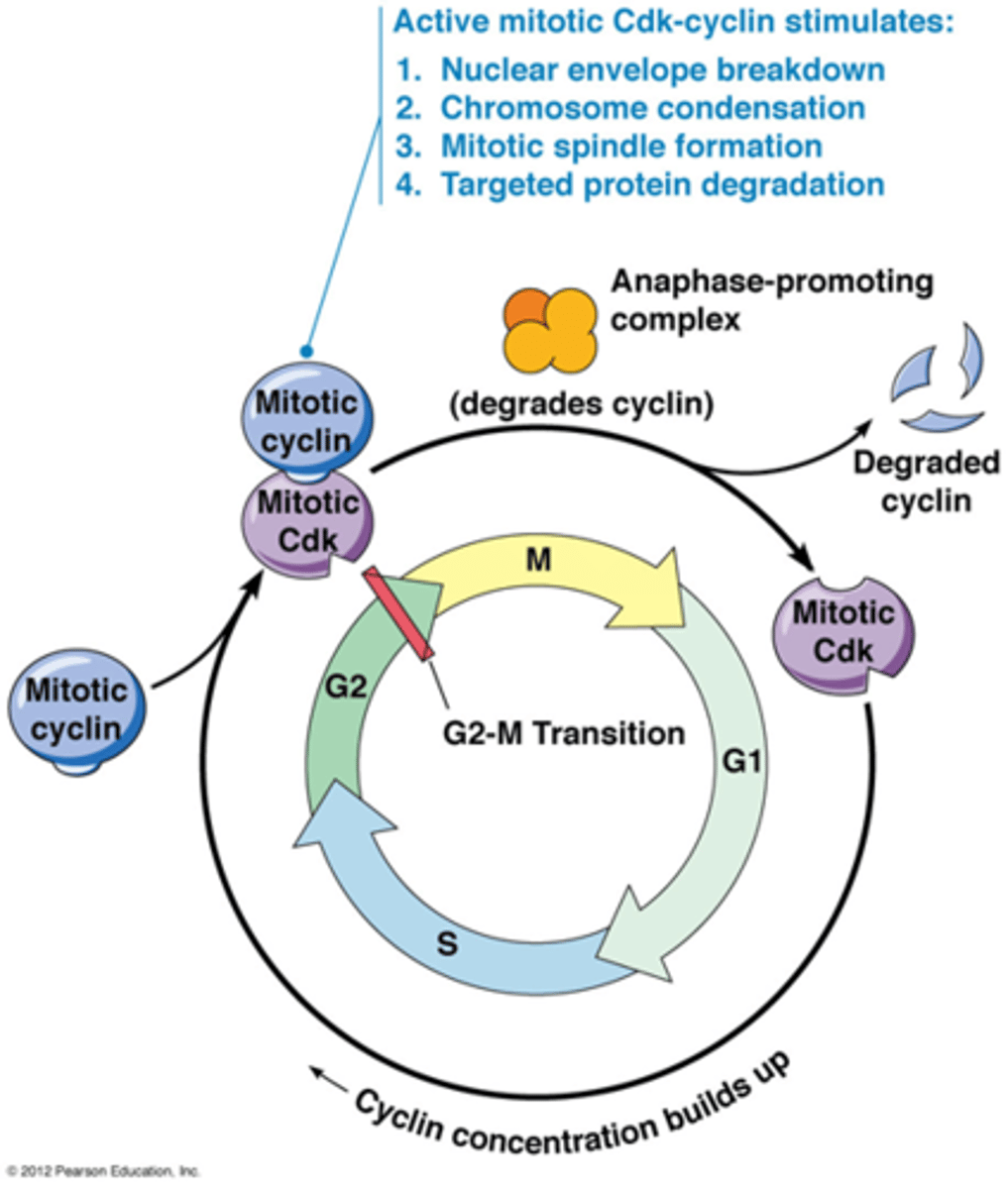
Hoe verloopt de G2/M transitie?
Fosforyleringen gebeuren door Cdk's
•Fosforylering laminen → uiteenvallen celkernmembraan
•Fosforylering condensine → chromosoomcondensatie
•Fosforylering van MT geassocieerde eiwitten → vorming kernspoel
•Fosforylering + inactivering van eiwitten betrokken in DNA herstel (voorkomen fusie van telomeren)*
Alle fosforyleringen kennen zonder te veel uitleg/details
*De DNA herstelmechanismen: zijn niet actief in mitose omdat de cel dan denkt dat de chromosoomuiteinden een ds breuk is waardoor die chromosomen aan elkaar gelinkt worden
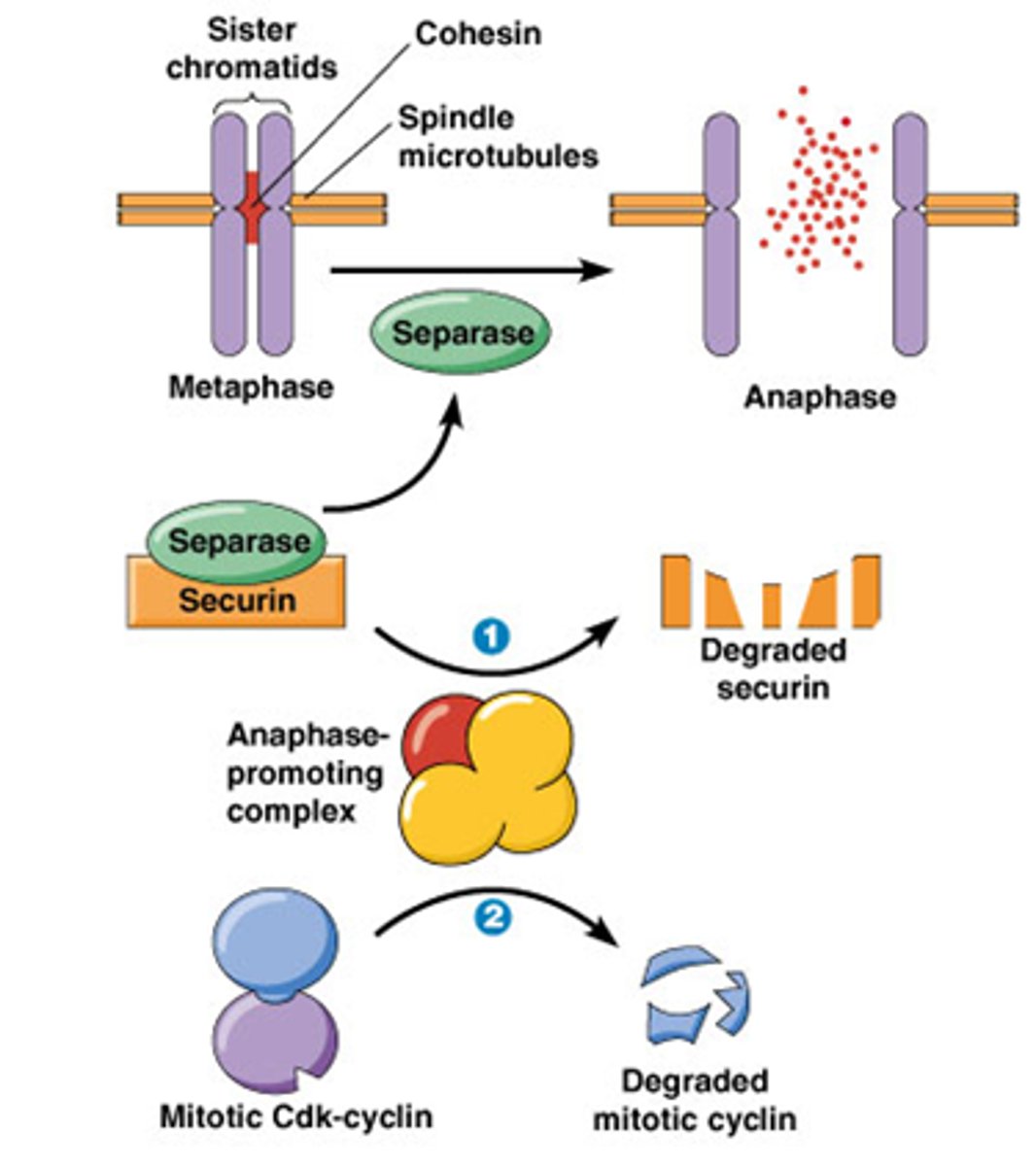
Wat is het Het anafase-promoting complex (APC) tijdens metafase-anafase transitie
Metafase anafase transitie, belangrijk want de zusterchromatiden moeten uit elkaar gaan
-Het enzyme separase zal cohesine afbreken
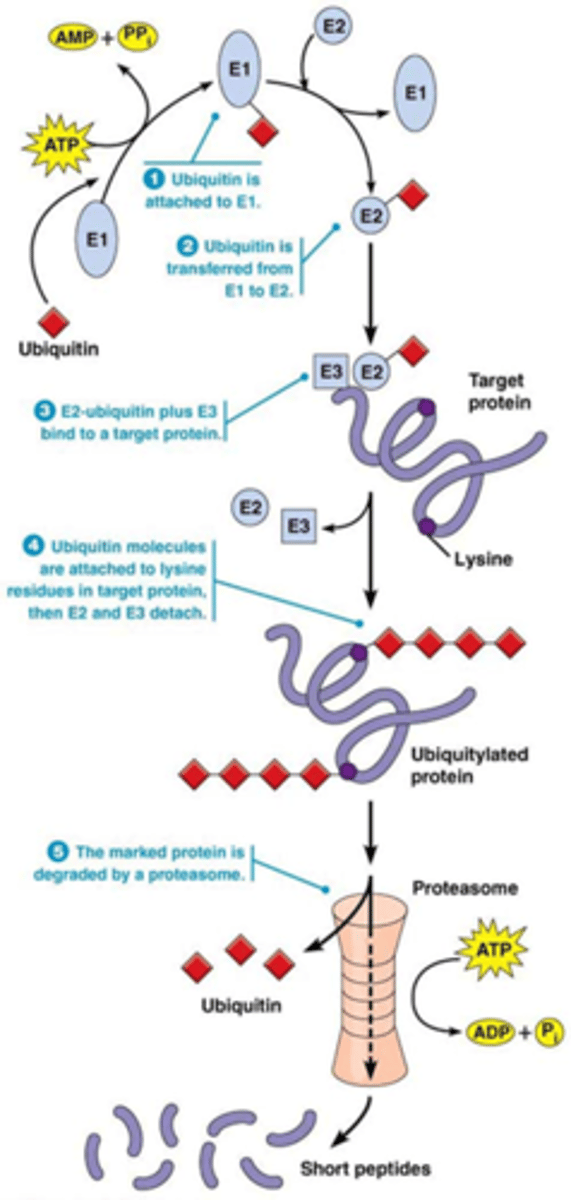
Securine en cycline is het substraat van het anafase promoting complex (APC) = E3 ubiquitine ligase: katalyseert de binding van ubiquitine aan eiwit; ge-ubiquitineerde eiwitten worden vervolgens afgebroken door proteasoom.
•APC ubiquitineert securine en cycline
