Pyruvate Dehydrogenase Complex & Krebs Cycle
1/59
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
60 Terms
There are multiple videos

explain what happens to Pyruvate under aerobic cell conditions, aerobic metabolism, discussed in glycolysis, glucose → pyruvate.
If the cell does NOT have mitochondria, it is called anaerobic metabolism.
then, we will identify the enzymes and cofactors required in the conversion of pyruvate to acetylCoA
















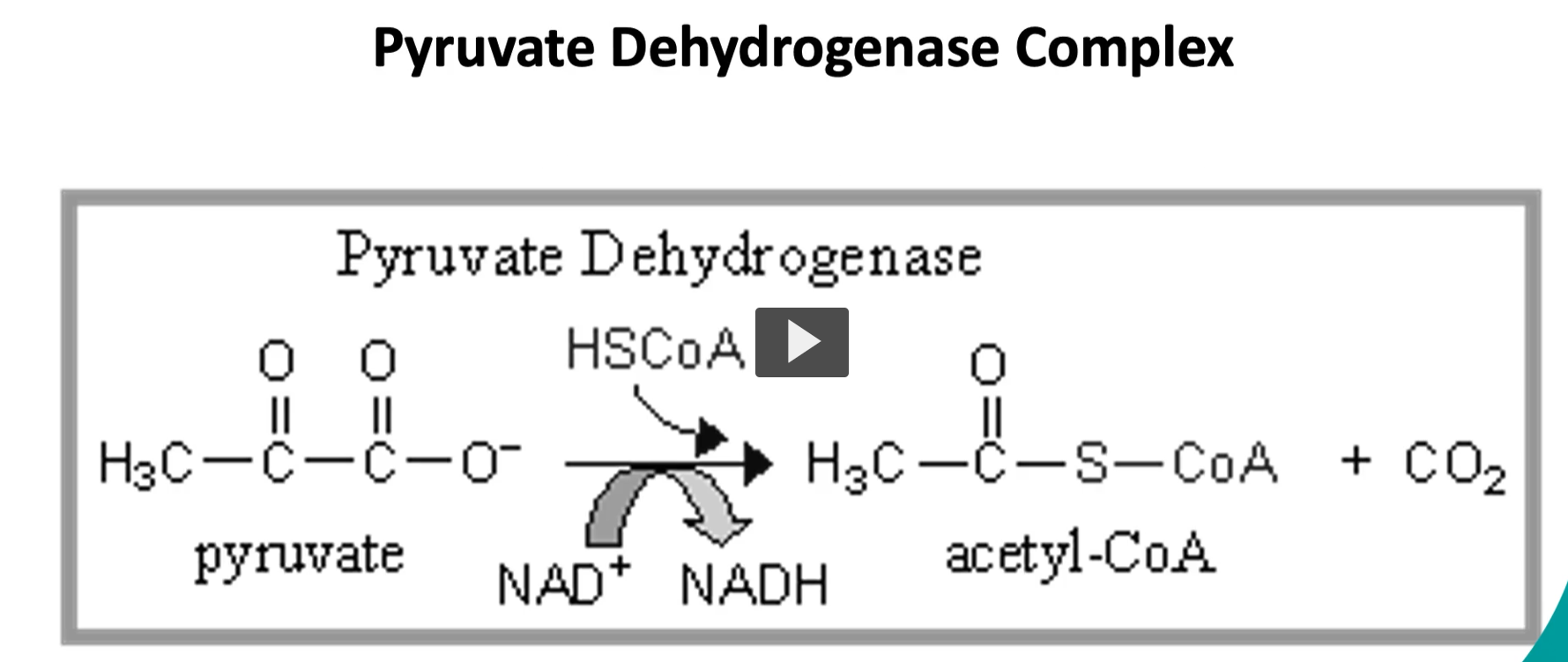
here is the pyruvate dehydrogenase complex, where the pyruvate on the left side is oxidized → acetyl-CoA and Co2 on the right side.
-this occurs via oxidative decarboxylation.
(pyruvate undergoes oxidative decarboxylation).
-since this pyruvate comes from glycolysis, there are two molecules of pyruvate entering this process, it is NECESSARY to remember to MULTIPLY BY 2.

you will find the pyruvate dehydrogenase complex located in the outer membrane of the mitochondria.
whereas Kreb’s Cycle and B-oxidation of fatty acids occurs in the mitochondrial matrix.
mito- = thread (Greek mitos)
chondrion = granule or grain
matrix = womb / source / container (Latin mater = mother)
So mitochondrial matrix means: the inner compartment inside the mitochondrion where important metabolic reactions occur.


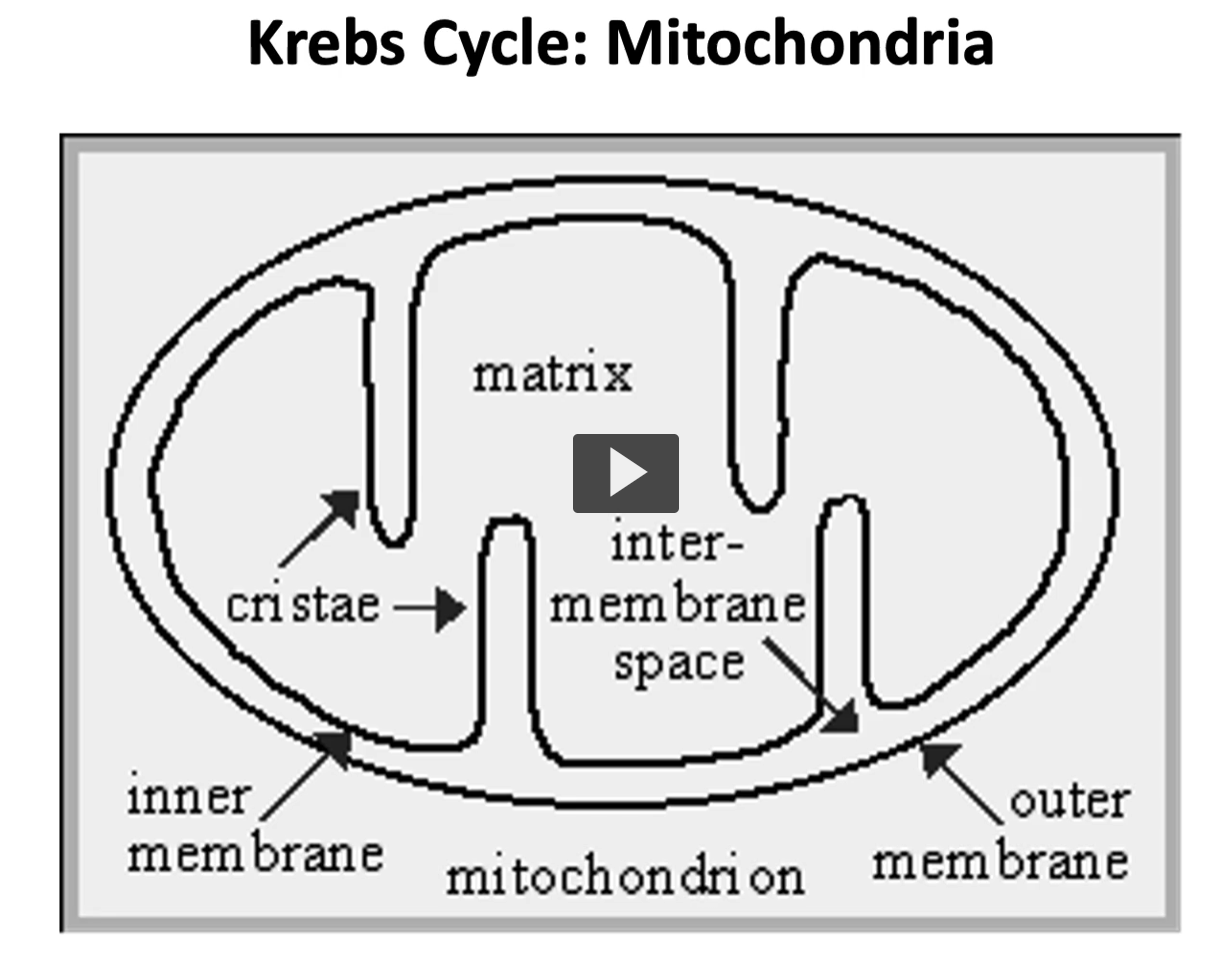
Here is an illustration of the mitochondria, the mitochondria is denominated as a semiautonomous organelle due to DNA replication and other important biological processes.
in this illustration, you can see the mitochondria inner and outer membrane, which have contrasting permeability characteristics.
The outer membrane is non-specifically permeable to all lower weight molecular solutes, whereas the inner membrane is impermeable to except through specific transporter.
then the matrix in the middle, is the soluble part where Kreb’s cycle and oxidative phosphorylation take place.

pyruvate dehydrogenase complex and the three enzymes are required are pyruvate dehydrogenase compound, denominated E1 with the coenzyme, TPP or thiamine pyrophosphate.
The Pyruvate Dehydrogenase Complex (PDC) is a large enzyme system that converts pyruvate into acetyl-CoA, which then enters the citric acid cycle (TCA cycle) in the mitochondrial matrix.
pyruvate dehydrogenase: Enzyme 1: E1 with coenzyme thiamine pyrophosphate (TPP).
dihydrolipyl transacetylase: Enzyme 2: E2 with coenzyme CoA and Lipoamide.
Dihydrolipoyl Dehydrogenase (E3) use NAD+ and FAD as coenzymes
WHAT DOES THIS MEAN?
1. TPP (Thiamine Pyrophosphate) Etymology
cofactor of pyruvate dehydrogenase (e1)
thiamine = vitamin B₁
pyrophosphate = two phosphate groups linked together
TPP = activated form of vitamin B₁.
What it does : TPP helps remove carbon dioxide (CO₂) from molecules.
In the pyruvate dehydrogenase complex it helps:
Pyruvate→ decarboxylation
So TPP stabilizes the reactive carbon intermediate after CO₂ is removed.
Simple role: TPP helps break carbon–carbon bonds.
2. Lipoamide Etymology
cofactor of dihydrolipyl transacetylase (e2)
lipo- = fat
amide = chemical bond involving nitrogen
It carries the acetyl group from one enzyme active site to another.
It also participates in redox reactions (accepting and donating electrons).
Simple role
Lipoamide transfers the acetyl group during the reaction.
3. CoA (Coenzyme A) Etymology
cofactor of dihydrolipyl transacetylase (e2)
coenzyme = enzyme helper
A originally meant acetylation
Simple role CoA is the carrier of acetyl groups.
4. FAD (Flavin Adenine Dinucleotide) Etymology
cofactor of Dihydrolipoyl Dehydrogenase (E3)
flavin = yellow compound derived from riboflavin
adenine = nitrogen base in nucleotides
dinucleotide = two nucleotide units
What it does: FAD accepts electrons during oxidation reactions.
FAD+2e−+2H+→FADH2
It helps transfer electrons to the next molecule in the chain.
Simple role: FAD is an electron carrier.
5. NAD⁺ (Nicotinamide Adenine Dinucleotide) Etymology
cofactor of Dihydrolipoyl Dehydrogenase (E3)
nicotinamide = derived from niacin (vitamin B₃)
adenine dinucleotide = nucleotide structure
What it does NAD⁺ accepts high-energy electrons.
NAD++2e−+H+→NADH
NADH then carries these electrons to the electron transport chain, which produces ATP.
Simple role: NAD⁺ is another electron carrier used to make energy.
Dihydrolipoyl Dehydrogenase (E3) use NAD+ and FAD as coenzymes
the delta G for the complex as a whole is -8.3 Kcal/mol, it is an exergonic and irreversible process.

1. What the Pyruvate Dehydrogenase Complex does
After glycolysis:
Glucose→Pyruvate
Pyruvate must be converted into acetyl-CoA to enter the TCA cycle.
The reaction:
Pyruvate+CoA+NAD+→Acetyl-CoA+CO2+NADH
This reaction occurs in the mitochondrial matrix and requires three enzymes working together.
Step 1 — E1 (Pyruvate Dehydrogenase)
Coenzyme: TPP (Thiamine pyrophosphate)
Function: E1 removes CO₂ from pyruvate.
Pyruvate→ Hydroxyethyl-TPP + CO2
This step is called decarboxylation.
TPP stabilizes the reactive intermediate.
Vitamin connection:
TPP comes from vitamin B₁ (thiamine).
Step 2 — E2 (Dihydrolipoyl Transacetylase)
Coenzymes:
Lipoamide
CoA
Function: The hydroxyethyl group is transferred to lipoamide, forming an acetyl group.
Then the acetyl group is transferred to CoA.
Result: Acetyl-CoA
This is the molecule that enters the TCA cycle.
Step 3 — E3 (Dihydrolipoyl Dehydrogenase)
Coenzymes:
FAD
NAD⁺
Function: E3 restores (reoxidizes) the lipoamide so the cycle can continue.
Electron flow:
Reduced lipoamide→FAD→NAD+
This produces: NADH
NADH later generates ATP through the electron transport chain
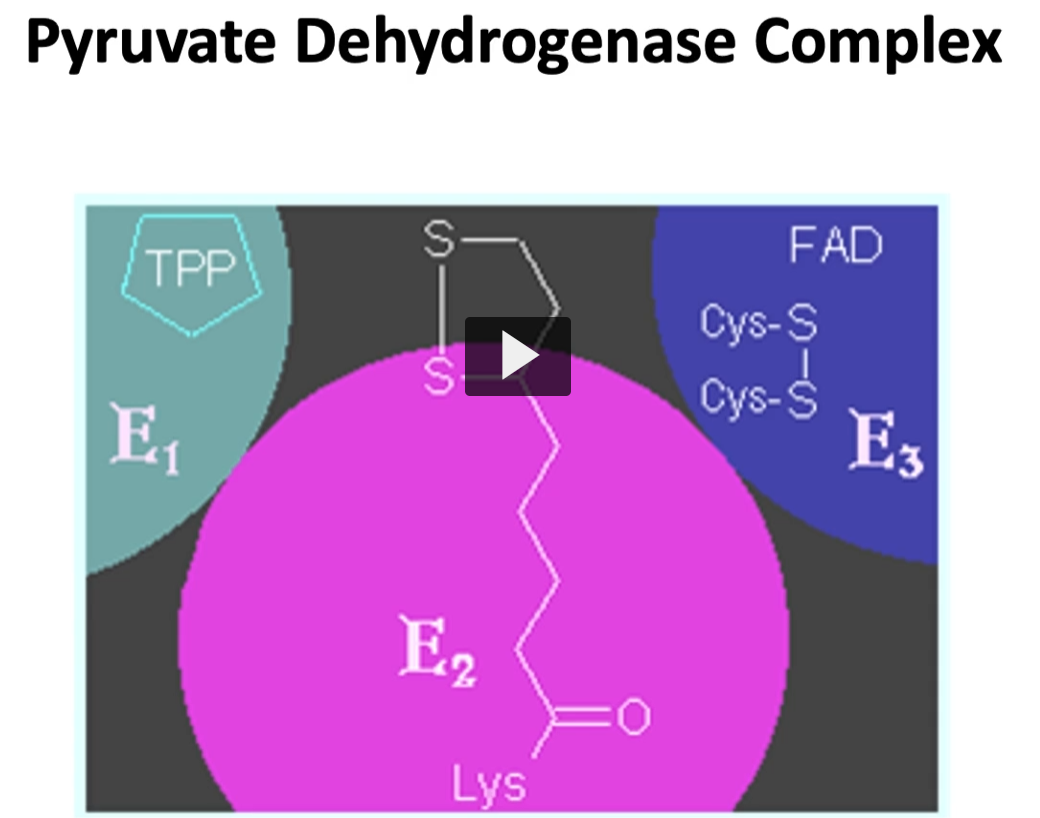
here are shown all three enzymes, previously mentioned, forming a multi-enzymatic complex.
all three enzymes are strategically located since the product of each reaction is the substrate to the following enzyme. This allows a quick turnover
again, this is another illustration of pyruvate dehydrogenase complex shows how pyruvate enters on the left and the net product is acetyl-CoA on the top and NADH per pyruvate molecule that it is oxidized, and you can see the NADH on the right side.


Step 1 — Enzyme 1 (E1): Pyruvate Dehydrogenase
Cofactor: TPP (Thiamine pyrophosphate)
What happens:
Pyruvate enters the enzyme.
CO₂ is removed (decarboxylation).
The remaining 2-carbon fragment attaches to TPP.
Intermediate formed: Hydroxyethyl-TPP
So:
Pyruvate→Hydroxyethyl-TPP+CO2
1. Etymology of the name Pyruvate
So the enzyme acts on pyruvate.
Dehydrogenase
Breakdown of the word:
de- = remove
hydrogen = H atoms (or electrons)
-ase = enzyme
dehydrogenase = an enzyme that removes hydrogen (electrons) from a molecule.
Why it’s called a dehydrogenase
Since the enzyme removes hydrogen/electrons from pyruvate and transfers them to NAD⁺:
NAD+→NADH\text{NAD}^+ \rightarrow \text{NADH}NAD+→NADH
the enzyme is classified as a dehydrogenase.
4. Important detail
Even though we call it pyruvate dehydrogenase, the reaction actually involves three enzymes working together (E1, E2, E3).
But the name comes from the first step, where pyruvate begins the reaction.
Step 2 — Enzyme 2 (E2): Dihydrolipoyl Transacetylase
dihydro: 2 hydrogen, “Two hydrogens added” or reduced form of a molecule.
In this enzyme, it refers to reduced lipoamide, which has two hydrogen atoms attached after it accepts electrons
Lipoyl comes from lipoic acid, a cofactor attached to the enzyme.
Trans-: “to move across” or “transfer.”
Acetyl refers to a 2-carbon group:
This is the same acetyl group that forms acetyl-CoA.
ase: enzyme
Cofactors:
Lipoamide
Coenzyme A (CoA)
1. Start: Oxidized lipoamide
Lipoamide initially exists in the oxidized form, which has a disulfide bond (-S-S-).
This is the starting form attached to E2.
Structure conceptually:
oxidized lipoamide
S—S (disulfide bond)
2. Transfer from TPP (cofactor of e1) → lipoamide (cofactor of e2)
From E1, the hydroxyethyl group attached to TPP, becomes oxidized to acetyl, and the acetyl is transferred to lipoamide.
During the hydroxyethyl transfer to Lipoamide:
the hydroxyethyl group becomes oxidized → acetyl
the lipoamide was in oxidized form, lipoamide disulfide bond is reduced (the hydroxyethyl group was oxidized to acetyl, the lipoamide was reduced to acetyl-lipoamide).
Result: acetyl-lipoamide (reduced) (NOT FULLY REDUCED yet).
Meaning:
the acetyl group is now attached to lipoamide
the disulfide bond has opened
So now we have: acetyl-lipoamide
3. Transfer of acetyl group to CoA
Next step: The acetyl group is transferred from lipoamide to CoASH.
(CoASH = Coenzyme A with a free sulfhydryl group).
Reaction:
acetyl-lipoamide + CoASH → acetyl-CoA + reduced lipoamide
Result:
acetyl-CoA is produced
lipoamide is now reduced (two -SH groups)
So now we have: FULLY reduced lipoamide
4. Regeneration of oxidized lipoamide
The enzyme complex must reset the lipoamide so the cycle can continue.
This happens in E3.
Steps:
Reduced lipoamide transfers electrons to FAD
FAD becomes FADH₂
FADH₂ transfers electrons to NAD⁺
NAD⁺ becomes NADH
During this process:
reduced lipoamide → oxidized lipoamide
Now the system is ready for another cycle.
This form is ready to accept the 2-carbon fragment from E1.
Result: Acetyl-CoA
This is the molecule that enters the Krebs cycle.


Step 3 — Enzyme 3 (E3): Dihydrolipoyl Dehydrogenase
Cofactors:
FAD
NAD⁺
What happens:
Lipoamide became reduced during step 2.
E3 re-oxidizes lipoamide so the enzyme can work again.
Electrons move:
Reduced lipoamide→ FAD→NAD+
This produces:
FADH₂
then NADH
Final product:
NADH
NADH carries electrons to the electron transport chain to make ATP.

