HC4: Bacterial genetics and infection
1/31
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
32 Terms
Neisseria meningitidis (meningokok)
= bacterie die hersenvliesontsteking en bloedvergiftiging veroorzaakt
A-symptomatisch dragerschap: Ongeveer 20% van de gezonde bevolking draagt deze bacterie gewoon bij zich in de bovenste luchtwegen (de neus- en keelholte) zonder er ziek van te worden (a-symptomatic carriage). Je kunt dus drager en verspreider zijn zonder dat je het weet.
Wanneer wordt het gevaarlijk? Als de bacterie erin slaagt om door de slijmvliesbarrière heen te breken en de bloedbaan binnen te dringen (entry into the bloodstream), ontstaat er acute, levensbedreigende ziekte: sepsis (bloedvergiftiging) en/of meningitis (hersenvliesontsteking). Dit gebeurt meestal heel snel, binnen 10 dagen na kolonisatie.
Overleving in de luchtwegen: Om in die neus- en keelholte te kunnen overleven, moet Neisseria zich continu aanpassen en ontsnappen aan de antistoffen van ons immuunsysteem. Dat doet hij via antigene variatie. Hij verandert simpelweg continu zijn 'uiterlijk' (de eiwitten op zijn jas), zodat het immuunsysteem hem niet meer herkent.
Neisseria meningitidis: interactie met PorA
= dit is een porine, een groot transportkanaal in het buitenmembraan van de bacterie
De normale biologische functie: Het dient als een passief kanaal voor de diffusie van voedingsstoffen (nutrients) naar binnen. De bacterie heeft PorA dus keihard nodig om te eten.
De medische relevantie: Omdat PorA massaal aanwezig is aan de buitenkant, is het de major immunogenic protein in mensen. Ons immuunsysteem herkent dit eiwit direct en maakt er antistoffen tegen. Daarom is het een heel belangrijk vaccinkandidaat (vaccine candidate).
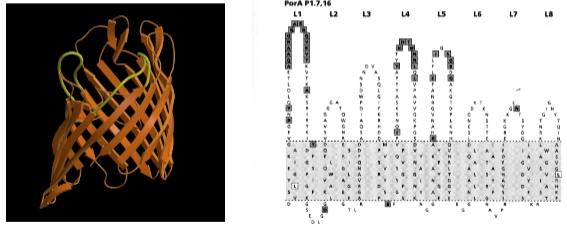
→ De genetische truc van PorA (De lussen variëren)
In de structuren onderaan de dia zie je hoe PorA is opgebouwd. Het is een beta-barrel (een koker van oranje bèta-sheets die in het membraan zit).
De lussen (loops): De delen van het eiwit die binnen het membraan zitten (het grijze vlak in het rechter diagram) veranderen nauwelijks. Maar de delen die aan de buitenkant uitsteken en blootgesteld worden aan de gastheer—de surface exposed loops (L1 t/m L8)—zijn hypervariabel.
Variatie in aminozuren: De bacterie verandert door mutaties en genetische variatie continu de volgorde van aminozuren (Amino Acid sequence) in deze lussen.
→ Wat is het gevolg hiervan?
Als jouw immuunsysteem antistoffen heeft gemaakt die precies passen op Lus 1 (L1) van PorA, dan overleeft de bacterie dat niet. Maar als er door een genetische variatie ineens een Neisseria ontstaat waarbij Lus 1 een net iets andere aminozuurvolgorde heeft, passen jouw antistoffen niet meer. Die bacterie ontsnapt aan de afweer, kan vrolijk blijven delen en infecteert de gastheer opnieuw.
Mogelijke gevolgen van mutaties
Affect active site of enzymes: Een mutatie verandert de aminozuren in het actieve centrum van een enzym. Hierdoor kan het enzym zijn werk niet meer doen (bijvoorbeeld het afbreken van voedingsstoffen).
Structural changes → instability or aggregation: De driedimensionale vorm van een eiwit verandert. Het eiwit wordt instabiel, vouwt verkeerd op, of klontert samen (aggregation), wat giftig kan zijn voor de cel.
Change binding sites: Belangrijke bindingsplaatsen veranderen, waardoor eiwitten niet meer kunnen communiceren met andere moleculen of receptoren.
Inactivate Promoters → expression lower: De 'aan-knop' van een gen (de promotor) raakt beschadigd. Hierdoor kan de RNA-polymerase het gen niet meer goed aflezen, en daalt de productie van dat specifieke eiwit (expression lower).
Selectiedruk
= de invloed van omgevingsfactoren (zoals de aanwezigheid van een antibioticum) die ervoor zorgt dat bacteriën met gunstige eigenschappen overleven en zich voortplanten, terwijl bacteriën zonder die eigenschappen afsterven.
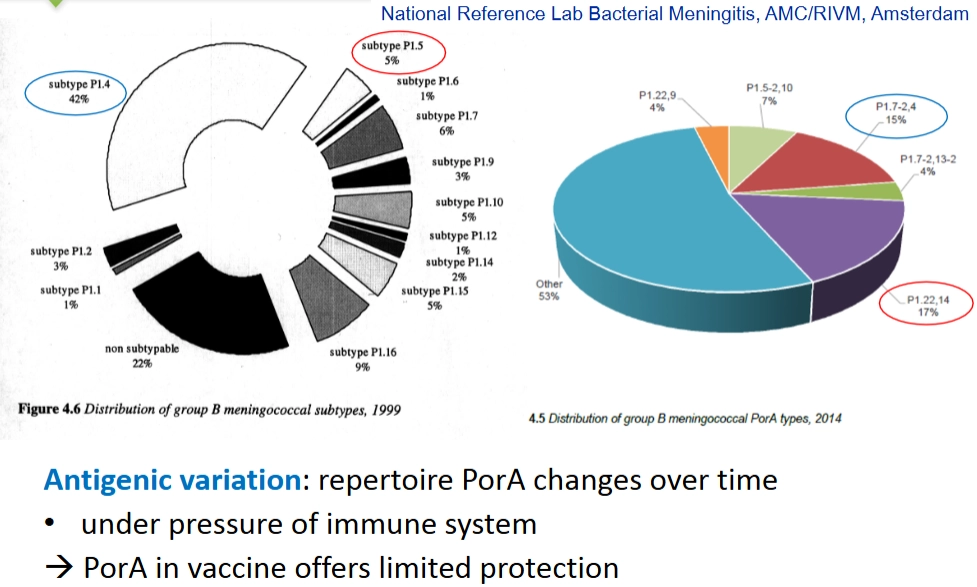
Selectiedruk: PorA
→ onder de druk van ons immuunsysteem verandert de totale verzameling (repertoire) van PorA-types over de jaren heen
Waarom moet DNA van een cel gerepareerd worden? (gevolgen)
Mutations in ORFs (Open Reading Frames): Een ORF is het specifieke stuk DNA dat daadwerkelijk codeert voor een eiwit (het gen zelf). Mutaties hierin kunnen leiden tot altered proteins (veranderde eiwitten). Dit tast de functie of de stabiliteit van het eiwit aan, waardoor vitale processen in de bacterie vastlopen.
Mutations outside ORFs: Stukken DNA buiten de genen coderen weliswaar niet voor eiwitten, maar bevatten wel de regelknoppen, zoals de promotor-regio's. Mutaties hierin veranderen de expressie: de bacterie maakt ineens veel te veel of veel te weinig van een bepaald eiwit.
