BCH311 L11 Ribosome, protein synthesis
1/30
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
31 Terms
similarities between eukaryotic and ribosomal compositions
general function:
full complex: translation
small subunit: mRNA binding and codon/anticodon pairing
large subunit: peptide bond formation
general composition:
full complex: 2/3 rRNA folded into duplex regions, 1/3 proteins connect rRNAs
small and large subunit: ONLY rRNA at subunit interface (no protein)
differences between eukaryotic and ribosomal compositions
eukaryotic ribosome composition: full complex 80S
small subunit: 40S: 30 proteins, 18S rRNA
large subunit: 60S: 40 proteins, 5S rRNA, 28S rRNA, 5.8S rRNA
prokaryotic ribosome composition: full complex 70S
small subunit: 30S: 21 proteins (S1-21), 16S rRNA
large subunit: 50S: 31 proteins (L1-31), 5S rRNA, 23S rRNA
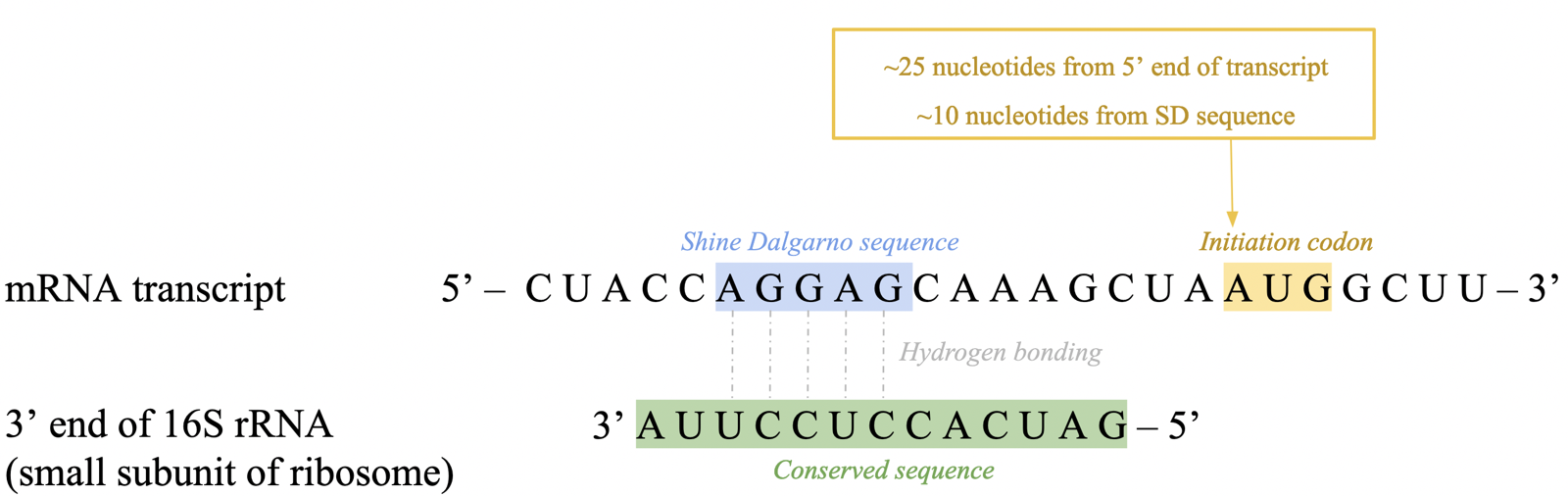
translation initiation: sequence specificity (prok)
translation begins before transcription is complete: co-transcriptional translation
first codon found ~25 nt downstream from 5’ end
ribosome binds ~10 nt upstream at purine-rich Shine Dalgarno (SD) sequence
recognized by 3’ end of 16S rRNA via bp, helping correctly position initiation codon (AUG) into ribosome’s P site
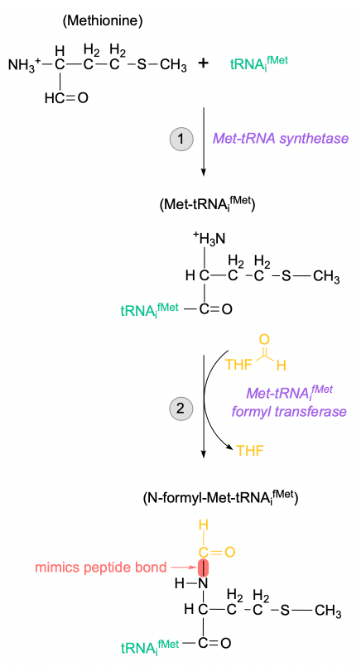
translation initiation: initiator tRNA + charging (prok)
AUG start codon after the SD signals for incorporation of formyl methionine (fMet) residue
carried exclusively by the initiator tRNA (tRNAifMet), differing from internal methionines
charging occurs in 2 distinct steps:
first, tRNAifMet is charged with unmodified methionine by the regular Met-tRNA synthetase
Met-tRNAifMet formyl transferase will attach a formyl group to the N-terminal of the tRNA-bound Met
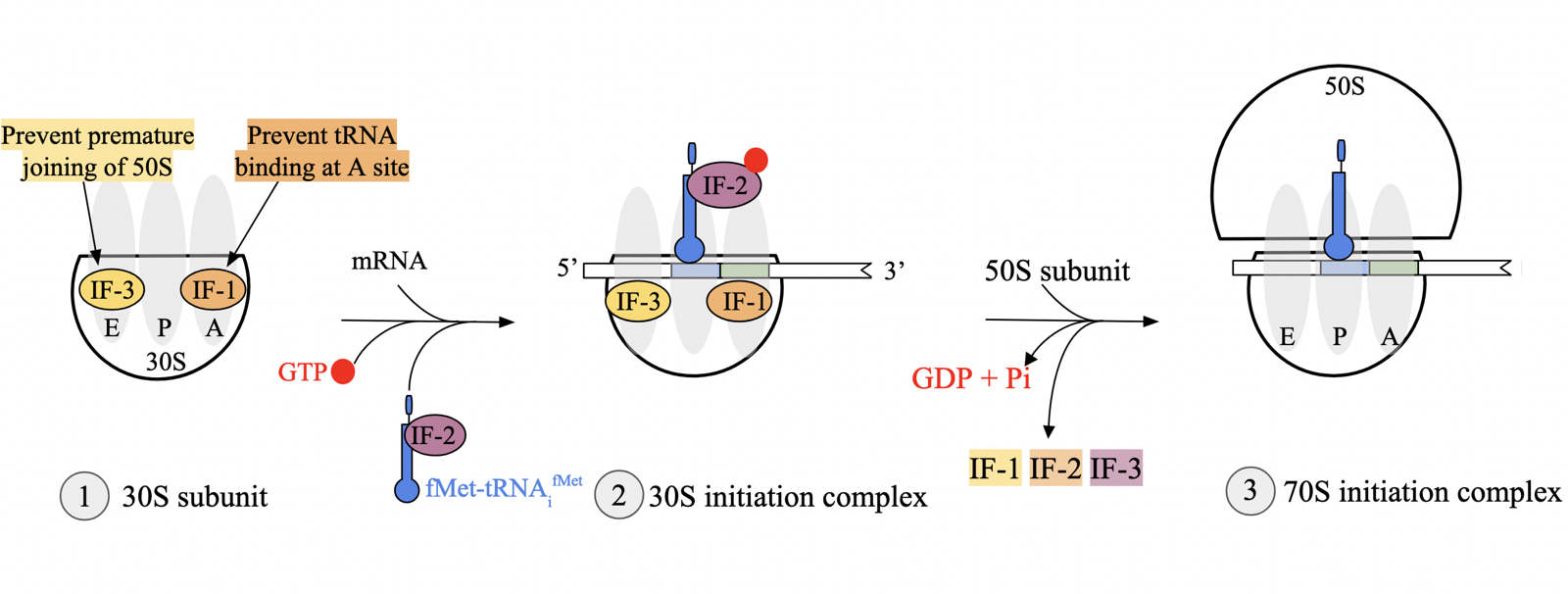
translation initiation: initiation factors (prok)
IF1, IF2, IF3: P-loop NTPases that undergo significant conformational changes upon binding and hydrolysis of NTPs
translation initiation via IFs occur in 3 key steps
IF1 and IF3 binding: IF1 binds near A site of 30S subunit, ensuring tRNAi binds P site. IF3 prevents premature binding of 50S, preventing formation of 70S complex without mRNA and tRNAi.
binding of mRNA and IF2 to form the 30S initation complex: 16S rRNA in 30S subunit H-bonds mRNA transcript for correct reading frame. IF2-fMet-tRNAifMet-GTP associates with 30S
transition to the 70S initiation complex: IF2 bound to 30S subunit undergoes GTP hydrolysis, releasing all IFs from complex. promotes binding of 50S large subunit, resulting in 70S initiation complex
IF2
initiation factor in prokaryotes
in complex with fMet-tRNAifMet and GTP, associates with 30S
when bound to 30S, undergoes GTP hydrolysis, releasing all IFs from complex.