The Pyruvate Dehydrogenase Complex (PDC) is a large enzyme system that converts pyruvate into acetyl-CoA, which then enters the citric acid cycle (TCA cycle) in the mitochondrial matrix.
what happens to acetyl-CoA once it is formed?
answer: acetyl-CoA can follow different directions depending on the metabolic needs of the cell.
One pathway is to go into the TCA cycle: where acetyl-CoA is combined with oxaloacetate to produce Citrate (acetyl-CoA + oxaloacetate= citrate).
another pathway leads to the formation of ketone bodies during starvation (acetyl-CoA can become ketone bodies during starvation)
or when blood glucose is high, acetyl-CoA can go into cholesterol, fatty acid synthesis, which is a fed state.
regulation
many reversible reactions need regulation for inhibition, and we will talk about the ones involved in pyruvate dehydrogenase complex
regulation of E1
Enzyme 1 (E1) is inhibited by high concentration of ATP, thus, we can say that ADP will cause the opposite and accelerate when there is low concentration of ATP. why?
Enzyme 1 (E1) of the pyruvate dehydrogenase complex converts:
Pyruvate→ Acetyl-CoA
This step allows pyruvate to enter the citric acid cycle, which produces ATP.
So activating this enzyme leads to more ATP production
regulation of E2
Enzyme 2 (E2) is inhibited by high concentration of its product, which is acetyl-CoA, by feedback inhibition as well.
1. What Enzyme 2 (E2) does
E2 in the pyruvate dehydrogenase complex is:
Dihydrolipoyl transacetylase
Its job is to transfer the acetyl group to CoA, producing: Acetyl-CoA
So E2 is the step that actually creates acetyl-CoA.


and the last enzyme, E3, is inhibited by its product NADH (why?)
What Enzyme 3 (E3) does
E3 is called:
Dihydrolipoyl dehydrogenase
Its job is to transfer electrons to NAD⁺, producing NADH.
Reaction:
NAD++2e−+H+→NADH
So E3 helps generate NADH, which carries high-energy electrons to the electron transport chain.
liver control over PDC with insulin, cAMP, glucagon and glucose.
another control seen in pyruvate dehydrogenase complex is in the liver, the first enzyme is phosphorylated and inactivated by the cyclic AMP (cAMP) cascade.
this cAMP cascade can be controlled by stress hormone glucagon since it is in the liver.
the presence of glucagon, during starvation will help preserve glucose and stop the pyruvate dehydrogenase complex, preventing formation of acetyl-CoA due to irreversibility.
then, in a fed state, insulin will be predominant and cyclic AMP will be activated by causing dependent phosphate.


as a summary, in this illustration, we see that E1 E2 and E3 are controlled by their products, and in the liver, is controlled by phosphorylation during starvation conditions
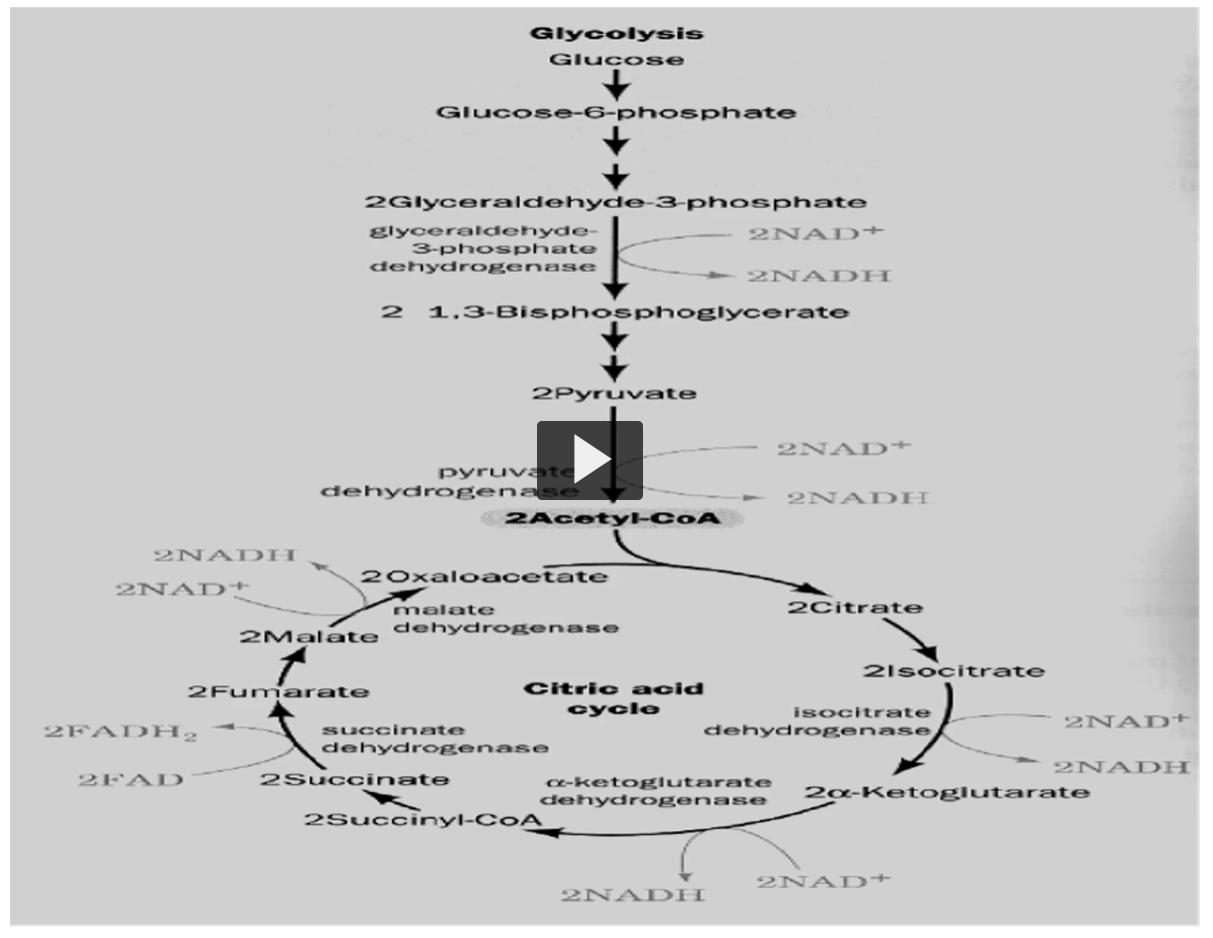
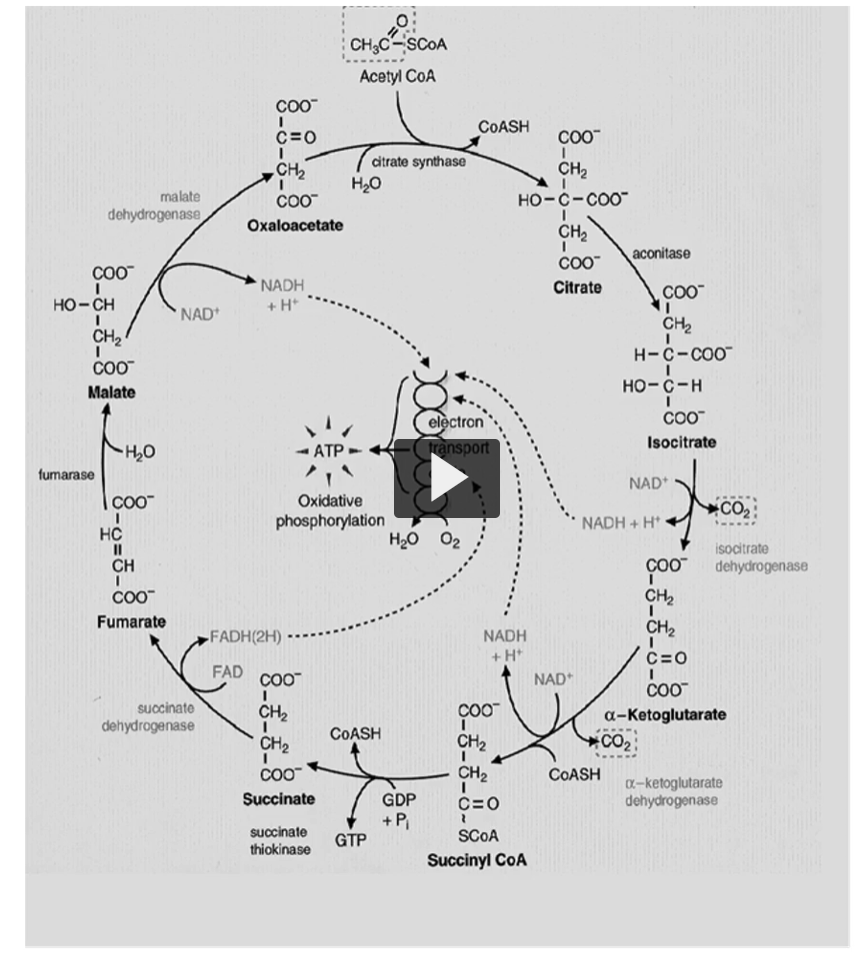
there are eight reactions or carbon versions in Kreb’s cycle, shown here.
oxidations are only four, in which three are made by NAD and one by FAD.
Here you can see the NAD (bottom right, 2 NAD+ AND 2NADH), another one here (bottom), and the last one on top (top left of the bottom circle), on the bottom left you can see the FAD (2 FAD, 2 FADH)
remember, this is once per cycle (he is referring to the big circle).

glucose is not the only source of acetyl-CoA.
Glucose is not the only source of acetyl-CoA .
as depicted here, the oxidation of many fields, like
1. fatty acids
2. ketone bodies
3. amino acids
will generate acetyl-CoA.
on top are the ones I just mentioned, and producing acetyl-CoA that will be into the kreb cycle
ATP not directly formed
ATP is not directly formed in Kreb’s cycle, but the reduction of NAD and FAD into NADH and FADH2 will carry the electrons needed in oxidation phosphorylation to produce ATP
1. What the Krebs cycle actually produces
When acetyl-CoA enters the Krebs (TCA) cycle, most of the energy is captured in reduced electron carriers:
Products per acetyl-CoA:
3 NADH
1 FADH₂
1 GTP (≈ ATP)
CO₂
So technically one ATP-equivalent (GTP) is produced, but the major energy yield comes later.
2. What NADH and FADH₂ are
NADH and FADH₂ are electron carriers.
During the Krebs cycle:
NAD⁺ accepts electrons → becomes NADH
FAD accepts electrons → becomes FADH₂
These electrons store high-energy chemical energy.
Think of them like charged batteries.
3. Where ATP is actually made
The electrons carried by NADH and FADH₂ go to the electron transport chain in the inner mitochondrial membrane.
There, the electrons move through a series of proteins.
As they move:
protons (H⁺) are pumped across the membrane
this creates a proton gradient
4. Oxidative phosphorylation
The proton gradient powers the enzyme ATP synthase.
Protons flow back through ATP synthase and this drives:
ADP+Pi→ATP
This process is called oxidative phosphorylation.
only high energy compound formed during Kreb’s cycle
the only high energy compound formed during Kreb’s cycle is GTP from GDP. This will then GDP is interconverted to ATP by a nucleoside, diphosphate kinase.
why do fatty acids produce Acetyl-CoA?
1. Fatty acids → acetyl-CoA
Fatty acids are broken down by a process called β-oxidation in the mitochondrial matrix.
What happens
The long fatty acid chain is cut into 2-carbon pieces repeatedly.
Each cycle of β-oxidation removes:
2 carbons→acetyl-CoA
Example:
A 16-carbon fatty acid (palmitate):
16C→8 acetyl-CoA
So fatty acids produce many acetyl-CoA molecules, which is why fats are a very energy-rich fuel.
why do ketone bodies produce acetyl-CoA?
2. Ketone bodies → acetyl-CoA
Ketone bodies are produced in the liver during fasting or starvation.
Main ketone bodies:
acetoacetate
β-hydroxybutyrate
Other tissues (brain, muscle) convert these ketone bodies back into acetyl-CoA.
Example pathway: Acetoacetate→ Acetoacetyl-CoA
Then: Acetoacetyl-CoA→2 acetyl-CoA
These acetyl-CoA molecules then enter the TCA cycle to produce ATP.
why do amino acids produce acetyl-CoA?
3. Amino acids → acetyl-CoA
Some amino acids are called ketogenic amino acids because their breakdown produces acetyl-CoA.
Examples of ketogenic amino acids:
Leucine
Lysine
Other amino acids are partially ketogenic, such as:
isoleucine
phenylalanine
tyrosine
tryptophan
When these amino acids are metabolized, their carbon skeletons are converted into acetyl-CoA or acetoacetate.
SECOND VIDEO
pyruvate dehydrogenase complex and krebs cycle part 2
we are continuing this topic, we discussed the pyruvate dehydrogenase complex, which is a multienzymatic, converting pyruvate → acetyl-CoA
-the localization is in the outer membrane of the mitochondria


as a quick recap, we talk about Kreb’s and beta oxidation of fatty acids occur in the mitochondrial membrane and how the mitochondria is a semiautonomous organelle.
here is an illustration of the mitochondria with its different spaces.
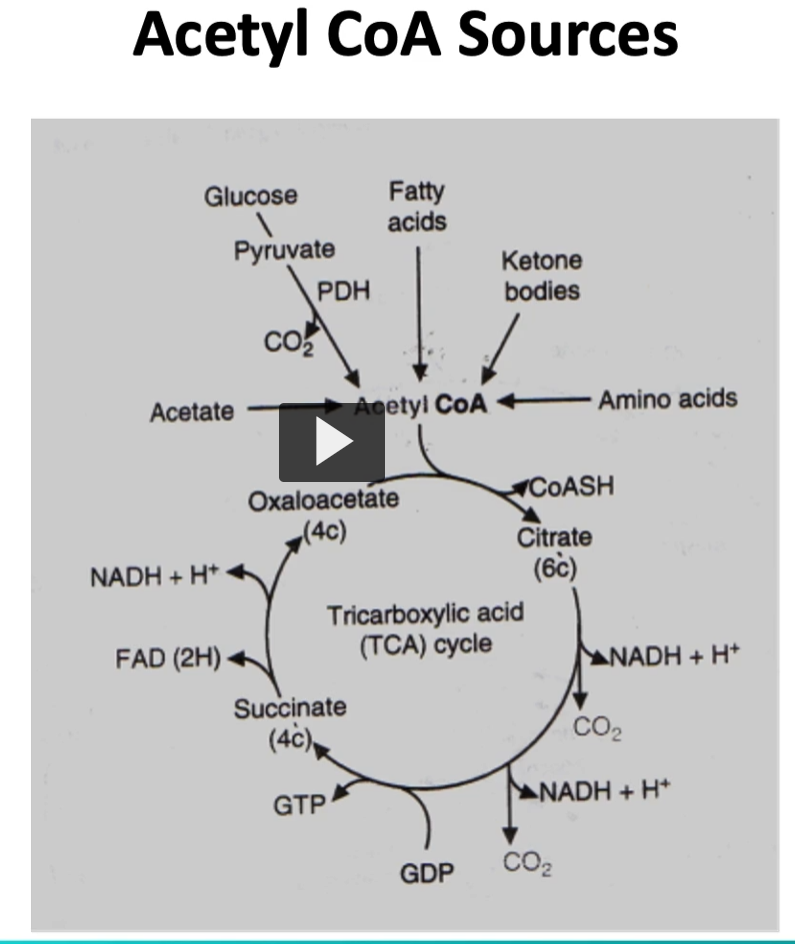
we mentioned the different fields that produce acetyl-CoA, such as fatty acids, ketone bodies, and amino acids.
Kreb’s cycle illustration shows oxidation, are only four, in which three are made by NAD and one by FAD.
Remember that this GTP is interconverted to ATP.
ATP is not directly formed in Kreb cycle, but the oxidation of NAD and FAD into NADH and FADH2 will carry the electrons needed in the oxidative phosphorylation process to produce ATP.
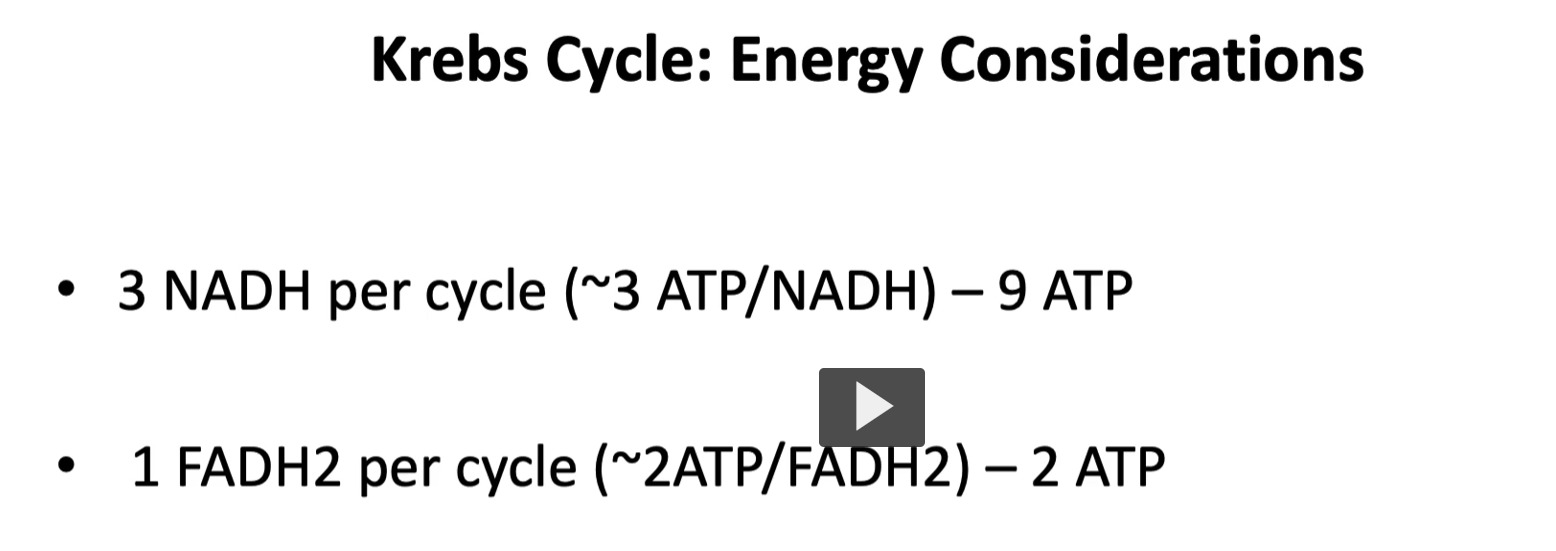
When talking of energy production, you will find different sources with different values.
but for this lecture, we will assume that one NADH will form three ATP.
1 NADH= 3 ATP
3 NADH= 9 ATP.
Since there are three NADH in a single Kreb cycle, the total ATPs formed are 9.
for each FADH2, the ATP produced is 2.
1 FADH2= 2 ATP.
in addition, and as mentioned in the last diagram, remember that 1 GTP is converted to ATP by nucleoside diphosphate kinase. ( GTP+ADP→GDP+ATP).
What GTP is
GTP is very similar to ATP.
Both contain:
a nucleotide
three phosphate groups
stored chemical energy
The only difference is the base:
Molecule | Base |
|---|---|
ATP | Adenine |
GTP | Guanine |
Both can store and transfer energy.
3. What nucleoside diphosphate kinase does
The enzyme nucleoside diphosphate kinase transfers a phosphate group.
Reaction:
GTP+ADP→GDP+ATP
What happens:
GTP donates one phosphate
ADP receives that phosphate
ATP is formed


the kreb’s cycle occurs under aerobic conditions (because oxygen is the final electron acceptor in oxidative phosphorylation). The reason is that NADH and FADH2 provides the electrons needed in the oxidative phosphorylation system, in the final phase, oxygen will accept that electron to be reduced and form water.
if oxygen is not present, as such in anaerobic conditions, there is no driving force to pull the electron through the oxidative phosphorylation system
anabolic aspect of the Kreb’s cycle
-the Krebs cycle is not only for energy (catabolism), it also supplies building blocks for biosynthesis (anabolism). This is why the cycle is often called an amphibolic (anabolic and catabolic) pathway.
the anabolic aspect of the kreb’s cycle means that the metabolites generated can also be precursors to other molecules like succinyl-CoA, can form heme or purines, alpha ketoglutarate can form glutamate, citrate to fatty acids, oxaloacetate to aspartate and then to amino acids.
as you can see, in this cycle, an H of the products.
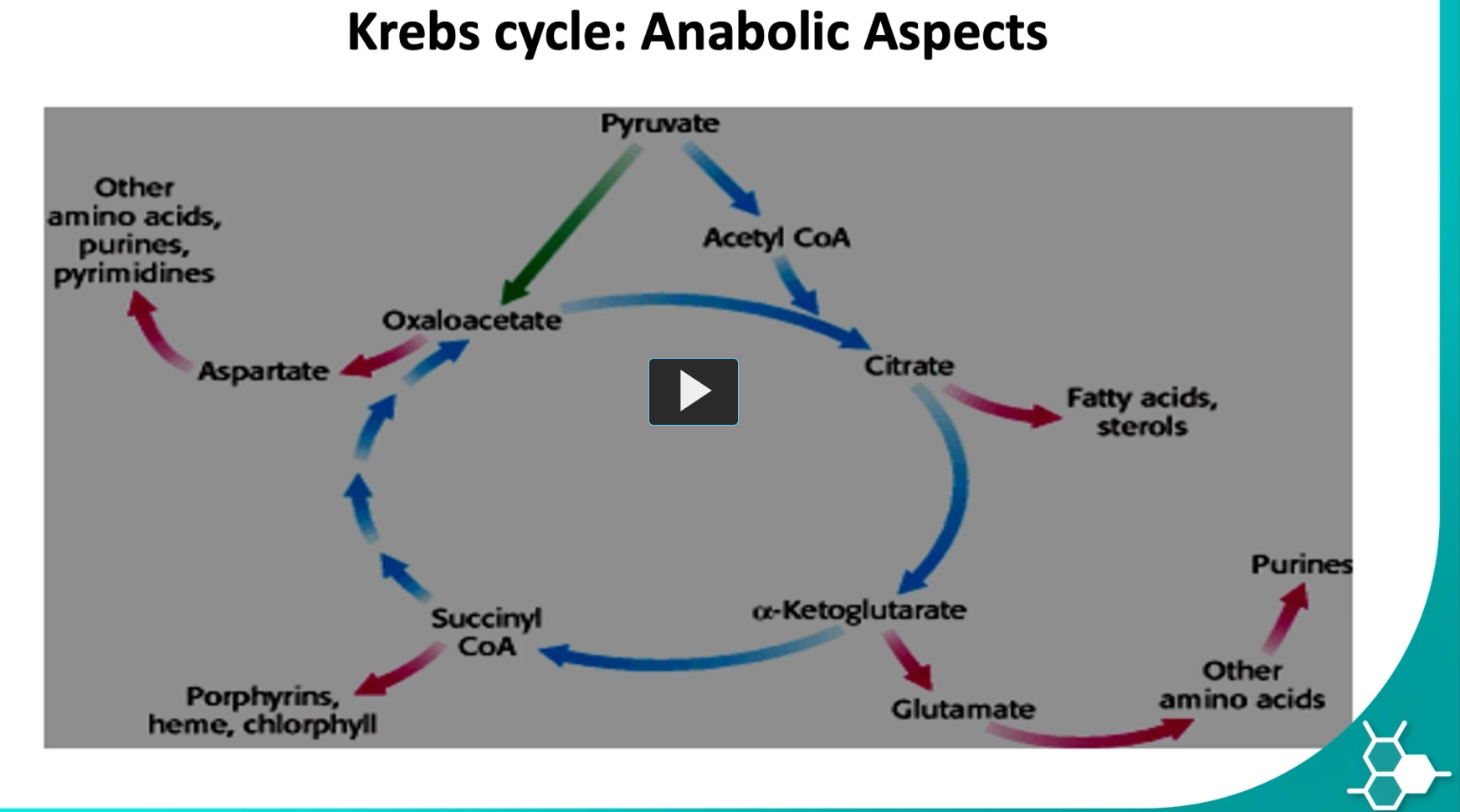

anaplerotic reactions
Anaplerotic
ana- = again / back
plērōsis = filling or replenishing
Anaplerotic reactions = reactions that refill or replenish the Krebs cycle.
we mentioned that the molecules of the Kreb’s cycle can be used in other places, but if that happens, if they are not replaced back, the Kreb’s cycle can collapse.
Thus, in order to replace the lost compounds, we have the anaplerotic reactions to form the lost metabolites.
example of anaplerotic reactions are:
pyruvate to Oxaloacetate
Propionyl to Succinyl-CoA
Aspartate to Oxaloacetate
Glutamate to a-ketoglutarate