->Securine en cycline wordt afgebroken, separase komt vrij
We zien nu al 2 eiwitten maar eigenlijk is er ook een derde substraat: geminine dat wordt geubiquitineerd door APC op het einde van de mitose (zie kaartje 40)
vierde substraat is Mad-Bub dat bindt op Cdc20 (subeenheid van APC) -> mid mitose controlepunt (zie kaartje 41)
Hoe verloopt de afbraak door het proteasoom? herhaling denk ik?
Hoe wordt het DNA gecontroleerd,
We hebben 5 controlepunten en 3 daarvan overlappen met het transitiepunt
Een transitiepunt is ook een controlepunt maar niet elk controlepunt is een transitiepunt
(1)G1/S transitiepunt -> E2F, Rb
(2)G2/M transitiepunt: replicatie-controlepunt (‘DNA replication checkpoint’) -> mitotisch CDK/mitotisch cyline, ATR kinase
(3) Metaphase-anafase transitiepunt : Mid-mitose controlepunt (‘Spindle assembly checkpoint’) -> APC complex + Mad, Bub, cdc20
(4)tijdens de hele S-fase: Replicatie licensing -> ORC/helicaseladers/MCM/geminine (zie volgend kaartje)
(5) DNA-schade controlepunt (actief in verschillende fasen) -> p53/Mdm2, p21, puma (zie volgend volgend kaartje)
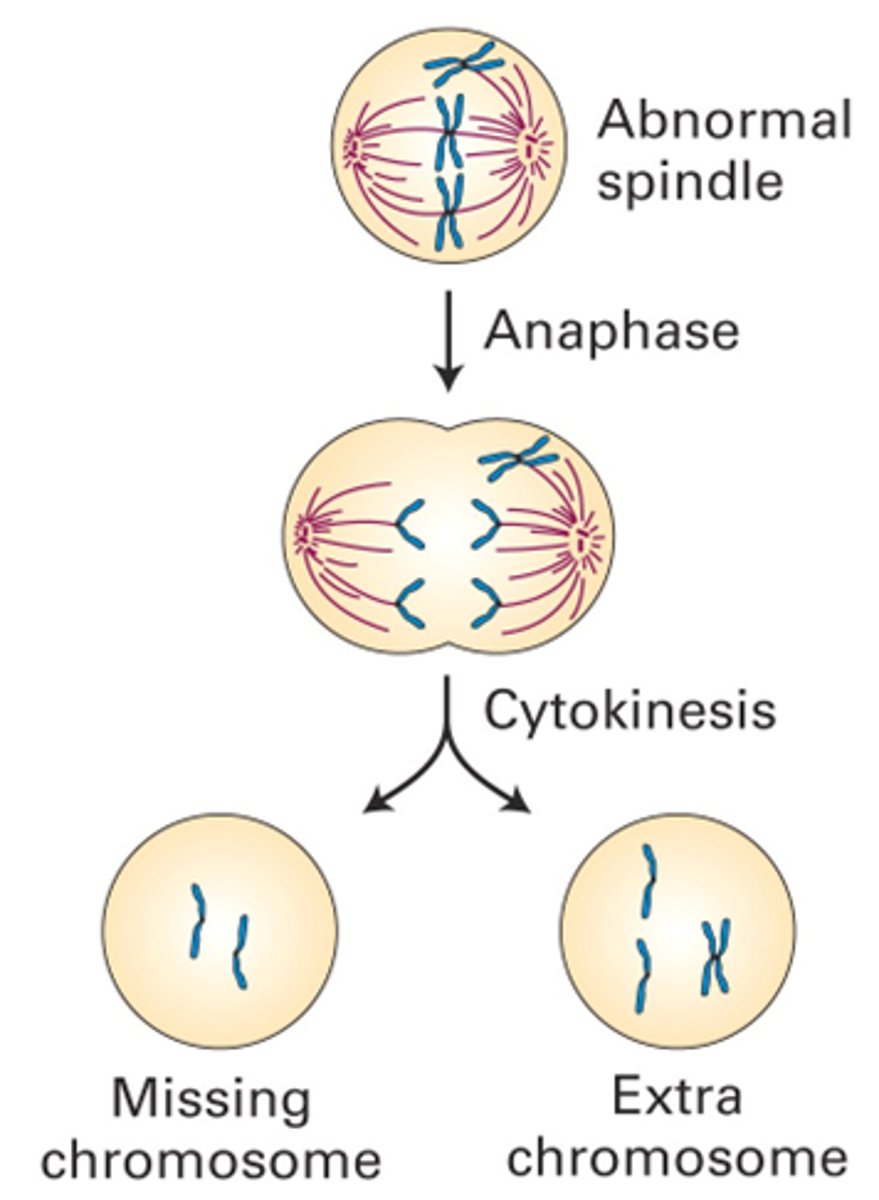
Controlepunten belangrijk: Als het fout loopt ga je iets fout krijgen in het genoom, bv foto