promotes binding of 50S large subunit, resulting in 70S initiation complex
IF1
initiation factor in prokaryotes
binds near A site of 30S subunit, ensuring tRNAi binds P site
IF3
initiation factor in prokaryotes
prevents premature binding of 50S without mRNA and initiator tRNA
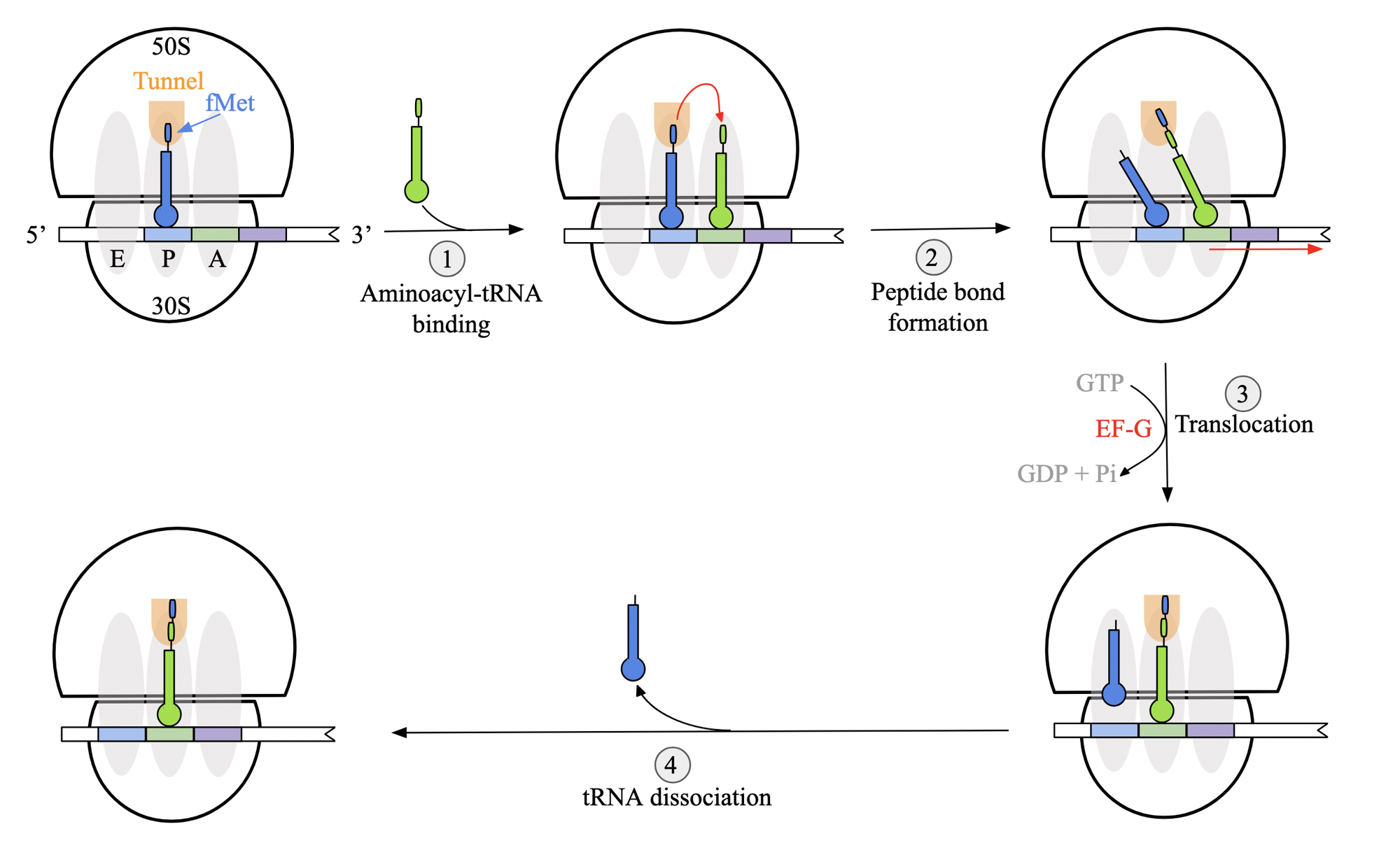
translation elongation stages: general mechanism (prok)
aminoacyl tRNA binding: binds at A site, thanks to fMet-tRNAifMet at P site.
peptide bond formation: takes place at P site of L subunit. chain is transferred from tRNA in P to tRNA in A. acceptor arms of tRNA undergo positional shift
anticodon regions initially remain in the same place
translocation: ribosome must shift by 3 nt towards mRNA 3’ end for next codon into A site. driven by GTP hydrolysis and facilitated by elongation factor G (EF-G)
after translocation, tRNA anticodons are properly aligned in respective site.
cycle: ribosome releases a deacylated tRNA molecule from E site and returns to initial state.
EF-Tu
elongation factor involved in aminoacyl tRNA binding
protects ester linkage between amino acid and tRNA molecule, ensuring aa remains attached to tRNA upon delivery
hydrolyzes GTP to GDP + Pi and releases tRNA molecule to ribosome when correctly paired with mRNA codon
must be regenerated by EF-T
EF-Tu does not interact with initiator tRNA, it is brought to ribosome by IF-2
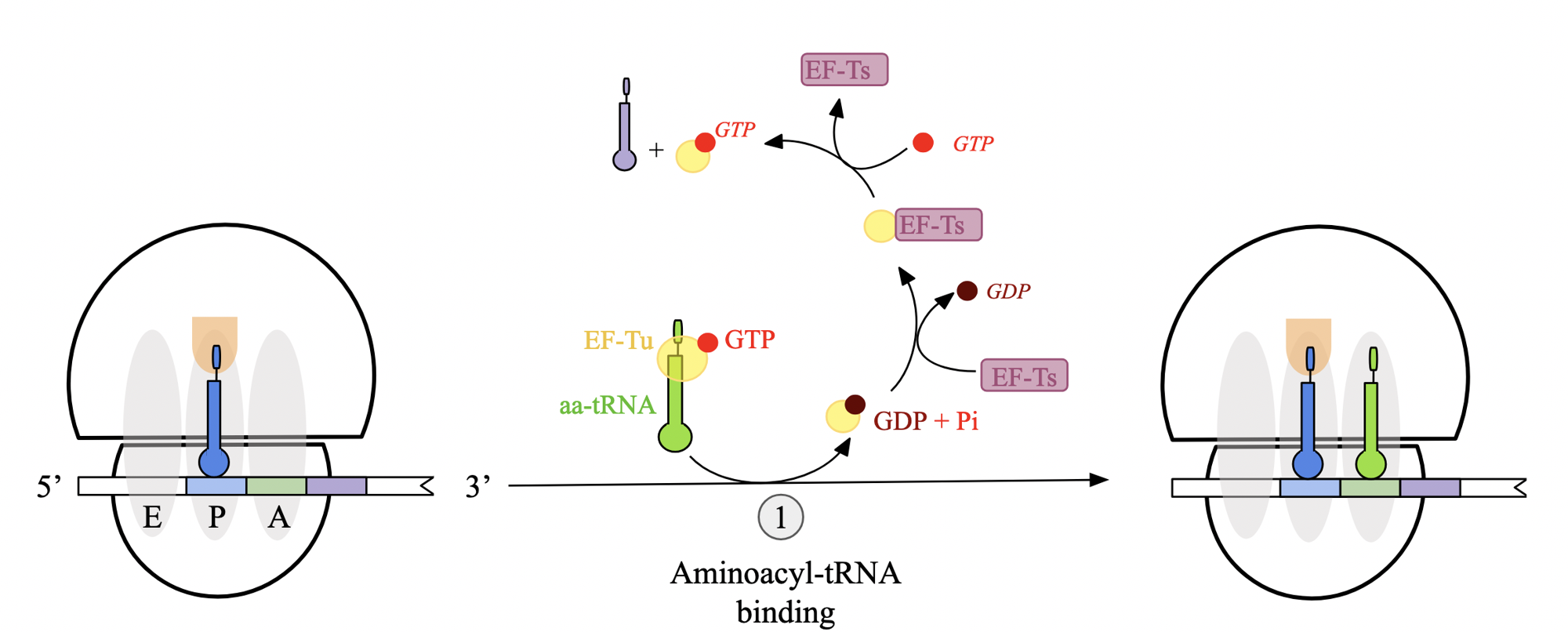
translation elongation stages: step 1 - aminoacyl tRNA binding (prok)
delivery of amino-acyl tRNA to A site of ribosome facilitated by GTP-bound elongation factor EF-Tu