Remember: anaplerotic reactions = “refilling reactions.”
When Krebs cycle molecules are taken out to make other things (amino acids, heme, fatty acids, etc.), the cell must replace them so the cycle can keep producing energy.
1. Pyruvate → Oxaloacetate
Reaction: Pyruvate+CO2+ATP → Oxaloacetate
Enzyme: Pyruvate carboxylase
Meaning:
Pyruvate (from glycolysis) is converted into oxaloacetate
Oxaloacetate is the molecule that starts the Krebs cycle
This reaction refills oxaloacetate when levels are low.
2. Propionyl-CoA → Succinyl-CoA
Propionyl-CoA comes from breakdown of:
odd-chain fatty acids
certain amino acids
It is converted into:
Propionyl-CoA → Succinyl-CoA
Succinyl-CoA is a Krebs cycle intermediate, so this reaction adds new material into the cycle
3. Aspartate → Oxaloacetate
Aspartate is an amino acid.
It can be converted into oxaloacetate through a reaction called transamination.
Aspartate → Oxaloacetate
This again replenishes oxaloacetate in the Krebs cycle.
*both pyruvate and aspartate can form Oxaloacetate
4. Glutamate → α-ketoglutarate
Glutamate is another amino acid.
It can lose an amino group through deamination:
Glutamate→α-ketoglutarate
α-ketoglutarate is a Krebs cycle intermediate, so this reaction refills the cycle.
5. Why these reactions are important
The Krebs cycle constantly loses intermediates because they are used to make other molecules.
Examples:
succinyl-CoA → heme
oxaloacetate → aspartate
citrate → fatty acids
If these molecules leave and are not replaced, the cycle slows down.
Anaplerotic reactions replace them so the cycle keeps running
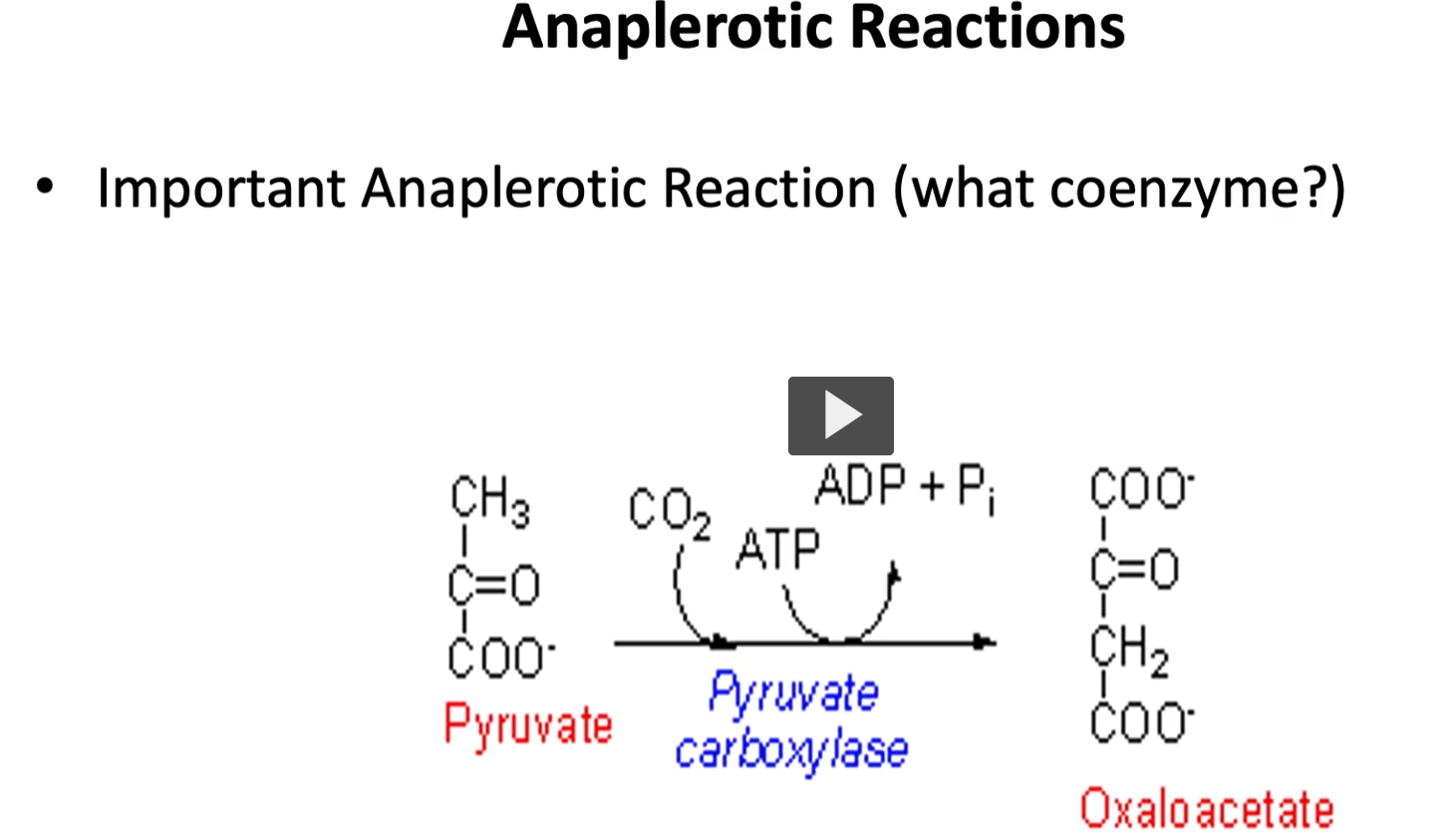
most important anaplerotic reaction
from all the anaplerotic reactions mentioned, the most important is the one that goes from pyruvate to oxaloacetate because oxaloacetate is the substrate needed in the first reaction, the coenzyme used by pyruvate carboxylase in this step is biotanimite along with ATP.
pyruvate →> oxaloacetate
enzyme (pyruvate carboxylase, The enzyme adds a carbon dioxide (CO₂) to pyruvate.)
This process is called carboxylation (adding CO₂).
Oxaloacetate
oxa- → refers to oxalic acid (a 2-carbon acid)
acetate → a 2-carbon acetyl-related compound
So oxaloacetate is a 4-carbon molecule formed from oxalo- and acetyl-type fragments.
Chemically it is a four-carbon dicarboxylic acid used in metabolism.
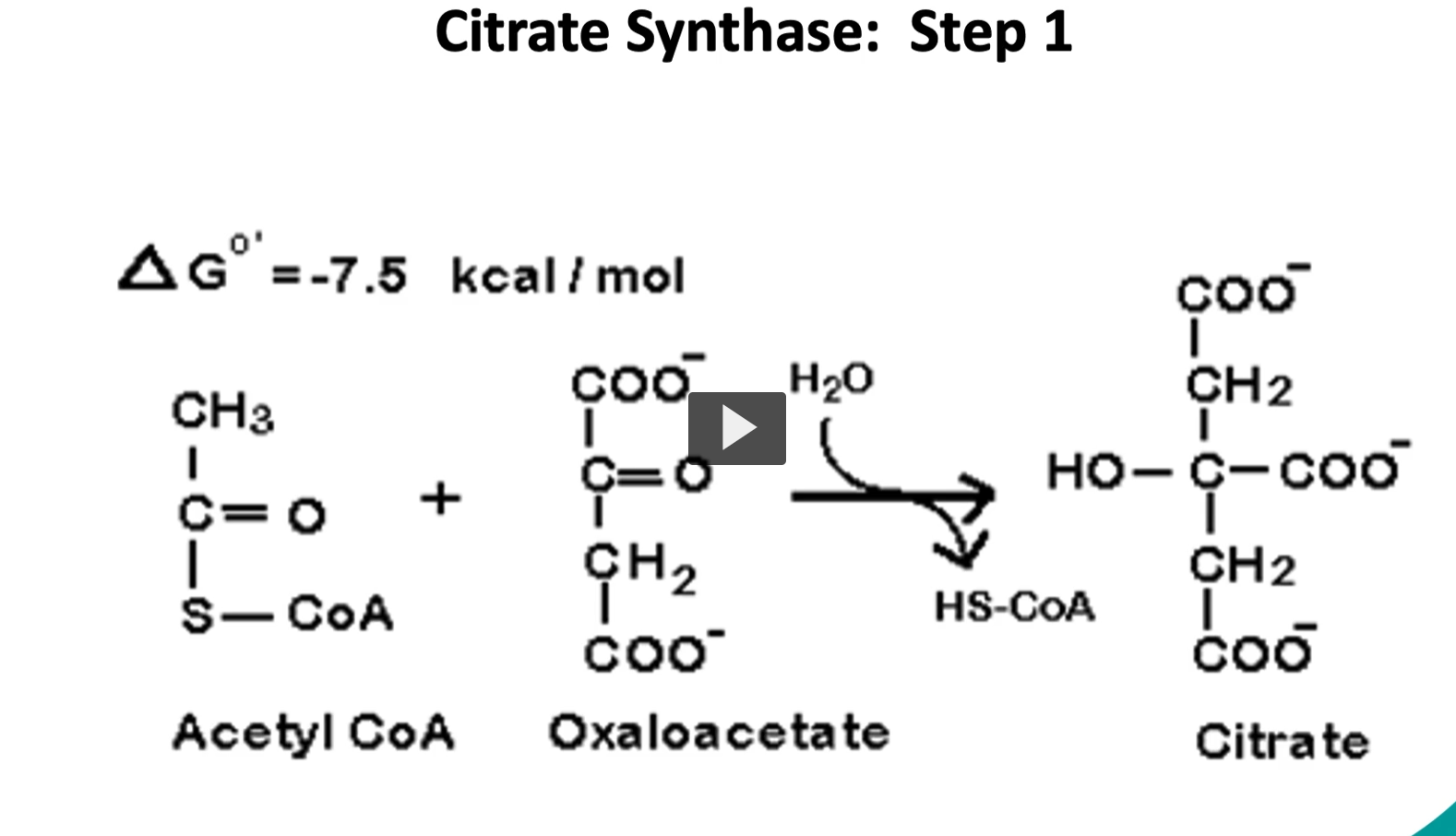
first step of the krebs cycle
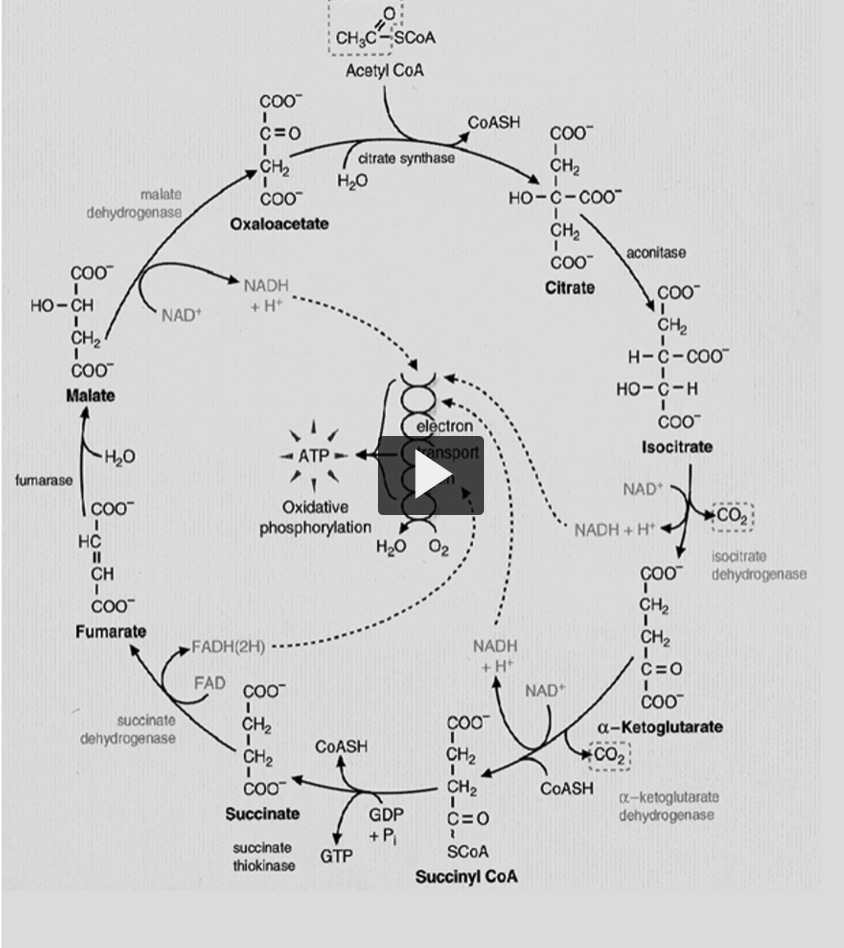
going step by step into the Kreb’s cycle, we start with the substrate of acetyl-CoA and oxaloacetate, the enzyme is called citrate synthase. through our condensation reaction, we will produce citrate. This is an exergonic reaction with a delta G of -7.5 Kcal/mole. This is an irreversible reaction or controlled reaction.
Acetyl-CoA + Oxaloacetate + H2O→ Citrate+ CoA-SH
Enzyme: Citrate synthase
Acetyl-CoA = Coenzyme A carrying a 2-carbon acetyl group, It is the main fuel molecule entering the Krebs cycle.
Oxaloacetate = a 4-carbon molecule related to oxalo and acetate groups
A 2-carbon acetyl group joins a 4-carbon oxaloacetate to form a 6-carbon citrate.
3. Water (H₂O)
Water participates in the reaction because the enzyme performs a hydrolysis step.
Water helps break the thioester bond between acetyl and CoA.
This releases CoA-SH.
4. CoA-SH Etymology
SH
sulfhydryl group
thiol group
When acetyl leaves, CoA regains its free sulfhydryl (-SH).
So: CoA-SH = free coenzyme A
Citrate
Derived from citric acid
citrus fruits contain citric acid
first isolated from lemons
Citrate is a 6-carbon molecule formed when:
2C+4C=6C2C + 4C = 6C2C+4C=6C
Acetyl group + oxaloacetate → citrate.
Citrate is the first molecule of the Krebs cycle.
But citrate has a hydroxyl group in the wrong position, so it must be rearranged before oxidation can occur in the cycle.
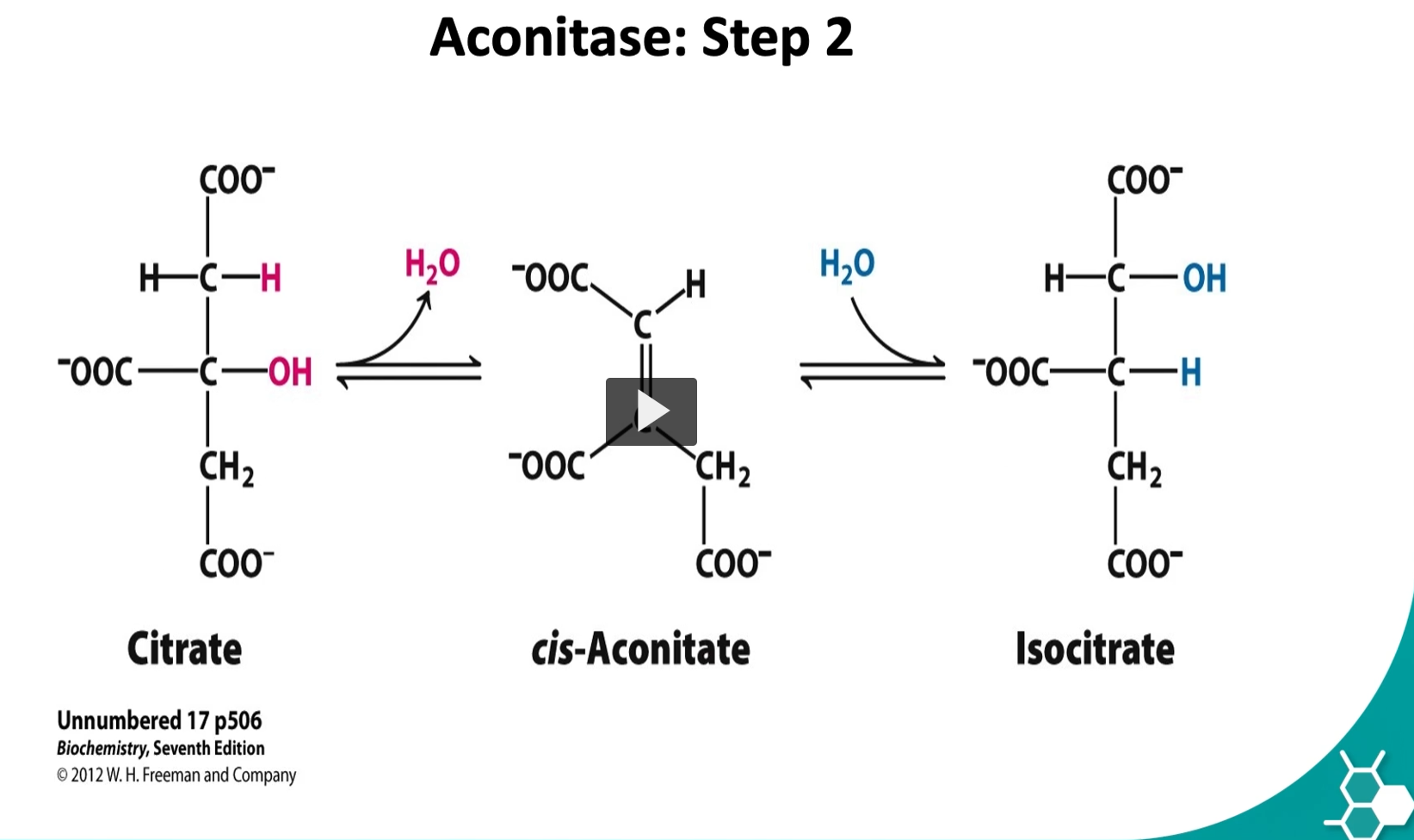
the second step is a reversible one. Here we have a conversion of citrate → iso-citrate by aconitase enzyme.
2. Aconitase Etymology
aconitic acid → a molecule related to citrate that contains a double bond
-ase → enzyme
Aconitase = the enzyme that converts citrate through the aconitate intermediate.
3. Cis-aconitate Etymology
aconitate → derived from aconitic acid, a compound with a carbon–carbon double bond
cis → Latin “on the same side”
Cis-aconitate is a temporary intermediate formed when water is removed from citrate.
4. Isocitrate Etymology
iso- = equal / rearranged
citrate = citric acid derivative
Isocitrate = a rearranged form of citrate.
5. What actually happens in Step 2
The enzyme aconitase performs two reactions:
Step A — Dehydration
Water is removed:
Citrate→cis-aconitate+H2O\text{Citrate} \rightarrow \text{cis-aconitate} + H_2OCitrate→cis-aconitate+H2O
This creates a double bond.
Step B — Rehydration
Water is added back, but in a different orientation:
cis-aconitate+H2O→Isocitrate\text{cis-aconitate} + H_2O \rightarrow \text{Isocitrate}cis-aconitate+H2O→Isocitrate
This moves the –OH group to a different carbon.
6. Why this step is important
Citrate cannot easily undergo oxidation.
Rearranging it to isocitrate puts the hydroxyl group in the correct position for the next step:
Isocitrate→α-ketoglutarate
This step produces NADH and CO₂.
![<p>The aconitase enzyme has an iron sulfur center forming an inorganic molecule, this molecule is bound to cysteine.</p><p>This kinds of molecules are seen in redox reactions as explained in the lecture of oxidative phosphorylation.</p><p>this case is an exception where it supports substrate binding. The importance of knowing this fact is knowing there are external compounds that can affect this binding.</p><p><u>1. What the iron–sulfur center is </u></p><p><strong>Aconitase contains an iron–sulfur cluster</strong> called a <strong>[4Fe–4S] cluster</strong>.</p><p>This cluster is made of:</p><ul><li><p><strong>4 iron atoms (Fe)</strong></p></li><li><p><strong>4 sulfur atoms (S)</strong></p></li></ul><p>These atoms form an <strong>inorganic metal cluster</strong> inside the enzyme.</p><p><u>2. Why cysteine is involved </u></p><p>The cluster is <strong>held in place by cysteine residues</strong> from the protein.</p><p>Cysteine contains a <strong>sulfur atom (-SH group)</strong>.</p><p>The sulfur of cysteine binds to the iron atoms in the cluster.</p><p>So the enzyme structure looks conceptually like:</p><pre><code>Cysteine-S → Fe
Cysteine-S → Fe
Cysteine-S → Fe
</code></pre><p>These cysteine residues <strong>anchor the iron–sulfur cluster inside the enzyme</strong>.</p><p><u>3. Iron–sulfur clusters in many enzymes </u></p><p>Iron–sulfur clusters are common in <strong>redox enzymes</strong>, especially in:</p><ul><li><p><strong>electron transport chain proteins</strong></p></li><li><p><strong>oxidative phosphorylation enzymes</strong></p></li></ul><p>In those systems, Fe–S clusters <strong>transfer electrons</strong>.</p><p>Example: <span>Fe2+</span><span data-name="left_right_arrow" data-type="emoji">↔</span><span>Fe3+</span></p><p>This allows them to participate in <strong>oxidation–reduction reactions</strong>.</p><p><u>4. Why aconitase is special </u></p><p>In <strong>aconitase</strong>, the Fe–S cluster is <strong>not mainly transferring electrons</strong>.</p><p>Instead, it helps <strong>bind the citrate molecule and orient it properly</strong>.</p><p>So here the cluster functions as a <strong>structural catalytic center</strong>, helping the enzyme:</p><ul><li><p>bind citrate</p></li><li><p>remove water</p></li><li><p>add water back to form isocitrate.</p></li></ul><p><u>5. Why external compounds can affect this </u></p><p>Because the enzyme relies on the <strong>iron–sulfur cluster</strong>, substances that bind iron or sulfur can <strong>inactivate the enzyme</strong>.</p><p>A classic example is <strong>fluoroacetate poisoning</strong>.</p><p>Fluoroacetate becomes <strong>fluorocitrate</strong>, which inhibits aconitase.</p><p>This blocks the <strong>Krebs cycle</strong>, stopping cellular energy production.</p><p><u>6. Why this matters biologically </u></p><p>If aconitase stops working:</p><ul><li><p>citrate cannot become isocitrate</p></li><li><p>the Krebs cycle stops</p></li><li><p>ATP production drops dramatically.</p></li></ul><p>This is why some toxins targeting this enzyme are <strong>very dangerous</strong>.</p>](https://knowt-user-attachments.s3.amazonaws.com/f4e7c4f9-945f-4e1e-9382-eb432959cf55.png)
The aconitase enzyme has an iron sulfur center forming an inorganic molecule, this molecule is bound to cysteine.
This kinds of molecules are seen in redox reactions as explained in the lecture of oxidative phosphorylation.
this case is an exception where it supports substrate binding. The importance of knowing this fact is knowing there are external compounds that can affect this binding.
1. What the iron–sulfur center is
Aconitase contains an iron–sulfur cluster called a [4Fe–4S] cluster.
This cluster is made of:
4 iron atoms (Fe)
4 sulfur atoms (S)
These atoms form an inorganic metal cluster inside the enzyme.
2. Why cysteine is involved
The cluster is held in place by cysteine residues from the protein.
Cysteine contains a sulfur atom (-SH group).
The sulfur of cysteine binds to the iron atoms in the cluster.
So the enzyme structure looks conceptually like:
Cysteine-S → Fe
Cysteine-S → Fe
Cysteine-S → Fe
These cysteine residues anchor the iron–sulfur cluster inside the enzyme.
3. Iron–sulfur clusters in many enzymes
Iron–sulfur clusters are common in redox enzymes, especially in:
electron transport chain proteins
oxidative phosphorylation enzymes
In those systems, Fe–S clusters transfer electrons.
Example: Fe2+↔Fe3+
This allows them to participate in oxidation–reduction reactions.
4. Why aconitase is special
In aconitase, the Fe–S cluster is not mainly transferring electrons.
Instead, it helps bind the citrate molecule and orient it properly.
So here the cluster functions as a structural catalytic center, helping the enzyme:
bind citrate
remove water
add water back to form isocitrate.
5. Why external compounds can affect this
Because the enzyme relies on the iron–sulfur cluster, substances that bind iron or sulfur can inactivate the enzyme.
A classic example is fluoroacetate poisoning.
Fluoroacetate becomes fluorocitrate, which inhibits aconitase.
This blocks the Krebs cycle, stopping cellular energy production.
6. Why this matters biologically
If aconitase stops working:
citrate cannot become isocitrate
the Krebs cycle stops
ATP production drops dramatically.
This is why some toxins targeting this enzyme are very dangerous.
the third step is known as the rate limiting step of the cycle. It is very spontaneous and irreversible reaction.
Step 3 of the Krebs cycle (isocitrate → α-ketoglutarate) is called the rate-limiting step and is considered essentially irreversible because of the large release of energy and the loss of CO₂, which drives the reaction strongly forward.
isocitrate → oxalosuccinate → α-ketoglutarate
with NAD⁺ becoming NADH and CO₂ being released.
Big picture
This step is catalyzed by:
isocitrate dehydrogenase
Overall reaction:
Isocitrate+NAD+→α-ketoglutarate+CO2+NADH
1. Start with isocitrate
Isocitrate is a 6-carbon molecule.
It has an alcohol group (-OH) on the carbon that will be oxidized.
That alcohol is important, because alcohols can be oxidized into carbonyls.
2. NAD⁺ becomes NADH
The enzyme removes 2 hydrogens / electrons from isocitrate.
So:
isocitrate is oxidized
NAD⁺ is reduced to NADH
Reaction idea:
Isocitrate→Oxalosuccinate
while
NAD+→NADH
So NADH is formed because it accepts the electrons taken from isocitrate
3. Oxalosuccinate forms
This first oxidation converts the alcohol on isocitrate into a ketone.
That gives the intermediate:
oxalosuccinate
Etymology
oxalo- = oxidized / oxalo-type acid relation
succinate = related to succinic acid
Meaning:
It is a more oxidized 6-carbon intermediate between isocitrate and α-ketoglutarate.
This intermediate is usually not isolated because it quickly moves to the next step.
4. Oxalosuccinate gives up CO₂
Now oxalosuccinate is a beta-keto acid–type intermediate, and those are unstable.
So it undergoes decarboxylation:
Oxalosuccinate→α-ketoglutarate+CO2
This means:
one carbon leaves as CO₂
the molecule goes from 6 carbons to 5 carbons
5. α-Ketoglutarate forms
After losing CO₂, the remaining 5-carbon molecule is:
α-ketoglutarate
Etymology
alpha (α) = ketone is on the carbon next to the carboxyl group
keto- = ketone group
glutarate = 5-carbon dicarboxylic acid backbone
So α-ketoglutarate is a 5-carbon keto acid.
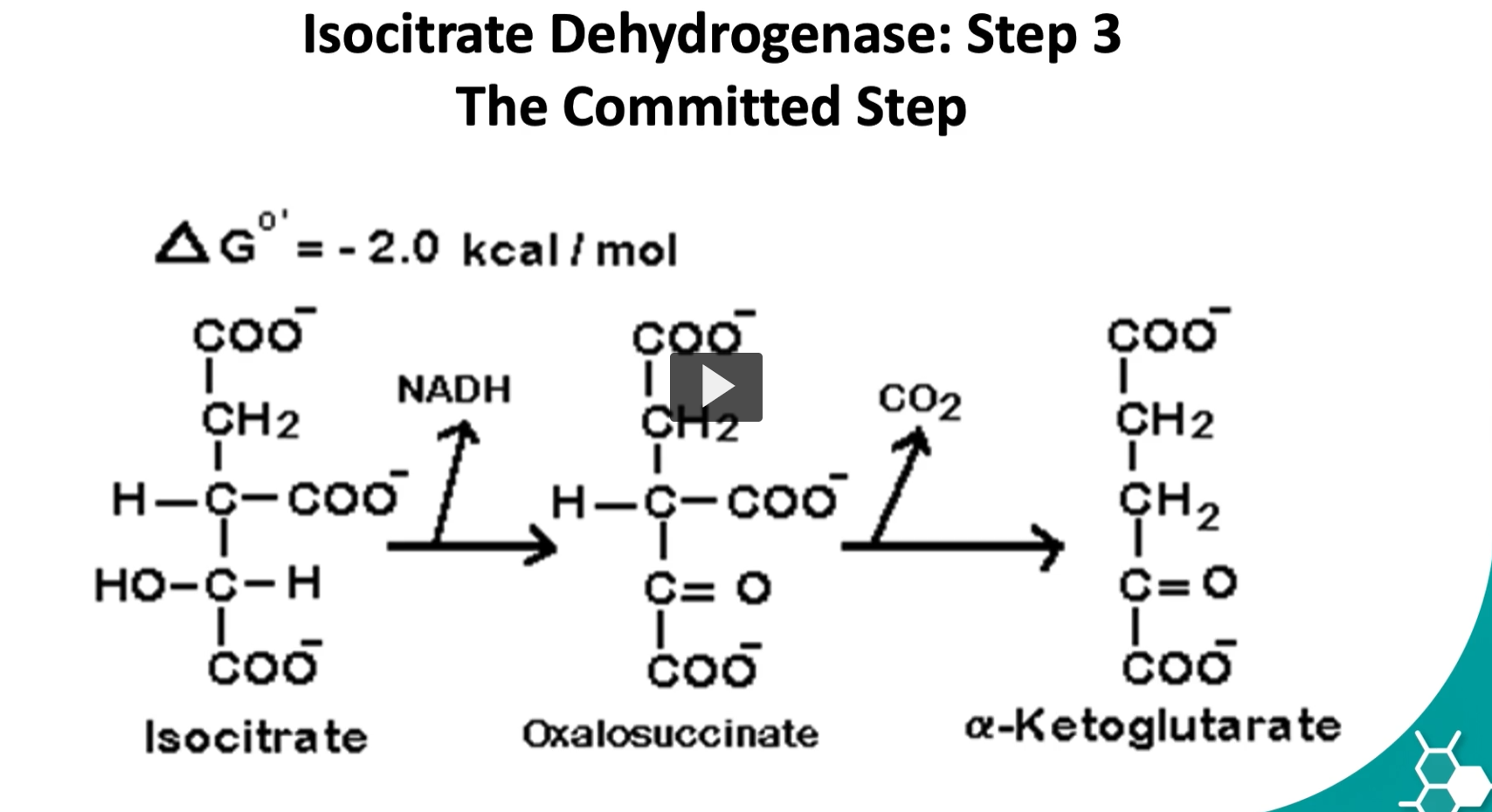
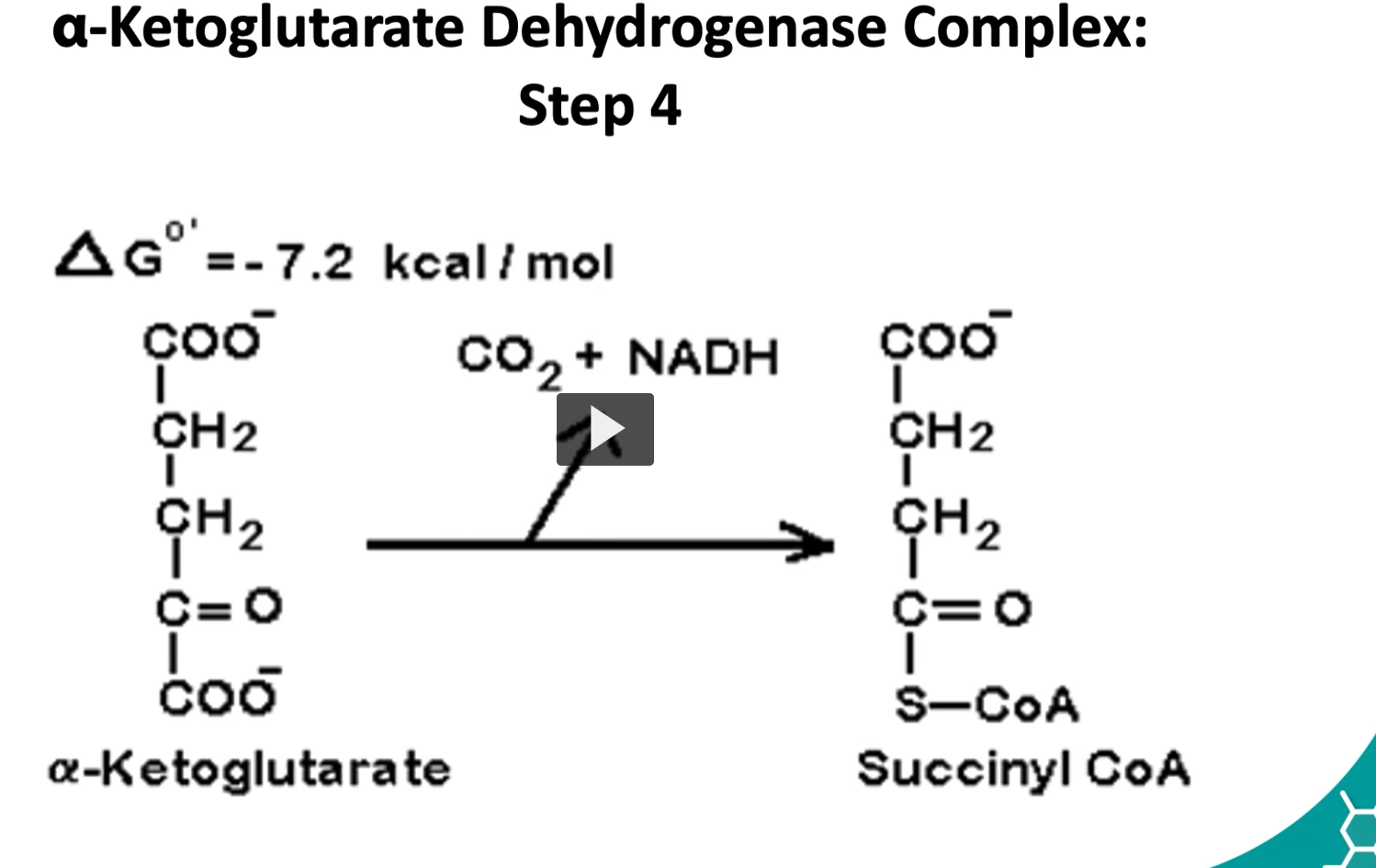
the fourth step is where alpha ketolglutarate is converted to succinyl CoA by a multisymatic complex
alpha ketoglutarate, CO2, NADH, Succinyl-CoA
Here we have an oxidative decarboxylation and the addition of coenzyme A.
The delta G is -7.2 Kcal/mole.
This is a controls step as well.
The Reaction (Step 4 of the Krebs Cycle)
α-ketoglutarate + CoA + NAD+→Succinyl-CoA+CO2+NADH
Enzyme complex: α-ketoglutarate dehydrogenase complex
Step-by-step puzzle
1. Start: α-ketoglutarate (5 carbons)
α-ketoglutarate is a 5-carbon molecule produced in Step 3.
Etymology:
alpha (α) → ketone is on the first carbon next to the carboxyl group
keto → ketone group (C=O)
glutarate → a 5-carbon dicarboxylic acid
So it is a 5-carbon keto acid.
2. Oxidation occurs
α-ketoglutarate loses electrons.
Those electrons are transferred to NAD⁺.
NAD+→NADH
So:
α-ketoglutarate is oxidized
NAD⁺ is reduced to NADH
3. Decarboxylation occurs
A carboxyl group (-COO⁻) is removed.
This releases: CO2
4. Coenzyme A attaches
After CO₂ leaves, the remaining 4-carbon succinyl group attaches to Coenzyme A (CoA-SH).
This forms: Succinyl-CoA
Succinyl-CoA contains a high-energy thioester bond, which will be used in the next step to produce GTP (ATP equivalent).
Why the reaction releases energy
The ΔG is:
−7.2 kcal/mol-7.2 \text{ kcal/mol}−7.2 kcal/mol
This means the reaction releases energy because:
oxidation releases energy
CO₂ leaves the system
formation of a high-energy thioester bond
This makes the reaction essentially irreversible.
Why this is a control step
The enzyme α-ketoglutarate dehydrogenase is regulated by the cell’s energy state.
Inhibited by
ATP
NADH
Succinyl-CoA (product inhibition)
Activated by
Ca²⁺ (especially in muscle)
So the cycle slows when the cell already has enough energy.
Similar to the pyruvate dehydrogenase complex, the alpha ketoglutarate complex uses 3 enzymes and 5 coenzymes.
the enzyme 1 or E1 is the alpha ketoglutarate dehydrogenase component with the coenzyme TPP or thiamine pyrophosphate.
the enzyme 2 or E2, then transsuccinylase, with two coenzymes with are CoA and lipoamide
the enzyme 3 or E3, is dihydrolipoyl dehydrogenase, which uses NAD and FAD as coenzymes
1. Enzyme 1 (E1): α-ketoglutarate dehydrogenase
Cofactor: TPP (Thiamine pyrophosphate)
What happens
α-ketoglutarate (5 carbons) binds the enzyme.
CO₂ is removed (decarboxylation).
The remaining 4-carbon fragment attaches to TPP.
Intermediate: Succinyl-TPP
So the molecule goes from:
5C→4C+CO2
2. Enzyme 2 (E2): Dihydrolipoyl transsuccinylase
Cofactors:
Lipoamide
Coenzyme A (CoA)
What happens
The succinyl group is transferred from TPP to lipoamide.
Lipoamide carries the group like a swinging arm.
The succinyl group is transferred to CoA.
Result: Succinyl-CoA
This molecule contains a high-energy thioester bond.
3. Enzyme 3 (E3): Dihydrolipoyl dehydrogenase
Cofactors:
FAD
NAD⁺
What happens
During step 2, lipoamide becomes reduced.
E3 restores it so the cycle can continue.
Steps:
Reduced lipoamide transfers electrons → FAD
FAD becomes FADH₂
FADH₂ transfers electrons → NAD⁺
NAD⁺ becomes NADH
Now the enzyme is reset
The 5 Cofactors (same as pyruvate dehydrogenase)
A common mnemonic:
TLCFN
Cofactor | Function |
|---|---|
TPP | removes CO₂ |
Lipoamide | carries the succinyl group |
CoA | forms succinyl-CoA |
FAD | accepts electrons |
NAD⁺ | final electron carrier |