->2 extra controlepunten die niet overeenkomen met de transitiepunten
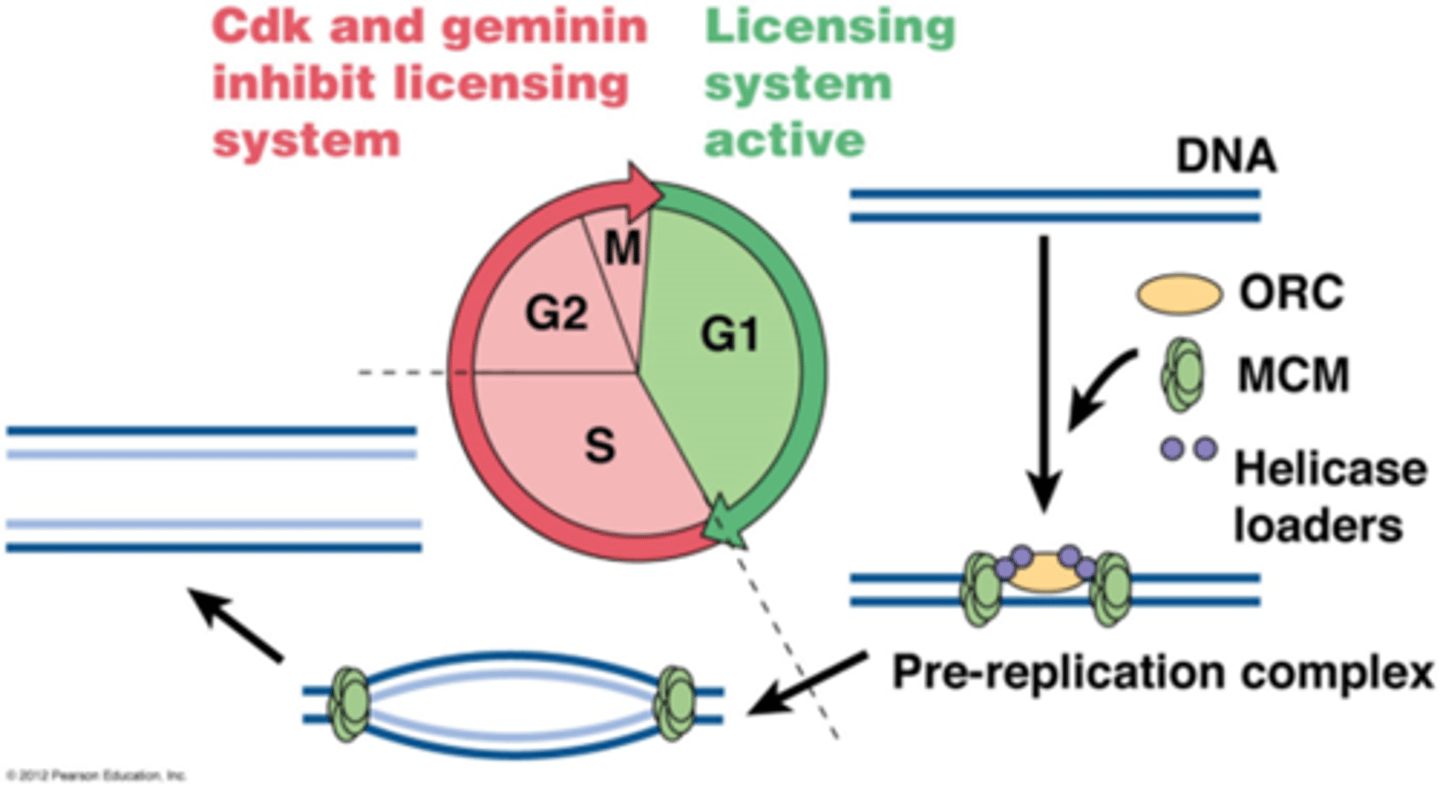
Hoe wordt het prereplicatiecomplex geactiveerd?
•DNA kan maar éénmaal worden gerepliceerd tijdens S-fase
->in de G1 fase worden eiwitten gevormd die gaan binden op de ORI: ORC, MCM en helicase loaders= pre replicatiecomplex
S-fase Cdks fosforyleert MCM, ORC en helicase laders
-> activatie van replicatie
-> fosfo-ORC and fosfo-helicase laders dissociëren van DNA, voorkomt hervergunning
-> fosfo-MCM (DNA helicase) haalt de 2 DNA strengen uiteen tijdens de replicatie
-> als de replicons samenkomen komt fosfo-MCM los van het DNA
-> geminine bindt fosfo-MCM en blokkeert de recrutering van het MCM complex naarhetDNA, geminine is aanwezig tot einde mitose.
• Geminine is substraat voor APC complex (anafase), geubiquitineerd geminine wordt afgebroken door proteasoom
• ORC and helicase laders worden gedefosforyleerd in mitose
• In G1 fase terug vorming van pre-replicatie complex
(aleyde is niet zo'n fan van de foto dus meer kijken naar de uitleg )
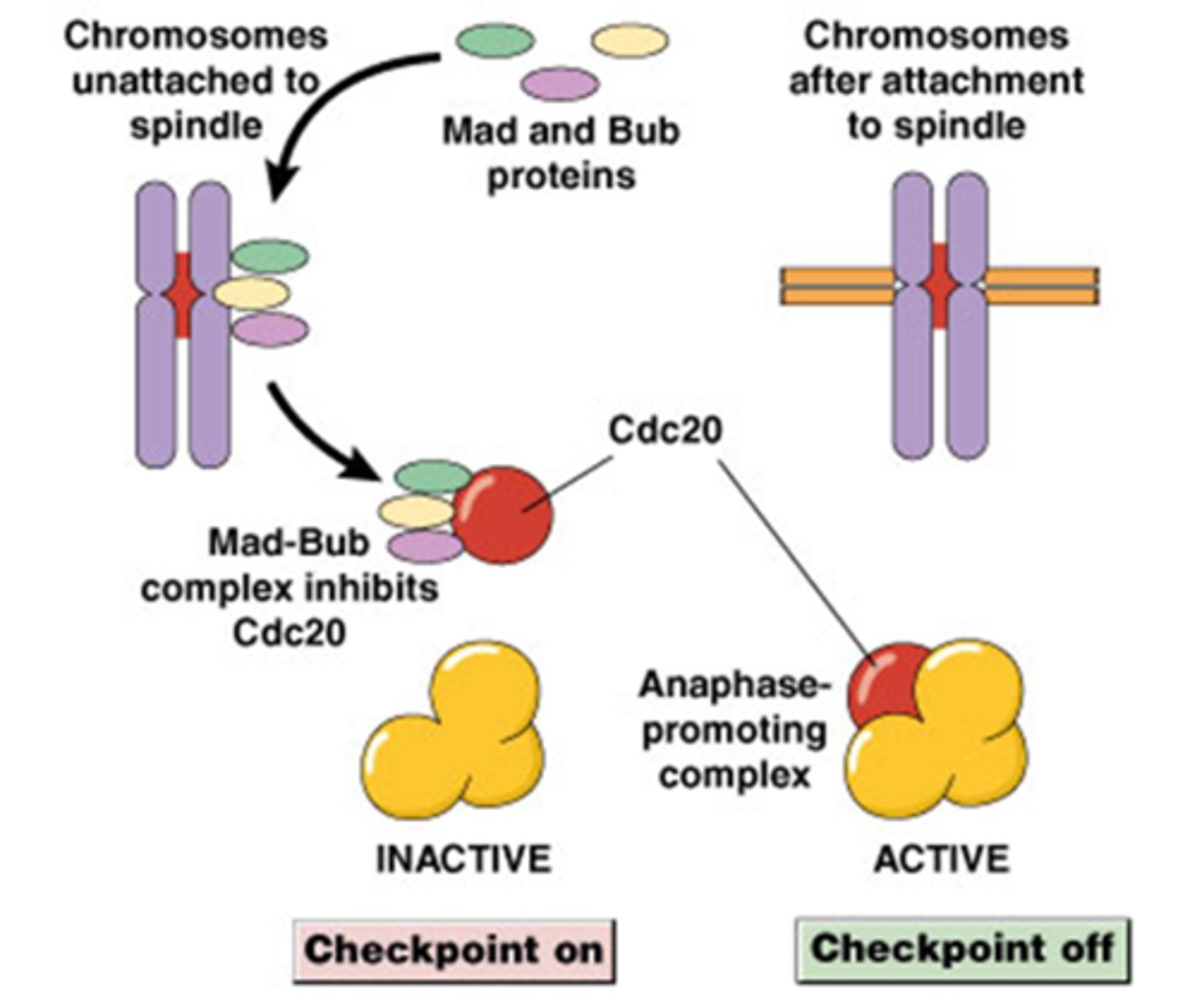
Wat is het mid-mitose controlepunt? Hoe gebeurt die regulatie?