EF-Tu protects ester linkage between aa and tRNA molecule, ensuring they are attached
when anticodon correctly pairs with mRNA codon at A site, EF-Tu hydrolyzes GTP to GDP + Pi and releases tRNA to the ribosome
triggers conformational change that allows tRNA to bind firmly to A site
EF-Tu is bound to GDP and must be regenerated by guanine exchange factor, EF-Ts
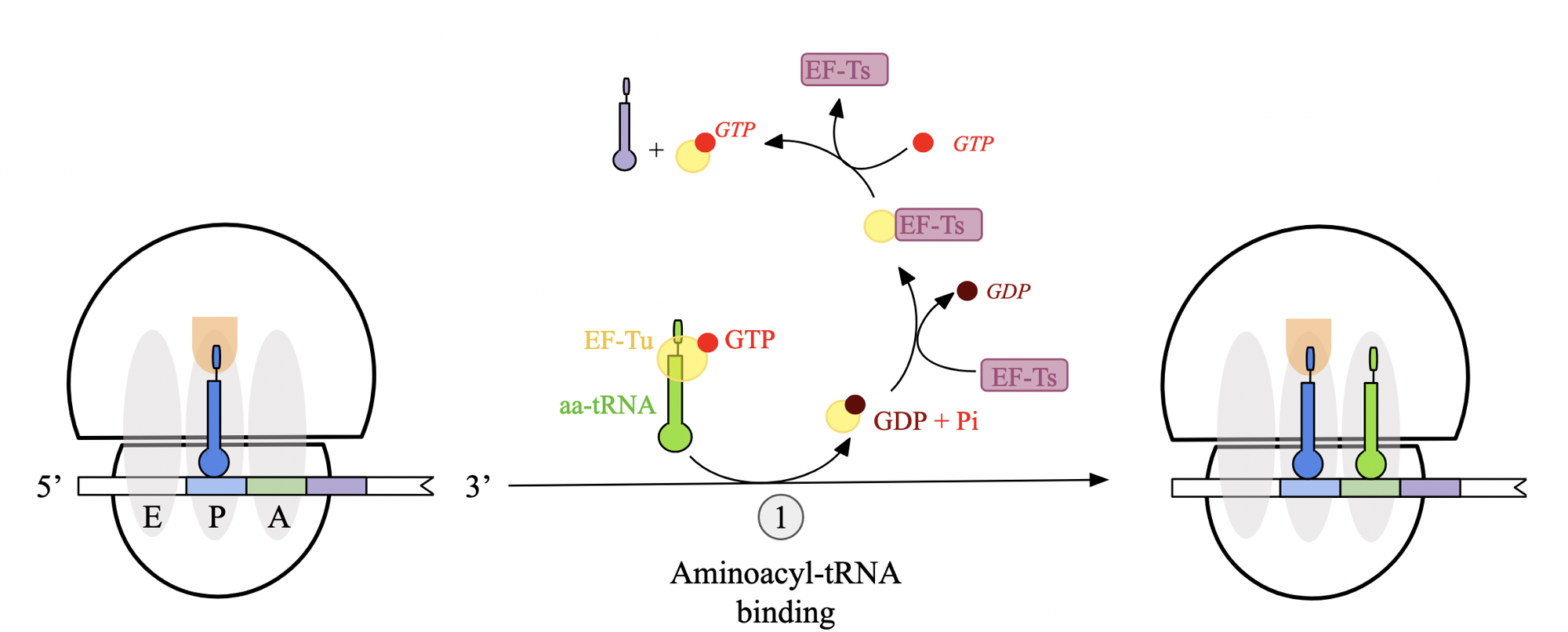
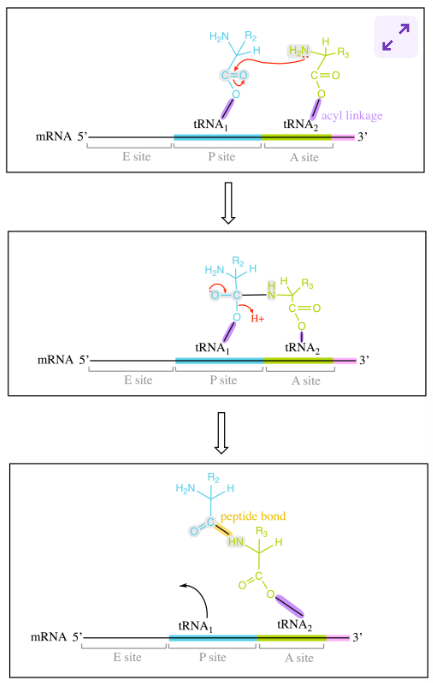
translation elongation stages: step 2 - peptide bond formation (prok)
once A and P site are occupied by aminoacyl-tRNA molecules, peptide bond formation takes place at P site of L subunit
amino group of aa at A attacks carbonyl carbon of aa at P.
formation of tetrahedral intermediate, polypeptide-chain and aa are linked as well as to tRNAs
undergoes bond rearrangement, leading to peptide bond formation and deacylated tRNA molecule
upon peptide bond formation, acceptor arms shift positions
acceptor arm of tRNA with growing pp chain → P site
deacylated tRNA accept arm → E site
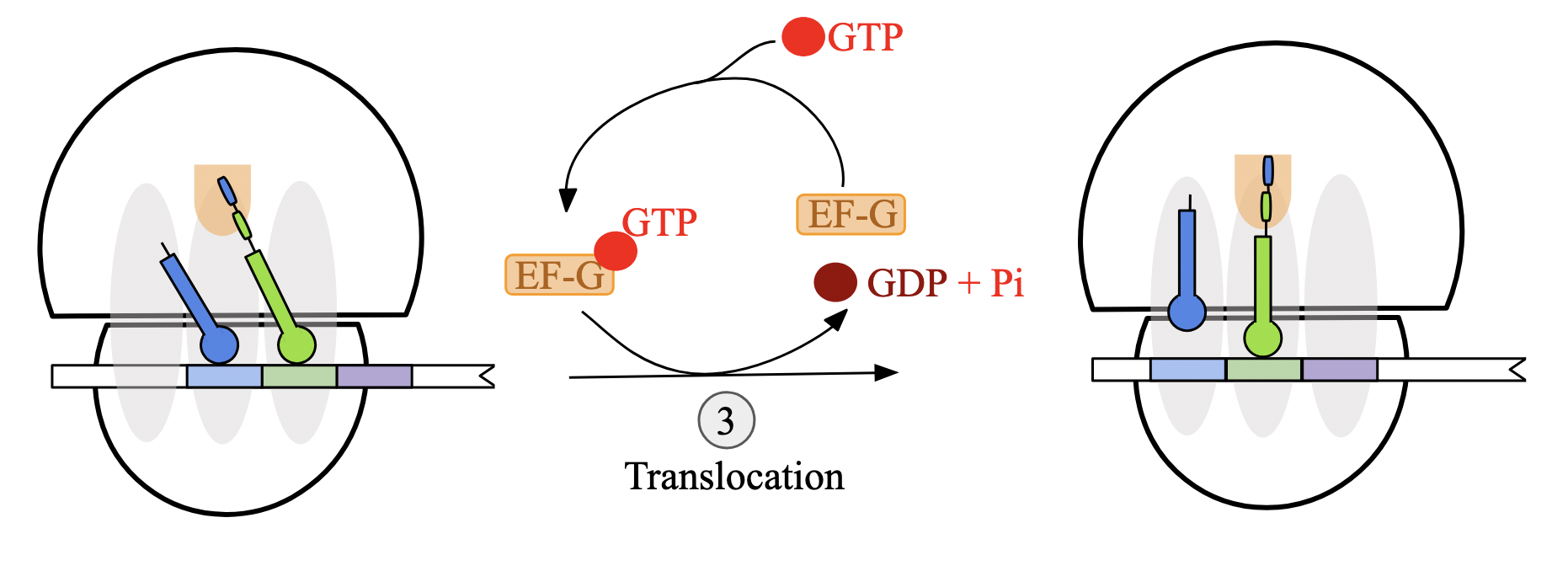
translation elongation stages: step 3 - translocation (prok)
GTP-bound elongation factor EF-G attaches to 50S ribosomal subunit near A site
EF-G hydrolyzes GTP, causing conformational change that simultaneously
transfers anticodon of tRNA at A site to the P site
moves the anticodon of the deacylated tRNA at the P site to the E site
ribosome translocates one codon towards 3’ end of mRNA transcript
EF-G
elongation factor involved in translocation
attaches to 50S ribosomal subunit near A site and hydrolyzes GTP, causing translocation-related conformational changes
