Biochem | Ch. 6 + Ch. 7
1/71
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
72 Terms
Need for Biological Catalysts
Review of peptide bond schematic on left!!! A peptide bond forms by the combination of a carboxyl group of one amino acid and the amine group of a second amino acid, thereby excluding water (condensation reaction). IF we want to break that bond, we do so by adding water (HYDROLYSIS) (that is, LYSIS by adding water; that’s illustrated in the bottom schematic!). We’ll add the oxygen to the CARBONYL carbon to form the CARBOXYL group on one end, and we add the 2 hydrogen atoms to the N to form the amine group on the other end!
Half-life of peptide bonds (peptide bond = an amide bond) in vitro [in glass opposed to in vivo] [outside the cell] is ~20 years. SO, if I had a test tube of protein after 20 years, HALF of those peptide bonds would still be intact! Very strong, very stable bond. Comforting since MOST of the structure, most of the activity in the cell relates to this very strong bond!
![<p>Review of peptide bond schematic on left!!! A peptide bond forms by the combination of a carboxyl group of one amino acid and the amine group of a second amino acid, thereby excluding water (condensation reaction). IF we want to break that bond, we do so by adding water (HYDROLYSIS) (that is, LYSIS by adding water; that’s illustrated in the bottom schematic!). We’ll add the oxygen to the CARBONYL carbon to form the CARBOXYL group on one end, and we add the 2 hydrogen atoms to the N to form the amine group on the other end!</p><p>Half-life of peptide bonds (peptide bond = an amide bond) in vitro [in glass opposed to in vivo] [outside the cell] is ~20 years. SO, if I had a test tube of protein after 20 years, HALF of those peptide bonds would still be intact! Very strong, very stable bond. Comforting since MOST of the structure, most of the activity in the cell relates to this very strong bond!</p>](https://assets.knowt.com/user-attachments/eb194d5e-8684-468f-a10f-dfadd52009f7.png)
Need for Biological Catalysts
Normal function of a cell is to recycle its components, including proteins and THUS PEPTIDE bonds. How can we make a chemical reaction happen more readily? Increase rate of a reaction by INCREASING the temperature (but as living organisms, we can only tolerate a small fluctuation in temperature). INCREASING [reactants] (we know from Le Chatelier’s, if we INCREASE the concentration of the reactants, [shift in equilibrium towards products], we can INCREASE the concentration of the products) (BUT also not feasible in a living cell since it [a living cell] doesn’t always control the availability of reactants or biological molecules [makes sense]). Add a catalysts (ONLY feasible option!!).
![<p>Normal function of a cell is to recycle its components, including proteins and THUS PEPTIDE bonds. How can we make a chemical reaction happen more readily? Increase rate of a reaction by INCREASING the temperature (but as living organisms, we can only tolerate a small fluctuation in temperature). INCREASING [reactants] (we know from Le Chatelier’s, if we INCREASE the concentration of the reactants, [shift in equilibrium towards products], we can INCREASE the concentration of the products) (BUT also not feasible in a living cell since it [a living cell] doesn’t always control the availability of reactants or biological molecules [makes sense]). Add a catalysts (ONLY feasible option!!).</p>](https://assets.knowt.com/user-attachments/bd8a2d6a-d738-48d6-badf-fb3af78cf874.png)
ENZYMES as Catalysts
“Enzyme” comes from the Greek, meaning “yeast.” Something smaller inside the cell was responsible for the reaction. Enzymes are TRUE catalysts; that is, they are NOT permanently altered or consumed. They may be modified in the course of the reaction; however, at the END of the reaction of the cycle, they must return to the original form. MOST enzymes are PROTEIN but SOME are made of RNA!!!!
Enzymes as Catalysts
Can speed up reactions to an amazing degree! Table from book, representing the rate enhancement of enzymes. First column = name of enzyme. 2nd column = half-time. 3rd column = rate of the uncatalyzed reaction (how MANY times it HAPPENS per second [?] [I don’t think so; Chat: the uncatalyzed rate = the rate at which A chemical reaction occurs without a catalyst [this makes more sense to me] [actually, no! the video is correct; the rate is how much product per unit of time!). Next column = catalyzed rate; how fast it happens with the aid of the enzyme. Last column = rate enhancement; THE CATALYZED RATE DIVIDED by the UNCATLYZED RATE; how much FASTER did the reaction go! (think: if we are looking at rate enhancement; we are looking at how much FASTER is the reaction going to go! put the FASTER in the numerator??). Units across Uncatalyzed Rate and Catalyzed Rate = s^-1.
Reminder: The enzyme carbonic anhydrase is responsible for maintaining the equilibrium between carbon dioxide and bicarbonate (CO2 + H2O <=> H2CO3 <=> H+ + HCO3-) are very important buffering system in the blood (think maintaining pH in the blood!).
Last row = carbonic anhydrase. Uncatalyzed rate = 0.13 per second. AKA several seconds would have to pass before the reaction occurred? (interesting wording?) (Clarification: If the uncatalyzed rate constant is 0.13 s⁻¹, that means each reactant molecule has about a 0.13 probability per second of reacting. A useful approximation is: average time for one reaction is approximately 1/k. So, 1/(0.13 s^-1) is approximately 7.7 seconds! This means that, on average, a given molecule would take about 8 seconds to react without the enzyme!) In the presence of the enzyme, the catalyzed rate is 1 M TIMES per second. The chemical reaction OCCURS 1 M times in 1 second!! That represents a rate enhancement of a trillion (a trillion = 1,000 billion) fold! A truly amazing degree of rate enhancement. Not all enzymes work this fast but with all the chemical reactions that need to occur in a living cell, without enzymes, we couldn’t carry out those reactions on a time scale that would make life possible.