Double-strand DNA breaks induce cell death: Dit is de gevaarlijkste vorm van schade: een volledige breuk door beide strengen van de DNA-helix. Dit blokkeert de replicatie (het kopiëren) en de transcriptie (het aflezen) van het DNA volledig, wat leidt tot acute celdood.
Link met antibiotica: Sommige antibiotica (zoals fluoroquinolonen of metronidazol) werken juist door dit soort dodelijke dubbelstrengsbreuken in bacteriën te forceren.
And for Eukaryotes (Het gevaar voor de gastheer): Bij meercellige organismen zoals mensen leiden ongerepareerde mutaties tot het verliezen van de celbalans (cell homeostasis). De cel stopt met luisteren naar weefselsignalen en kan veranderen in een kankercel.
DNA reparatiemethoden
Tijdens het kopiëren: Editing of DNA during replication
→ Proofreading (Kwaliteitscontrole): Terwijl het enzym DNA-polymerase een nieuwe DNA-streng bouwt, controleert het direct zijn eigen werk. Als het per ongeluk een verkeerde base (bouwsteen) plaatst, merkt het enzym dat aan de foute vorm van de helix. Het stopt, reist een stukje achteruit, knipt de foute base eruit en zet de goede erin. Dit voorkomt veruit de meeste mutaties.
Na het kopiëren: Removal of DNA damage after replication
Als er tóch schade door de kwaliteitscontrole glipt, of als het DNA later beschadigd raakt (door UV-straling, chemicaliën of radicalen), activeert de cel 3 hoofdtypen reparatie:Repair chemical adducts attached to DNA: Soms plakken schadelijke chemicaliën (adducten) vast aan de basen van het DNA (zoals bij blootstelling aan toxische stoffen). Speciale enzymen herkennen deze 'vlekken' op het DNA en knippen de beschadigde basen heel specifiek weg (bijvoorbeeld via Nucleotide Excision Repair of Base Excision Repair).
Remove gaps (missing bases in one strand): Als er gaten vallen in één van de twee strengen, gebruikt de cel de andere, tegenoverliggende streng als een blauwdruk. Omdat de basen altijd vaste paren vormen, weet de cel precies welke bouwstenen er in het gat gekopieerd moeten worden.
Join double-strand breaks: Als het DNA volledig in tweeën is gebroken, gebruikt de cel noodroutes om de losse eindjes weer aan elkaar te lijmen. Dit kan heel nauwkeurig door de blauwdruk van een ander chromosoom te kopiëren (Homologous Recombination), of snel en slordig door de uiteinden simpelweg aan elkaar te plakken (Non-Homologous End Joining), wat soms extra mutaties oplevert maar wel acute celdood voorkomt.
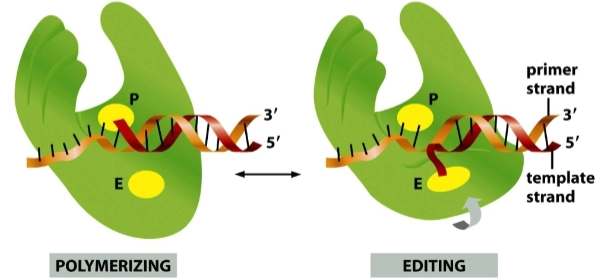
DNA polymerase tijdens de DNA-replicatie
= enzym heeft twee verschillende actieve locaties (sites): de P-site (Polymerizing) en de E-site (Editing/Exonuclease).
Links (Polymerizing): Onder normale omstandigheden bouwt het enzym de nieuwe DNA-streng op in de P-site. Het glijdt over de mal (template strand) heen en plakt de juiste basen aan de nieuwe streng (primer strand).
Rechts (Editing): Als het enzym per ongeluk een verkeerde base inbouwt, past de DNA-helix fysiek niet meer lekker in de P-site. Het DNA-molecuul is flexibel (uses DNA flexibility): de nieuwe streng raakt een soort van 'geknikt' en zwenkt uit naar beneden, rechtstreeks de E-site in.
In deze E-site (de exonuclease-afdeling) wordt de foute base er direct afgeknipt. Zodra de fout is weggesneden, veert de DNA-streng weer terug naar de P-site om de normale bouw voort te zetten.
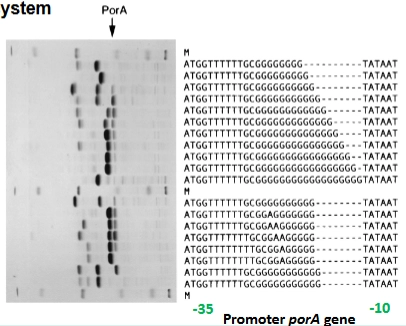
Falen van proofreading → phase-variation
→ laat zien hoe dit werkt in de promotor (de aan/uit-knop) van het porA-gen van Neisseria:
Promoter spacer sequences: Tussen de herkenningspunten van de promotor (zoals de TATAAT-box) zit een stukje variabele ruimte (de spacer). De lengte van deze spacer bepaalt hoe strak of los de RNA-polymerase aan de aan-knop kan binden.
De DNA-codes op de dia: Als je naar de letters kijkt, zie je dat de lengte van de rij G's per bacterie verschilt (sommige hebben 9 G's, anderen 11 of 12 G's).
Het gevolg voor expressie: * Heeft de spacer de perfecte lengte? Dan kan de RNA-polymerase er goed bij→ PorA expressie staat AAN.
Zorgt een proofreading-fout voor een G te veel of te weinig, waardoor de spacer te lang of te kort wordt? Dan kan de RNA-polymerase niet meer binden → PorA expressie staat UIT.
Phase-variation
= het willekeurig en herprikkelbaar aan- of uitzetten van een eiwit.
→ Als een hele kolonie Neisseria in jouw neusholte leeft en PorA aan heeft staan, maakt jouw immuunsysteem antistoffen tegen PorA. De bacteriën die PorA aan hebben staan, gaan allemaal dood (selectiedruk!). Maar door de ingebouwde 'foutjes' in de genetica, zijn er in die kolonie altijd een paar bacteriën waarbij het porA-gen door een slip van de polymerase net op UIT is gesprongen. Deze bacteriën hebben tijdelijk geen PorA op hun jas. De antistoffen herkennen ze niet, ze overleven de immuunaanval, en kunnen de infectie feilloos voortzetten.
Phase-variation in het ORF
ORF = Open Reading Frame, betekent in het midden van het gen
→ hierbij verandert de bacterie niet de hoeveelheid promotor-binding, maar saboteert hij direct de genetische code via een frameshift-mutatie
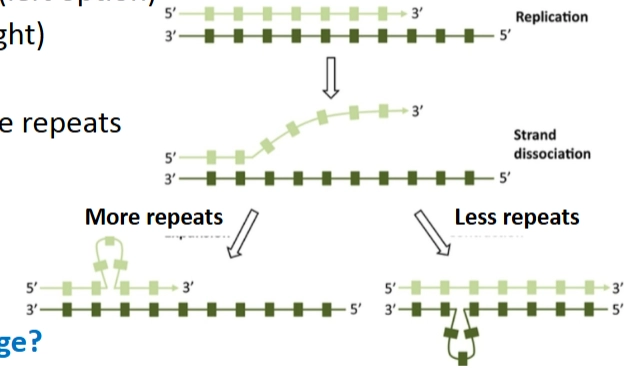
Het mechanisme
Strand dissociation (Loslaten): Tijdens het kopiëren laten de twee DNA-strengen heel even lokaal van elkaar los.