structure mimics EF-Tu-bound amino-acyl tRNA complex, allowing to fit easily near A site
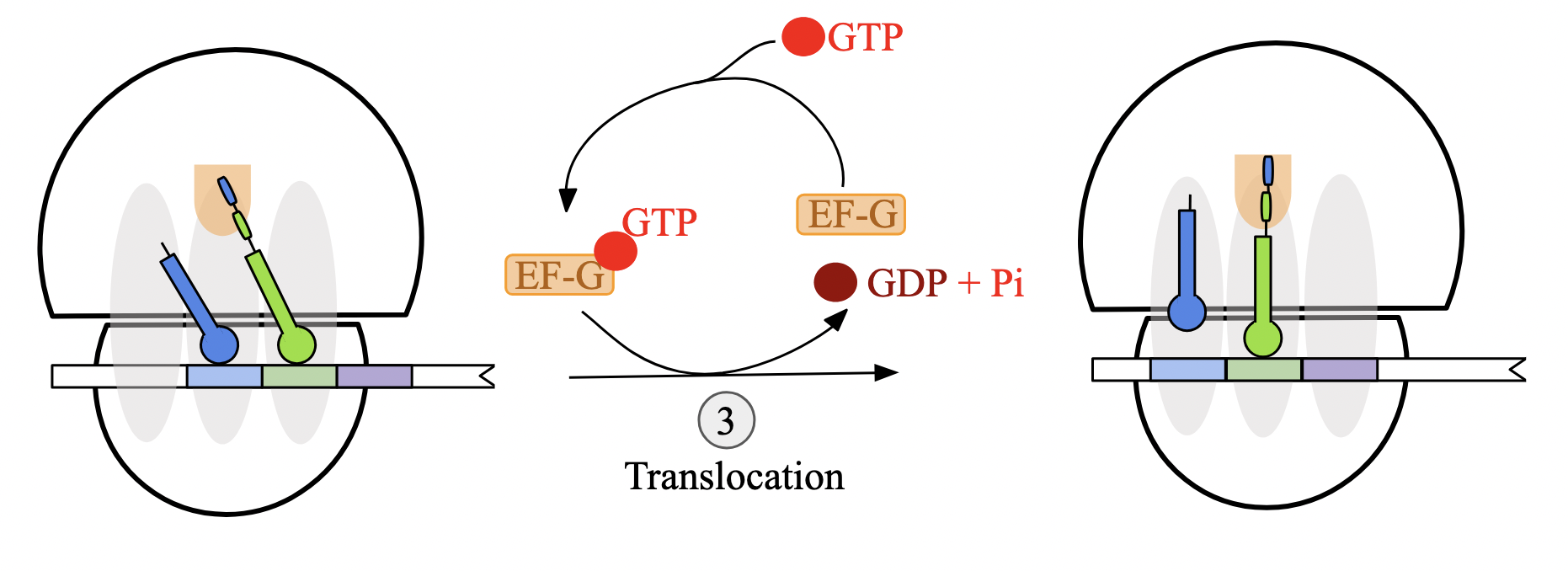
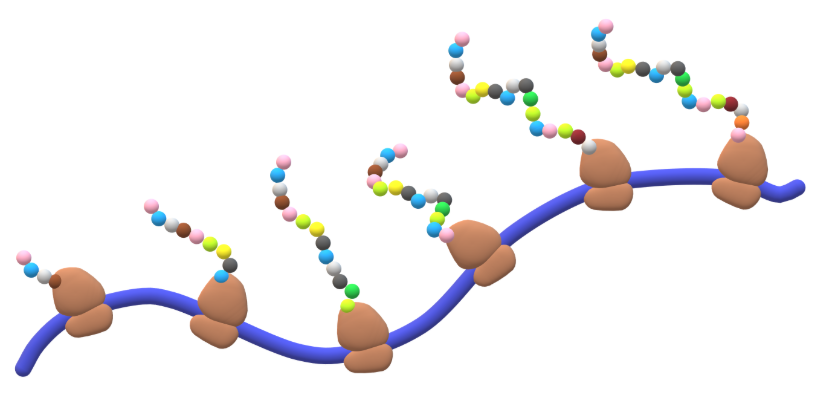
translation elongation stages: step 4 - cycle
multiple ribosomes can be cycling through the translation elongation process on the same mRNA transcript at the same time
groupings called polysomes
enhances translational efficiency for both prokaryotes and eukaryotes
can be visualized using electron microscopy
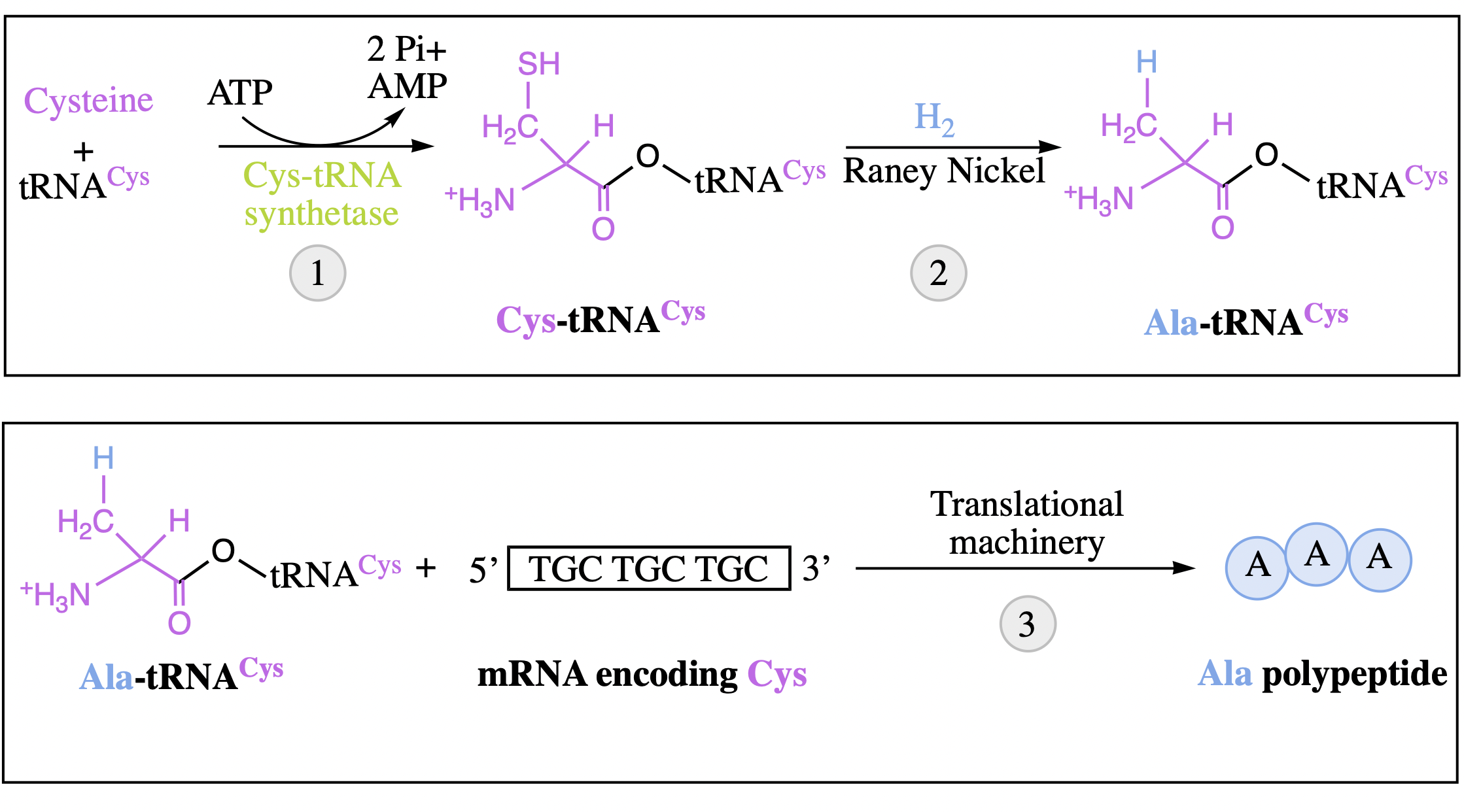
mRNA/tRNA pairing: mischarging tRNA molecules
base pairing interactions between mRNA and tRNA codons dictates which amino acid will be added to the growing polypeptide chain
specific amino acid plays no role in this process
if tRNA is mischarged, anticodon will still interact, and incorrect aa will be incorporated into the polypeptide chain
amino acids do not play a role in codon pairing: experiment
tRNACys correctly charged with Cysteine using cysteinyl-tRNA synthetase and ATP
the sulfur of the cysteine can then be converted into a H atom using Raney nickel, creating Ala-tRNACys, a mischarged tRNA
combining the Ala-tRNACys molecules with a synthetic mRNA template specific for cysteines + the necessary translational machinery produces a polypeptide chain composed of Ala-s
demonstrates that protein synthesis is independent of the amino acid charged to a particular tRNA
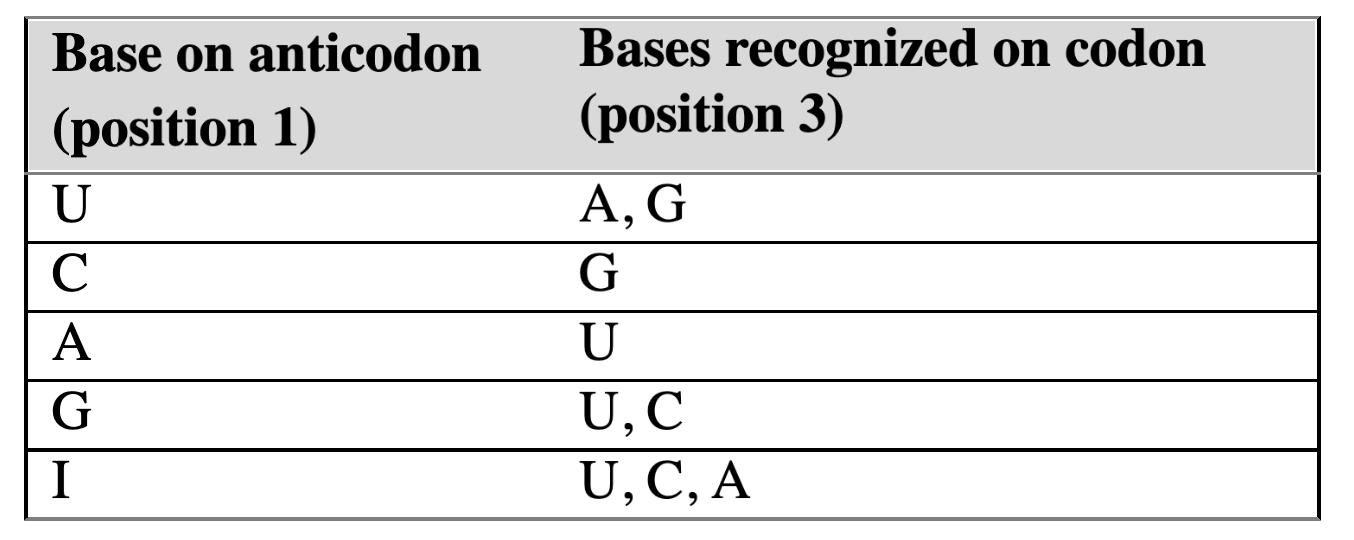
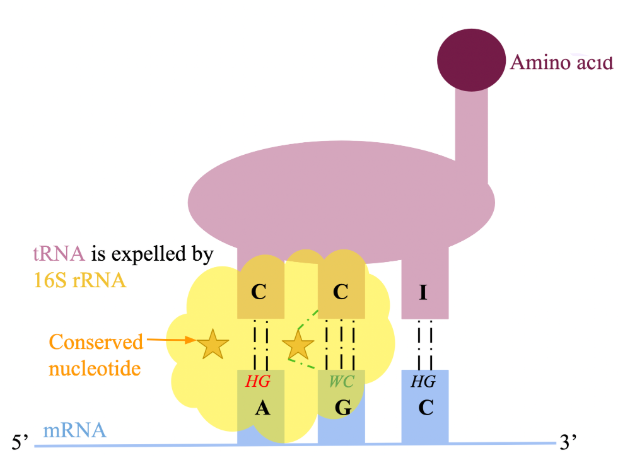
wobble hypothesis
codons that differ only in their third base can bind to the same tRNA anticodon