![<p>Can speed up reactions to an amazing degree! Table from book, representing the rate enhancement of enzymes. First column = name of enzyme. 2nd column = half-time. 3rd column = rate of the uncatalyzed reaction (how MANY times it HAPPENS per second [?] [I don’t think so; Chat: the uncatalyzed rate = the rate at which A chemical reaction occurs without a catalyst [this makes more sense to me] [actually, no! the video is correct; the rate is how much product per unit of time!). Next column = catalyzed rate; how fast it happens with the aid of the enzyme. Last column = rate enhancement; THE <strong>CATALYZED RATE </strong>DIVIDED by the UNCATLYZED RATE; how much <strong>FASTER </strong>did the reaction go! (think: if we are looking at rate enhancement; we are looking at how much FASTER is the reaction going to go! put the FASTER in the numerator??). Units across Uncatalyzed Rate and Catalyzed Rate = s^-1.</p><p>Reminder: The enzyme carbonic anhydrase is responsible for maintaining the equilibrium between carbon dioxide and bicarbonate (CO2 + H2O <=> H2CO3 <=> H+ + HCO3-) are very important buffering system in the blood (think maintaining pH in the blood!).</p><p>Last row = carbonic anhydrase. Uncatalyzed rate = <strong>0.13 per second</strong>. <strong>AKA several seconds would have to pass before the reaction occurred? </strong>(interesting wording?) (Clarification: If the <strong>uncatalyzed rate constant</strong> is <strong>0.13 s⁻¹</strong>, that means each reactant molecule has about a <strong>0.13 probability per second</strong> of reacting. A useful approximation is: average time for one reaction is approximately 1/k. So, 1/(0.13 s^-1) is approximately 7.7 seconds! This means that, on average, a given molecule would take about 8 seconds to react without the enzyme!) In the presence of the enzyme, the catalyzed rate is 1 M <strong>TIMES </strong>per second. The chemical reaction OCCURS 1 M times in 1 second!! That represents a rate enhancement of a trillion (a trillion = 1,000 billion) fold! A truly amazing degree of rate enhancement. Not all enzymes work this fast but with all the chemical reactions that need to occur in a living cell, without enzymes, we couldn’t carry out those reactions on a time scale that would make life possible.</p>](https://assets.knowt.com/user-attachments/20c4f334-61b8-447b-a361-7470f3eff6d4.png)
Enzymes as Catalysts
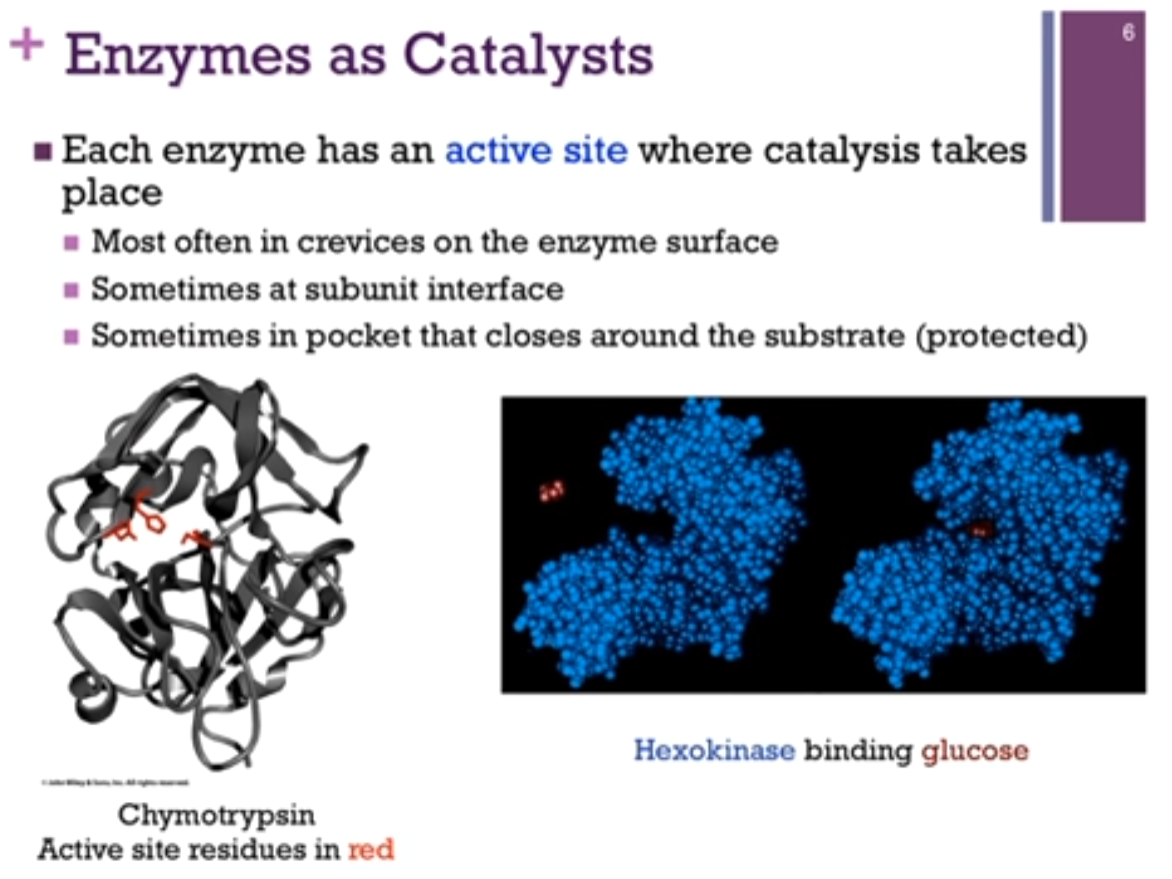
Each enzyme has an active site where catalysis (Chat: chemical reaction) takes place. MOST often in crevices on the enzyme surface, sometimes at subunit interface! (if talking about a multi-subunit complex), sometimes in pocket that closes around the substrate (protected).
Enzyme Chymotrypsin; active residues in red. IT IS in a CREVICE or buried portion (see schematic!) of the protein.
Enzyme Hexokinase. Sometimes there is a pocket in which the substrate (substrate = the specific reactant that an enzyme binds to and transforms during a reaction) binds and then the enzyme and then the enzyme closes around the substrate (see schematic!). The space-filling model in blue is hexokinase; the red is its substrate, glucose. There is a binding pocket for glucose but then, there’s a conformational change in the enzyme after it binds substrate. This gives it a protected environment. We OFTEN find with enzymes that catalysis occurs IN a protected environment (so that nothing interferes with the proper completion of the reaction cycle).
Enzymes as Catalysts
Biological catalysts differ from metal catalysts! Biological catalysts have complex, specific structures: catalysis may be associated with conformational changes; Work under mild conditions (T [temp] and P [pressure]). In contrast, with metal catalysts, they often require elevated temperature or pressure or both.
Also, biological catalysts only act on a specific substance (substrate) (the specific reactant that an enzyme binds to and transforms during a reaction) (Some are so specific, they will act on the L-enantiomer BUT not the D!) (some are more forgiving in what substrates they will binds) (can be important for detecting and measuring enzyme activity) (think: earlier table, where we measured how much product was formed per unit of time; we do that by measuring enzyme activity).
Example: Chymotrypsin: Normal function is to hydrolyze a peptide bond; it adds water across a peptide or amide bond; however, it can use the substrate (specific reactant that an enzyme binds to) para-Nitrophenylacetate, will HYDROLYZE the bond connecting the acetate to the Nitrophenol part of the molecule so clearly NOT a peptide bond and yet SIMILAR ENOUGH that chymotrypsin will work on this substrate! Product then is p-NitrophenoLATE (after = product), which is yellow in color, and we can detect that by its absorbance at a wavelength of 410 nm. SO, the substrate (specific reactant that an enzyme binds to) is colorless, the product is colored. We can measure the strength of the color as indicating the CONCENTRATION of product. We can measure how much product forms within a given period of time. AND this is how those measurements were arrive at with regard to HOW MUCH PRODUCT per UNIT TIME in the table we saw in the earlier slide!
![<p>Biological catalysts differ from metal catalysts! Biological catalysts have complex, specific structures: catalysis may be associated with conformational changes; Work under mild conditions (T [temp] and P [pressure]). In contrast, with metal catalysts, they often require elevated temperature or pressure or both. </p><p>Also, biological catalysts only act on a specific substance (substrate) (the specific reactant that an enzyme binds to and transforms during a reaction) (Some are so specific, they will act on the L-enantiomer BUT not the D!) (some are more forgiving in what substrates they will binds) (can be important for detecting and measuring enzyme activity) (think: earlier table, where we measured how much product was formed per unit of time; we do that by measuring enzyme activity). </p><p>Example: <strong>Chymotrypsin</strong>: Normal function is to hydrolyze a peptide bond; <strong>it adds water across a peptide or amide bond</strong>; however, it can use the substrate (specific reactant that an enzyme binds to) para-Nitrophenylacetate, will HYDROLYZE the bond connecting the acetate to the Nitrophenol part of the molecule so clearly NOT a peptide bond and yet SIMILAR ENOUGH that chymotrypsin will work on this substrate! Product then is p-NitrophenoLATE (after = product), which is yellow in color, and we can detect that by its absorbance at a wavelength of 410 nm. SO, the substrate (specific reactant that an enzyme binds to) is colorless, the product is colored. We can measure the strength of the color as indicating the CONCENTRATION of product. We can measure how much product forms within a given period of time. AND this is how those measurements were arrive at with regard to HOW MUCH PRODUCT per UNIT TIME in the table we saw in the earlier slide!</p>](https://assets.knowt.com/user-attachments/8d15d311-5171-4af1-b914-6eff282607cc.png)
Classes of Enzymes
Enzymes are often regulated by a number of mechanisms: Knowing “when” and “why” they work is as important as “how.” Video: Enzymes are often regulated by a number of mechanisms depending on the importance of the reaction which they catalyze, SO it’s not just a matter of knowing how they work, that is, what is their mechanism of action but also, under what conditions do they work, what regulates, and when they work, and why is that significant.