Slipped mispairing (Verkeerd terugplakken): Omdat er veel opeenvolgende identieke basen zijn, kunnen de strengen verschoven weer aan elkaar plakken.
More repeats (Linker optie): De nieuwe streng vormt een lusje naar buiten toe. Bij de volgende ronde zijn er extra nucleotides toegevoegd (+1, +2, etc.).
Less repeats (Rechter optie): De mal-streng (template) vormt een lusje. Hierdoor slaat de polymerase een base over en zijn er nucleotides verwijderd (-1, -2, etc.).
→ Waarom niet 3? De dia vermeldt specifiek: 1-, 2-, 4- of 5-nucleotide repeats (not 3). Omdat het biologische translationele systeem (het ribosoom) DNA afleest in groepjes van drie (codons), zou het toevoegen of verwijderen van exact 3 nucleotiden slechts één aminozuur toevoegen of weghalen. Het gen blijft dan grotendeels intact. De bacterie gebruikt juist herhalingen van 1, 2 of 4 letters om de boel bewust te ontregelen.
Consequentie van de verandering:
Shift of Reading frame: De complete afleesfase verschuift, wat leidt tot een totaal andere aminozuurvolgorde vanaf dat punt.
Premature stopcodon: Vrijwel altijd zorgt zo'n verschuiving ervoor dat er ergens verderop per ongeluk veel te vroeg een stopteken verschijnt. Dit resulteert in een afgebroken, niet-functioneel eiwit (truncated protein). Dit zet het eiwit dus effectief hard op UIT.
Phase variation inside ORF: voorbeeld
De normale situatie (0): Het ribosoom leest de codons netjes af:
GUG(Valine),CCU(Proline),GUU(Valine). Dit levert het normal protein op (bijvoorbeeld een functioneel oppervlakte-eiwit waar het immuunsysteem antistoffen tegen heeft).De Insertion (+1): Er glipt door een foutje een extra cytosine (
C, rood omrand) in de DNA-streng. Het mRNA verandert nu. Het tweede codon is nog steedsCCC(Proline), maar de extraCduwt de rest van de letters één positie op. Het derde codon verandert daardoor vanGUUinUGU(Cysteïne) en er blijft aan het eind een losseUover. Het eiwit verandert direct van structuur óf breekt vroegtijdig af.De Deletion (-1): Er wordt een
Cweggehaald uit het normale DNA. Het mRNA mist een letter. Het tweede codon slikt nu deGvan de buren in en wordtCUG(Leucine). Het derde codon schuift op en wordtUU...(Fenylalanine).
De evolutionaire truc voor de bacterie Als het eiwit door een +1 of -1 verandering compleet misvormd raakt of vroegtijdig stopt met groeien (truncated), verdwijnt dit eiwit van de buitenkant van de bacterie. Zodra de gastheer antistoffen maakt tegen het normale eiwit (0), overleven alleen de bacteriën met de +1 of -1 mutatie. En het mooie is: omdat dit een willekeurig proces is van 'uitglijden', kan een nakomeling van de mutant bij een volgende celdeling nóg een keer uitglijden (bijvoorbeeld weer -1 doen bij de +1 mutant), waardoor het frame weer terugspringt naar de normale situatie (0). Het is dus een volledig omkeerbare aan/uit-schakelaar die puur op geniaal gokwerk en kansberekening van de DNA-polymerase draait!
Types schade door het immuunsysteem
Oxidation: Radicalen veranderen Guanine (G) in het beschadigde 8-oxoG. Dit herstelt de bacterie met enzymen zoals MutM en MutY.
Deamination: Cytosine (C) verliest een aminogroep en verandert spontaan in Uracil (U). Uracil hoort in RNA, niet in DNA! Dit wordt gerepareerd door het enzym Ung.
Alkylation: Chemische groepen plakken vast aan het DNA. Dit herstelt de bacterie met Tag.
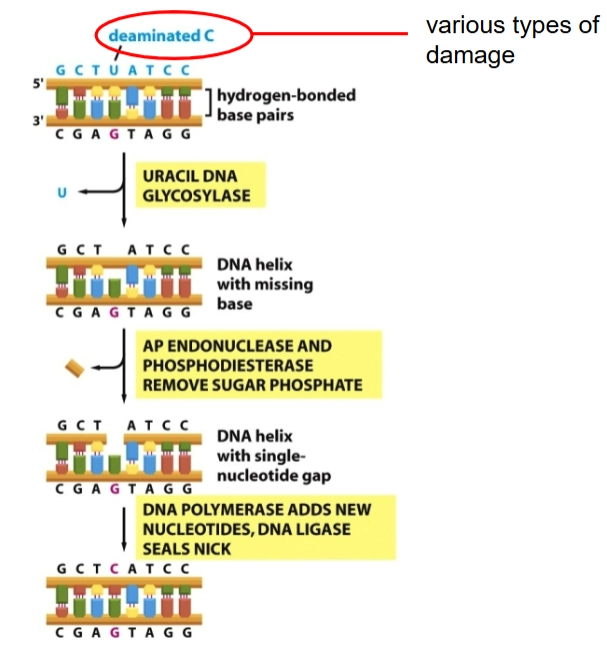
Base Excision Repair (BER)
= Als er één specifieke base beschadigd of veranderd is (zoals de deaminated C die nu een Uracil (U) is geworden), start de cel de Base Excision Repair (BER). Dit is een heel secuur proces:
Herkenning door Glycosylase: Het enzym Uracil DNA Glycosylase (Ung) reist over het DNA. Het controleert de basen door ze letterlijk naar buiten te flippen (base flipping). Omdat een beschadigde base geen waterstofbruggen (no h-bonding) kan vormen met de overkant, is de streng lokaal flexibel. Het enzym merkt dit, flipt de foute base eruit en knipt alleen de base (de letter) weg.
AP Endonuclease: Er blijft nu een DNA-streng over waar wel de suiker-fosfaatkabel zit, maar de letter mist (DNA helix with missing base / AP-site). Het enzym AP endonuclease knipt de suiker-fosfaatverbinding op die plek door.
DNA Polymerase & Ligase: DNA polymerase komt langs, ziet het gapende gat, leest de gezonde tegenstreng af en zet de juiste, gloednieuwe nucleotide (de letter + het suikervoetje) erin. DNA ligase plakt de boel weer definitief aan elkaar (seals nick).
Nucleotide Excision Repair (NER)
= Als de schade groter is, zoals bij Thyminediementen (caused by UV light) of grote chemische adducten, raakt de complete structuur van de dubbele helix vervormd (distortion). Dit kan niet met één letter tegelijk worden opgelost. De bacterie zet dan het UVR-systeem (NER) in:
Stap a (Scannen): Het eiwitcomplex UvrA en UvrB scant het DNA. Zodra ze de fysieke bocht/vervorming (distortion) tegenkomen, blijft UvrB op de schade zitten en gaat UvrA weg (kost ATP).
Stap b & c (Knippen): UvrB rekruteert het 'schaareiwit' UvrC. UvrC maakt aan weerszijden van de beschadiging een knip in de DNA-streng (nick). Het knipt dus een heel herstel-segment van zo'n 12 nucleotiden los.