dit is bij de metafase-anafase transitie
De Mad en Bub eiwitten (namen kennen)
Cdc 20 is een subeenheid van APC complex (dit complex moet ook geactiveerd worden)
Mad-Bub complex inhibeert Cdc20
-Als Cdc 20 geassocieerd is met Bub eiwitten dan is het APC complex inactief (het zal niet ubiquitineren)
Als alle kinetochoren gebonden zijn aan MTs, kan Cdc20 het APC activeren
-Als Cdc 20 niet gebonden met Bub is gaat het APC complex actief zijn
Hoe gebeurt die regulatie?
-Een kinetochoor dat geen MT gebonden heeft heeft een fibreuze corona, deze worden herkent door Mad-Bub
->Als er nog 1 kinetochoor is dan zal er geen APC geactiveerd worden!
Wat is het DNA-replicatie controlepunt?
Een controlepunt dat ligt bij de G2-M transitiepunt
Je moet zien dat het volledig gerepliceerd is, en geen DNA schade aanwezig is
Werking door eiwitkinase ATR: herkent replicatievorken, inactiveert het fosfatase (pijl is niet getekent)
- als het kinase geinactiveerd is ga je de mitotische cdk complex kunnen activeren
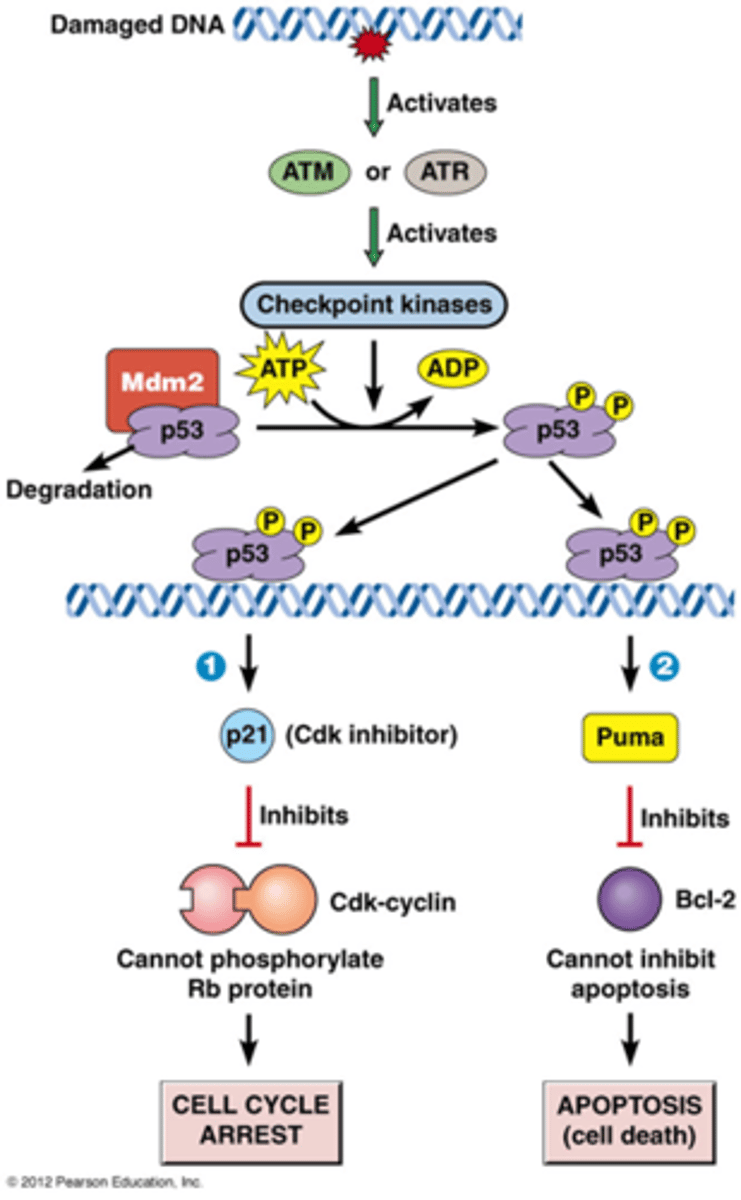
Wat is het DNA-schade controlepunt?