There are 6 classes of enzymes! (responsible to know!!) (know 6 classes AND the type of reactions they catalyze!) (if gave a reaction, you should be able to tell what type of reaction that was and the CLASS OF ENZYME THAT CATALYZES IT). Most of them involve the addition or removal of some functional group!
Classes of enzymes (6):
Addition or removal of functional group:
Oxidoreductases: They CATALYZE reactions that involve an oxidation and reduction.
Transferases: Involve the transfer of a functional group. They’re transferring that group from one molecule to another!
Hydrolases: They CATALYZE hydrolysis reactions. They are breaking (lysis) a bond by adding water.
Lyases: They also eliminate a group, but they thereby form double bonds.
Ligases: They involve bond FORMATION, and they couple with that ATP hydrolysis.
Rearrangement of substance:
Isomerases: They ALSO in a sense, add and remove a functional group; they remove some group from ONE location on the molecule and attach it to a different location (BUT WITHIN ONE MOLECULE so different from transferase!), so it’s an isomerization reaction (isomerization reaction: a type of chemical reaction in which a molecule is transformed into another molecule that has the SAME chemical formula but a different arrangement of atoms) (oh! makes sense!)!

Memory of Classes of Enzymes:
Old Teachers Hate Lazy Idle Learners:
Oxidoreductases, Transferases, Hydrolases, Lyases (note spelling!), Isomerases (rearrangement of substance), Ligases.
Oxidoreductase: transfers electrons (oxidation/reduction); O=oxidation. Transferase: Transfers a functional group BETWEEN molecules; T = Transfer. Hydrolase: Breaks bonds using water (lysis = break bond); Hydro = Water. Lyase = breaks bonds WITHOUT water/oxidation; OFTEN FORMS a DOUBLE BOND; L = Leaves behind a double bond!! Isomerase: Rearranges atoms WITHIN on molecule; I = Internal rearrangement. Ligase = joins two molecules together, USUALLY using ATP!! Ligate = Link. Think: When we form a bond, it takes energy, so we often COUPLE ligases with ATP hydrolysis!
Classes of Enzymes: Examples
Top right = Reaction where pyruvate is decarboxylated!! to produce acetaldehyde. You can see the CO2 group in red has been removed, AND we have THEREFORE formed a double bond; fits our classification of LYASE!! (interesting! note, this reaction is NOT oxidation/reduction! and also, lyase = leaves behind a double bond; note; leave behind is not in [kinda original] larger molecule; double bond is in the CO2 [I think] [or not sure?]). See schematic! Chat: When the carboxylate group (-COO^-) LEAVES as CO2, the electrons form the BROKEN bond HAVE to go somewhere. A simplified way to think about it is: 1.) The C-C bond to the carboxylate group breaks. 2.) The electrons left behind create a DOUBLE bond between atoms in the REMAINING fragment. 3.) THAT double bond is then rearranged/protonated, giving the final product acetaldehyde (note spelling!): CH3-CHO. SO => when your instructor says “we have therefore formed a double bond,” they usually mean: During the elimination of CO2, the electrons from the broken bond are used to CREATE a NEW pi bond (double bond) somewhere in the intermediate?? The final product acetaldehyde does contain a double bond - the carbonyl bond (but so did the reactant??): CH3 - C(=O)H. The C=O bond is a double bond. Connection to Lyases: A hallmark of lyases is: remove a group (CO2, H2O, NH3, etc.) and generate a double bond or use a double bond in the reaction. That’s why decarboxylation reactions are often classified as lyase reactions! Honestly: Really check this or ask with the teacher; seems confusing??? Where is the double bond (that is typically left behind or formed?). What was meant in class by oxidation?
Bottom right = We can see we’re removing this amine group (blue) from Ala, and we’re transferring that to alpha-Ketoglutarate to form glutamate => transferase.
BOTTOM LEFT = Reaction that is catalyzed by ALCOHOL dehydrogenase, so are converting ethyl alcohol into acetaldehyde (note spelling). Look at schematic!!! We’re REPLACING a CH with a CO bond; THAT’S an OXIDATION (note that it is NOT a ligase; we are not linking two molecules together!) (also not an isomerase!). SO, that would classify as an OXIDOREDUCTASE!
![<p><strong>Top right </strong>= Reaction where pyruvate is decarboxylated!! to produce acetaldehyde. You can see the CO2 group in red has been removed, AND we have <strong>THEREFORE formed a double bond</strong>; fits our classification of <strong>LYASE</strong>!! (interesting! note, this reaction is NOT oxidation/reduction! and also, lyase = leaves behind a double bond; note; leave behind is not in [kinda original] larger molecule; double bond is in the CO2 [I think] [or not sure?]). See schematic! Chat: When the carboxylate group (-COO^-) LEAVES as CO2, the electrons form the BROKEN bond HAVE to go somewhere. A simplified way to think about it is: 1.) The C-C bond to the carboxylate group breaks. 2.) The electrons left behind create a DOUBLE bond between atoms in the REMAINING fragment. 3.) THAT double bond is then rearranged/protonated, giving the final product <strong>acetaldehyde </strong>(note spelling!): CH3-CHO. SO => when your instructor says “we have therefore formed a double bond,” they usually mean: During the elimination of CO2, the electrons from the broken bond are used to CREATE a NEW pi bond (double bond) somewhere in the intermediate?? The final product acetaldehyde does contain a double bond - the carbonyl bond (but so did the reactant??): CH3 - C(=O)H. The C=O bond is a double bond. Connection to Lyases: A hallmark of lyases is: remove a group (CO2, H2O, NH3, etc.) and generate a double bond or use a double bond in the reaction. That’s why decarboxylation reactions are often classified as lyase reactions! Honestly: Really check this or ask with the teacher; seems confusing??? Where is the double bond (that is typically left behind or formed?). What was meant in class by oxidation?</p><p>Bottom right = We can see we’re removing this amine group (blue) from Ala, and we’re transferring that to alpha-Ketoglutarate to form glutamate => <strong>transferase</strong>.</p><p>BOTTOM LEFT = Reaction that is catalyzed by ALCOHOL dehydrogenase, so are converting ethyl alcohol into <strong>acetaldehyde </strong>(note spelling). Look at schematic!!! We’re REPLACING a CH with a CO bond; THAT’S an OXIDATION (note that it is NOT a ligase; we are not linking two molecules together!) (also not an isomerase!). SO, that would classify as an <strong>OXIDOREDUCTASE</strong>!</p>](https://assets.knowt.com/user-attachments/136c57a4-6851-409c-a8c1-54e023f5bbf9.png)
Naming Enzymes
Enzymes are often names for the substrate (the reactant that binds to the enzyme) that it acts on OR on the product it produces!