Stap d (Verwijderen en Opbouwen): Het enzym DNA helicase (UvrD) trekt het losgeknipte, beschadigde stuk DNA er tussenuit. Nu is er een flink gat in één streng. DNA polymerase I vult dit grote gat weer netjes op met gezonde nucleotiden aan de hand van de onbeschadigde tegenstreng, en DNA ligase lijmt het vast.
Regulatie van genexpressie van een bacterie
→ 2 soorten genen:
Constitutive promoter (Huishoudgenen): Sommige genen staan onder controle van een constitutieve promotor. Dit betekent dat ze een constant niveau van expressie hebben (constant level of expression). Dit zijn de genen die de bacterie altijd nodig heeft om te ademen en te leven.
Gereguleerde genen: Virulentiegenen vallen hieronder. Het is energetisch ongunstig (energetically unfavourable) om deze altijd aan te hebben staan. De bacterie zet ze pas aan wanneer hij gevaren of kansen herkent:
Veranderingen in de omgeving (aanwezigheid van nutriënten of toxische stoffen).
De afweer van de gastheer (zodra het immuunsysteem aanvalt).
Competitie met de normale bewoners (commensale flora) in de darm of keel.
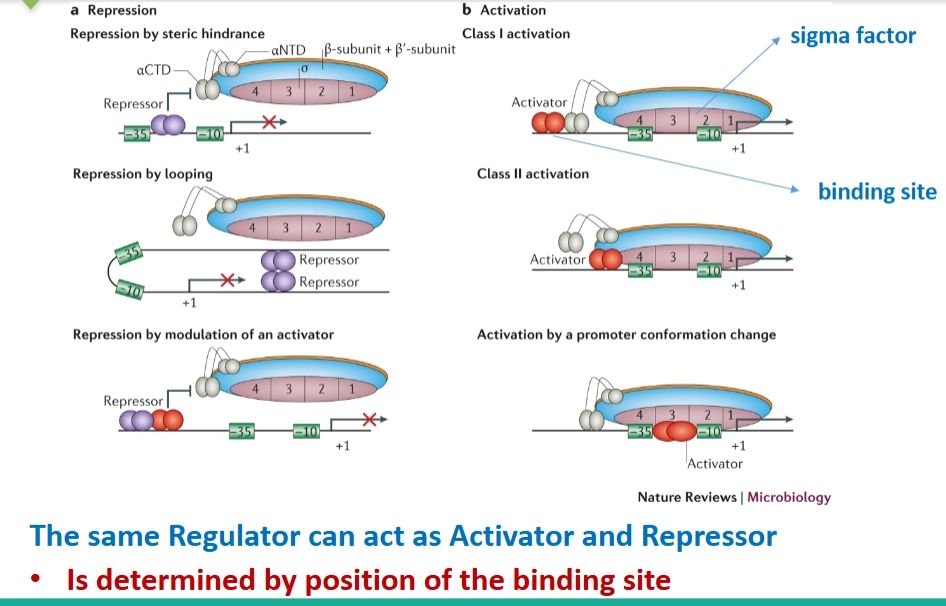
Regulatoren van bacteriën kunnen → activation & repression
→ laat zien hoe eiwitten (regulatoren) fysiek op het DNA binden om RNA-polymerase (geholpen door de sigma factor) tegen te houden of juist te helpen:
Repression (Onderdrukking/UIT-schakelaars):
Steric hindrance: Een repressor gaat fysiek boven op de promotor zitten. De RNA-polymerase kan er simpelweg niet langs (blokkade).
Looping: Twee repressors binden aan het DNA en trekken het DNA in een lus, waardoor de promotor onbereikbaar wordt.
Activation (Activering/AAN-schakelaars):
Class I & II activation: Een activator bindt vlak voor de promotor en trekt de RNA-polymerase fysiek aan om te helpen starten.
Cruciale opmerking: The same Regulator can act as Activator and Repressor. Eén en hetzelfde eiwit kan voor gen A een activator zijn, en voor gen B een repressor. Dit wordt puur bepaald door de positie van de bindingssite ten opzichte van de promotor!
Genregulatie op basis van omgeving
→ hoe weet de regulator wat er buiten de cel gebeurt? Kan op 2 manieren:
Cytoplasmic sensing (Binnenin de cel meten) Kleine metabole moleculen (zoals suikers of aminozuren) diffunderen de bacteriecel in. Ze binden direct aan een transcriptieregulator in het cytoplasma. Dit zorgt voor een vormverandering (conformational change), waardoor de regulator het DNA loslaat of juist vastgrijpt.
Sensing outside of cell: Two-component signal transduction (Buiten de cel meten) Wanneer een signaalstof niet door het membraan heen kan, gebruikt de bacterie een twee-componenten-systeem. De onderste dia laat deze prachtige cascade stap voor stap zien:
Component 1: De Sensor kinase Dit eiwit zit ingebed in het cytoplasmatische membraan. De buitenkant vangt een environmental signal op (bijvoorbeeld een verandering in pH, temperatuur, of een specifiek immuun-eiwit van de gastheer).
De actie: Zodra het signaal bindt, activeert de kinase zichzelf aan de binnenkant van de cel met behulp van ATP. Het plakt een fosfaatgroep op zichzelf (autofosforylering).
Component 2: De Response regulator Het fosfaatje (P) wordt vervolgens overgedragen op een los eiwit in het cytoplasma: de response regulator.
De actie: Door deze fosforylering verandert de response regulator van vorm en wordt hij actief. Hij reist direct naar het DNA toe en bindt aan de operator vlak voor de structural genes (de virulentiegenen).
Het effect: In het getekende voorbeeld zie je dat de actieve response regulator de weg blokkeert voor de RNA-polymerase (transcription blocked). Dit specifieke systeem staat dus op UIT zodra er een signaal is. (Bij andere systemen werkt het precies andersom en activeert het juist de transcriptie).
De Reset-knop (Phosphatase activity): Als het gevaar buiten de cel geweken is, haalt een ingebouwde enzymfunctie (fosfatase) het fosfaatje weer van de regulator af, zodat het systeem keurig terugkeert naar de ruststand.
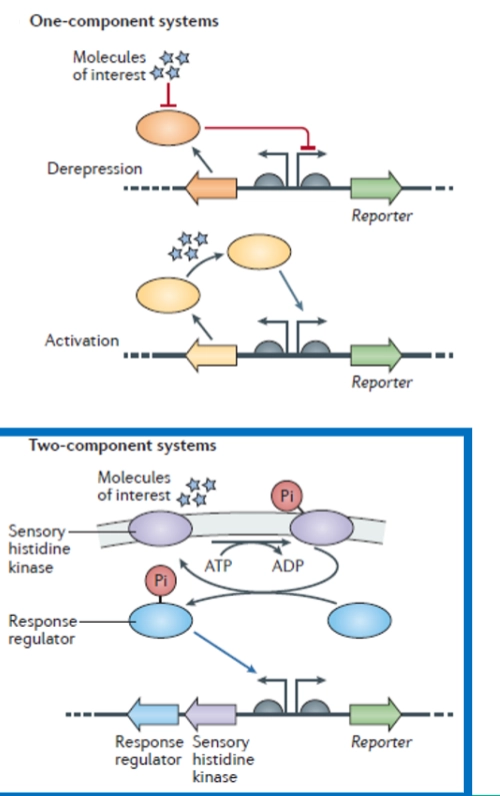
Voorbeelden van twee-componenten-systemen:
PhoP/Q systeem in myobacteria
De werking: PhoQ is de sensor in het membraan. Hij voelt een lage pH of een lage concentratie magnesium. Dit zijn de exacte kenmerken van een fagosoom/lysosoom. Zodra PhoQ dit voelt, fosforyleert hij zichzelf en activeert hij de response regulator PhoP via de bekende domino-cascade.