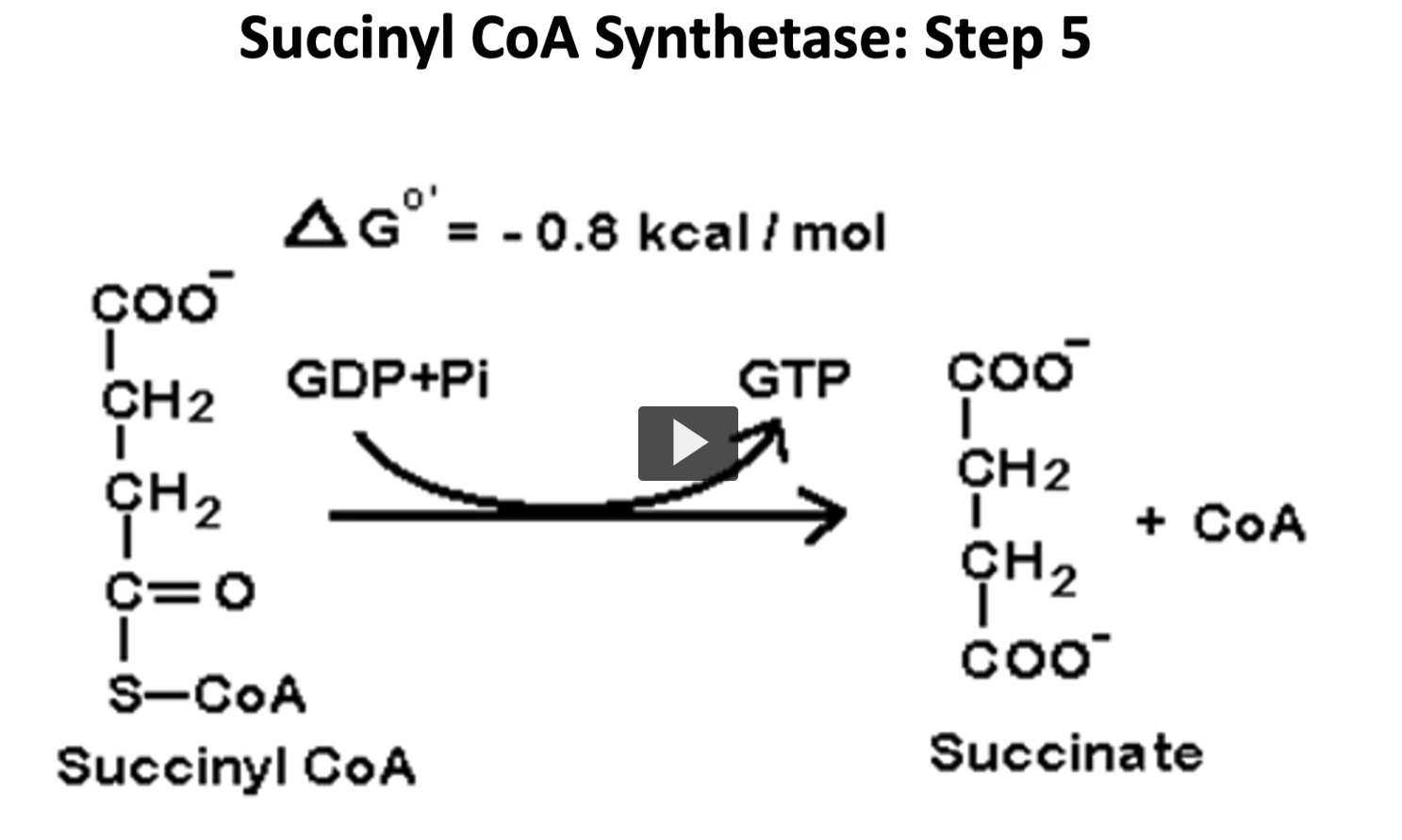
the fifth step is a reversible reaction in which succinyl coa is converted to succinate, and GTP is formed by substrate level phosphorylation.
this GTP is then converted to ATP by the nucleoside triphosphate kinase.
succinyl Coa, GDP + Pi, GTP, succinate
The Reaction (Step 5)
Succinyl-CoA +GDP +Pi → Succinate + GTP + CoA−SH
Enzyme: Succinyl-CoA synthetase (also called succinate thiokinase)
This reaction uses the energy of the thioester bond in succinyl-CoA to make GTP.
1. Succinyl-CoA Etymology
Succinyl
from succinic acid
Latin succinum = amber
(succinic acid was first isolated from amber)
-yl
chemical suffix meaning derived group
So succinyl = the acyl group derived from succinic acid.
CoA
Coenzyme A
co- = helper
enzyme = assists enzymes
A originally referred to acetyl transfer
CoA carries acyl groups using a sulfhydryl (-SH).
Succinyl-CoA = a 4-carbon succinyl group attached to coenzyme A
It contains a high-energy thioester bond.
Use
The energy of that bond is used to produce GTP.
2. GDP Etymology
G = guanine (nitrogenous base)
D = di (two)
P = phosphate
So: GDP = guanosine diphosphate
Structure:
guanine base
ribose sugar
2 phosphates
Use
GDP receives a phosphate to become GTP
3. Pi Meaning
Pi = inorganic phosphate
Etymology:
P = phosphate
i = inorganic
Use
Pi provides the phosphate that converts GDP → GTP.
4. GTP Etymology
G = guanine
T = tri (three)
P = phosphate
GTP = guanosine triphosphate
Use
GTP is an energy molecule, similar to ATP.
It can:
power some cellular reactions
or be converted into ATP.
5. Succinate Etymology
Succinate
From succinic acid (amber acid).
Succinate is a 4-carbon Krebs cycle intermediate.
It continues to the next step of the cycle.
6. Substrate-level phosphorylation Etymology
substrate
the molecule being acted upon in the reaction.
phosphorylation
phospho = phosphate
-ylation = adding a group
A phosphate group is directly transferred to GDP to make GTP.
This is different from oxidative phosphorylation.
6. Substrate-level phosphorylation Etymology
substrate the molecule being acted upon in the reaction.
phosphorylation
phospho = phosphate
-ylation = adding a group
Meaning A phosphate group is directly transferred to GDP to make GTP.
This is different from oxidative phosphorylation.
7. Nucleoside triphosphate kinase Etymology
nucleoside: base + sugar
triphosphate: three phosphates
kinase: enzyme that transfers a phosphate
Reaction GTP+ADP→GDP+ATP
Meaning: The enzyme transfers the phosphate from GTP to ADP, producing ATP
Putting the whole puzzle together
Step-by-step:
Succinyl-CoA has a high-energy thioester bond.
Breaking that bond releases energy.
That energy adds Pi to GDP, forming GTP.
CoA is released.
The remaining molecule becomes succinate.
GTP can then transfer its phosphate to ADP → ATP.
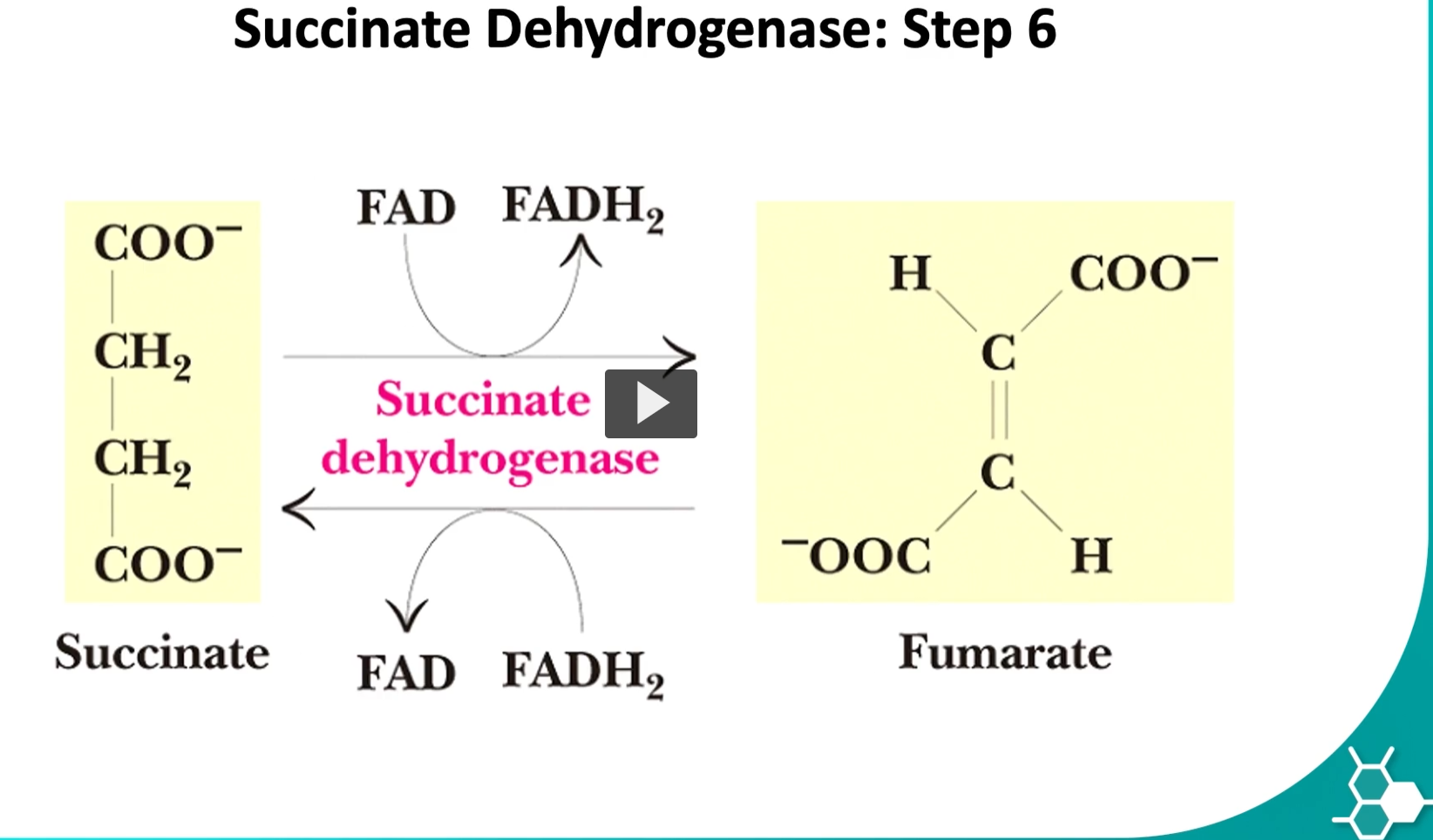
the sixth step is where succinate is converted to fumarate by the enzyme called succinate dehydrogenase
this is the only step where FAD will be oxidized and produce FADH2.
the enzyme is bound in the inner membrane of the mitochondria.
the reason is this enzyme participates in kreb’s cycle but also in oxidative phosphorylation.
succinate, FAD, FADH2, succinate dehydrogenase, fumarate
You are describing Step 6 of the Krebs (TCA) cycle, where succinate is oxidized to fumarate and FAD is reduced to FADH₂. This step is unique because the enzyme is also part of the electron transport chain (Complex II).
Let’s break down the etymology, meaning, and role of each term, and then put the biochemical puzzle together.
The Reaction (Step 6)
Succinate+FAD→Fumarate+FADH2
Enzyme: Succinate dehydrogenase
1. Succinate Etymology
Succinate comes from succinic acid.
Latin succinum = amber
Succinic acid was first isolated from amber resin
The suffix -ate means the ion or salt of an acid.
Meaning
Succinate is a 4-carbon molecule in the Krebs cycle.
Structure conceptually:
HOOC–CH₂–CH₂–COO⁻
Use Succinate is the substrate that gets oxidized in Step 6.
During the reaction, it loses two hydrogens and becomes fumarate.
2. FAD Etymology
FAD = Flavin Adenine Dinucleotide
Breakdown: Flavin
from Latin flavus = yellow
flavin molecules are yellow pigments
Adenine
a nitrogenous base in nucleotides
Dinucleotide
two nucleotides linked together.
Meaning
FAD is a coenzyme that accepts electrons and hydrogen atoms.
Use In this step: FAD→FADH2
It captures 2 hydrogens and 2 electrons from succinate.
3. FADH₂ Etymology
Same root as FAD, but with H₂ meaning two hydrogens added.
Meaning
FADH₂ = the reduced form of FAD
It now carries high-energy electrons.
Use FADH₂ transfers those electrons to the electron transport chain, producing ATP.
4. Succinate Dehydrogenase Etymology
Succinate the substrate molecule.
Dehydrogenase
de- = remove
hydrogen
-ase = enzyme
Meaning: An enzyme that removes hydrogen from succinate.
Meaning: This enzyme removes two hydrogens from succinate, transferring them to FAD.
Use It catalyzes: Succinate→Fumarate
while producing FADH₂
5. Fumarate Etymology
Fumarate comes from fumaric acid.
Latin fumus = smoke
Fumaric acid was first isolated from fumitory plants.
Meaning Fumarate is another 4-carbon Krebs cycle intermediate.
Structure: HOOC–CH=CH–COO⁻
Notice the double bond between the carbons.
Use Fumarate continues into the next step of the Krebs cycle: Fumarate→Malate
Why this step is special
1. Oxidation occurs
Succinate loses hydrogen atoms.
Succinate→Fumarate
2. FAD is reduced
FAD accepts the electrons and hydrogens: FAD→FADH2
3. Enzyme location
Succinate dehydrogenase is embedded in the inner mitochondrial membrane.
This is unique because all other Krebs cycle enzymes are in the mitochondrial matrix.
4. Link to oxidative phosphorylation
Succinate dehydrogenase is also:
Complex II of the electron transport chain.
So this enzyme participates in both:
Pathway | Role |
|---|---|
Krebs cycle | oxidizes succinate |
Electron transport chain | transfers electrons to ubiquinone |
Putting the puzzle together
Step-by-step:
Succinate loses two hydrogens.
The molecule forms a double bond, becoming fumarate.
FAD accepts the hydrogens, becoming FADH₂.
The electrons from FADH₂ enter the electron transport chain.
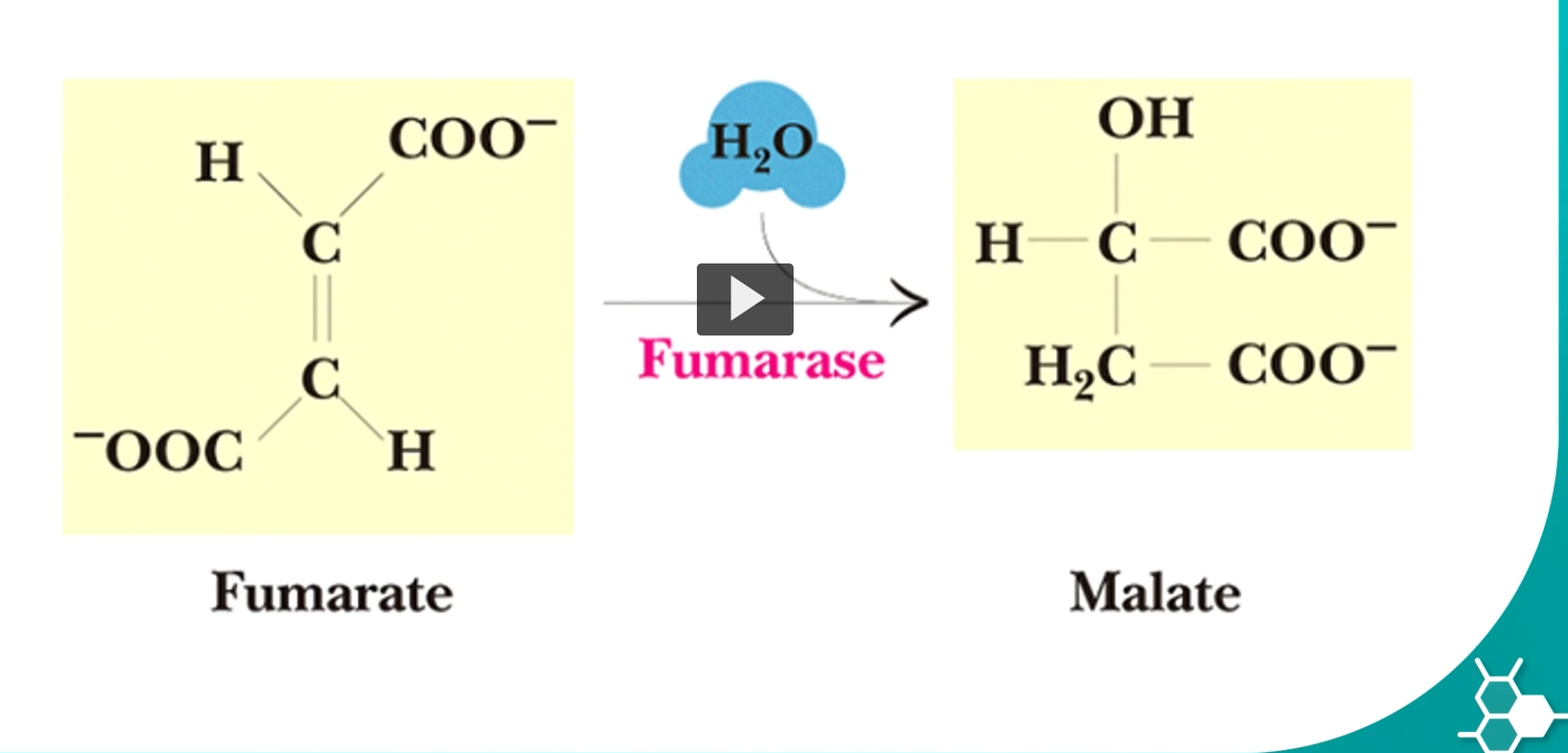
step 7 will convert fumarate into malate, by using fumarase enzyme
this hydration reaction can be a reversible reaction. the malate is known as the L-malate.
Malate comes from malic acid.
Latin malum = apple
Malic acid was first found in apple juice.
Suffix -ate
the ionized form of malic acid.
Malate is a 4-carbon molecule with a hydroxyl group (-OH).
Structure conceptually:
HOOC–CH₂–CHOH–COO⁻
Use Malate continues into the final step of the Krebs cycle, where it will be oxidized to:
Oxaloacetate
producing NADH.
The enzyme fumarase adds water in a very specific orientation.
This produces only the L-isomer of malate.
Etymology:
L
refers to Levo configuration, describing the spatial orientation of the molecule.
Only L-malate can be used in the next Krebs cycle reaction.
What actually happens in the reaction
The double bond in fumarate is broken.
Water adds across the bond:
OH group attaches to one carbon
H attaches to the other carbon
Result: Fumarate+H2O→L-malate
Putting the puzzle together
Step-by-step:
Fumarate has a carbon-carbon double bond.
Fumarase adds water across that double bond.
The double bond becomes a single bond.
A hydroxyl group forms, producing L-malate.
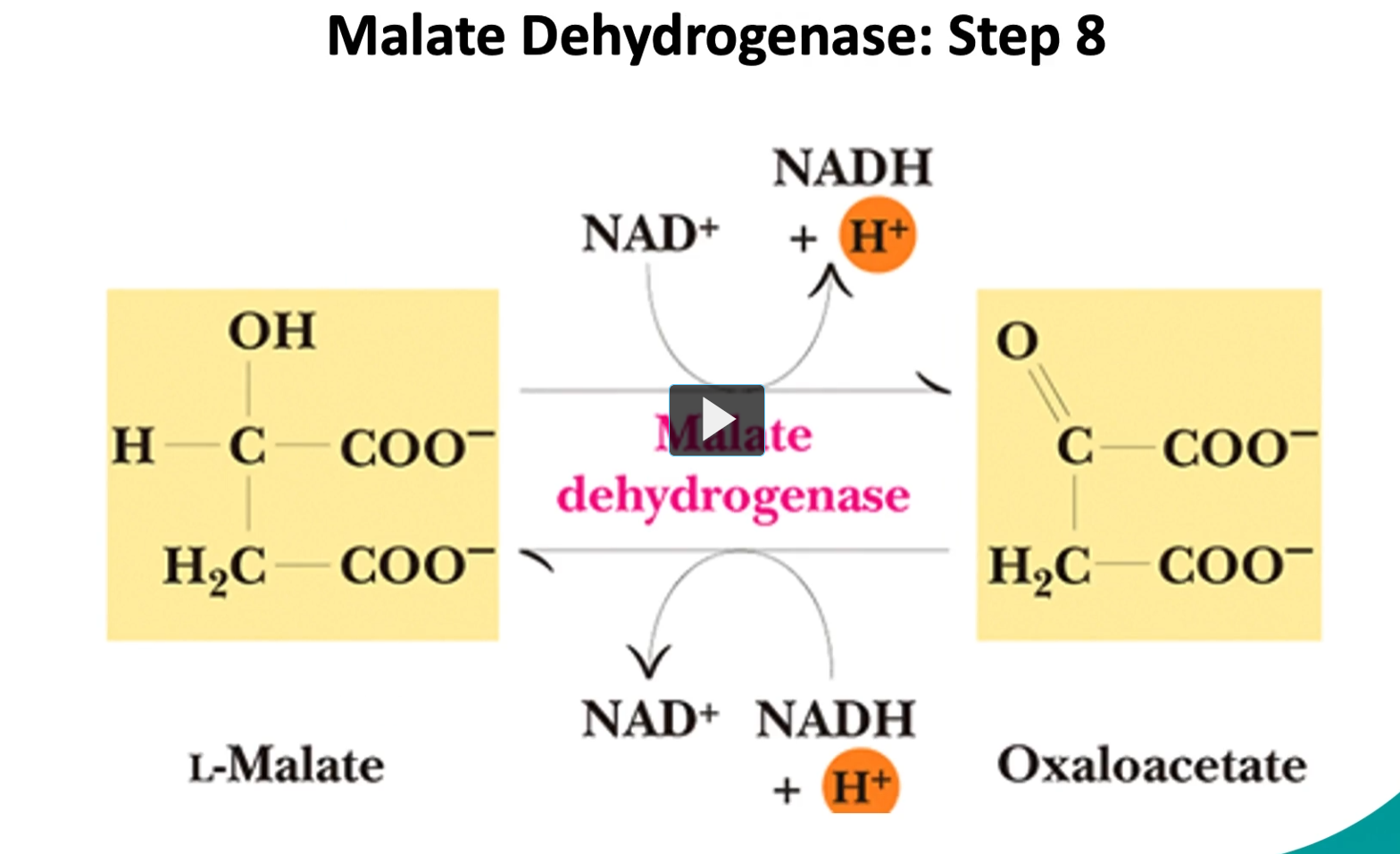
the final step (step 8), is the conversion of the L-malate into oxaloacetate by malate dehydrogenase.
it is reversible as well.
The Reaction (Step 8)
L-malate + NAD+→Oxaloacetate + NADH+H
Enzyme: Malate dehydrogenase
Type of reaction: Oxidation (dehydrogenation)
This reaction is reversible, although under normal conditions it proceeds forward because oxaloacetate is quickly used in the next step of the cycle.
Step-by-step:
L-malate contains a hydroxyl group (-OH).
The enzyme malate dehydrogenase removes hydrogens.
Those electrons go to NAD⁺, forming NADH + H⁺.
The alcohol group becomes a carbonyl (C=O).
The molecule becomes oxaloacetate.
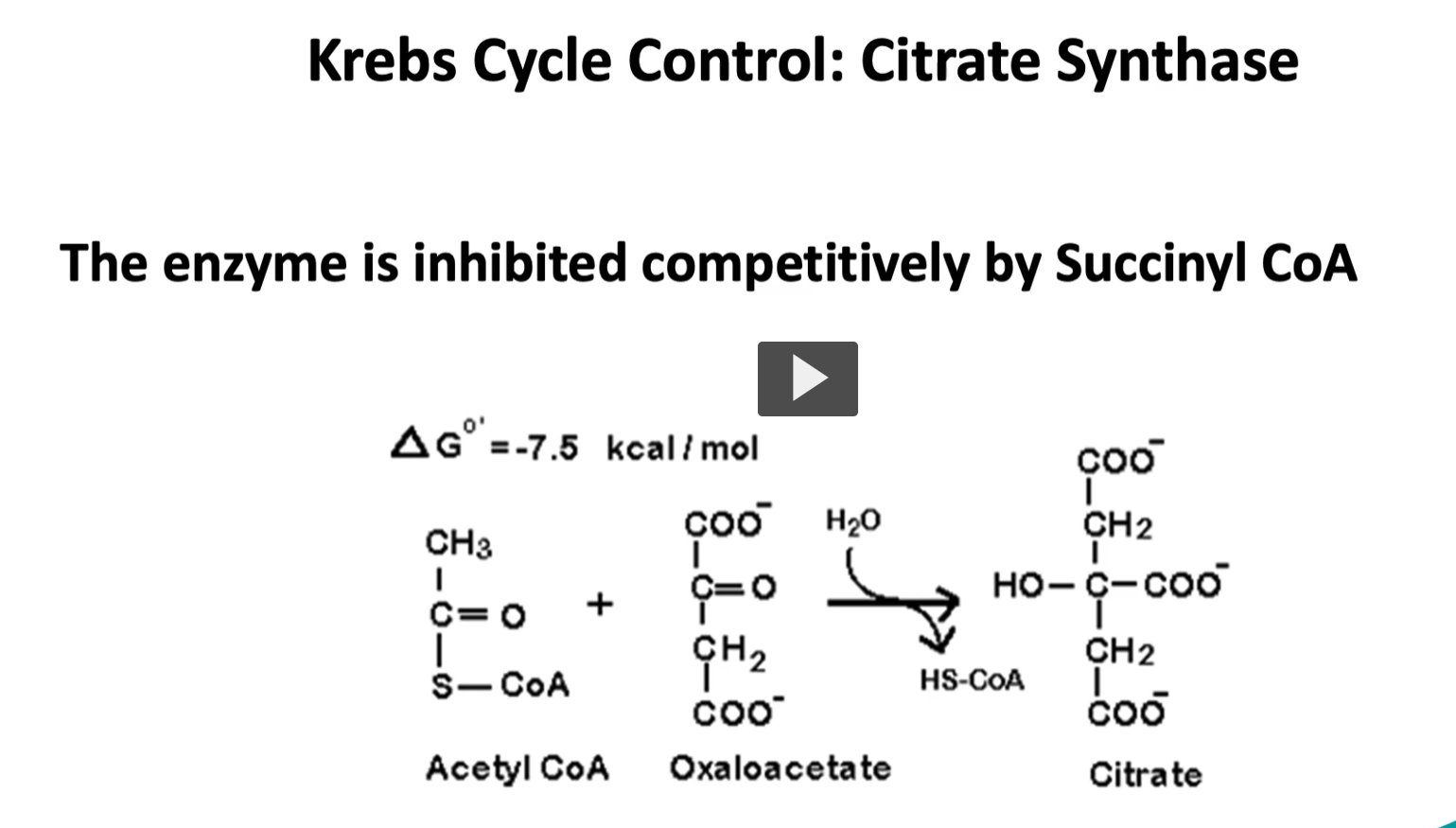
control points in Kreb cycle
to summarize, there are three control points in the Kreb cycle.
the first one is the first step of the cycle, where the enzyme is citrate synthase is controlled by the level of succinyl-CoA, where a high amount will inhibit in a competitively manner.
the second one, pacemaker, is isocitrate dehydrogenase. (step 3)
this enzyme catalyzes the committed step, the enzyme can be inhibited allostetically, by high concentration of ATP and ADH.
vice versa, if ADP and NADH are high, then the enzyme is activated.
Why this step is called the “pacemaker”
It is considered the main regulatory step of the Krebs cycle because it determines how fast the cycle runs.
When this enzyme is active → the cycle runs faster.
When it is inhibited → the cycle slows.