Enzyme name most often ends in “-ase.”
Going to look into detail into the enzyme chymotrypsin.
Name often tells you what it does.
Look at schematics! We’re converting pyruvate to acetaldehyde (LYSIS!), AND it was DECARBOXYLATED, so the name of the enzyme is pyruvate decarboxylase since it removes a carboxyl (-COOH / -COO- group) from pyruvate!
On the bottom right, the enzyme is called alanine aminotransferase since it is transferring an amino group FROM alanine to alpha-Ketoglutarate.
Also note: There is OFTEN MORE THAN ONE VERSION of the same enzymes. MULTIPLE enzymes that CATALYZE the SAME reaction. These are called isozymes; iso = same; that is, they have the same relative function, but might operate in different tissues or they might be expressed under different conditions [perhaps they are expressed depending on pH or temperature]). Isozymes often differ in catalytic properties. Clarification: “differ in catalytic properties” => for example, one isozyme might work best at different pH or temperature; different isozymes may respond differently to hormones, etc.
![<p>Enzymes are often names for the <strong>substrate </strong>(the reactant that binds to the enzyme) that it acts on OR on the product it produces!</p><p>Enzyme name most often ends in “-ase.”</p><p>Going to look into detail into the enzyme chymotrypsin. </p><p>Name often tells you what it does. </p><p>Look at schematics! We’re converting pyruvate to acetaldehyde (<strong>LYSIS</strong>!), AND it was DECARBOXYLATED, so the name of the enzyme is pyruvate decarboxylase since it removes a <strong>carboxyl (-COOH / -COO- group) </strong>from pyruvate!</p><p>On the bottom right, the enzyme is called alanine aminotransferase since it is transferring an amino group FROM alanine to alpha-Ketoglutarate. </p><p>Also note: There is OFTEN MORE THAN ONE VERSION of the same enzymes. <strong>MULTIPLE enzymes that CATALYZE the SAME reaction</strong>. These are called <strong>isozymes</strong>; iso = same; that is, they have the same relative function, but might operate in different tissues or they might be expressed under different conditions [perhaps they are expressed depending on pH or temperature]). Isozymes often differ in <strong>catalytic properties</strong>. Clarification: “differ in catalytic properties” =><strong> </strong>for example, one isozyme might work best at different pH or temperature; different isozymes may respond differently to hormones, etc.</p>](https://assets.knowt.com/user-attachments/ba86befd-763d-4b3d-81e2-abfdad05d5fa.png)
Chemistry of Catalysis
Catalysts do NOT change the DIRECTION of a chemical reaction; they cannot influence the equilibrium or balance of that reaction.
For 2 compounds to react, the two molecules have to get close enough to react, and there are ALWAYS some repulsive forces there. That represents an energy barrier that we must OVERCOME if we’re going to form those products!

Chemistry of Catalysis
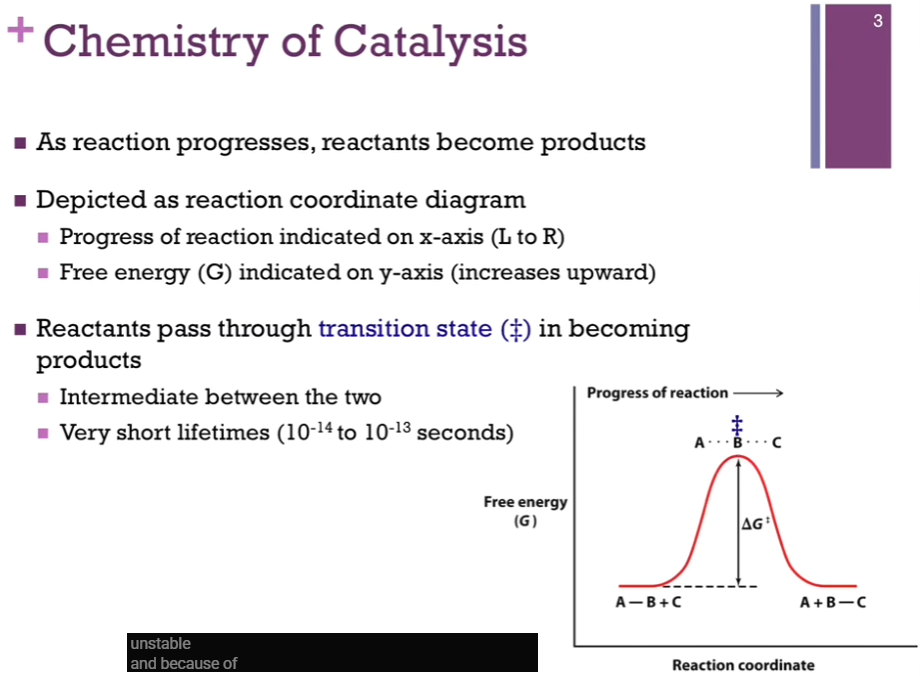
X-axis => reaction coordinate.
Y-axis => free energy (G).
Halfway point (in reaction coordinate between reactants and products) represents our intermediate => this is our transition state AND that would be the TOP of our energy hill, DEPICTED by this double dagger (look at schematic!). AS if our group B - halfway between our A and C. (In our reaction B is being transferred to group C). Notice it’s at a VERY high energy level => that means it’s VERY unstable! AND because of that, the lifetime is very short (less than a picosecond)!
Review on Reaction Coordinate Diagrams
This diagram shows how the energy of a reaction changes as reactants turn into products. Y-axis = higher up = higher energy = less stable. Lower down = lower energy = more stable. Top of the hill = the transition state (‡)!!
The activation energy is: ΔG‡. Big hill = slow reaction. Smaller hill = faster reaction.
Blue curve (uncatalyzed) = higher activation energy = reaction happens very slowly = deltaGdoublecorssuncat. Red curve = enzyme lower activation energy = reaction becomes faster. deltaGdoublecrosscat.
IMPORTANT POINT = deltaG does NOT change: the overall energy difference between reactants and products. Since products are lower in energy here: deltaG = negative, reaction is thermodynamically favorable, there is a release in free energy. BTW the enzyme does not change deltaG, does not change equilibrium, only changes the speed by lowering activation energy! Connection to kinetically stable: A metabolite can be kinetically stable if the activation barrier is HIGH. (metabolite is kinetically stable). Even if products are lower in energy (deltaG = negative): the molecule stays around; because climbing the activation hill is slow. Enzymes lower the hill and allow the reaction to proceed quickly (ah! makes sense now!).
Other previous: Metabolite is kinetically stable => it does not change or react quickly, even if converting into something else would be energetically favorable; the REACTION is SLOW because of a high ACTIVATION energy barrier.
More Chat: “Kinetically stable” does not mean “thermodynamically stable.” A molecule can be unstable in terms of energy (deltaG < 0) BUT STILL stick around for a long time. That’s because of activation energy barrier (Ea). A metabolite is kinetically stable if: The reaction it could undergo has a high activation energy barrier, so it happens very slowly. Even if: Products are lower energy (deltaG is negative => reaction is favorable) => It still doesn’t proceed quickly because it can’t easily “climb the hill.”