De kracht van activatie: Gekloonde PhoP bindt aan het DNA vlak voor de doelwitgenen (target genes). De afstand tussen de PhoP-bindingssite en de start van het gen (de promotor) bepaalt hoe sterk het gen geactiveerd wordt.
→ Mutaties bepalen virulentie: Niet elke Mycobacterie is even gevaarlijk. Sommige stammen hebben mutaties:
In de promotor van phoP: Er wordt minder PhoP geproduceerd (less PhoP produced).
In de ORF (de code) van phoP/phoQ: Het DNA-bindingsdomein of het fosforyleringsdomein raakt beschadigd. Dit leidt tot less activation van de virulentiegenen.
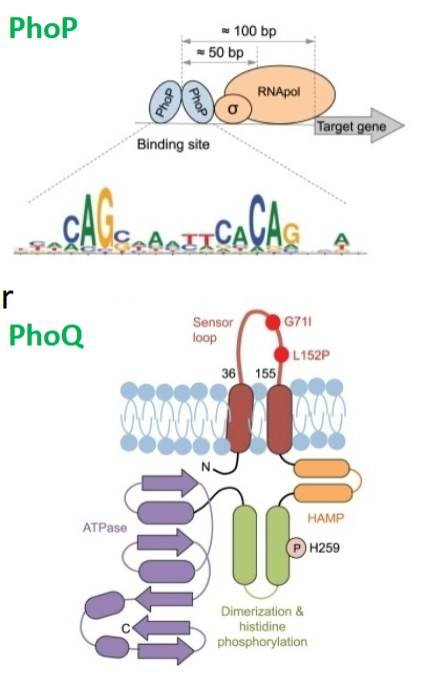
PhoP/Q regulon in myobacteria
Regulon
= het totale pakket aan genen dat door 1 regulator wordt aangestuurd Verschillen in dit regulon bepalen hoe ziek je van de bacterie wordt
Mycobacterium tuberculosis (Wildtype - Links): Heeft een perfect werkend PhoP/Q-systeem. Zodra hij in het fagosoom zit, activeert PhoP twee cruciale zaken:
De productie van speciale vetten (lipids of outer membrane) die de bacterie beschermen.
Het Esx-1 secretiesysteem. Dit is een specifieke moleculaire pomp waarmee de bacterie gaten boort in het fagosoom om te ontsnappen naar het cytoplasma (used to escape phagosme).
Mycobacterium bovis / BCG-vaccin (Midden en Rechts): * M. bovis (verwekker van runder-tbc) heeft een natuurlijke mutatie in zijn sensor (PhoR/Q mutant), waardoor het systeem veel minder goed werkt (midden).
De mens gemaakte MTBVAC vaccin-stam (rechts) is een tbc-bacterie waarin wetenschappers het phoPgen bewust kapot hebben gemaakt (PhoP mutant). Omdat PhoP ontbreekt, kan de bacterie het Esx-1 secretiesysteem niet aanzetten (het rode kruis). Hij kan niet uit het fagosoom ontsnappen, richt geen schade aan, maar traint wel veilig jouw immuunsysteem!
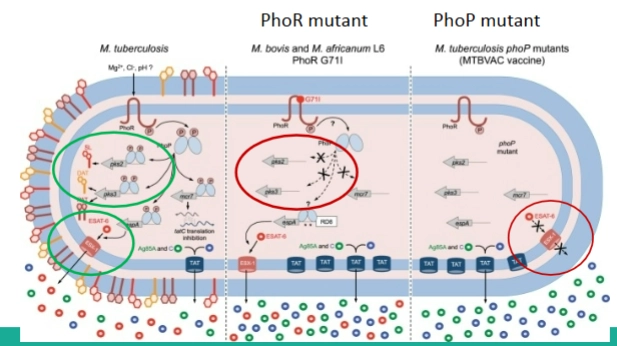
Voorbeelden van twee-componenten systeem:
BvgA/S systeem in Bordetella pertussis (kinkhoest)
→ gebruikt een vergelijkbaar systeem (BvgS is de sensor kinase, BvgA is de response regulator), maar reageert op een andere prikkel: lichaamstemperatuur.
→ temperatuur als aan-schakelaar
Buiten de gastheer (<37): Het systeem staat op Inactive (links in het diagram). De bacterie maakt geen dure gifstoffen aan, want hij zit toch nog niet in een mens.
Binnen de gastheer (37): Zodra de bacterie wordt ingeademd en de menselijke lichaamstemperatuur voelt, klapt de sensor BvgS om naar Active. Er loopt een fosfaatcascade via BvgS naar BvgA.
De fasen van infectie (De grafiek rechtsonder): De bacterie activeert zijn genen niet allemaal tegelijk, maar in opeenvolgende golven (klassen):
Class 4 (Buiten de gastheer): Genen voor beweeglijkheid staan aan; genen voor virulentie staan uit.
Class 3 & 2 (Transmission & colonisation): Zodra de bacterie binnenkomt, maakt hij eerst heel snel adhesines (plakeiwitten) aan om zich stevig vast te zetten in de keel.
Class 1 (Infection established): Als de bacterie eenmaal stevig vastzit, gaan de zware wapens (toxines) pas aan om het weefsel aan te vallen en de gastheer te laten hoesten.
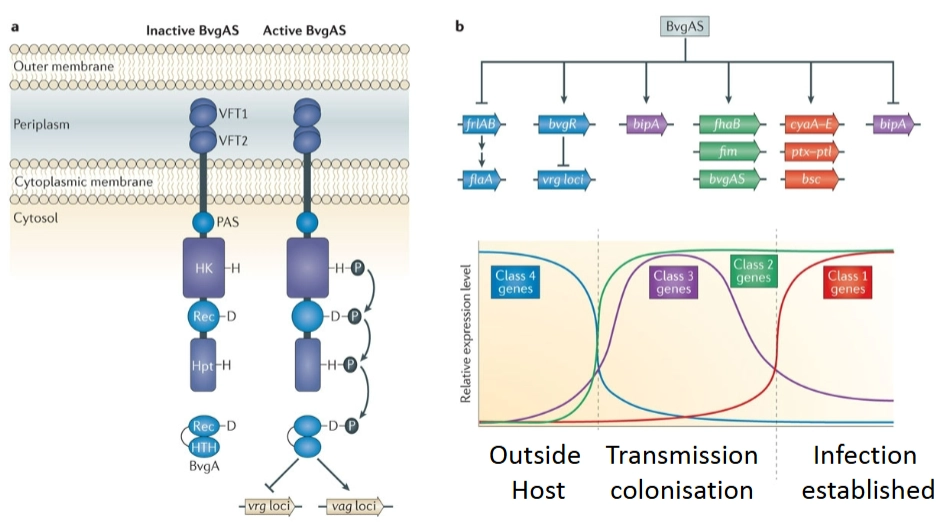
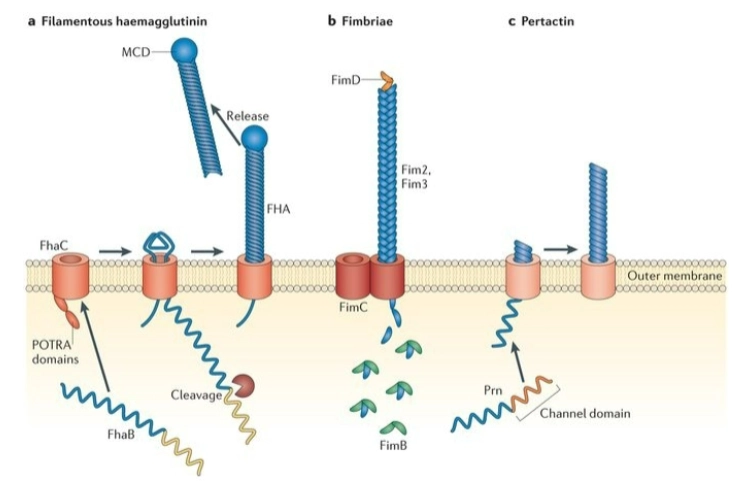
BvgA/S-systeem: wat bouwt het tijdens zo’n Class 1/2 infectie op het oppervlak van de bacterie?