Resulteert in inhibitie van G1, S of G2/M Cdks
• Checkpoint kinasen fosforyleren en stabiliseren p53
• p53 (regulerende transcriptiefactor) induceert expressie van p21 (Cdk inhibitor) en DNA-hersteleiwitten of Puma (apoptose stimulator)
• MDM2= E3 ubiquitine ligase
DNA schade -> herstelmechanismen
DNA schade in de celcyclus ->celcyclus moet gestopt worden om dan hersteld te worden
Massieve DNA schade -> apoptose
Als er mutaties zijn in deze pathway heb je een groot probleem want dan werkt bestraling enzo niet meer voor een bepaalde kanker
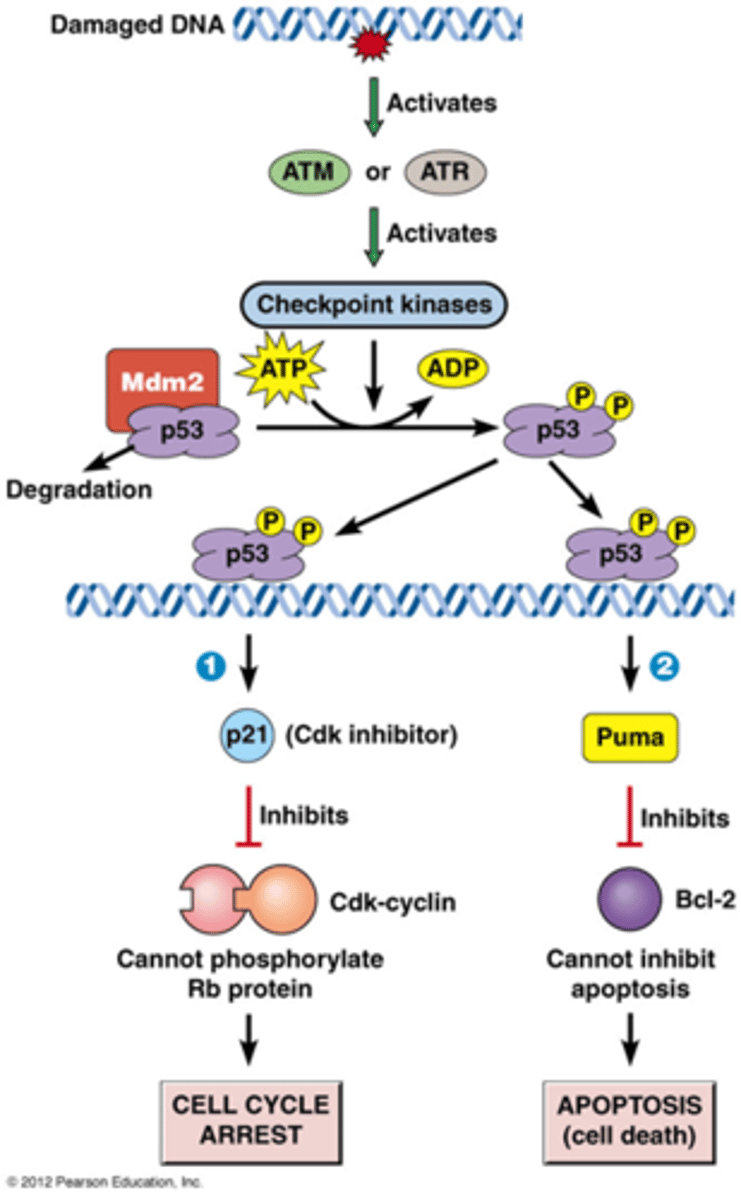
Hoe DNA schade leiden tot een 'cell cycle arrest'?
-ATM of ATR (afhankelijk van de schade) zijn eiwitkinases die geactiveerd worden door DNA schade
-ATM of ATR fosforyleert checkpoint kinases: deze cascade zorgt voor amplificatie
-checkpoint kinasen fosforyleren verschillende substraten bv p53: een regulerende transcriptiefactor
->p53 is normaal gezien geassocieerd met Mdm2 (een E3 ubiquitine ligase), normaal gezien wordt p53 bijna onmiddellijk afgebroken, het wordt ook constant gesynthetiseerd
->als p53 gefosforyleerd is dan wordt het eiwit gestabiliseerd
-p53 bindt op het DNA en regelt dan het gen p21 (een Cdk inhibitor)
-p21 inhibeert dus deze Cdk-cycline waardoor de cel in pauze gaat: cell cycle arrest
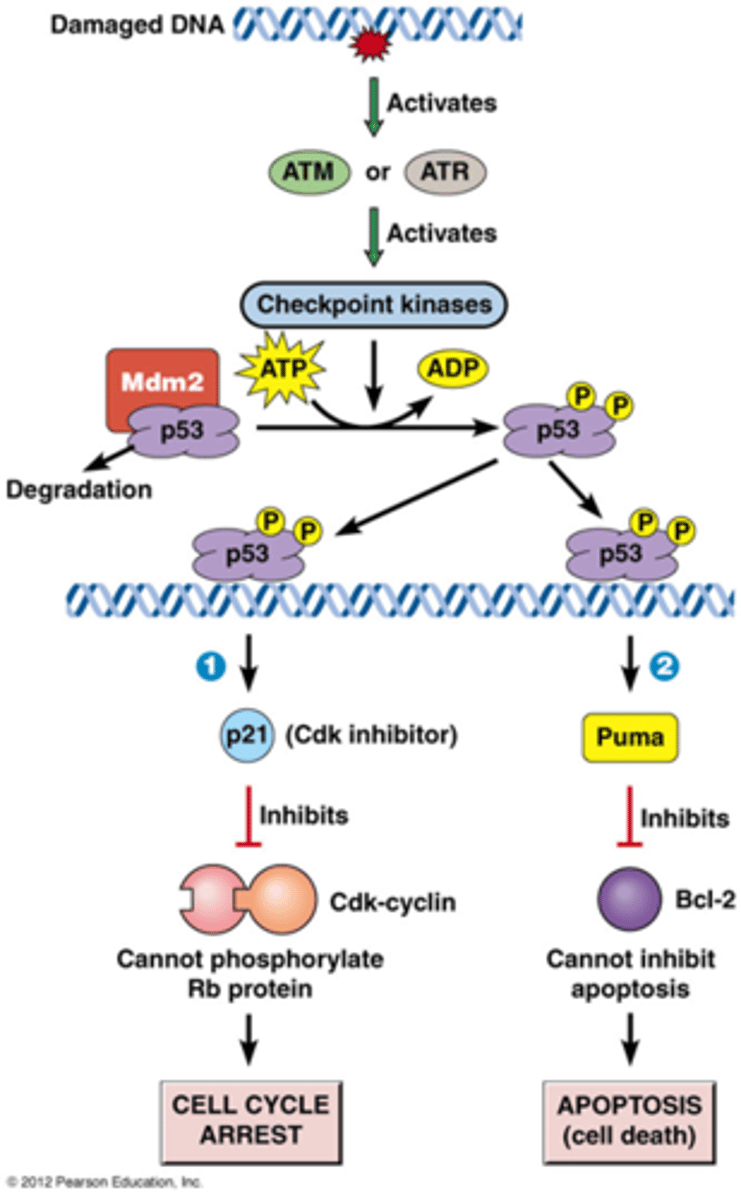
Hoe kan DNA schade leiden tot apoptose
eerste deel is herhaling van vorige kaartje:
-ATM of ATR (afhankelijk van de schade) zijn eiwitkinases die geactiveerd worden door DNA schade
-ATM of ATR fosforyleert checkpoint kinases: deze cascade zorgt voor amplificatie
-checkpoint kinasen fosforyleren verschillende substraten bv p53: een regulerende transcriptiefactor
->p53 is normaal gezien geassocieerd met Mdm2 (een E3 ubiquitine ligase), normaal gezien wordt p53 bijna onmiddellijk afgebroken, het wordt ook constant gesynthetiseerd
->als p53 gefosforyleerd is dan wordt het eiwit gestabiliseerd
2. p53 kan bij massieve DNA schade (bv bestraling) kiezen voor apoptose (geprogrammeerde celdood)
-p53 zorgt ervoor dat Puma tot expressie komt, Puma is een pro apoptose, het bevordert de apoptose
-Puma gaat het Bcl 2 eiwit inhiberen, dit is een eiwit dat de apoptose inhibeert
-het inhiberende ding wordt geinhibeerd waardoor het wel door gaat
->cel gaat in apoptose!
Wat zijn de uitdagingen en oplossingen van de regeling van de celcyclus?
Uitdagingen:
•Juiste volgorde van de fasen
•Voorgaande fase moet beëindigd zijn
•Respons aan interne en externe stimuli (DNA schade; kinetochoor-microtubule interacties; omgevingsfactoren..)
Oplossingen:
•Specifieke kinasen (CDKs) die overgangen regelen
•Controlepunten: G1-S, S, G2-M, mid-mitose, DNA schade)
•Apoptose
Waarom kan je de celcyclus zien als een controleerbare klok? (domme vraag ma idk hoe ik dit er anders in zet)
•Klok: cycline synthese + afbraak
•Fine-tuning: via (de)fosforylering + Cdk inhibitoren
•Interne controle: checkpoints
•Externe controle: groeifactoren
24.4. Groeifactoren en celproliferatie
Hoe wordt de celcyclus geregeld bij verschillende soorten organismen?