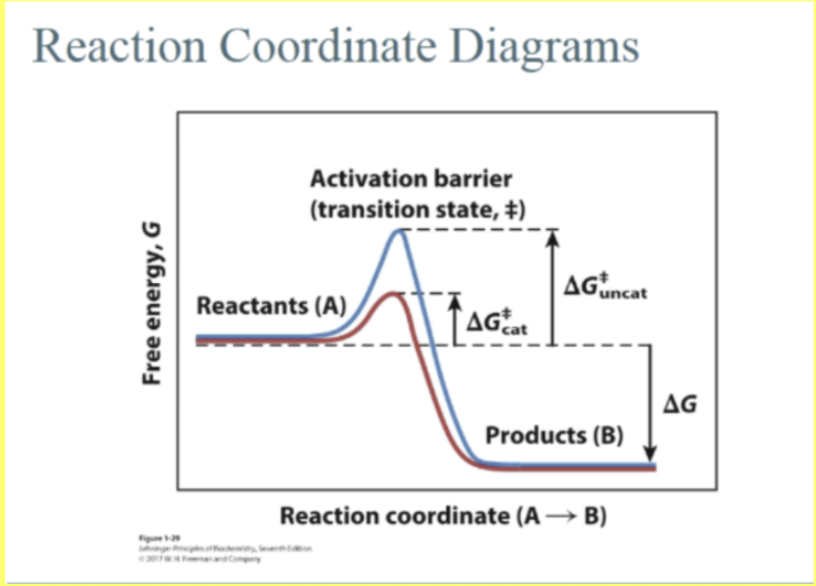
Chemistry of Catalysis
This is the energy hill. This is the barrier that represents repulsion of the forces as these molecules come together. On left, this is our deltaGcross (or deltaGdoubledagger) or Ea or activation energy. REMEMBER, that DOUBLE DAGGER is OUR TRANSITION STATE!! deltaGdoubledagger is the change associated with our top of the hill here, our transition state, AND our STARTING POINT, the reactants. deltaGdoubledagger represents the amount of energy our REACTANTS must have to reach the bottom of the hill on the other side and make products!
Higher the hill, the fewer molecules we will have THAT will have enough energy to climb the hill, and make the products. What we want is some way to lower that energy hill SO we can make MORE product in the same amount of TIME.
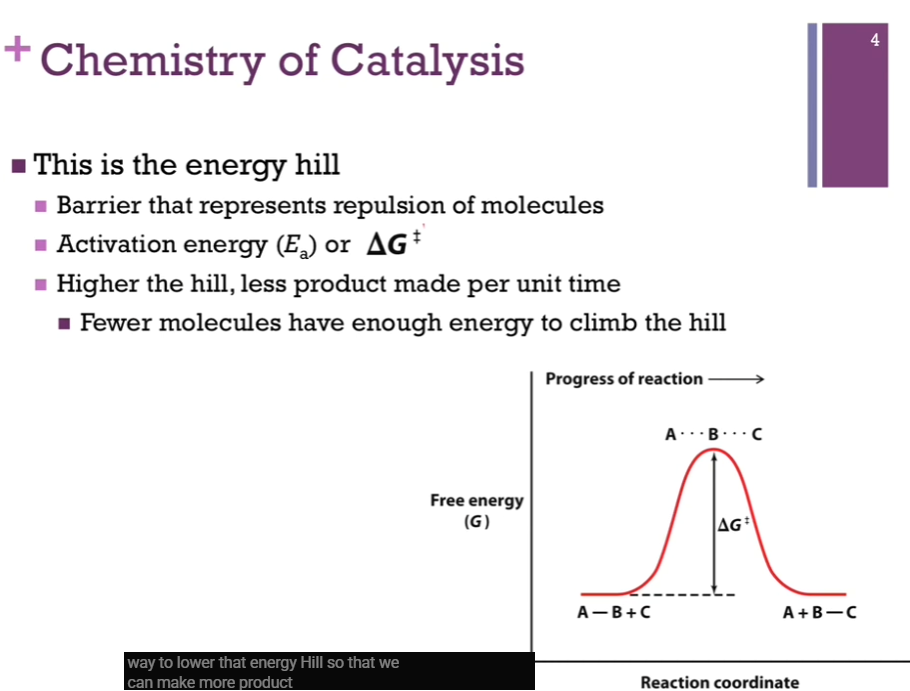
Chemistry of Catalysis
If you are at the top of the hill, our transition state, we could easily roll either way down the side of that hill. We can reach that transition state (unstable) and instead of forming product, we could slide down the other way and go back to reactants!
In this idealized case (see left!), the energy of our reactants AND product is the same! (deltaG = 0)! There is no preference to go one way or the other; this rarely happens. Usually, a reaction favors either products or reactants.
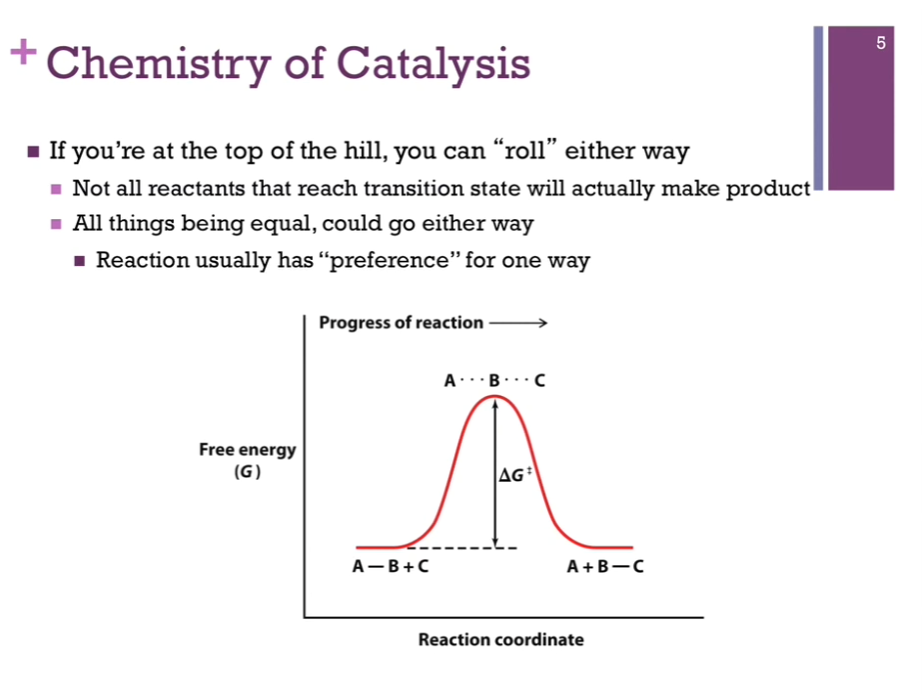
Chemistry of Catalysis
If products on free energy versus reaction coordinate is lower that reactants, deltaG = negative, reaction is spontaneous! Release [free] energy.
When deltaG = positive, need to input [free] energy to make that reaction happen.
Reminder of schematics is on the left!
![<p>If products on free energy versus reaction coordinate is lower that reactants, deltaG = negative, reaction is spontaneous! Release [free] energy. </p><p>When deltaG = positive, need to input [free] energy to make that reaction happen. </p><p>Reminder of schematics is on the left!</p>](https://assets.knowt.com/user-attachments/bfd446a9-3080-44df-b00d-eb593856e1f5.png)
Making It Happen
Xdoubledaggercat, our intermediate, has a lower free energy and is far less unstable than Xdoubledaggeruncat => AKA Xdoubledaggercat is more STABLE than Xdoubledaggeruncat => (with catalyst) more molecules can get to the top of the hill and make it to products; deltaGdoubledaggercat is much less than deltaGdoubledaggeruncat.