first two nucleotides of an mRNA codon must form strict WC bp with tRNA anticodon
3rd position has greater steric freedom and allows for both Hoogsteen and WC bp between tRNA and mRNA
wobble hypothesis regulation in ribosome
difference in flexibility of 3rd position is dictated by 16S rRNA of the S subunit
features 3 conserved nucleotides that H-bond with position 1 and 2 of the codon, 2 and 3 of the anticodon
correct H-bonding between these 3 can only occur if WC bp are formed
image: correct WC bp at position 2 enables 16S rRNA to form H-bonds with both anticodon & codon, unlike at position 1. this prevents a conserved nucleotide from making the correct H-bonds, resulting in expulsion. position 3 is not subject to scrutiny
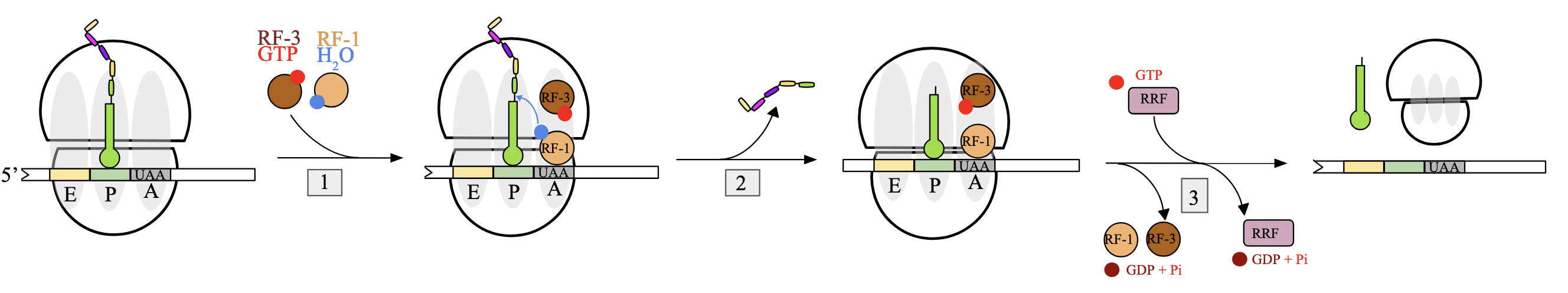
translation termination (prok)
no aminoacyl-tRNA molecules with anticodons complementary to 3 STOP codons; UAA, UGA, UAG.
when encountered at A site of the ribosome, they are instead recognized by release factors (RFs)
binding of release factor: when STOP codon is encountered in A site, RF-1 or RF-2 binds assisted by GTP-bound RF-3. RF will carry a molecule of water with them
peptide release: water molecule provided by RF used by ribosome to hydrolyze ester bond between pp chain and P site tRNA, releasing polypeptide
complex dissociation: RF-3 hydrolyzes GTP to GDP, leading to dissociation of RF. ribosomal release factor (RRF) binds ribosome and triggers release of entire translation complex by GTP hydrolysis
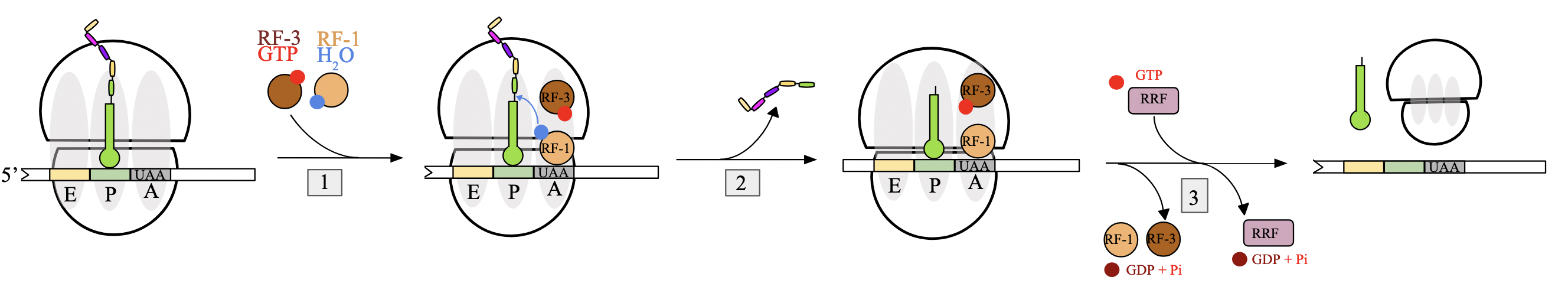
RF-1, RF-2, RF-3, RRF
RF-1: recognizes STOP codons UAA and UAG
RF-2: recognizes STOP codons UAA and UGA