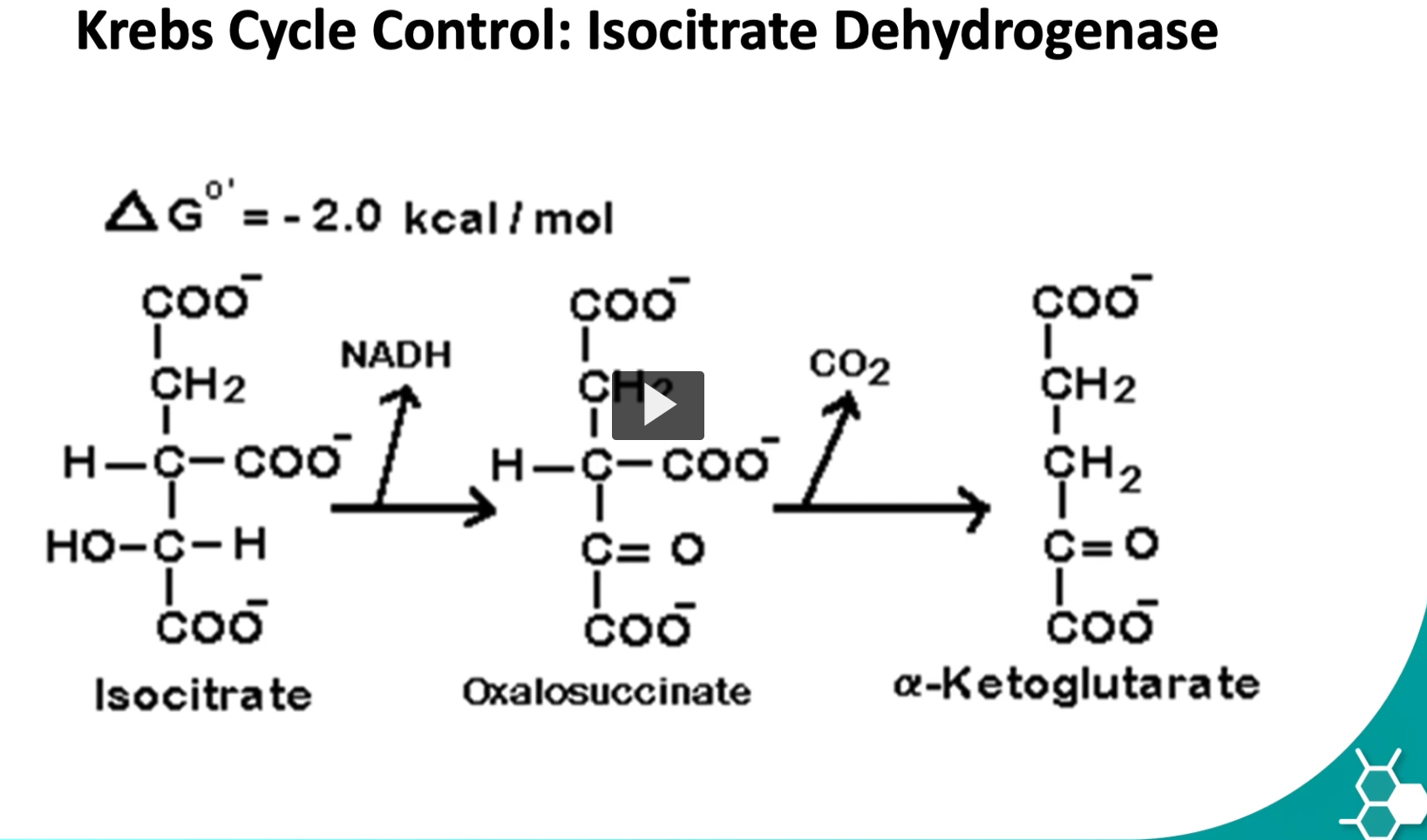
the last control point is in the step where isocitrate is converted to alpha ketoglutarate by isocitrate dehydrogenase.
remember that this step is requires a multienzymatic complex
steps 1 and 3 are the control points in the Kreb’s cycle.
as we discussed before, the multi-enzymatic complex is regulated by three enzymes, where enzyme one, alpha ketoglutarate dehydrogenase complex with a cofactor TPP is inhibited allosterically by high concentration of ATP activated by ADP.