Enzymes (catalysts) only change deltaGdoubledagger! Make deltaGdoubledagger lower!
Catalysts do NOT change the equilibrium; catalysts only make it more likely that the reactants will form the products; catalysts lower the energy hill.
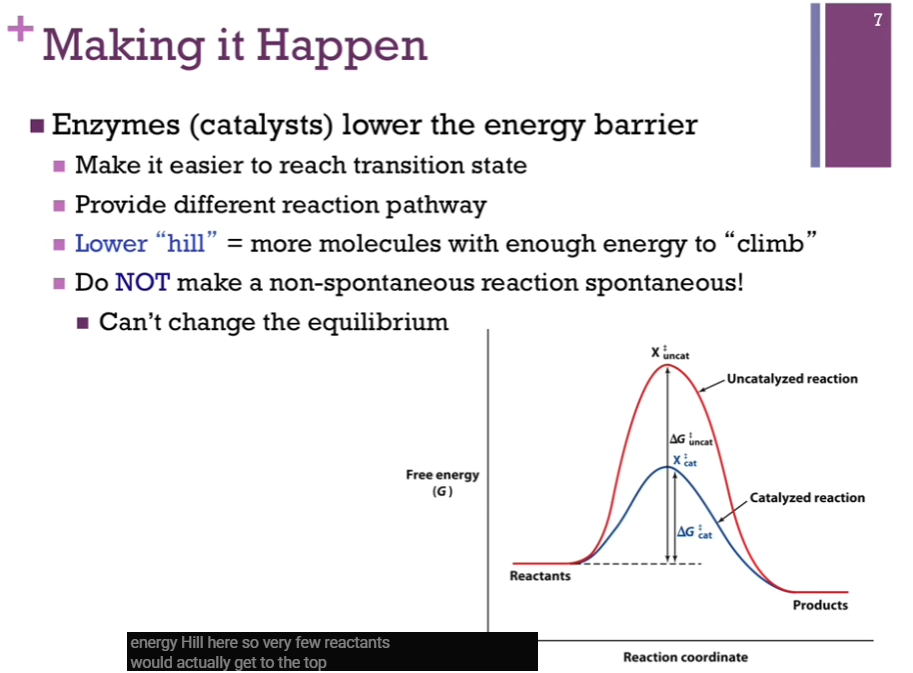
Catalytic Helpers
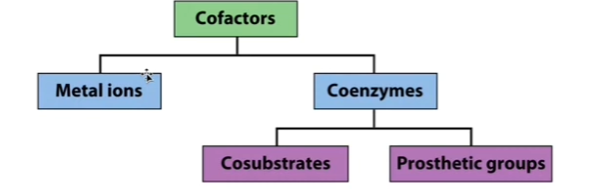
Enzymes don’t work alone. They may need cofactors (Chat: components that some enzymes need in order to function properly) to carry out the catalytic mechanisms. Sometimes, these cofactors carry chemical groups that simply aren’t present on amino acid side chains. COFACTORS actually participate in the catalytic mechanisms! Sometimes, they are inorganic (like metal ions! [capable of multiple oxidation states] [helpful in oxidation reduction reactions]).
![<p>Enzymes don’t work alone. They may need cofactors (Chat: components that some enzymes need in order to function properly) to carry out the <strong>catalytic </strong>mechanisms. Sometimes, these cofactors carry chemical groups that simply aren’t present on amino acid side chains. COFACTORS actually participate in the catalytic mechanisms!<strong> Sometimes, they are inorganic (like metal ions! [capable of multiple oxidation states] [helpful in oxidation reduction reactions]). </strong></p>](https://assets.knowt.com/user-attachments/7435e789-9186-4578-ac4d-330e6a683093.png)
Catalytic Helpers
Cofactor can ALSO be a COENZYME (an organic molecule!). Can classify these coenzymes (organic molecules that are cofactors) as either prosthetic groups (prosthetic groups = the coenzyme remains associated with the enzyme, like heme in Mb & Hb!) OR cosubstrates (cosubstrates = coenzymes that may enter AND exit the active site [reminder: [almost] each enzyme has an active site where catalysis [chemical reaction] takes place!] [Clarification: Almost every enzyme has an active site WHERE: the substrate binds AND catalysis [the chemical reaction] occurs!] in the course of the reaction).
![<p>Cofactor can ALSO be a <strong>COENZYME (an organic molecule!</strong>). Can classify these coenzymes (organic molecules that are cofactors) as either prosthetic groups (prosthetic groups = the coenzyme remains associated with the enzyme, like heme in Mb & Hb!) OR <strong>cosubstrates </strong>(cosubstrates = coenzymes that may enter AND exit the active site [reminder: [almost] each enzyme has an active site where catalysis [chemical reaction] takes place!] [Clarification: Almost every enzyme has an active site WHERE: the substrate binds AND catalysis [the chemical reaction] occurs!] in the course of the reaction).</p>](https://assets.knowt.com/user-attachments/6ccda270-cb23-4bd8-9c02-db2203768120.png)
Cofactors table
Catalytic Mechanisms
3 catalytic mechanisms: acid-base catalysis, covalent catalysis, metal ion catalysis. ALL catalytic mechanisms involve one or more of these three types. Remember, the transition state is often unstable, so the role of enzyme is to help stabilize that transition state intermediate.
Clarification: Chat: Catalysis is the process by which a substance called a catalyst speeds up a chemical reaction without prematurely being permanently consumed or changed in the reaction.

Acid-base Catalysis
Involves transfer of H+. Some enzymes use acid catalysis, some use base catalysis, and some use a combination of the two. Top schematic: We have a chemical reaction where we’re converting a ketone to an enol. Our transition molecule is in brackets; it involves an unstable carbanion. We will see how we will use catalysis to stabilize this intermediate!
Acid catalysis (H-A) is capable of donating a proton. Important to recognize that this is an amino acid chain on the enzyme. In the bottom schematic, you see in the intermediate, our acid catalyst has done its job of donating the proton (that blue proton!) to neutralize the negative charge on that oxygen atom. BEFORE that final product forms, however, that acid group must extract a proton from the OTHER end of the intermediate SO THAT it REGENERATES the acid form. Enzymes are catalysts! => means they cannot be permanently modified in the course of the reaction. If we started with an acid group protonated, we have to end with protonating that same acid! NOTE! In the bottom schematic, the acid regenerated HAS a red proton!!!
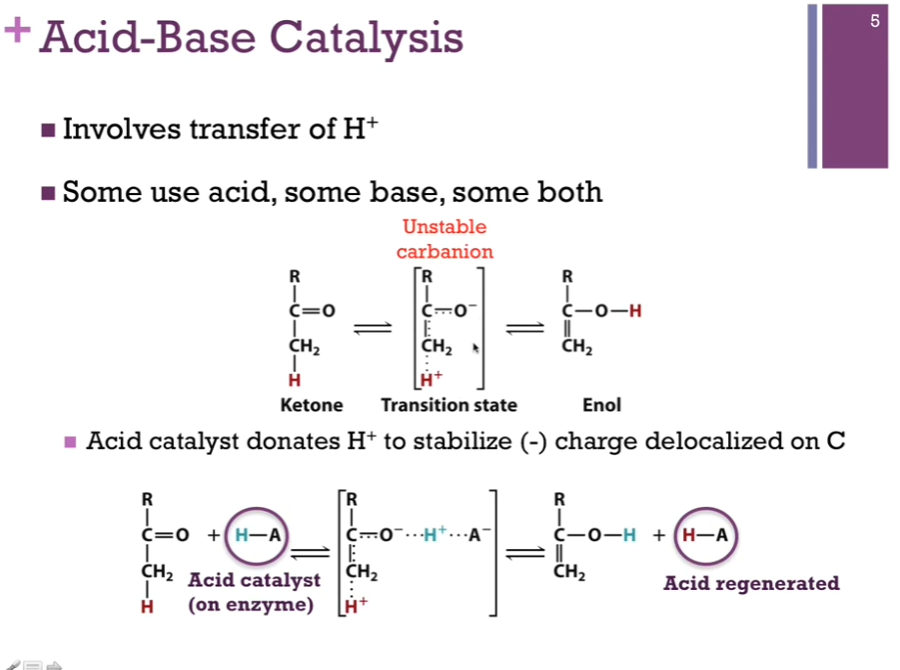
Acid-Base Catalysis
If it carries a proton, it can be an acid catalyst. If it’s lacking that proton, it can play the role of a base catalyst and accept a proton!