both carry a water molecule into the A site to hydrolyze the ester bond between pp chain and tRNA in P site
RF-3: hydrolyzes GTP→GDP, leading to dissociation of RFs
RRF: hydrolyzes GTP→GDP, binds to the ribosome and triggers the release of the entire translation complex
prokaryotic translation ATPs GTPs for 9 amino acid pp chain
charging tRNAs: 2 molecules per charging event - 2×9=19 ATP
translation initiation: 1 molecule to bring in initiator tRNA - 1 GTP
aminoacyl-tRNA binding: 1 molecule for additional amino acids brought in by EF-Tu, hydrolyzed upon binding - 1×8=8 GTP
translocation: EF-G and GTP hydrolysis for each translocation - 1×8=8 GTP
translation termination: requires 1 GTP from RF-3 - 1 GTP
ribosome disassembly: occurs using RRF and GTP hydrolysis - 1 GTP
18 ATPs and 19 GTPs required to synthesize a 9 amino acid protein
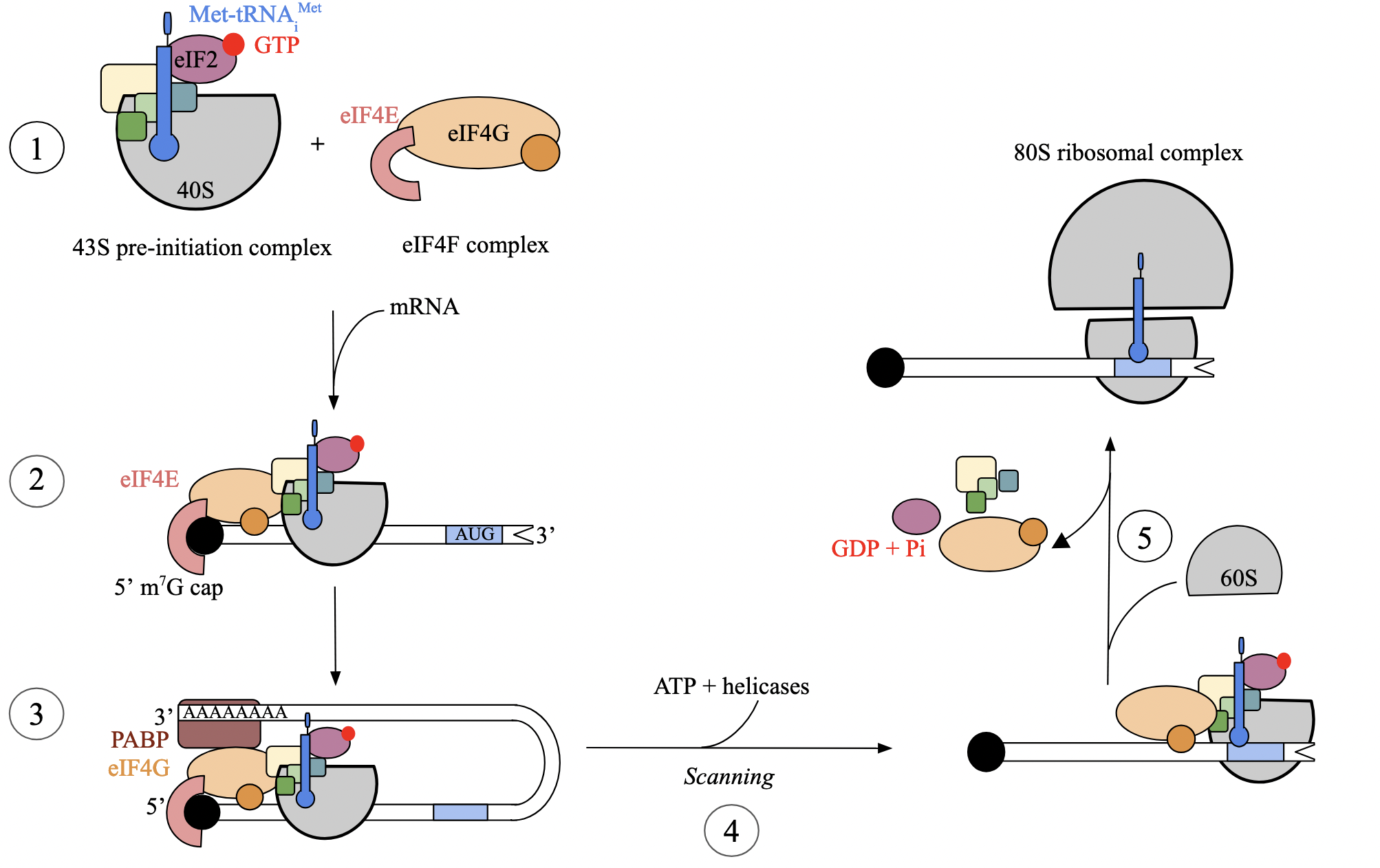
translation initiation: step 1 - assembly of complexes (euk)
independent assembly of two essential complexes:
eIF4F heterotrimer complex
includes eIF4E (binds to 7-methyl-G cap of mRNA)
43S pre-initiation complex
consists of small ribosomal subunit, Met-tRNAiMet, and several eukaryotic initiation factors (eIFs, like eIF2)
eIF2, bound to GTP, facilitates interaction between tRNAi and S subunit
tRNAiMet differs from tRNA species that carries internal methionines, but is not formylated. associates with S subunit devoid of mRNA
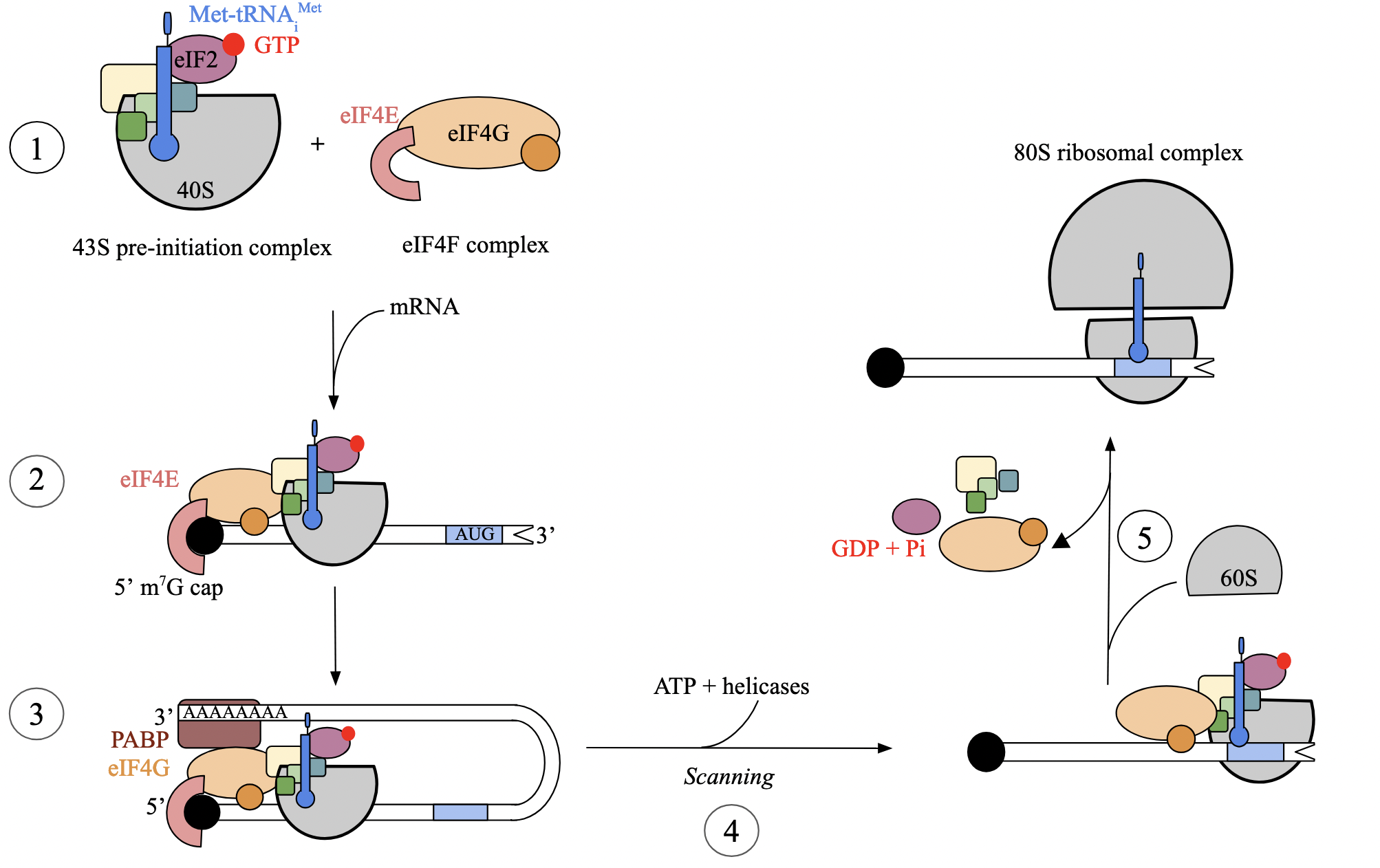
translation initiation: step 2 - complexes bind mRNA (euk)
both complexes bind to the mRNA at the 5’ cap
eIF4F, which binds to 7-me-G cap of mRNA