Prokaryoten, unicellulaire eukaryoten: voeding
Meercellige eukaryoten:
•Voeding en coördinatie tussen cellen (in tegenstelling tot kankercellen)
•Groeifactoren (mitogenen): nodig om restrictiepunt (G1/S) te passeren (serum, EGF, PDGF)
•Groei-inhibitoren: TGFβ → CDK inhibitoren
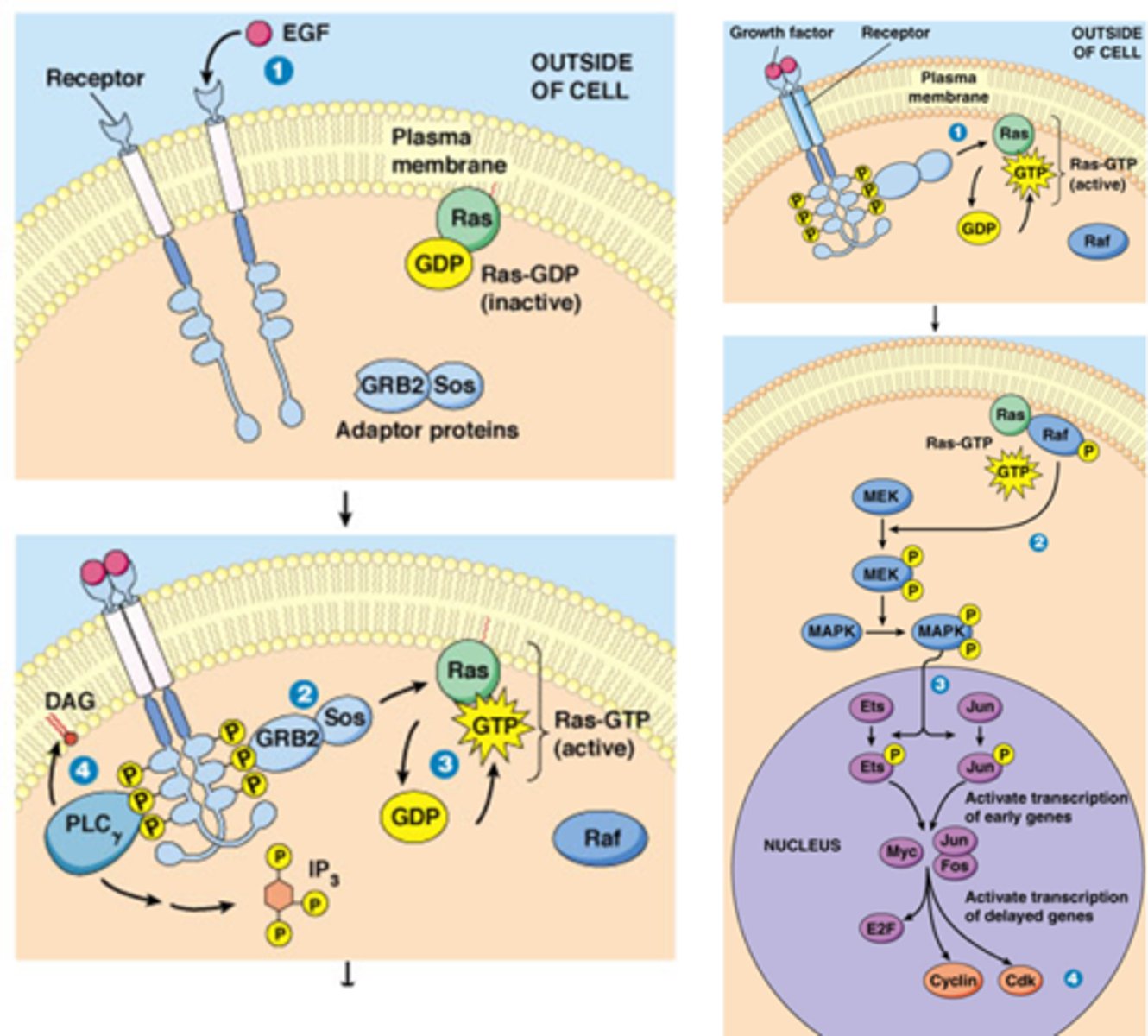
Hoe kunnen groeifactoren leiden dat een cel die in de G0 fase naar de S fase gaat?
- Groeifactor bindt op receptor ->dimerisatie
- activatie Ras GTP
- Raf eiwitkinase (autofosfolyryling)
- MEK eiwitkinase
- MAPK eiwitkinase
->gefosforyleerd naar de nucleus
- MAPK zal Ets transcriptie factor fosforyleren
-ets reguleert de expressie van early genes
- Early genes: coderen voor TF die nodig zijn voor G0-G1 overgang
- Late genes: coderen voor TF G1-S (E2F) en voor S-fase CDKs en S-fase cyclines
Enkel rechtsonder kennen maar alles voor celbio dusja
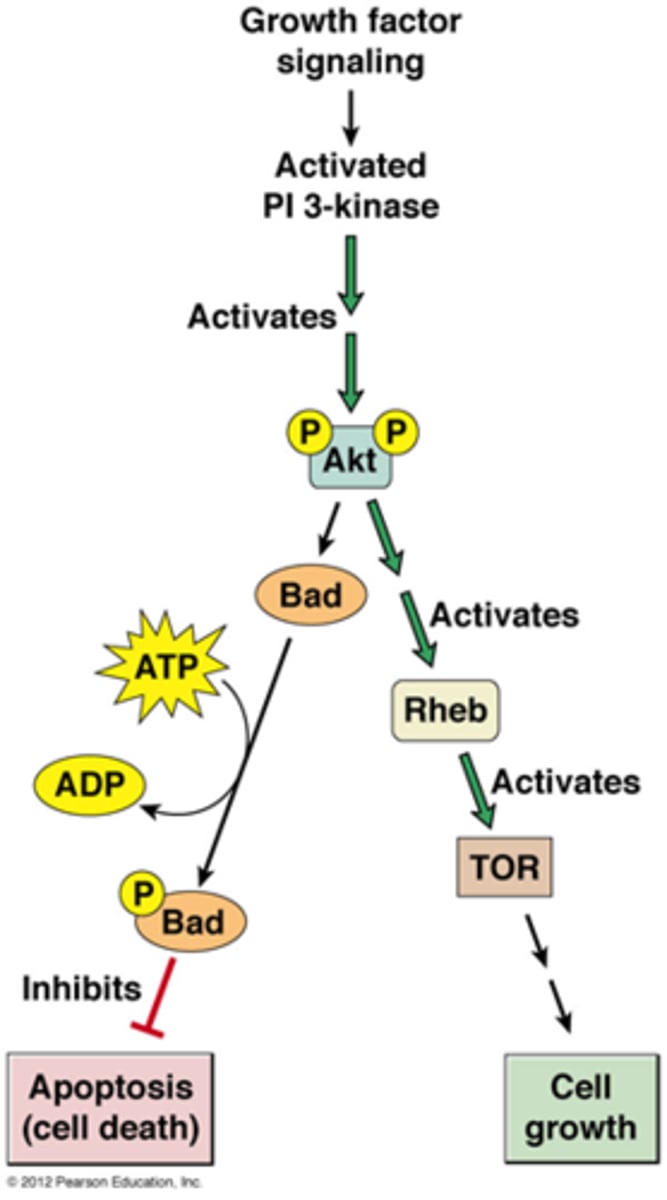
Hoe kan insuline zorgen voor celgroei of tot het inhiberen van apoptose
achtergrondkennis:
AKT=een overlevingskinase die apoptose verhindert
Bad= pro-apoptotic, fosfo-Bad is inactief
TOR kinase stimuleert eiwitsynthese en leidt tot celgroei, nodig omdat de cel voldoende groot zou zijn voor G1/S transitie
pathway:
*een AKT kinase wordt gefosforyleerd
-een actief AKT gaat Bad fosforyleren zodat de apoptose geinhibeerd wordt
-een actief AKT kan ook het eiwitkinase TOR activeren ->dit leidt tot celgroei
*zie celbio:
insuline bindt op receptor -> PIP2 wordt PIP3 ->activeert AKT door het te fosforyleren
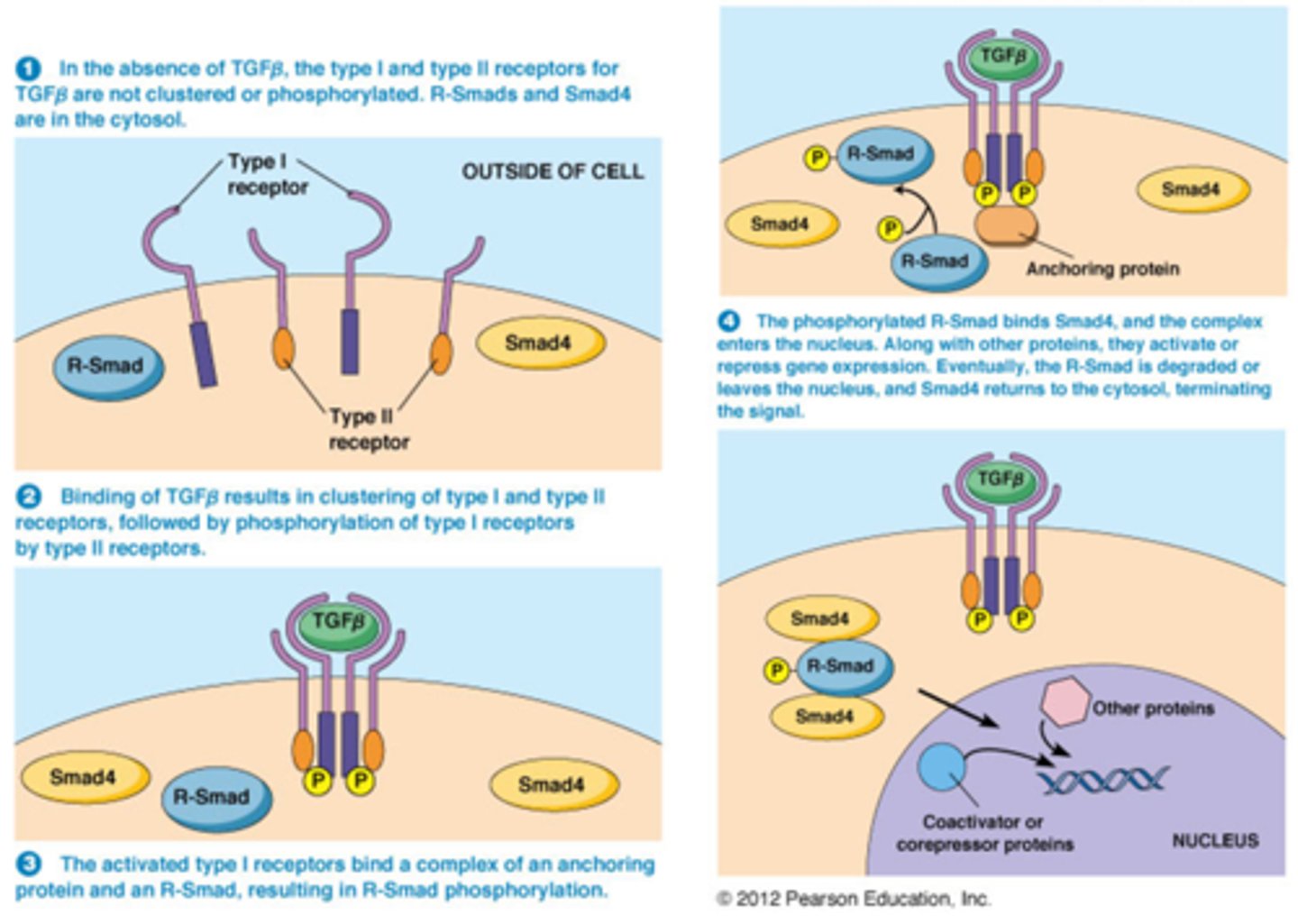
Hoe kunnen inhibitorische groeifactoren leiden tot de inhibitie van de celcyclus
• TGFβ transforming growthfactor
• TGFβ leidt tot synthese van inhibitoren van CDKs
Hoe?
• Binding van TGFβ op zijn receptoren leidt tot activatie van Smads
• Smads transloceren naar de kern en activeren genen bv. Genen die inhibitoren van CDKs tot expressie brengen bv p21 (de derde regulator van activiteit van CDK's)
24.5. Apoptosis
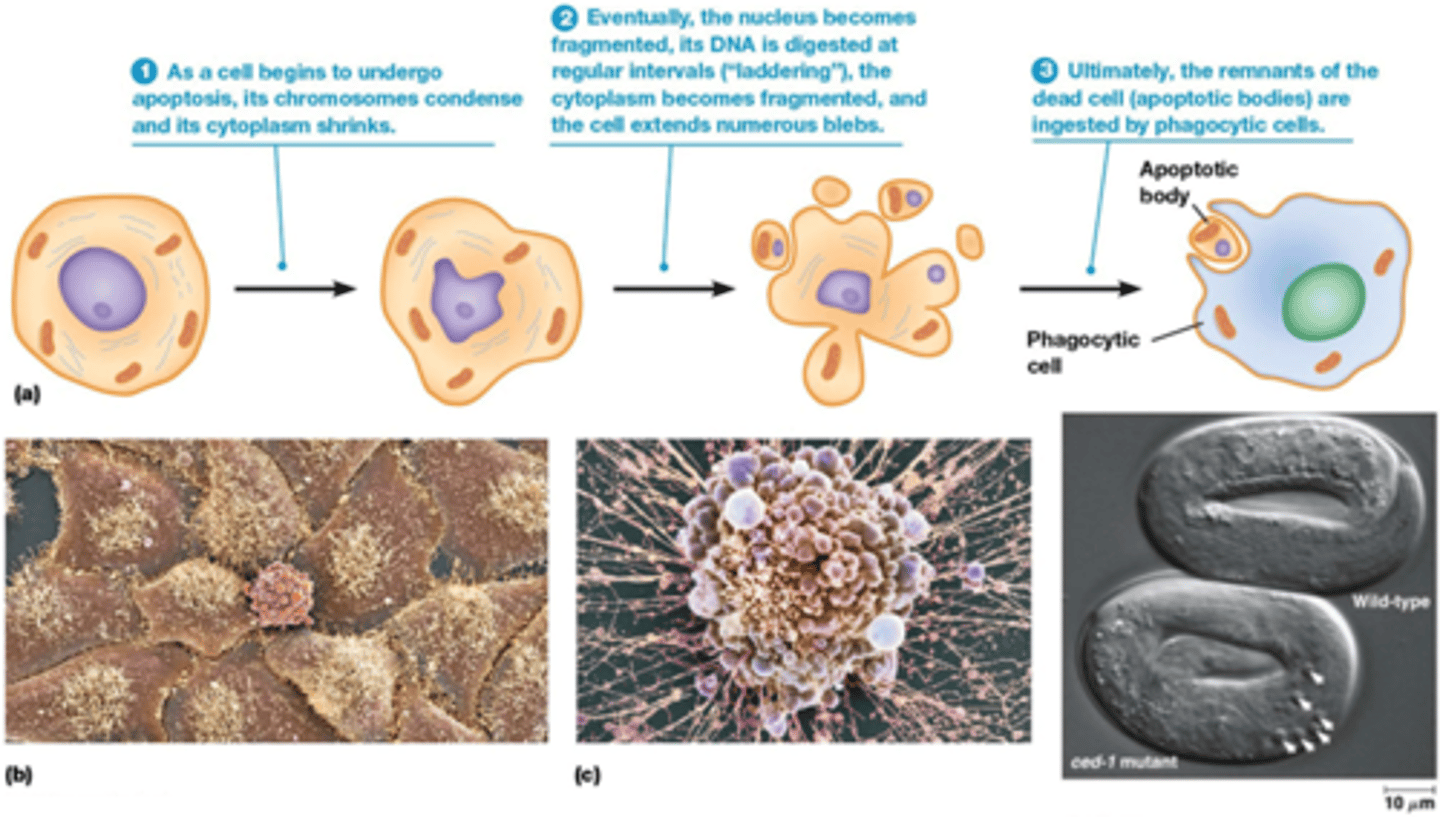
Wat is apoptose?