a. Filamentous haemagglutinin (FHA): Een gigantisch, staafvormig plakeiwit dat door het membraan heen naar buiten wordt geduwd (via FhaC) om de bacterie muurvast te ankeren aan de trilharen in je luchtwegen.
b. Fimbriae: Lange, harige uitsteeksels (gemaakt van FimB/C/D) die ook helpen bij de hechting.
c. Pertactin (Prn): Nóg een belangrijk adhesie-eiwit aan de buitenkant.
Pertussis Toxin: Naast deze plakeiwitten activeert BvgA de productie van het beruchte kinkhoest-toxine. Dit gifstofje verlamt de trilharen in de luchtpijp en vernietigt cellen, wat leidt tot die typische, gierende en uitputtende hoestbuien (causes the characteristic coughs).
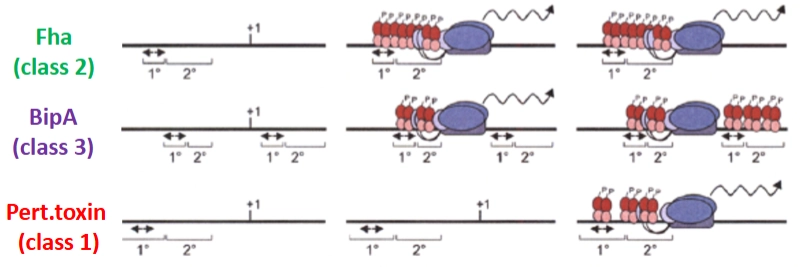
Hoe zorgt 1 en dezelfde actieve regulator (BvgA-P) ervoor dat sommige genen heel snel (Class 2) en andere genen pas veel later (Class 1) aangaan?
= bindingsaffiniteit (hoe goed het eiwit plakt) & de positie van de primaire en secundaire bindingsplaatsen op het DNA.
primaire vs. secundaire bindingsplaatsen
De actieve, gefosforyleerde regulator (BvgA-P, de rode bolletjes op de dia) werkt met een tweetrapsraket:
Primair: Heeft een hoge bindingsaffiniteit. Dit betekent dat zelfs als er nog maar heel weinig BvgA-P in de cel aanwezig is, het direct muurvast aan deze plek plakt. Binding aan deze eerste plek helpt vervolgens om andere regulatoren naar de minder aantrekkelijke secundaire site te trekken.
BvgA als Activator of Repressor: Net zoals we bij de algemene transcriptiedia zagen, bepaalt de positie van deze bindingssites ten opzichte van de promotor (+1) of de RNA-polymerase (de blauwe ovaal) of het gen aan- of uitgezet wordt.
mate van bindingsaffiniteit per eiwit
Fha (Class 2 genen - De vroege plakeiwitten)
Het DNA: De primaire en secundaire bindingssites liggen vóór de promotor (+1).
Het mechanisme: Omdat de primaire site een superhoge affiniteit heeft, binden de eerste paar rode bolletjes direct (middelste plaatje). Dit rekruteert meteen de RNA-polymerase. Fha gaat dus extreem snel AAN zodra de temperatuur 37 wordt. Het fungeert als een pure activator.
BipA (Class 3 genen - De tussenfase)
Het DNA: Heeft bindingssites vóór de promotor én achter de promotor.
Het mechanisme: Bij een lage/medium concentratie BvgA-P (middelste plaatje) activeert het de transcriptie vanaf de linkerkant. Maar zodra er heel veel BvgA-P is (rechter plaatje), bezet het ook de rechter bindingssites. De RNA-polymerase wordt nu ingesloten en geblokkeerd. BvgA-P switcht hier dus van activator naar repressor naarmate de infectie vordert.
Pertussis Toxin (Class 1 genen - De late zware wapens)
Het DNA: De primaire site heeft hier een lagere affiniteit omdat de DNA-volgorde net iets anders is (different sequence).
Het mechanisme: Als de bacterie net binnenkomt, is er nog te weinig BvgA-P om aan deze promotor te binden; het gen blijft volledig UIT (middelste plaatje). Pas wanneer de bacterie helemaal gesetteld is en de fosfaatcascade op volle toeren draait, is er zoveel BvgA-P in de cel dat het dwingend naar de primaire en secundaire site van het toxine-gen wordt geduwd (rechter plaatje). Pas dán start de productie van het kinkhoest-toxine.
Horizontal Gene Transfer and Genome Stability
= de manier waarop bacteriën genen met elkaar uitwisselen zonder zich te hoeven delen.
→ evolutionaire processen op DNA niveau
Vertical transmission
= celdeling → replicaties en mutatiesHorizontal gen transfer
= DNA overdracht tussen organismen
→ gevolgd door verticale transmissie
Horizontale genoverdracht: genetische elementen van bacteriën die van belang zijn:
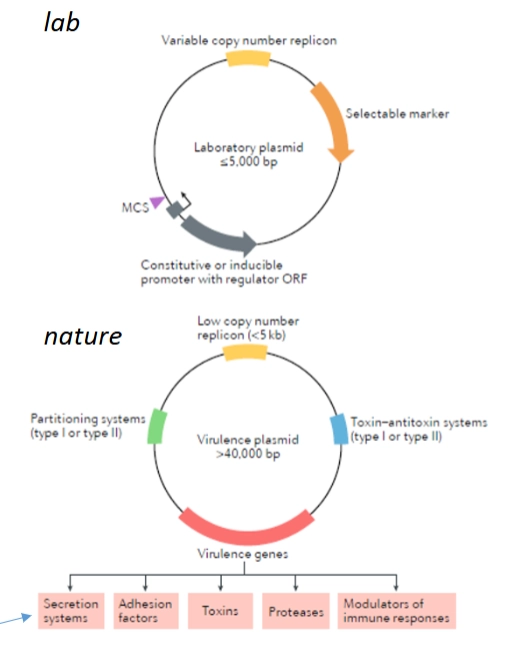
Plasmiden
= kleine, cirkelvormige stukjes DNA die losstaan van het grote bacteriële chromosoom.