43S pre-initiation complex, which consists of several initiation factors and tRNAiMet
eIF4F complex attaches first through eIF4E subunit
followed by binding of 43S pre-initiation complex
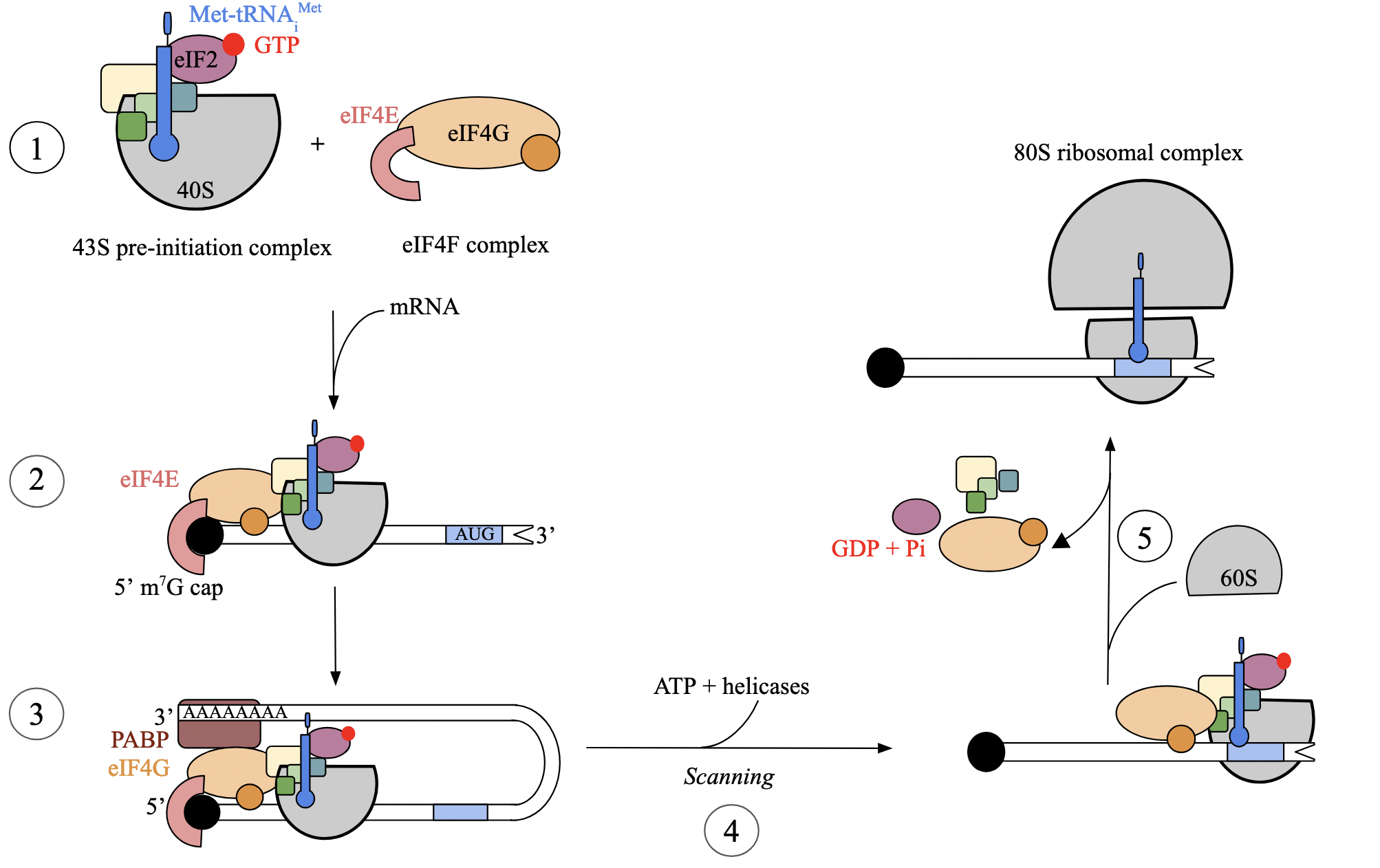
translation initiation: step 3 - circularization of mRNA (euk)
once both complexes are bound to the mRNA, transcript undergoes circularization
thought to improve ribosomal recycling by facilitating the re-binding of ribosome after the mRNA is finished translating
eIF4G component of eIF4F complex acts as bridge between cap-binding protein eIF4E and poly(A)-binding protein at 3’ end of mRNA
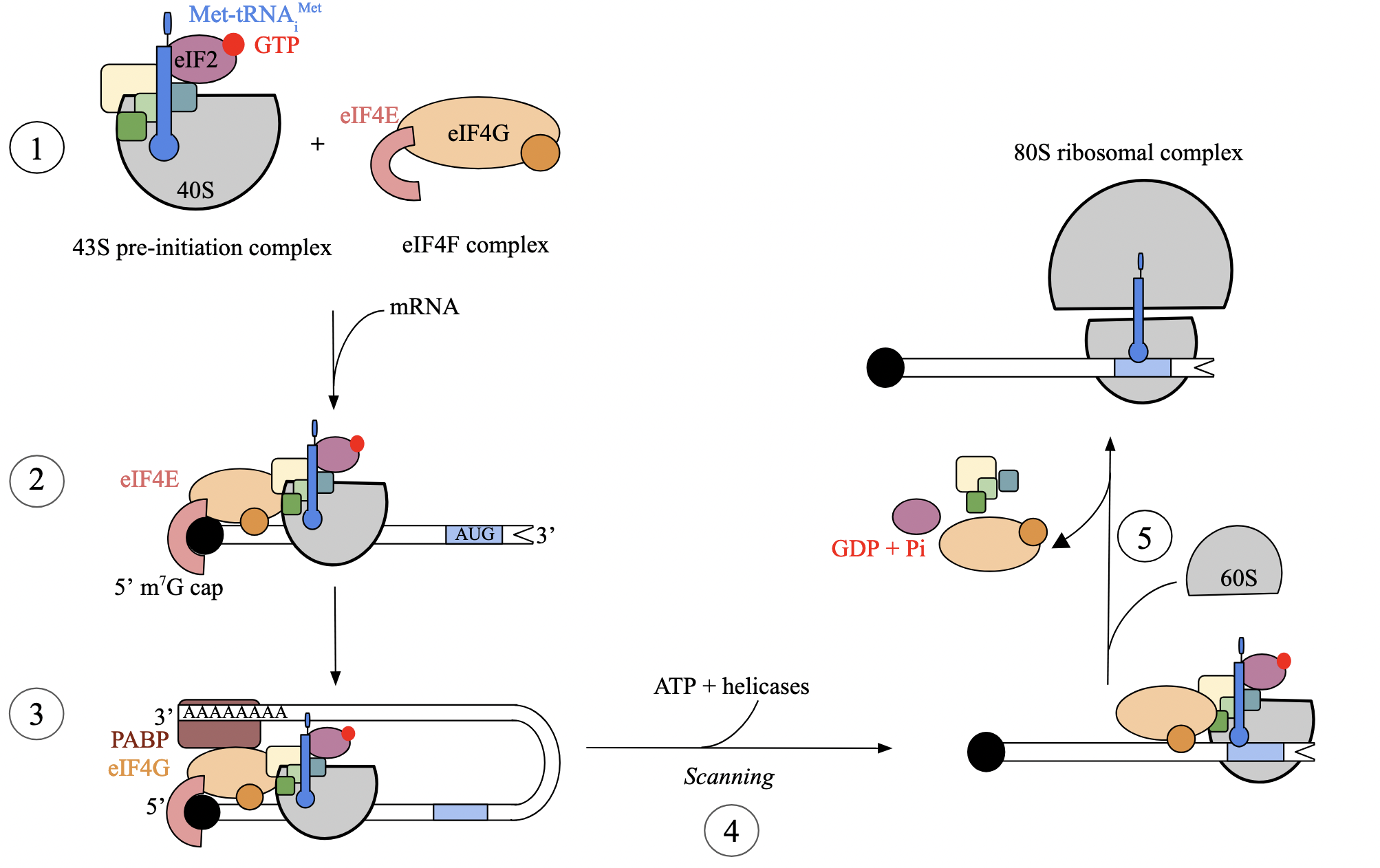
translation initiation: step 4 - scanning of mRNA (euk)
circularization initiates 5’ to 3’ scanning of the mRNA by the S subunit in search of the start codon
scanning driven by helicases and ATP hydrolysis
start codon is simply the first AUG located by the complex, closest to the 5’ end
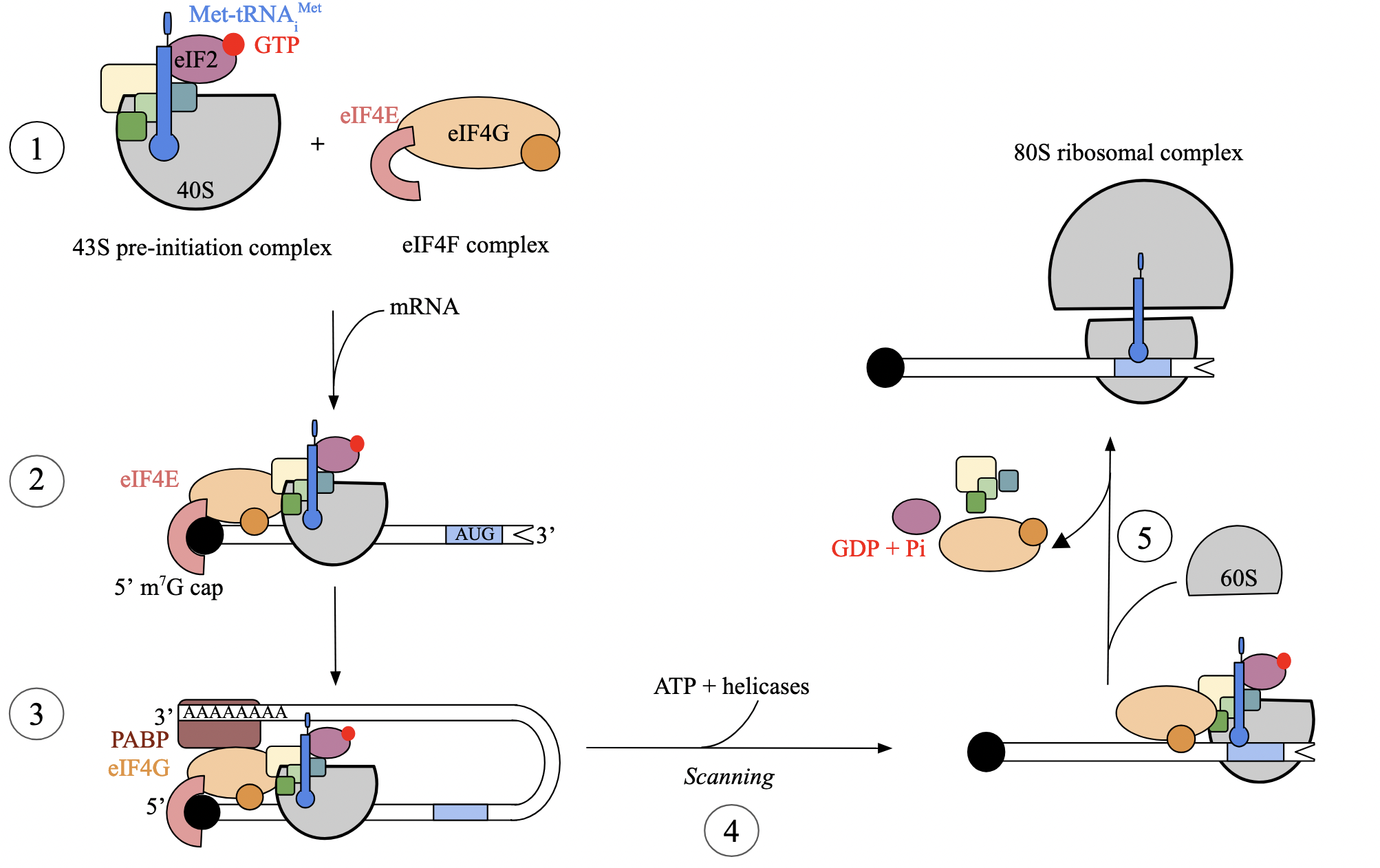
translation initiation: step 5 - recognition of initiation codon (euk)
when first AUG is encountered by S subunit, interaction between mRNA codon and cognate tRNA anticodon triggers GTP hydrolysis of eIF2
hydrolysis facilitates binding of L (60S) subunit and positioning of tRNAi at P site, forming 80S ribosome
once assembled, all eIFs dissociate, ribosome is now ready to proceed to the elongation phase
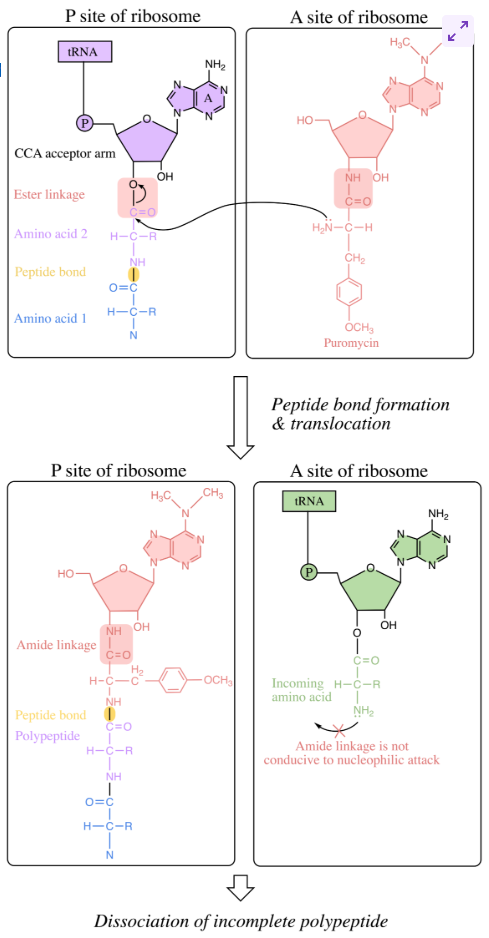
puromycin: ribosomes as drug targets
closely resembles A of tRNA acceptor arm (CCA), attached to an amino acid. features amide linkage instead of ester linkage