Slide: Base catalyst donates electron pair to stabilize H+.
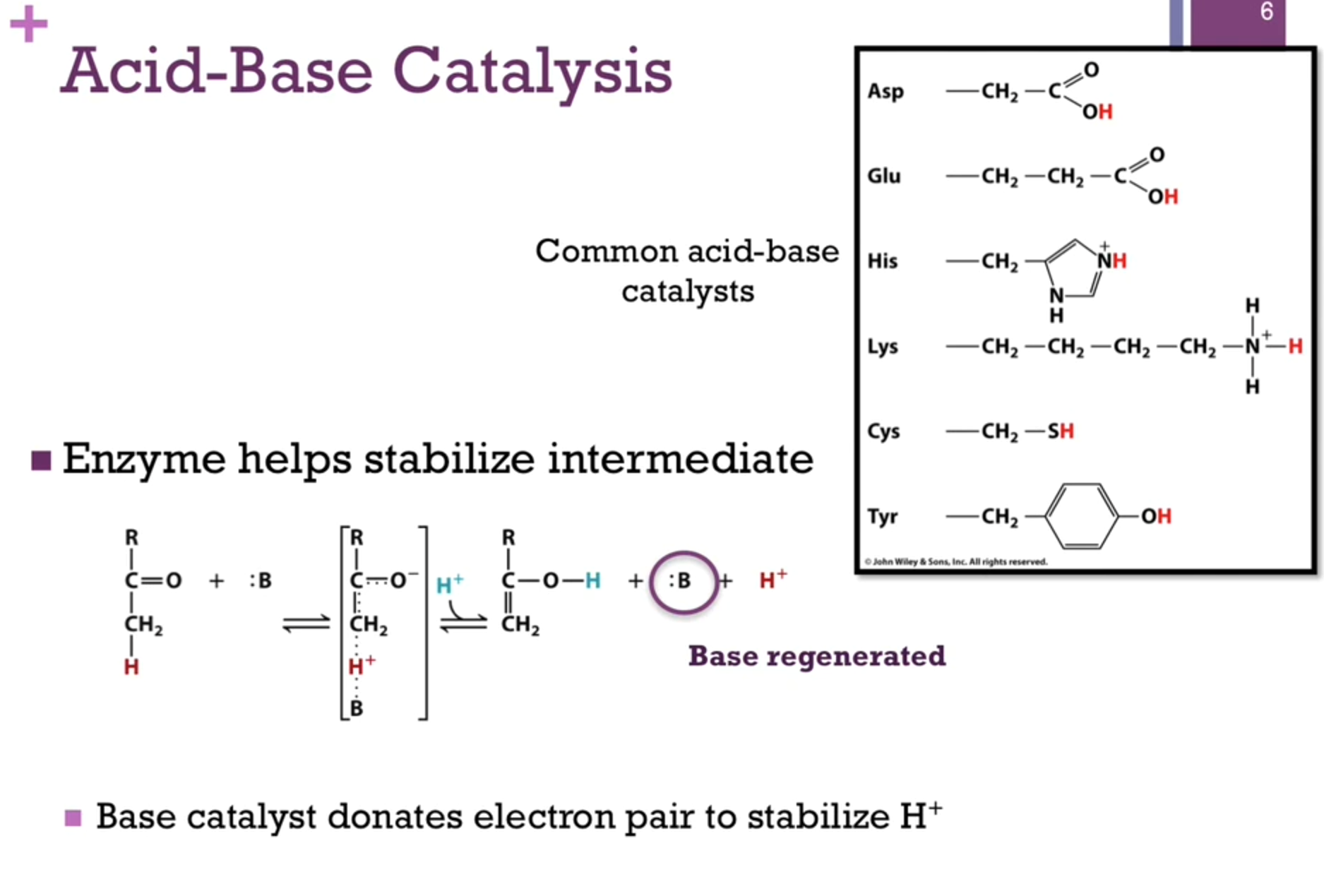
Covalent Catalysis
Case where the enzyme CYCLE between two states. In the first step, it actually forms a covalent bond with the substrate or a portion of the substrate. In the second step, it must break that bond because remember, we must regenerate the original form of the enzyme. Here’s a reaction where we are converting acetoacetate through decarboxylation to acetone! Unstable enolate intermediate, another carbanion.
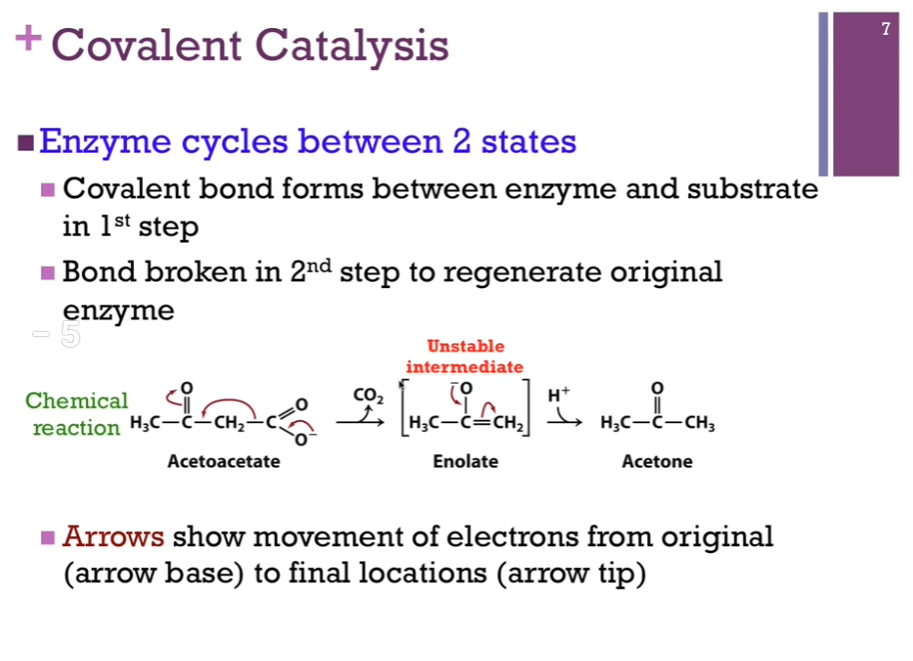
Covalent Catalysis
Substrate = acetoacetate. Primary amine will play the part of our covalent catalyst; reminder! this is an amine group side chain on our enzyme! We have nucleophile (acetoacetate) and the electron rich nitrogen atom (on our primary amine). Our primary amine will attack our carbonyl carbon atom that carries a partial positive dipole. In the process, we form this imine (see schematic!) (Schiff base) (= our intermediate!) (a C=N) and release hydroxide.