enzyme 2, transuccinylase with cofactors as coenzyme A lipoamide is inihibited allosterically by high concentrations of succinyl-CoA
enzyme 3, dihydrolipoyl dehydrogenase with cofactor of FAD and NAD is inhibited by allosterically by high concentration of NADH

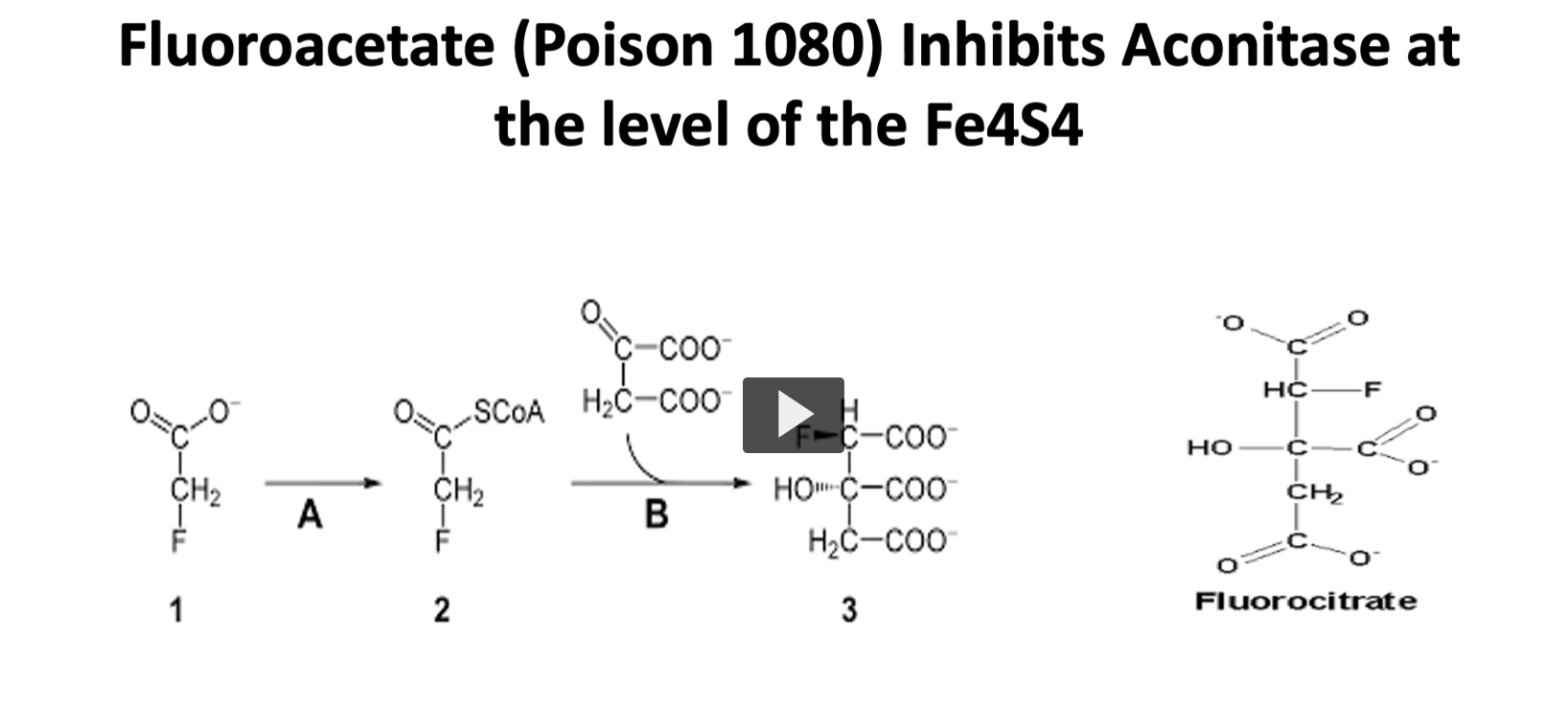
a part of the inhibition of the kreb’s cycle we just mentioned, there is a compound called fluoroacetate (poison 1080) inhibits aconitase at the level of Fe4S4
if you recall in the step two of the Kreb’s cycle, the enzyme is aconitase. this enzyme is inhibited at the level of cofactor with the iron sulfur center. this inhibitions is not direct. what happens is that fluoroacetate is used as a substrate instead of acetate, which is converted to fluorocitrate, which is a very electronegative atom.
the floor of fluorocitrate binds tightly to aconitase, this is a very tight bond, so the enzyme is unable to work in the Kreb’s cycle.
remember, that the Kreb’s cycle is important in the central nervous system, and if there is a high concentration of this compound, it will lead to the death of the organism it is exposed to.
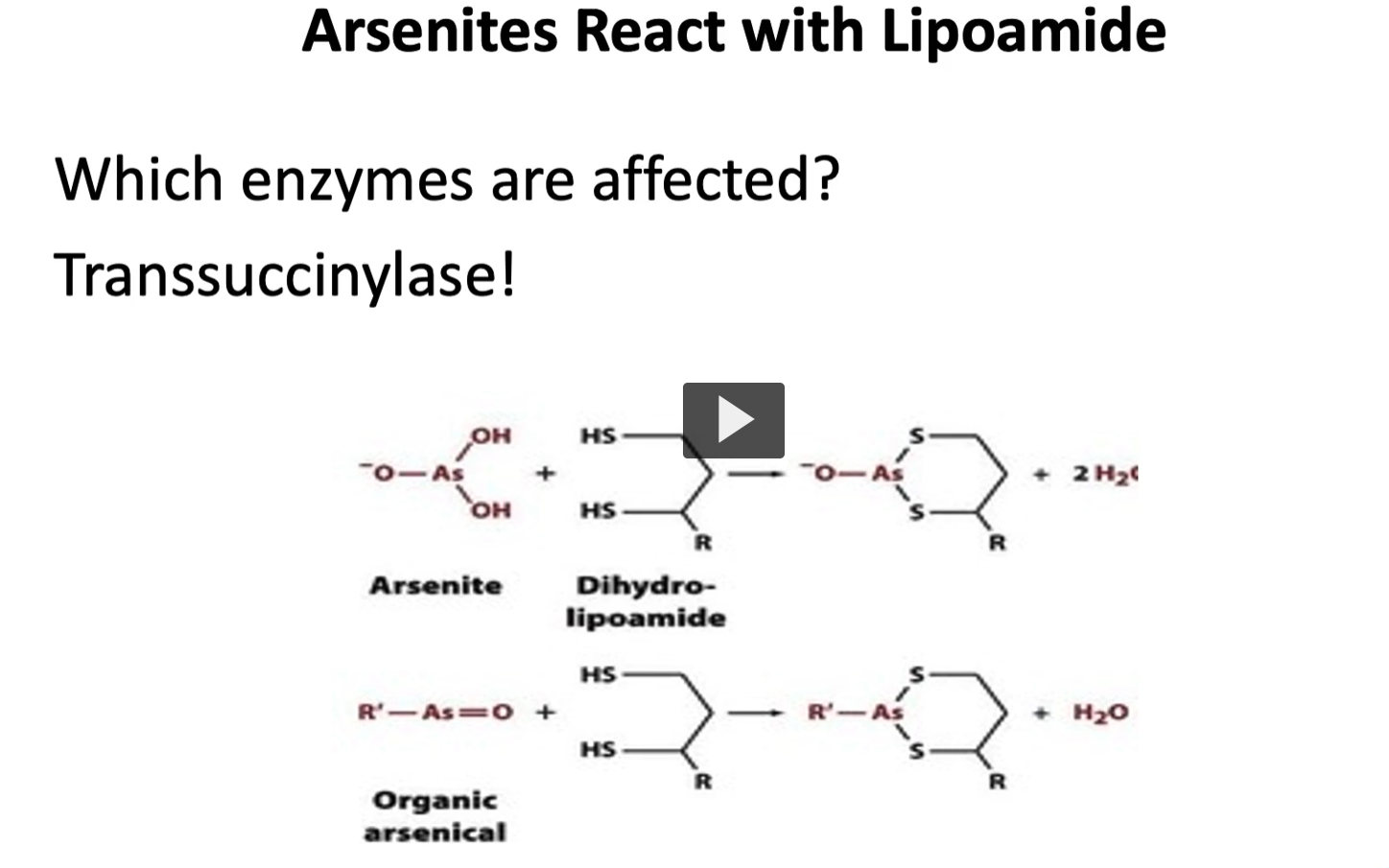
lastly, we have arsenites, which react with cofactor lipoamide.
we talk about alpha ketoglutarate dehydrogenase complex
arsenates will bind and inactivate the coenzyme lipoamide needed, the enzyme that it will affect is transuccinylase

THIRD VIDEO.
krebs cycle = citric acid cycle= TCA cycle


in the previous video, we discussed the pyruvate dehydrogenase complex mechanism and important cofactors.
it is important to remember that one product of the PDC reaction was acetyl-CoA
the acetyl-CoA produced during the PDC reaction will be used during the KREBS cycle
this happens because pyruvate cannot directly enter the Krebs cycle.
the PDC mechanism will ensure that pyruvate is converted to acetyl-CoA, the key substrate for energy production in the krebs cycle.
specifically, in the first step of the krebs cycle, acetyl-CoA combines with oxaloacetate to form citrate, which will start the cycle.
for a little bit more background on the kreb’s cycle, it is also known as the tri-carboxylic acid cycle, or the citric acid cycle.
it is a central metabolic pathway, that occurs in the mitochondrial matrix. as mentioned previously, it’s major key role is to generate energy by oxidizing acetyl-CoA.
this acetyl-CoA can be obtained from multiple sources like carbohydrates, fats, proteins, and, as previously discussed, the PDC.
once oxidized, acetyl-CoA will be converted to carbon dioxide, and with this cycle, electrons are transferred into the carrier molecules.