=geprogrammeerde celdood
• Belangrijk tijdens embryonale ontwikkeling en voor verwijdering van beschadigde cellen (C. elegans, ced genes; 1090 cellen, 131 cellen ondergaan apoptose)
->De cellen tussen de vingers gaan apoptose ondergaan waardoor je vingers krijgt
Hoe verloopt de apoptose?
-Chromatine condenseert
-cytoplasmasma krimpt
-Fragmentatie van dna
Blaasjes worden gevormd (ze hebben een membraan en er zitten stukjes dna in)
-Apoptotische body's worden opgegeten door een fagocyterende cel bv de macrofagen
->de celinhoud komt niet vrij in het cytosol
Wat is het verschil tussen apoptose en necrose?
bij apoptose gaat de celinhoud niet vrijkomen -> geen ontstekingsreactie/inflamatie
bij necrose gaat de celinhoud wel vrijkomen waardoor er wel een ontstekingsreactie start
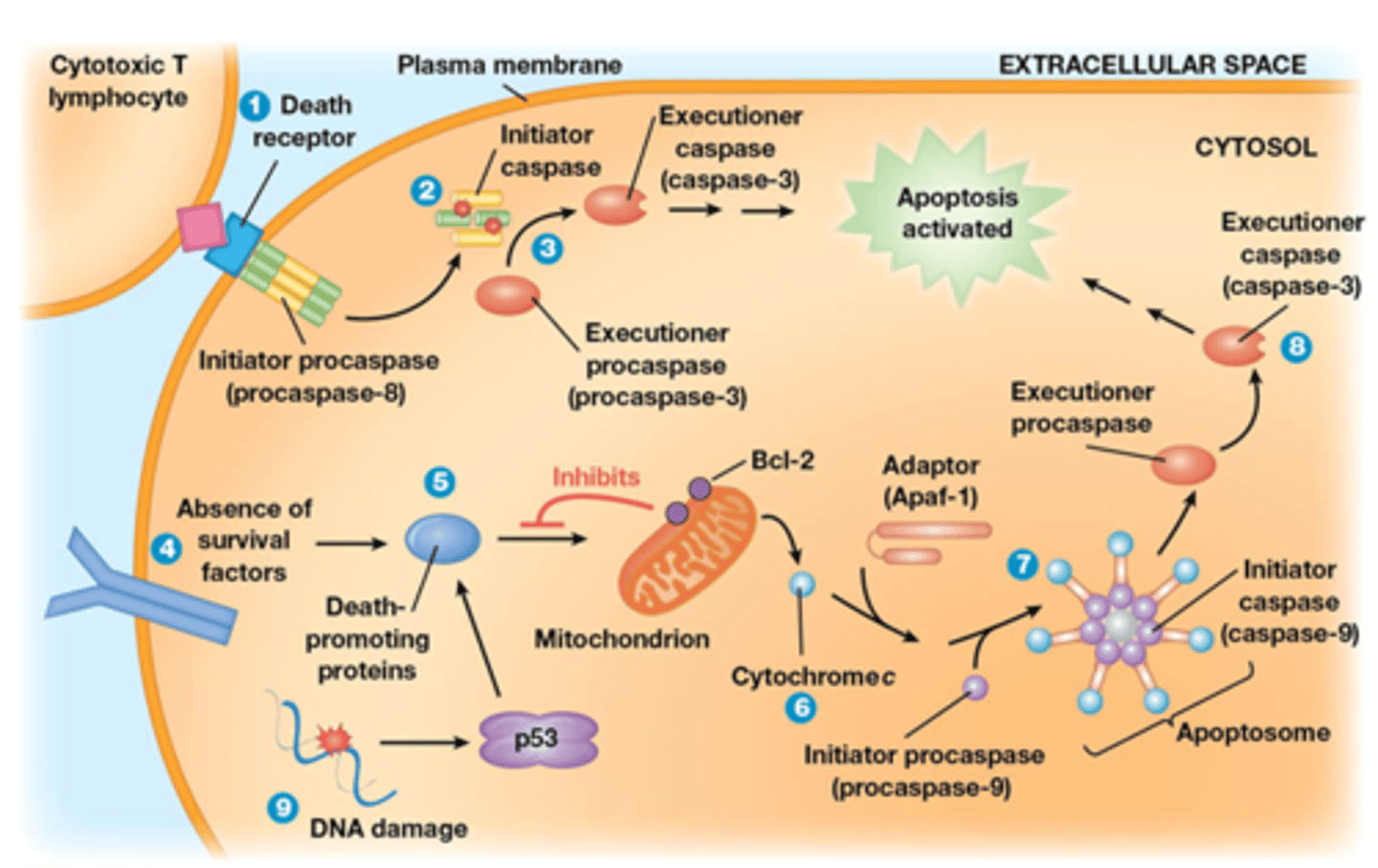
Hoe wordt de apoptose geregeld? Hele figuur kennen en ook eigenlijk alle moeilijke naampjes (niet de nummers achter de namen)
De regeling van apoptose gebeurt via 3 verschillende manieren:
1. death receptor vormt zich als trimeer het ligand (een witte bloedcel die leidt tot celdood) bindt
-death receptor is een procaspase dat geactiveerd is vanaf het een trimeer is
-actieve death receptor is een protease, dit zal eiwitten afbreken
-de actieve death receptor zal zichzelf proteoliseren
2. na de zelf proteolise gaat het initiator caspase vrijkomen
3. initiator caspase gaat een executioner caspase proteoliseren
->de caspase gaat nog andere eiwitten proteoliseren en dit gaat apoptose uitlokken
2 voorbeelden kennen van de Death receptor:
bv. Fas receptor (ligand is FasL)
Bv. TNF receptor (ligand is TNF)
4. Absence of SV (bv insuline) kan leiden tot apoptose Als AKT niet gefosforyleerd/geactveerd is zal Bad (blauw eiwit 5.) niet gefosforyleerd worden
-Bcl2 is anti-apoptose: op het buitenste membraan van mitochondrien
Bad wint: apoptose
Bcl2 wint: geen apoptose
Cyt C heeft nog functies naast de elektronen transportketen: het is een moonlighting eiwit!
6. Cyt C heel belangrijk in de apoptose, het wordt gesecreteerd
- Cyt C gaat interageren met een adaptor en met een initiator procaspase
7. Deze 3 dingen vormen een complex: het apoptosoom, het proteoliseert het executioner procaspase
8. dit leidt tot de vorming van executioner caspase
9. DNA damage
-leidt tot stabilisatie van p53
-expressie van PUMA (vb van blauwe eiwit 5), het inhibeert Bcl2
... zie hierboven bij absence of SV voor stap 6,7 en 8
https://mediaplayer.pearsoncmg.com/assets/sci-fwt-induction-of-apoptosis-by-cell-death-signals