Kenmerken:
Lab vs. Natuur: In het laboratorium gebruiken we kleine, kunstmatige plasmiden met een variabele copy number en een selectable marker (zoals antibioticaresistentie). In de natuur zijn plasmiden van pathogenen vaak gigantisch (>40.000 basisparen).
Natuurlijke snufjes: Natuurlijke plasmiden bevatten slimme systemen, zoals Toxin-antitoxin systemen (die ervoor zorgen dat de bacterie het plasmid niet verliest; als hij het plasmid verliest, sterft hij door het achtergebleven toxine) en partitioning systems om het plasmid netjes te verdelen tijdens celdeling.
Overdracht: Ze worden overgedragen via transformatie (los DNA opnemen, zie hieronder) of conjugatie (bacterieseks via een pilus).
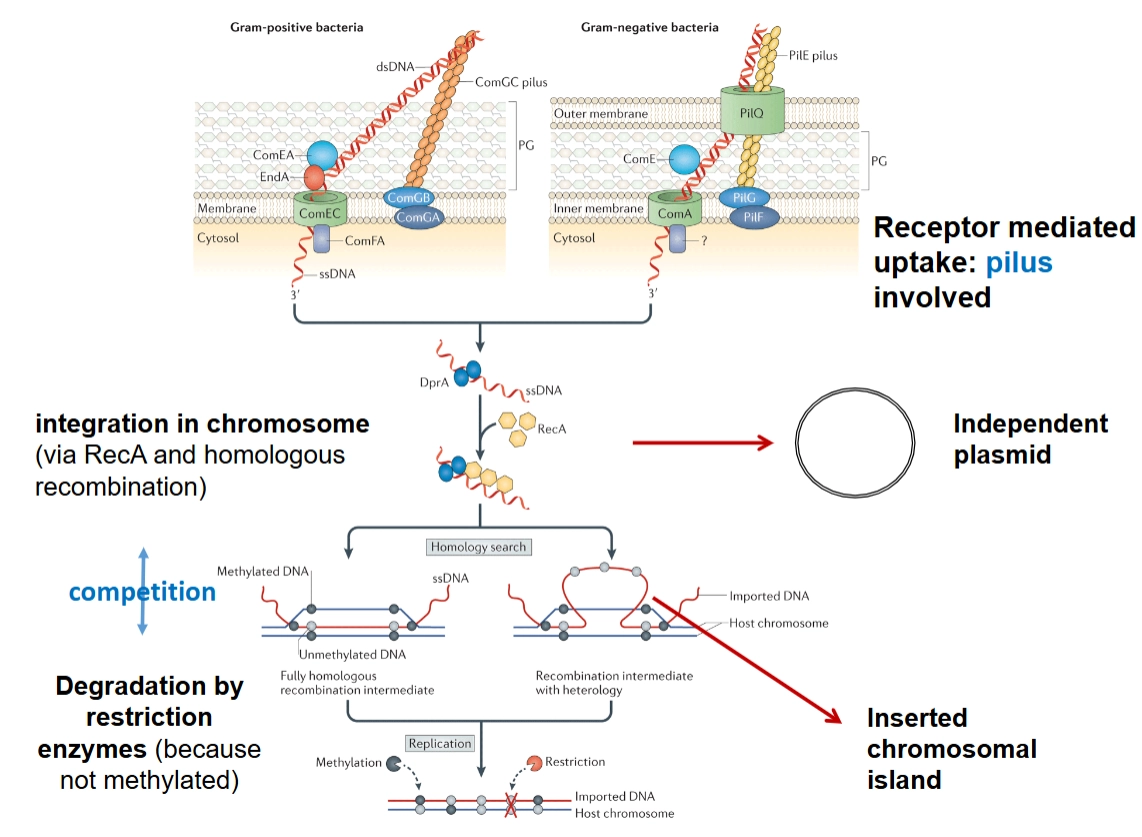
Opname proces plasmides
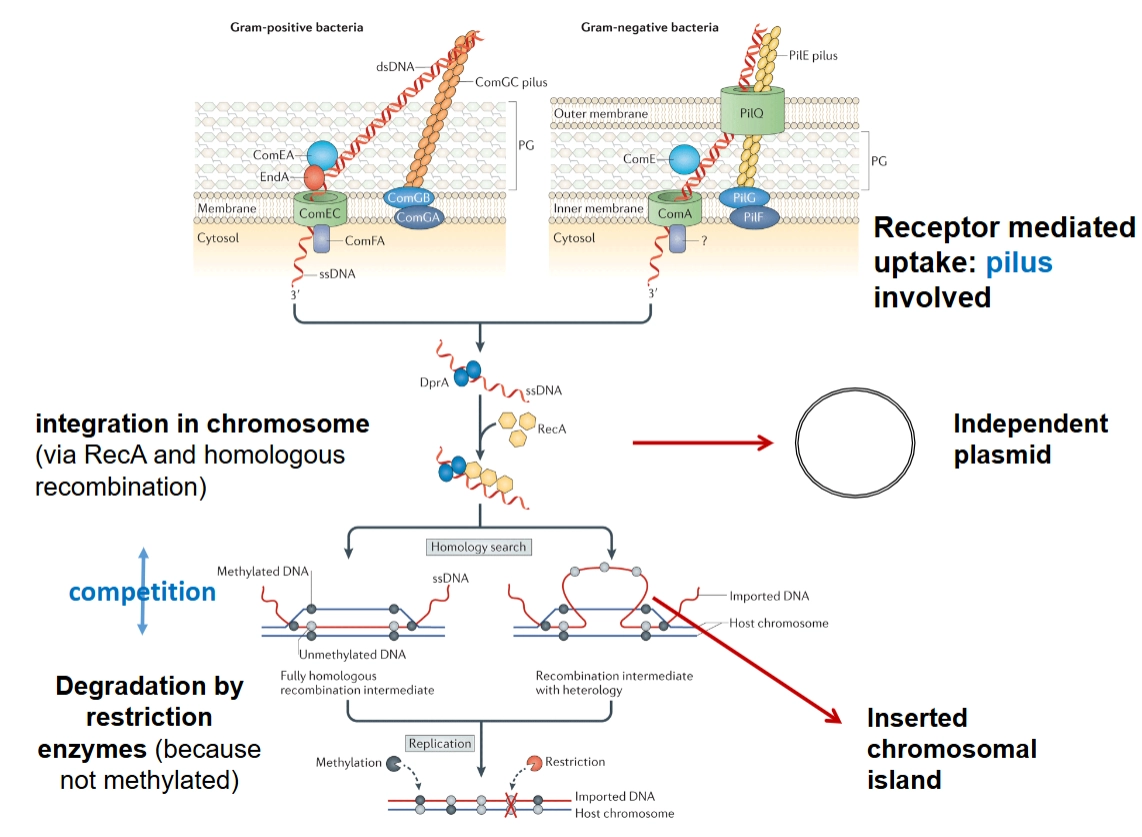
Bacteriën die van nature vreemd DNA kunnen opnemen, noemen we naturally competent. Het mechanisme verschilt per bacterietype:
→ Gram-positief vs. Gram-negatief: Beide gebruiken een samentrekkende haar (pilus) aan de buitenkant om het losse, dubbelstrengs DNA (dsDNA) te grijpen (receptor mediated uptake).