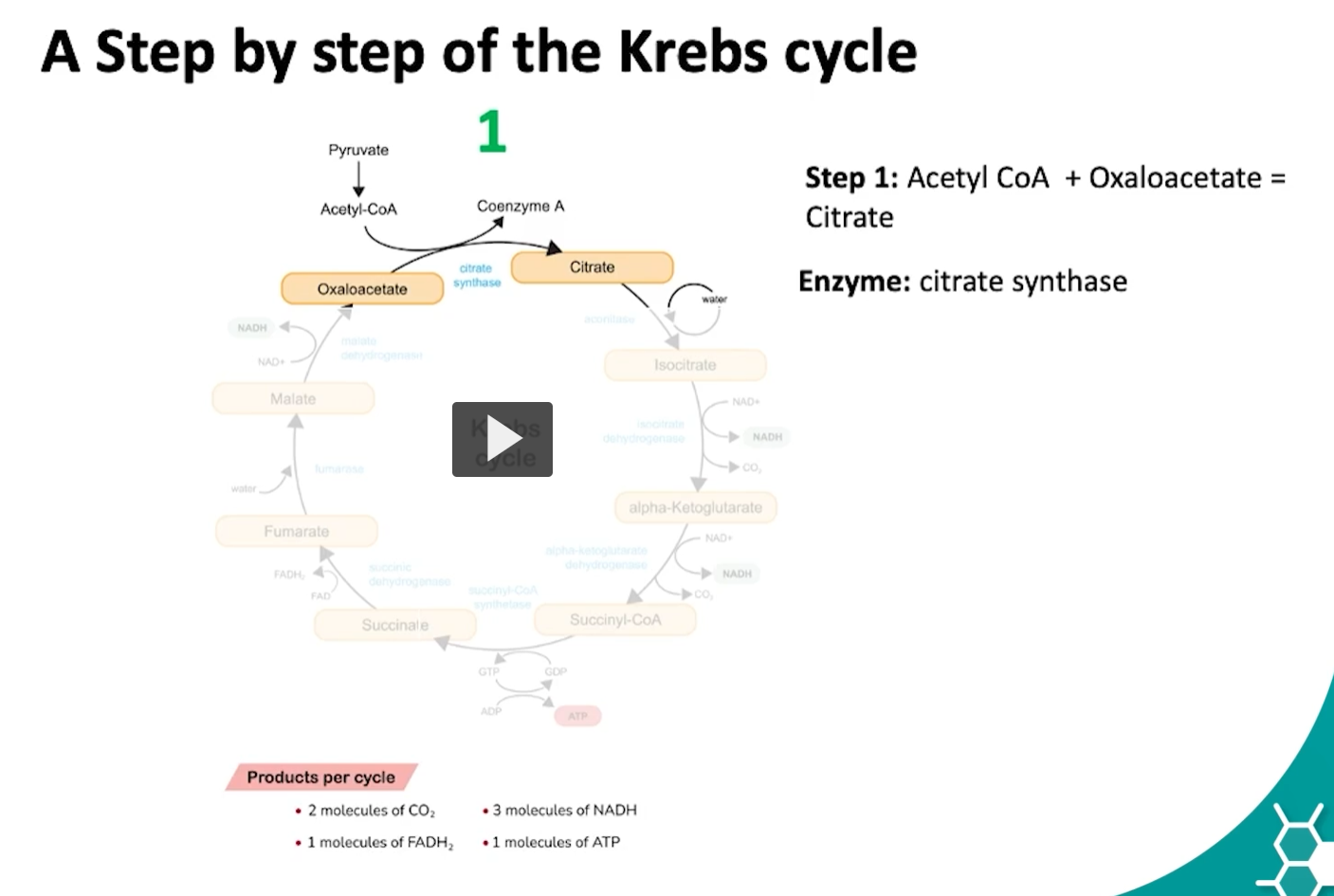
moving onto the steps of the kreb’s cycle.
the first step of the kreb’s cycle begins with acetyl-CoA
the acetyl-CoA, which has two carbons, combines with oxaloacetate, which has four carbons, by the action of citrate synthase, to form citrate, a six carbon intermediate.
what is happening during this reaction is the methyl group of the acetyl-CoA reacts with the carbonyl group of the oxaloacetate, this is known as an aldol condensation followed by a hydrolysis of carbon dioxide.
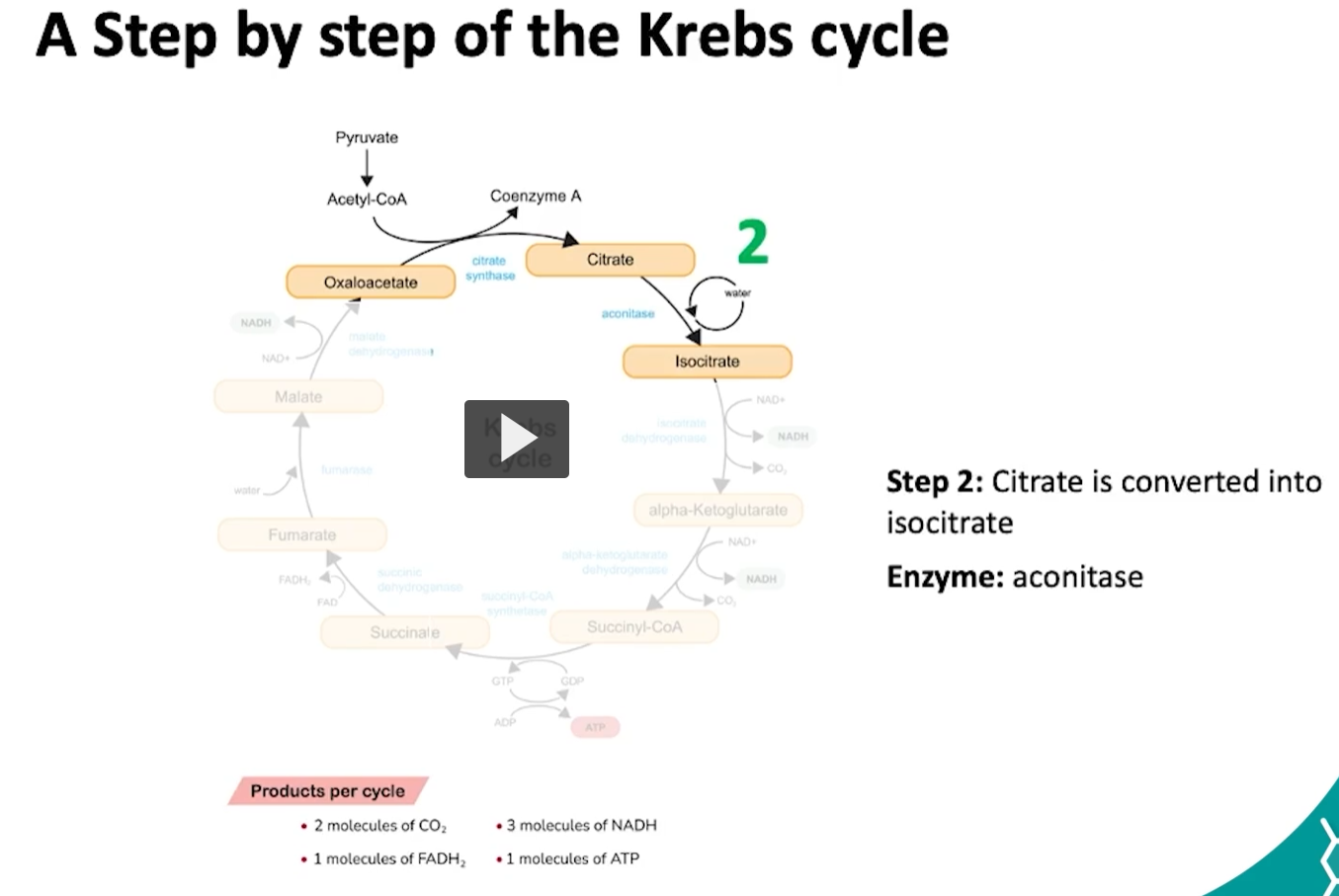
the second step of the krebs cycle is the conversion of citrate to isocitrate. this conversion is done by the aconitase enzyme.
what is happening during this reaction is: the citrate will undergo a dehydration to form a cis aconitase, which is then hydrated into isocitrate.
this ensures that the molecule is more reactive for oxidation.
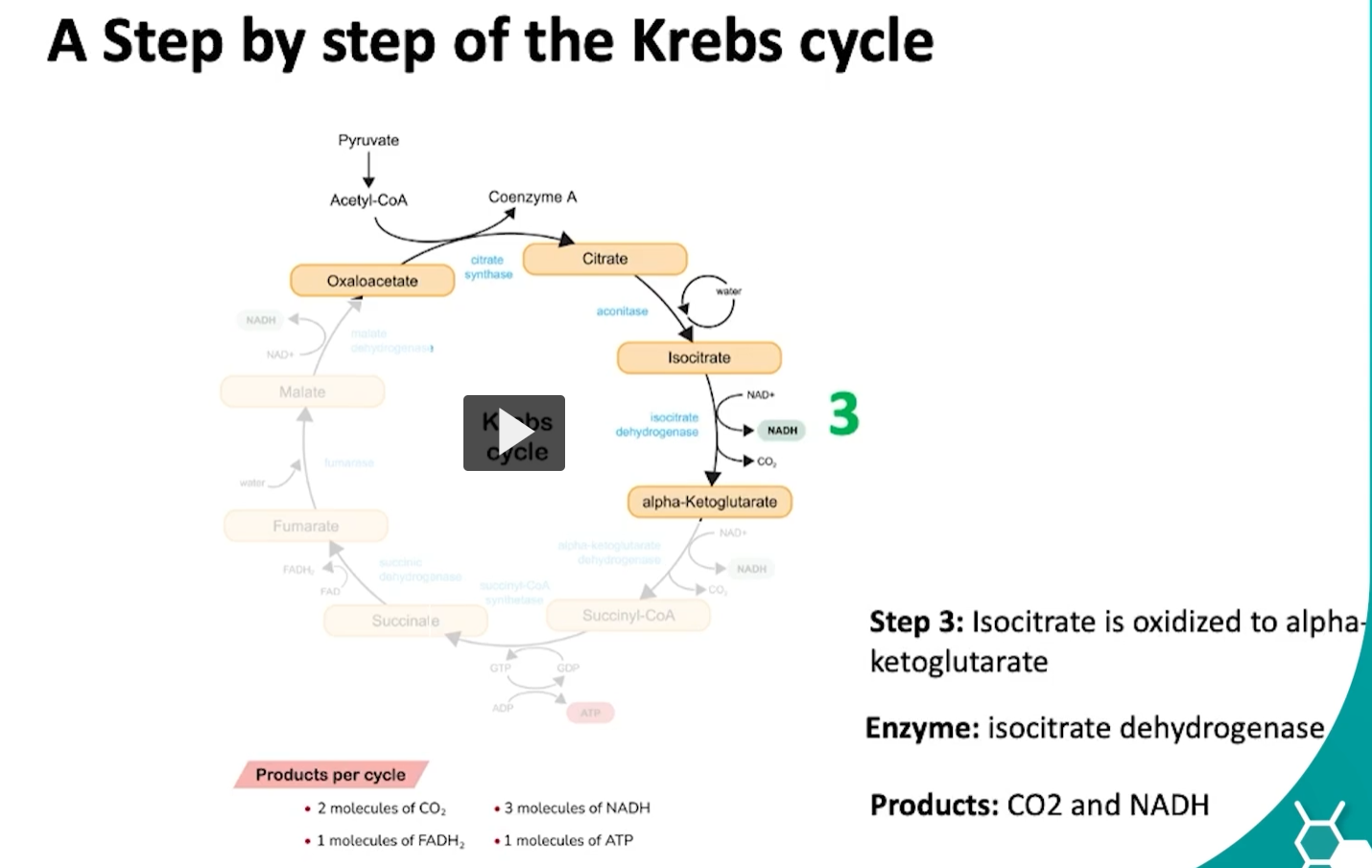
during the third step of the cycle, isocitrate is oxidized into alpha ketoglutarate via the isocitrate dehydrogenase.
what is happening here is the hydroxyl group of isocitrate is oxidized to a carbonyl group forming oxalosuccinate.
then oxalosuccinate undergoes decarboxylation which will release CO2 and form alpha-ketoglutarate.
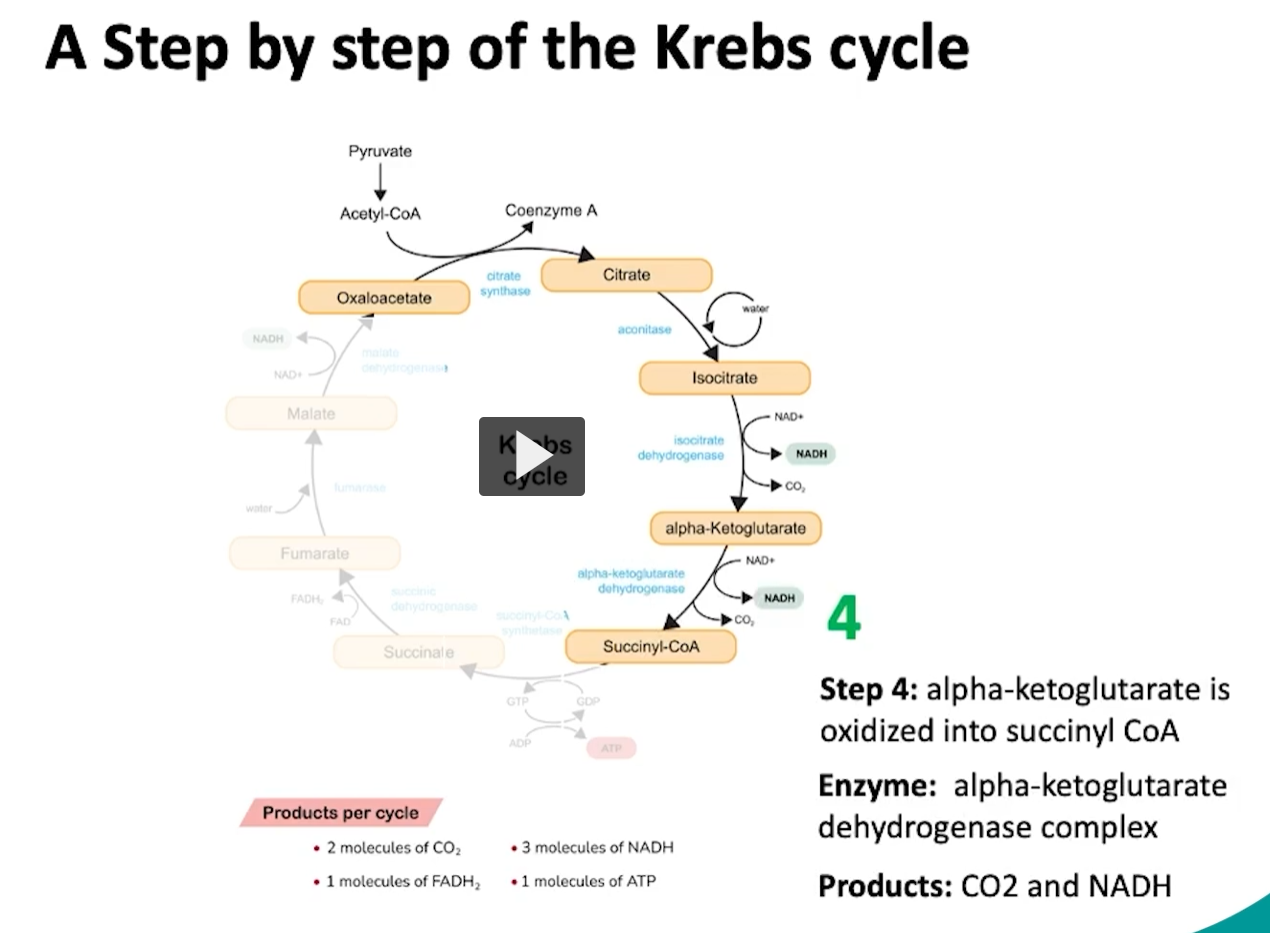
during the fourth step of the cycle, alpha ketoglutarate will be oxidized to form succinyl CoA
what happens here is a decarboxylation or removal of a carbon group of alpha ketoglutarate, which will form succinyl.
then this succinyl is attached or oxidized to coenzyme A via the alpha ketoglutarate enzyme which will form succinyl CoA.
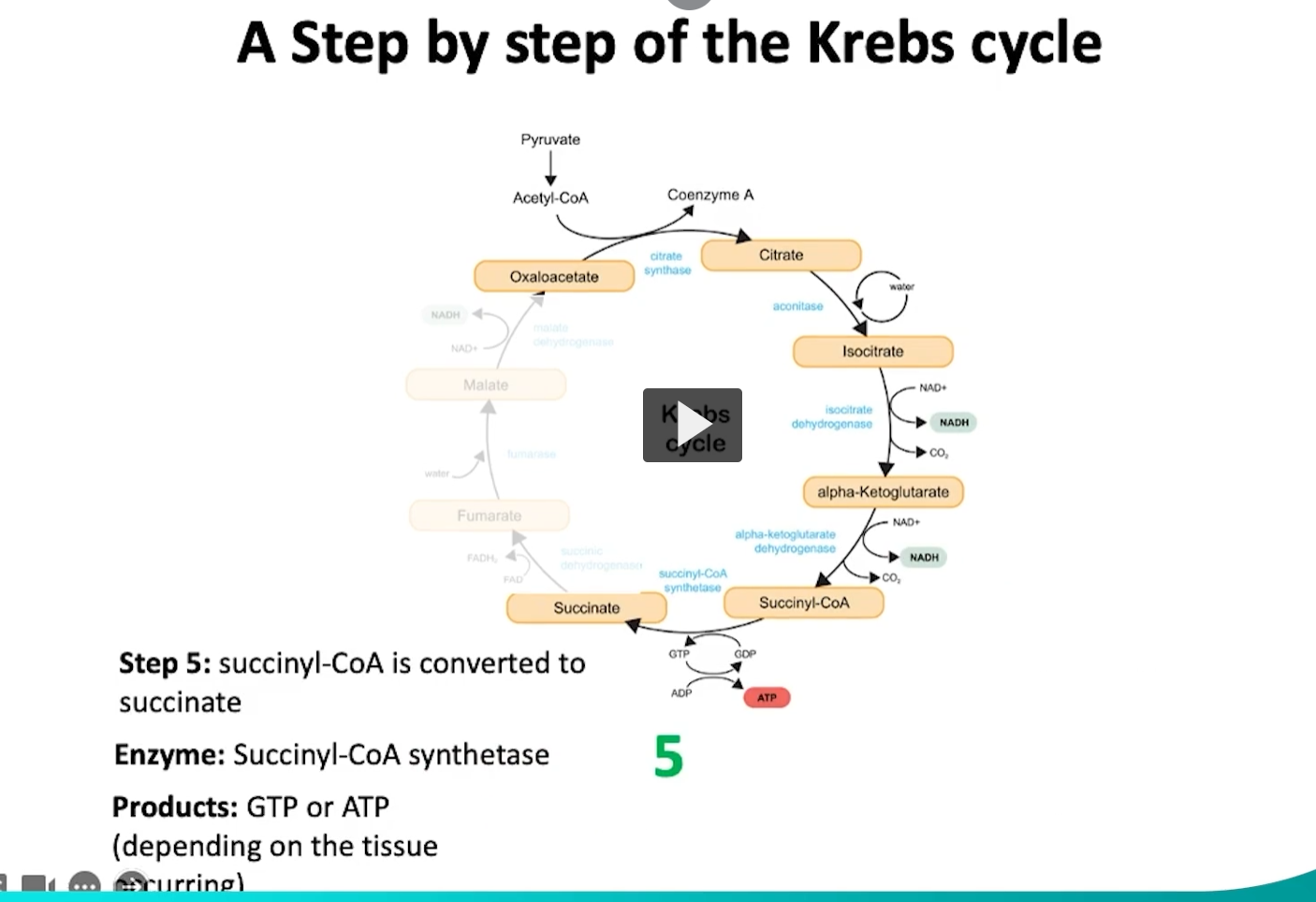
during the fifth step of the krebs cycle, succinyl CoA will converted into succinate.
what is happening in this reaction is that the succinyl CoA synthase will hydrolyze the high energy thioester bond in succinyl CoA.
This hydrolysis releases energy which helps with the ATP or GTP production depending on the tissue.
this step is highly significant given that it is the only one that directly will form ATP or GTP.
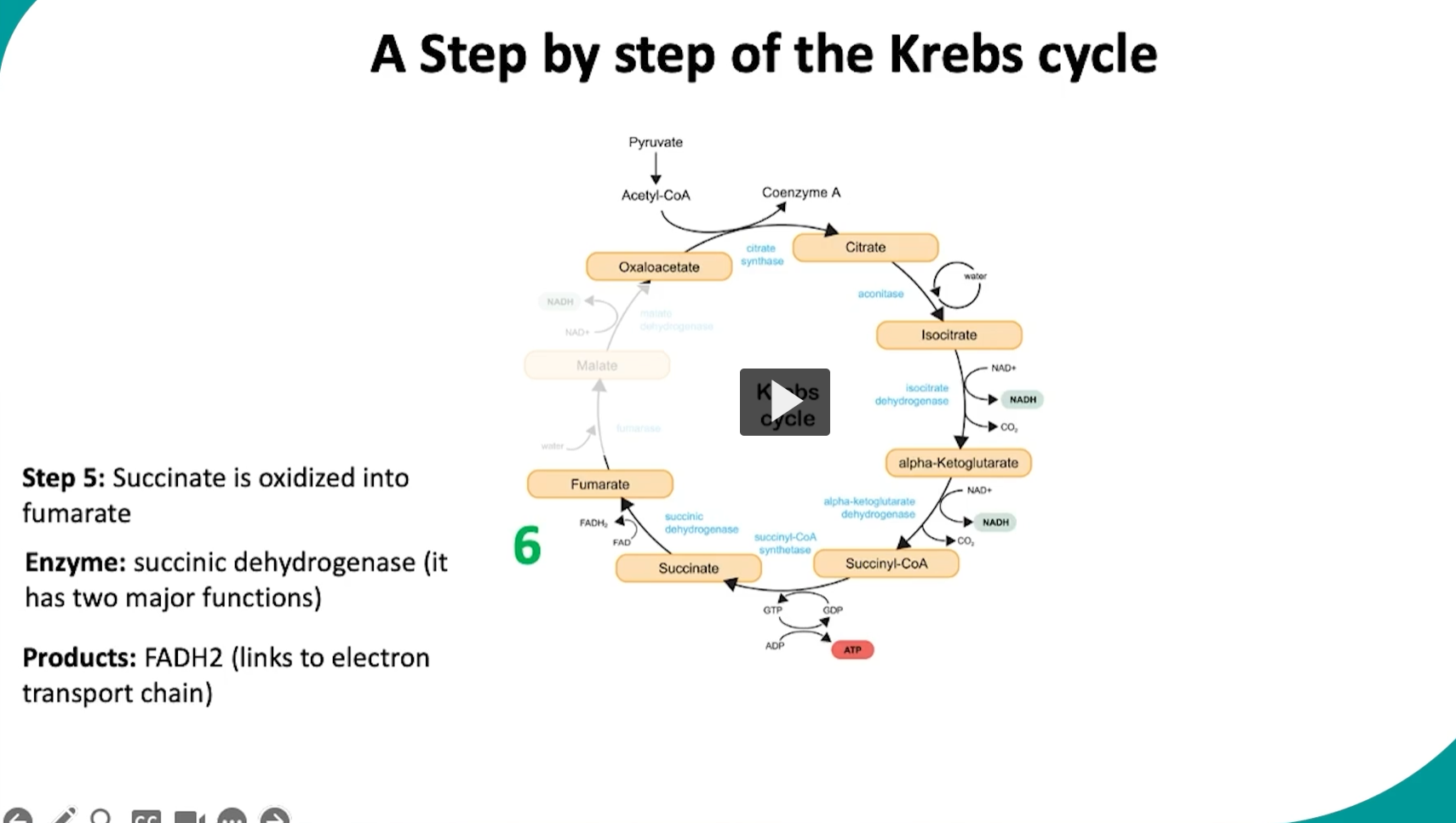
during the sixth step of the cycle, succinate will be oxidized to fumarate via the succinate dehydrogenase
what is happening here is unlike other steps of the Krebs cycle, the succinate dehydrogenase enzyme is located in the inner mitochondrial membrane
this allows the enzyme to have two functions,
1) oxidation of succinate, which means succinate will lose two hydrogen atoms and form fumarate
2) to work as part of the electron transport chain, the two removed hydrogen groups from succinate will be transferred to FAD to produce FADH2. then FADH2 will donate its electron directly to the electron transport chain.
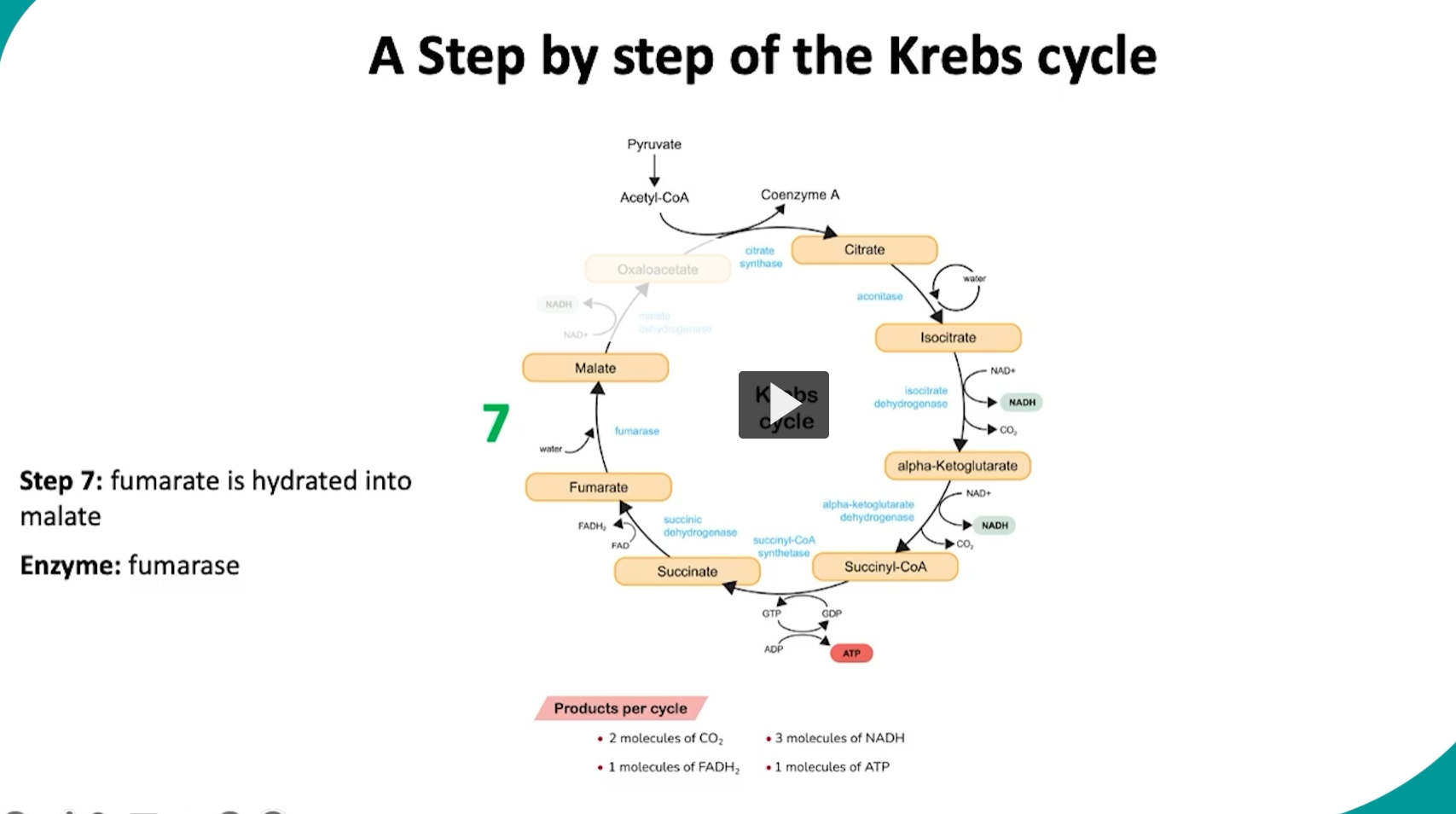
during the seventh step in the cycle, fumarate will be hydrated into malate via the fumerase enzyme.
what is happening in this step is that the fumarase will add a hydroxyl group and a hydrogen atom across the double bond in fumarate.
this step is mainly in preparation for the final oxidation step.
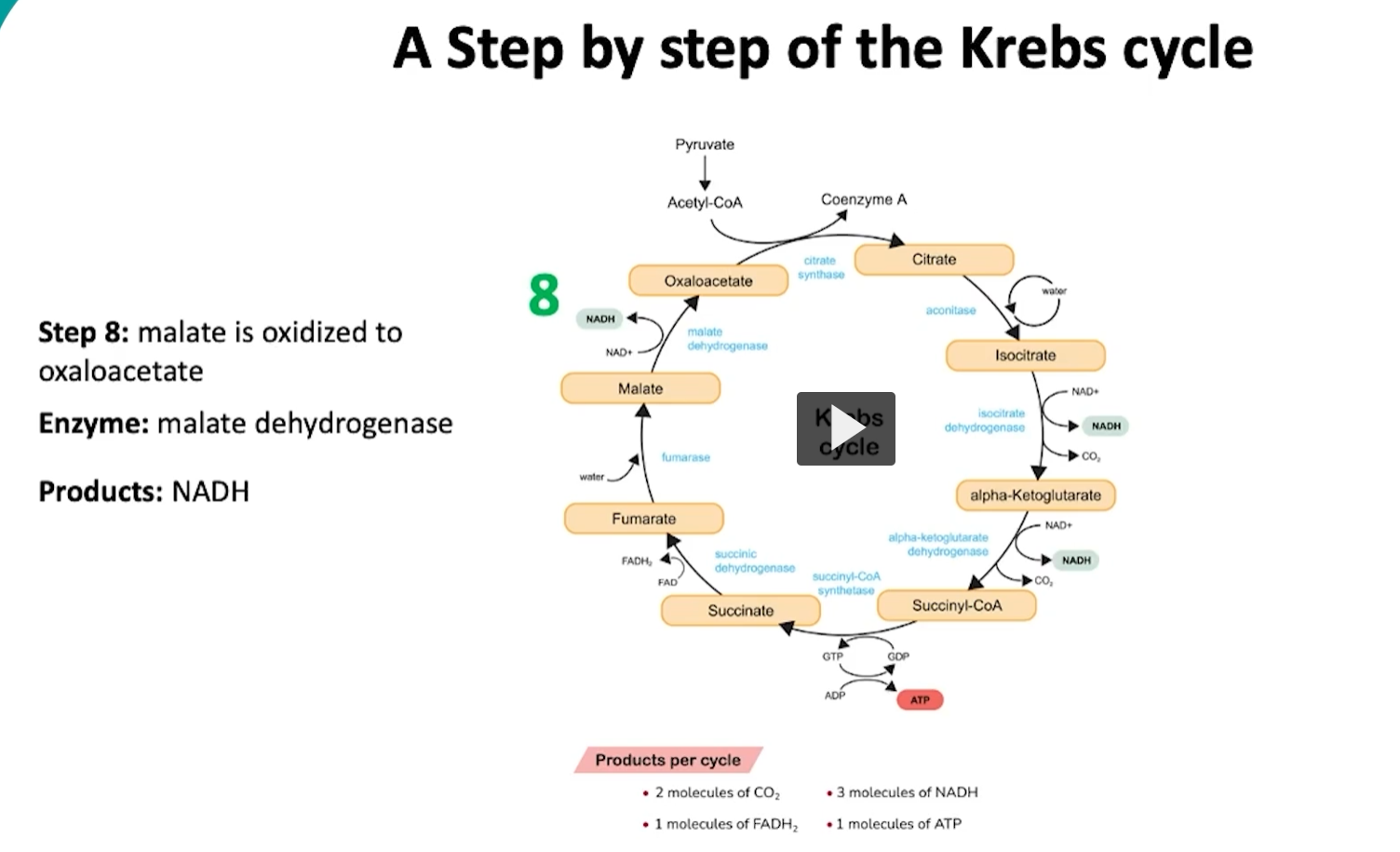
during the eighth and last step of the cycle, malate will be oxidized into oxaloacetate via the malate dehydrogenase.
what is happening here is that the hydroxyl group is malate is oxidized to a carbonyl group, during this oxidation, a pair of electrons REMOVED from the hydroxyl group of malate to form oxaloacetate.
then, the removed electrons are transferred to NAD reducing it to NADH.
this will finalize one cycle of the kreb cycle, specially producing an overall of two molecules of CO2, three molecules of NADH, one molecule of FADH2 and one molecule of ATP.
it’s very important to also keep in mind that 2 acetyl-CoA molecules are produces from one glucose molecule, which means the kreb cycle will run twice per glucose. This will double the product amount.