allows puromycin to fit into A site of ribosome as though it were a charged tRNA to be incorporated into pp chain
its N atom attacks C-terminal end of growing pp chain - forming normal peptide bond - pp chain is transferred to puromycin
incoming aa is unable to attack during the next round of translation elongation due to amide linkage
puromycin binds weakly to ribosome, leading to dissociation and halting of translation
affects both eukaryotes and prokaryotes
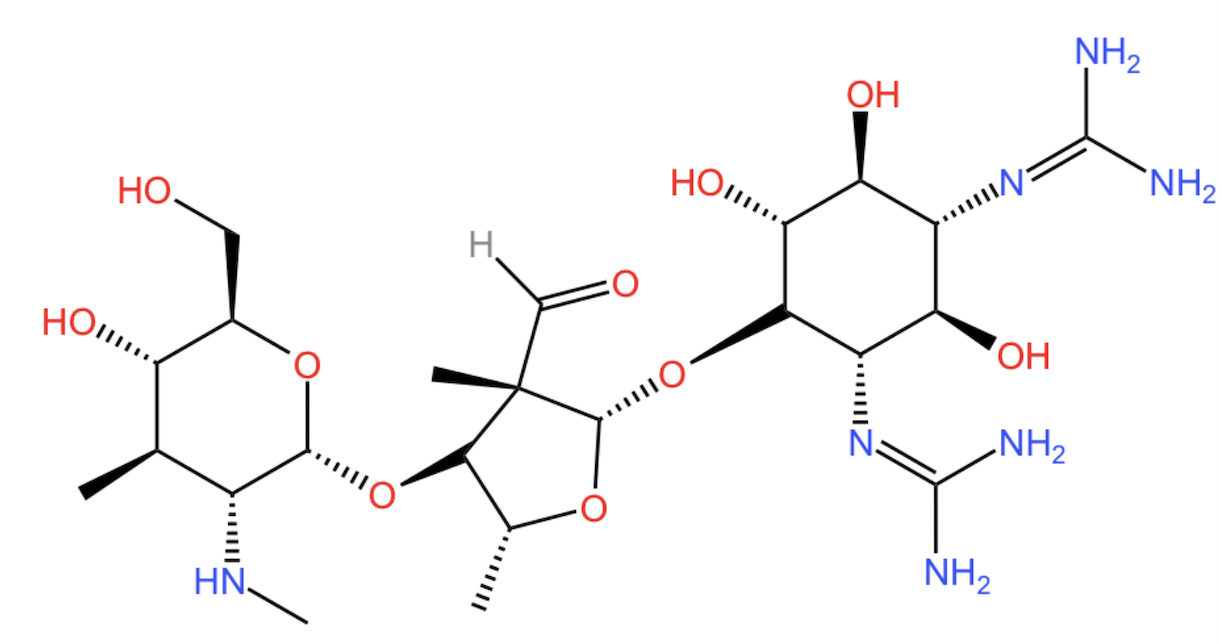
streptomycin: ribosomes as drug targets
streptomycin prevents correct binding of tRNAi in prokaryotes
due to streptomycin’s basic nature, conferred by many amino groups
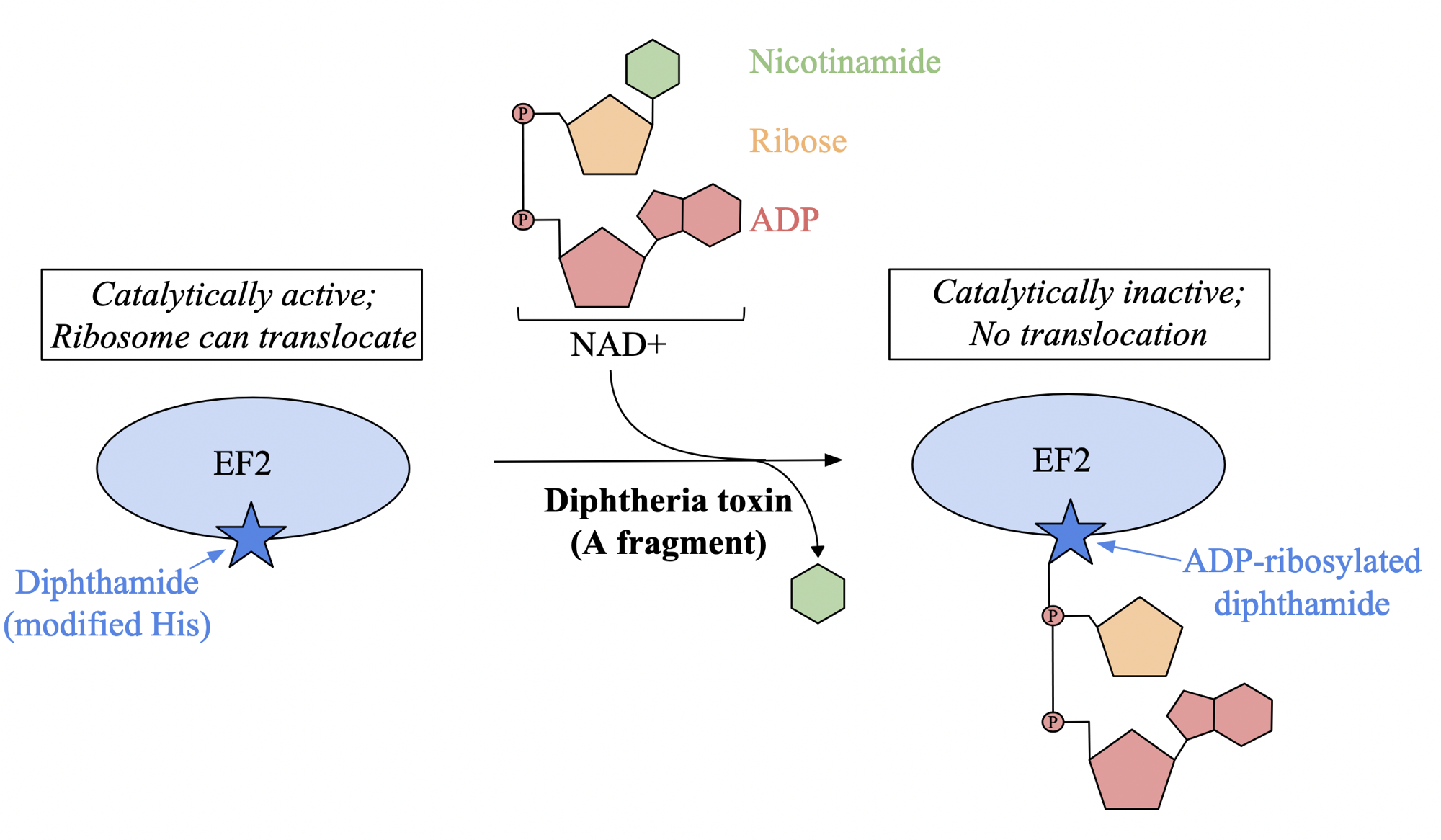
diptheria toxin: ribosomes as drug targets
affects eukaryotes
upon entering cell, toxin is cleaved into two fragments: A and B
A fragment acts as enzyme that catalyzes NAD+ cleavage into ADP-ribose and nicotinamide
toxin attaches ADP-ribose moiety to modified histidine residue in elongation factor 2 called dipthamide
modification inhibits EF2 ability to translocate to ribosome during protein synthesis, halting transformation
ricin: ribosomes as drug targets
N-glycosidase that inhibits protein synthesis in eukaryotes
heterodimer composed of two chains
A chain, has catalytic activity
B chain, responsible for cellular uptake, connected to A chain by single disulfide bond
when taken up by cells through endocytosis, A chain is released into cytoplasm
specifically targets a conserved adenine residue in 28S rRNA of ribosome, cleaving N-glycosidic bond between base and respective ribose sugar
prevents binding of EFs and inactivates ribosome