Naar binnen halen: Het DNA wordt door de membraanpoortjes geleid (zoals ComEC of ComA) en omgezet in enkelstrengs DNA (ssDNA).
De bestemming: Eenmaal binnen kan dit ssDNA zich weer omvormen tot een stabiele, cirkelvormige structuur die onafhankelijk blijft bestaan: het independent plasmid.
Ontstaan van genetische eilanden (& pathogeniciteit eiland)
→ Wat gebeurt er als dat opgenomen DNA niet als een los plasmid blijft rondzwemmen? Dan kan het integreren in het hoofdchromosoom van de bacterie.
→ hoe ontstaat zo’n eiland?
Homologe recombinatie: Het binnengekomen enkelstrengs DNA wordt beschermd door het eiwit RecA (integration via RecA and homologous recombination). RecA helpt om te zoeken naar stukken DNA op het bacteriechromosoom die ergens lijken op het nieuwe DNA (homology search). Het nieuwe stuk DNA wordt er tussen geplakt: een inserted chromosomal island (een genetisch eiland) is geboren.
Pathogenitietseilanden (PAIs): Als dit eiland vol zit met virulentiegenen (zoals codes voor secretion systems, adhesion factors, toxins, proteases), spreken we van een Pathogenicity Island.
De verdediging van de bacterie (Competitie): De gastheerbacterie slikt niet zomaar alles. Hij heeft restrictie-enzymen die vreemd DNA direct in stukken hakken (degradation). Hoe herkent de bacterie zijn eigen DNA? Eigen DNA is gecodeerd met methylgroepen (methylated). Het binnengekomen DNA mist deze bescherming (unmethylated) en wordt vaak vernietigd, tenzij het heel snel integreert via recombinatie.
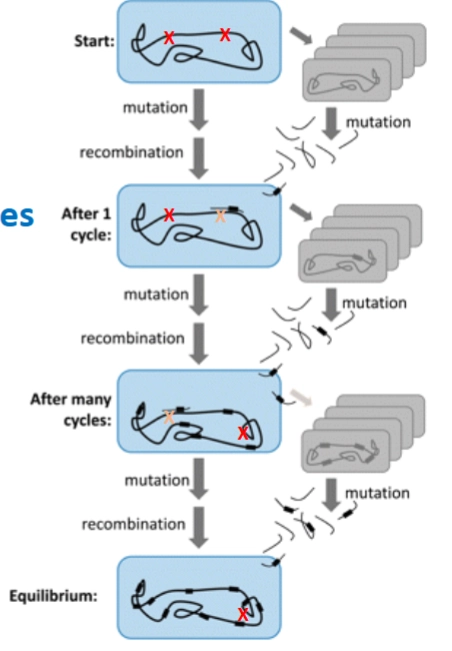
Transformatie van bacteriën → ook om eigen fouten te herstellen
(transformation for genome maintenance)
→ Neisseria meningitidis gebruikt DNA uptake sequences: Neisseria is extreem kieskeurig. Hij neemt niet zomaar elk DNA op. Hij herkent een heel specifieke code van 10 letters (GCCGTCTGAA) op het losse DNA. Deze code zit vast aan de household genes van zijn eigen soortgenoten. Zo neemt hij alleen nuttig DNA van overleden 'familieleden' op om mutaties of defecten in zijn eigen genoom te repareren via recombinatie.
Bacteriofaag (& transductie)
= virussen die specifiek bacteriën infecteren
→ wanneer zij per ongeluk bacterie-DNA meenemen van de ene naar andere bacterie
= transductie
Bacteriofaag: 2 levenscycli
→ Een faag landt op de bacterie, boort gaatjes en spuit zijn virale DNA naar binnen (injection). Vanaf hier zijn er twee routes:
Lytic pathway (De sloper): Het virale DNA kaapt de bacterie, dwingt de productie van duizenden nieuwe fagen af, en laat de bacterie exploderen (lysis).
Lysogenic pathway (De slaper): De zogenaamde temperate virussen slopen de bacterie niet direct. Het faag-DNA integreert geruisloos in het bacteriële chromosoom (phage genome integrated in chromosome). De bacterie leeft gewoon door, deelt zich, en kopieert het geïntegreerde faag-DNA (nu een profaag of dormant virus genoemd) trouw mee naar alle nakomelingen.
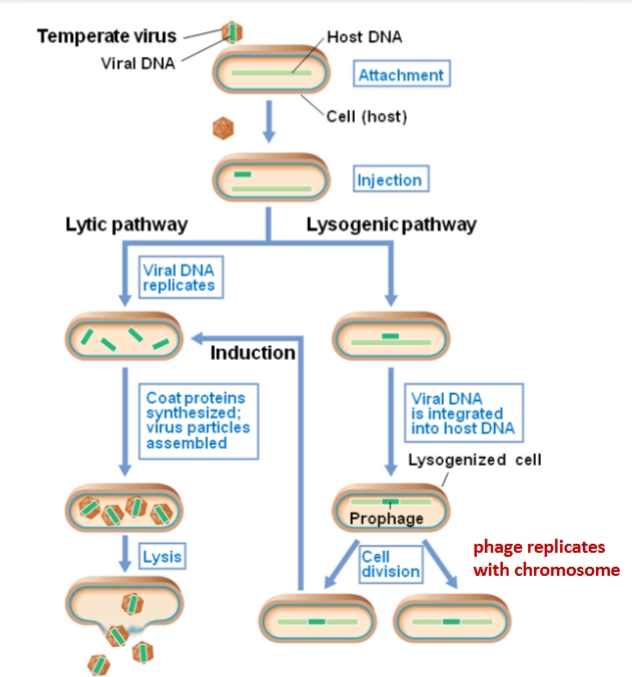
Lysogenic pathway: lysogene conversie
→ dit slapende virus-DNA is niet zomaar junk-DNA; het kan functionele genen bevatten waar de bacterie enorm veel voordeel van heeft. Dit fenomeen noemen we lysogene conversie: de bacterie krijgt ineens superkrachten dankzij het virus.
Voorbeelden van fagentoxines
Het Cholera-toxine (veroorzaakt diarree bij Vibrio cholerae).
Vibrio cholerae zwemt rond in het water. Er zijn ontzettend veel verschillende stammen (serogroepen). Maar alleen de stammen die én een kapsel hebben (zoals O1 en O139) én toxigeen zijn (het choleratoxine-gen van de faag bezitten), kunnen de mens infecteren en de ziekte cholera veroorzaken.
Waarom stimuleert het faag-toxine diarree? Het is een evolutionaire verspreidingsstrategie. Door het toxine ontstaat er gigantische diarree. Hierdoor spoelt de bacterie massaal uit de gastheer (bacteria escape host) terug het water in, samen met heel veel nutriënten uit de darm.
Horizontale genoverdracht (HGT): Wanneer zo'n slapende profaag in de darm ineens weer actief en lytisch wordt, verlaat hij de bacterie, neemt het toxine-gen mee en infecteert andere Vibrio's in de buurt die het toxine nog niet hadden. Zo verspreidt de virulentie zich razendsnel door de bacteriepopulatie.
Het Botuline-toxine (veroorzaakt verlamming bij Clostridium botulinum).
Het Difterie-toxine (DT toxin, zie de rode cirkel op de genenkaart van Dia 3).