Here, we include a table with a summary of all the steps of the krebs cycle and important details to remember.

moving onto the regulation of the kreb’s cycle, it will occur at key steps of the involving enzymes that will catalyze irreversible reactions.
these checkpoints ensure that the cycle functions efficiently based on the the cell’s energy status and substrate availability.
specifically, we will have enzyme regulation step one, which will be inhibited by high levels of ATP, NADH or citrate.
activation on the other hand will be due to high levels of ADP, oxaloacetate or acetyl-CoA
regulation will also occur on step 3, specifically, inhibition will occur again at high levels of ATP or NADH, while activation occurs by the presence of ADP or calcium ions.
more regulation can occur in step four, which the reaction of the alpha ketoglutarate dehydrogenase is inhibited by high levels of ATP, high NADH levels, and high succinyl CoA levels.
and activation will occur by high levels of of calcium ions, other regulatory mechanisms like allosteric regulation, feedback inhibition, and energy charge of the cells will inhibit or activate this cycle.
all of these are important to prevent overprodcution of ATP to ensure intermediate of other pathways and to maintain metabolic flexiblity.

and finally we include the table with disorders and diseases related to dysfunction of the kreb cycle.
cancer
The cancer row in the table is highlighting that mutations in certain Krebs cycle enzymes can contribute to tumor growth. These enzymes are IDH, SDH, and FH, which are normally part of the TCA (Krebs) cycle in mitochondria.
Let’s unpack it step-by-step.
1. The enzymes involved
The table lists three enzymes:
IDH
Isocitrate Dehydrogenase
Reaction in the Krebs cycle:
Isocitrate→α-ketoglutarate+CO2+NADH\text{Isocitrate} \rightarrow \alpha\text{-ketoglutarate} + CO_2 + NADHIsocitrate→α-ketoglutarate+CO2+NADH
SDH
Succinate Dehydrogenase
Reaction:
Succinate→Fumarate\text{Succinate} \rightarrow \text{Fumarate}Succinate→Fumarate
This enzyme is also Complex II of the electron transport chain.
FH
Fumarate Hydratase
Reaction:
Fumarate→Malate\text{Fumarate} \rightarrow \text{Malate}Fumarate→Malate
2. What happens when these enzymes mutate
Normally these enzymes help generate energy.
But mutations can cause metabolic rewiring that favors tumor growth.
Mutated IDH
Mutated IDH produces an abnormal metabolite:
2-hydroxyglutarate (2-HG)
This molecule interferes with:
DNA methylation
gene regulation
cell differentiation
This can push cells toward cancerous behavior.
IDH mutations are seen in:
gliomas
acute myeloid leukemia (AML)
Mutated SDH
If succinate dehydrogenase is defective:
Succinate accumulates.
High succinate can activate hypoxia signaling pathways even when oxygen is present.
This stimulates:
angiogenesis
tumor growth.
SDH mutations are associated with:
paragangliomas
pheochromocytomas.
Mutated FH
If fumarate hydratase is defective:
Fumarate accumulates.
Excess fumarate can alter:
cellular signaling
oxidative stress pathways
gene regulation.
FH mutations are linked to:
renal cancers
hereditary leiomyomatosis.
3. Why altered metabolism helps cancer
Cancer cells often change their metabolism to support:
rapid cell growth
biosynthesis
survival in low oxygen environments.
These metabolic changes are sometimes called oncometabolite pathways.
Examples of oncometabolites:
2-hydroxyglutarate
succinate
fumarate
These molecules alter gene expression and promote tumor growth.
4. Clinical approach (from the table)
Because these mutations are specific, doctors can use targeted therapies.
Example:
IDH inhibitors
These drugs block the abnormal enzyme activity and reduce production of 2-hydroxyglutarate.
Simple summary
Enzyme mutated | What accumulates | Effect |
|---|---|---|
IDH | 2-hydroxyglutarate | alters gene regulation |
SDH | succinate | activates hypoxia signaling |
FH | fumarate | alters cell signaling |
All of these changes can promote tumor growth.
✅ Key idea
The table shows that mutations in Krebs cycle enzymes can create abnormal metabolites that disrupt gene regulation and cellular metabolism, helping drive cancer development.


TCA mitochondrial disorders
The “Mitochondrial Disorders” row in the table is explaining diseases where the mitochondria do not function properly, which disrupts the Krebs cycle and ATP production. Since mitochondria are the main site of cellular energy production, problems with them can affect many organs.
Let’s break down what the table means.
1. What mitochondrial disorders are
Mitochondrial disorders are diseases caused by defects in:
mitochondrial DNA (mtDNA), or
nuclear genes that encode mitochondrial proteins.
Because mitochondria produce ATP, these disorders mainly affect tissues that need a lot of energy.
Examples of high-energy tissues:
brain
muscles
heart
liver
2. Krebs cycle connection (Defective ATP production)
The table says:
“Defective ATP production.”
This happens because mitochondria contain:
the Krebs (TCA) cycle
the electron transport chain
oxidative phosphorylation
If mitochondria malfunction:
The Krebs cycle cannot efficiently produce NADH and FADH₂.
The electron transport chain cannot produce ATP efficiently.
Cells become energy deficient.
Result:
Low ATP → poor cellular function.
3. Symptoms of mitochondrial disorders
Because the body lacks energy, symptoms often involve:
Neurologic problems
seizures
developmental delay
stroke-like episodes
Muscle problems
muscle weakness
exercise intolerance
Organ dysfunction
cardiomyopathy
liver disease
vision problems
4. Why mitochondria are especially vulnerable
Mitochondria have their own DNA:
mtDNA
Characteristics:
inherited from the mother
more susceptible to mutations
codes for proteins involved in energy production.
Mutations in mitochondrial genes disrupt ATP synthesis.
5. Clinical approach (from the table)
The table lists treatments aimed at supporting mitochondrial function, not curing the genetic defect.
Ketogenic diet
Provides fat-derived ketone bodies as an alternative energy source for the brain.
Ketones can enter the Krebs cycle as acetyl-CoA.
CoQ10 (Coenzyme Q10)
CoQ10 is part of the electron transport chain.
It helps transport electrons between:
Complex I / II
Complex III.
Supplementation can improve mitochondrial electron transport.
Antioxidants
Defective mitochondria produce excess reactive oxygen species (ROS).
Antioxidants help reduce oxidative damage.
Examples:
vitamin C
vitamin E
alpha-lipoic acid.
6. Why this relates to the Krebs cycle
The Krebs cycle feeds electrons into the electron transport chain.
It produces:
NADH
FADH₂
If mitochondria are damaged:
these molecules cannot generate ATP efficiently
the Krebs cycle slows
energy production collapses.
Big picture
Problem | Result |
|---|---|
mitochondrial dysfunction | low ATP |
impaired Krebs cycle | less NADH / FADH₂ |
defective electron transport | poor oxidative phosphorylation |
cells lack energy | tissue damage |
✅ Key idea
The table is showing that mitochondrial disorders disrupt the Krebs cycle and oxidative phosphorylation, leading to defective ATP production, which particularly harms energy-demanding tissues like the brain and muscles.


The Neurodegenerative Diseases row in the table explains how problems in the TCA (Krebs) cycle and mitochondrial metabolism can contribute to diseases where neurons gradually die or lose function.
Let’s connect the puzzle step-by-step.
1. What neurodegenerative diseases are
Neurodegenerative diseases are disorders where neurons progressively deteriorate.
Common examples include:
Alzheimer’s disease
Parkinson’s disease
Huntington’s disease
ALS (amyotrophic lateral sclerosis)
These diseases mainly affect the brain and nervous system.
2. Why neurons depend heavily on the TCA cycle
Neurons require huge amounts of ATP because they constantly maintain electrical signaling.
Energy is required for:
Na⁺/K⁺ ATPase pumps
neurotransmitter release
axonal transport
synaptic activity.
Most of this ATP comes from mitochondrial metabolism, specifically:
TCA (Krebs) cycle
Electron transport chain
Oxidative phosphorylation
3. Role of the TCA cycle in neuronal energy
The TCA cycle generates:
NADH
FADH₂
These molecules deliver high-energy electrons to the electron transport chain.
This process produces the majority of ATP in neurons.
4. What happens if the TCA cycle is dysfunctional
If the TCA cycle is impaired:
less NADH and FADH₂ are produced
electron transport slows
ATP production decreases
This causes neuronal energy deficits, which is exactly what the table states.
Since neurons have extremely high energy demands, they are very sensitive to ATP shortages.
5. Mitochondrial dysfunction in neurodegenerative diseases
Many neurodegenerative diseases involve mitochondrial damage.
Examples:
Parkinson’s disease
Defects in mitochondrial Complex I lead to reduced ATP and increased oxidative stress.
Alzheimer’s disease
Impaired mitochondrial metabolism and TCA cycle enzymes lead to energy deficits in neurons.
Huntington’s disease
TCA cycle dysfunction and mitochondrial damage contribute to neuronal death.
6. Oxidative stress also increases
When mitochondria malfunction:
reactive oxygen species (ROS) accumulate
oxidative damage occurs in neurons.
This further damages:
mitochondrial DNA
proteins
membranes.
7. Clinical approach (from the table)
The table lists:
“Mitochondria-targeted therapies.”
These treatments aim to improve mitochondrial function and energy metabolism.
Examples include:
antioxidants (reduce oxidative stress)
mitochondrial cofactors (e.g., CoQ10)
metabolic therapies that improve energy production.
Big picture
Problem | Effect |
|---|---|
TCA cycle dysfunction | less NADH/FADH₂ |
ETC slows | less ATP |
neurons lack energy | neuronal damage |
mitochondrial ROS | oxidative stress |
✅ Key idea
Neurodegenerative diseases are linked to TCA cycle and mitochondrial dysfunction, which reduces ATP production and increases oxidative stress, leading to neuronal energy deficits and progressive neuron loss.

TCA cycle diabetes
The Diabetes row in the table is explaining how diabetes affects the TCA (Krebs) cycle and cellular energy metabolism, and how some treatments help restore mitochondrial function.
Let’s connect the biochemistry puzzle.
1. What diabetes is (metabolically)
Diabetes mellitus is a condition where:
insulin is deficient (Type 1), or
cells are resistant to insulin (Type 2).
Insulin normally allows cells to take up glucose and use it for energy.
When insulin signaling is abnormal:
glucose metabolism is altered
energy metabolism shifts toward fat metabolism.
2. How diabetes affects the TCA cycle
The table says:
“TCA cycle imbalance affects glucose metabolism.”
This happens through several mechanisms.
Reduced glucose utilization
Normally:
Glucose → glycolysis → pyruvate → acetyl-CoA → TCA cycle
In diabetes:
cells cannot use glucose efficiently
less pyruvate enters mitochondria
the TCA cycle becomes imbalanced.
Increased fatty acid metabolism
Because glucose cannot be used well, the body relies more on fatty acid oxidation.
Fat breakdown produces large amounts of:
acetyl-CoA
If too much acetyl-CoA accumulates:
the TCA cycle becomes overloaded
excess acetyl-CoA is diverted into ketone body production.
This is why uncontrolled diabetes can lead to:
diabetic ketoacidosis (DKA).
Mitochondrial stress
High glucose and fatty acids can also cause:
mitochondrial dysfunction
increased reactive oxygen species (ROS).
This damages the electron transport chain and worsens ATP production efficiency.
3. Why the TCA cycle imbalance matters
If the TCA cycle and mitochondria are stressed:
ATP production becomes inefficient
oxidative stress increases
cells become metabolically dysfunctional.
This contributes to long-term diabetic complications such as:
neuropathy
nephropathy
retinopathy
cardiovascular disease.
4. Clinical approach mentioned in the table
The table lists:
Metformin (enhances mitochondrial function).
Metformin is one of the most common treatments for Type 2 diabetes.
It works by:
reducing glucose production in the liver
improving insulin sensitivity
modifying mitochondrial metabolism
reducing excess glucose entering metabolic pathways.
This helps restore metabolic balance, including effects on the TCA cycle.
5. Big picture connection
Normal metabolism | Diabetes metabolism |
|---|---|
Glucose → TCA cycle → ATP | Glucose not used efficiently |
Balanced acetyl-CoA supply | Excess acetyl-CoA from fat |
Efficient ATP production | mitochondrial stress |
✅ Key idea
In diabetes, impaired glucose metabolism and increased fatty acid oxidation disrupt the TCA cycle and mitochondrial energy balance, contributing to metabolic stress and diabetic complications.

TCA cycle ischemia/hypoxia
The Ischemia / Hypoxia row in the table explains how lack of oxygen slows the TCA (Krebs) cycle and ATP production, which can damage tissues.
Let’s connect the metabolic puzzle step-by-step.
1. What ischemia and hypoxia mean Ischemia
From Greek:
isch- = to hold back
haima = blood
Meaning: reduced blood flow to tissue.
Example:
heart attack (myocardial infarction)
stroke.
Hypoxia
From Greek:
hypo = low
oxia = oxygen
Meaning: low oxygen levels in tissues.
Ischemia often causes hypoxia because blood carries oxygen.
2. Why oxygen matters for the TCA cycle
The TCA cycle itself does not directly use oxygen, but it depends on oxygen indirectly.
Why?
Because the TCA cycle produces:
NADH
FADH₂
These molecules deliver electrons to the electron transport chain (ETC).
3. Oxygen is the final electron acceptor
In the electron transport chain:
O2+electrons+H+→H2OO_2 + electrons + H^+ → H_2OO2+electrons+H+→H2O
Oxygen accepts electrons at Complex IV.
This allows:
NADH → NAD⁺
FADH₂ → FAD
These oxidized cofactors (NAD⁺ and FAD) are required for the TCA cycle to continue.
4. What happens during hypoxia
If oxygen is low:
Electron transport chain stops
NADH cannot be oxidized to NAD⁺
NAD⁺ becomes depleted
Without NAD⁺, several TCA reactions stop:
Isocitrate dehydrogenase
α-ketoglutarate dehydrogenase
Malate dehydrogenase
Result:
➡ TCA cycle slows or stops
5. Energy crisis in the cell
If the TCA cycle stops:
less NADH
less oxidative phosphorylation
ATP production collapses
Cells switch to anaerobic glycolysis, which produces:
lactate
This causes lactic acidosis.
6. Why ischemia damages tissues
High-energy tissues are especially vulnerable:
brain
heart
kidneys.
Without ATP:
ion pumps fail
calcium accumulates
cells swell
cell death occurs.
7. Clinical approaches in the table Hyperbaric oxygen
Patient breathes high-pressure oxygen.
This increases oxygen delivery to tissues and restores:
electron transport
ATP production.
Ischemic preconditioning
This is a protective strategy where tissues are exposed to short, controlled episodes of low oxygen.
This activates protective pathways that make tissues more resistant to severe ischemia later.
Big picture connection
Step | What happens |
|---|---|
Oxygen decreases | ETC stops |
NADH accumulates | NAD⁺ decreases |
TCA cycle slows | ATP production drops |
Cells rely on glycolysis | lactate accumulates |
Energy failure | tissue damage |
✅ Simple key idea
In ischemia or hypoxia, lack of oxygen stops the electron transport chain, preventing regeneration of NAD⁺ and FAD, which causes the TCA cycle to slow and ATP production to collapse, leading to cellular injury.