We’re starting to form that covalent bond? We decarboxylate. Get another intermediate after (away CO2 arrow). Then, we get structure on the bottom of the screen. Notice, we have our product, but it is covalently attached to the enzyme. Have to release the product.
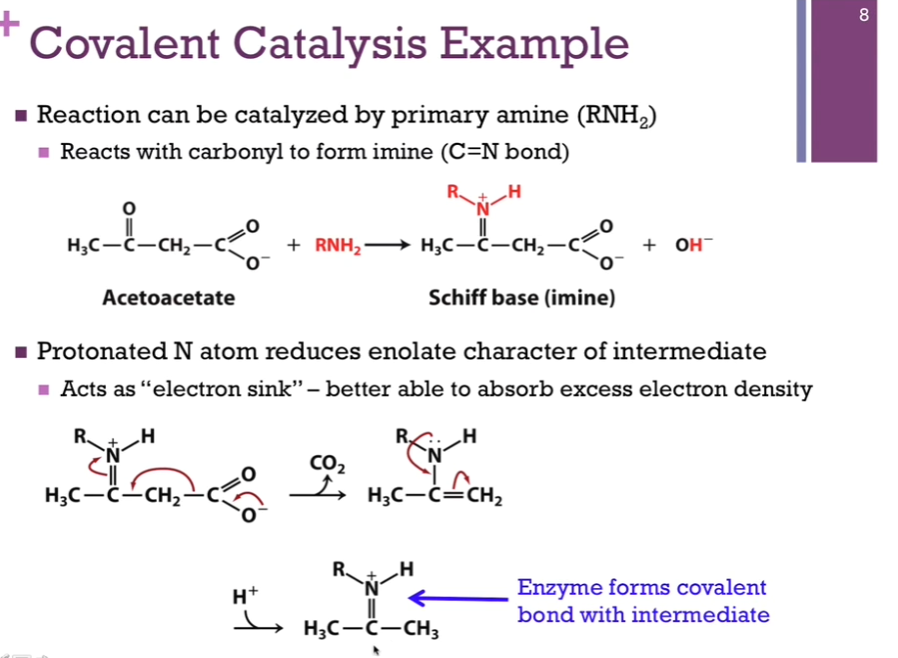
Covalent Catalysis
Our Schiff base has our product covalently linked to that amino acid side chain. A hydroxide ion that will react with that to form our final product, acetone, and we have regenerated the amine form of our SIDE CHAIN!
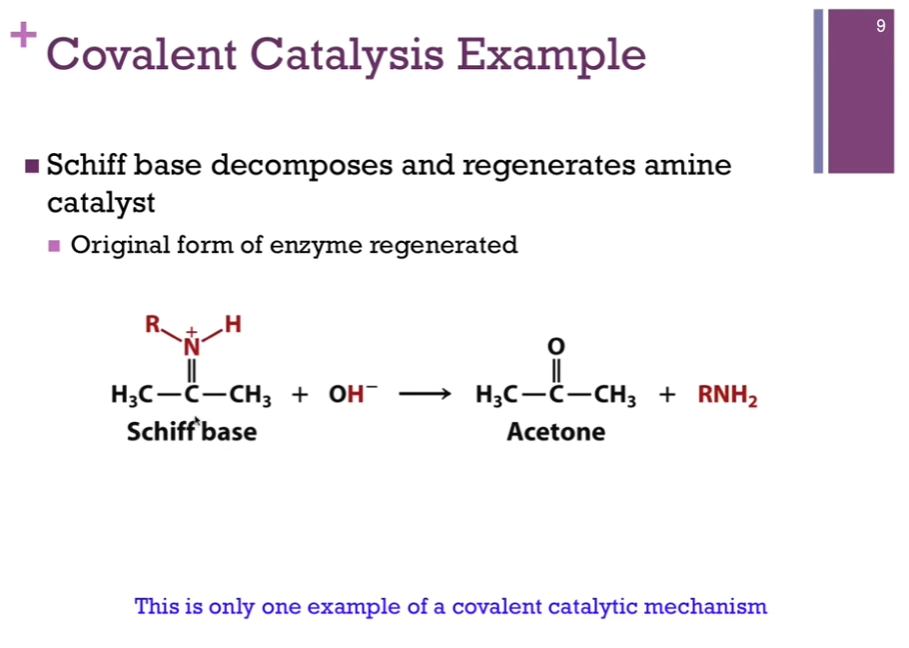
Covalent Catalysis
Need nucleophile (electron-rich) to stabilize carbonyl group.
Attack an electrophile.
Nucleophiles common in covalent catalysis: Ser, Tyr, Cys, Lys, His.
Electrophiles: Protons, metal ions, carbonyl carbon atoms, cationic imine (Schiff base).
Look at schematic!
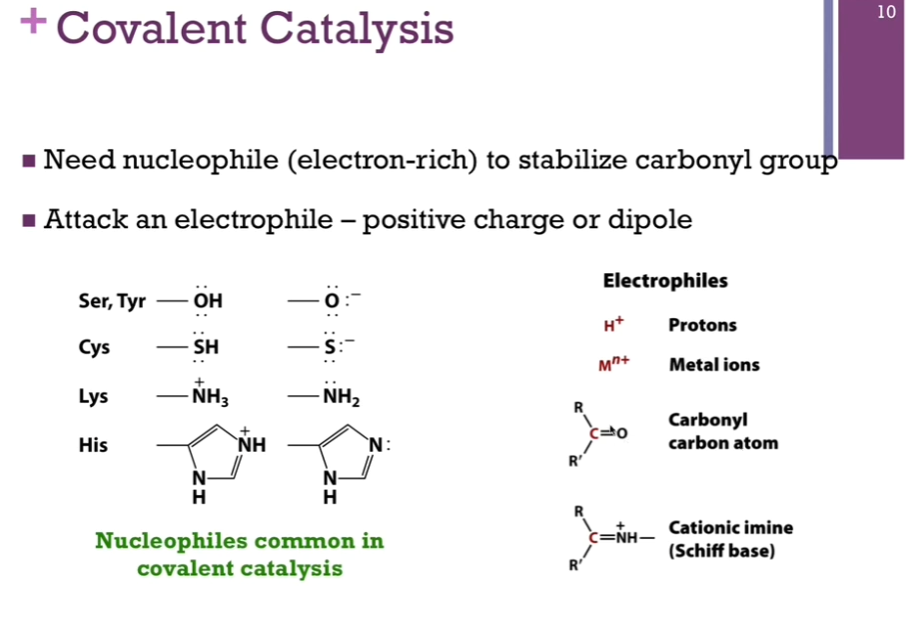
Covalent Catalysis
Covalent catalysis is a 2 part reaction! 2 energy hills! One first energy hill to form Schiff base, represents the hill to form our covalent bond (that hill = deltaG1doubledagger) (first intermediate is X1doubledagger). AFTER we form that covalent bond with our substrate and covert it to product, we have to break that bond; that’s our second energy hill! (that second hill = deltaG2doubledagger) (second intermediate = X2doubledagger).
NOTE “Intermediate” on the diagram! The valley BETWEEN the two hills represents the point at which we formed that covalent bond with the substrate or product before it’s released. It is at a lower energy level that EITHER of our two intermediates (X1doubledagger and X2doubledagger) and yet it is at a higher energy level that either the reactants or products.
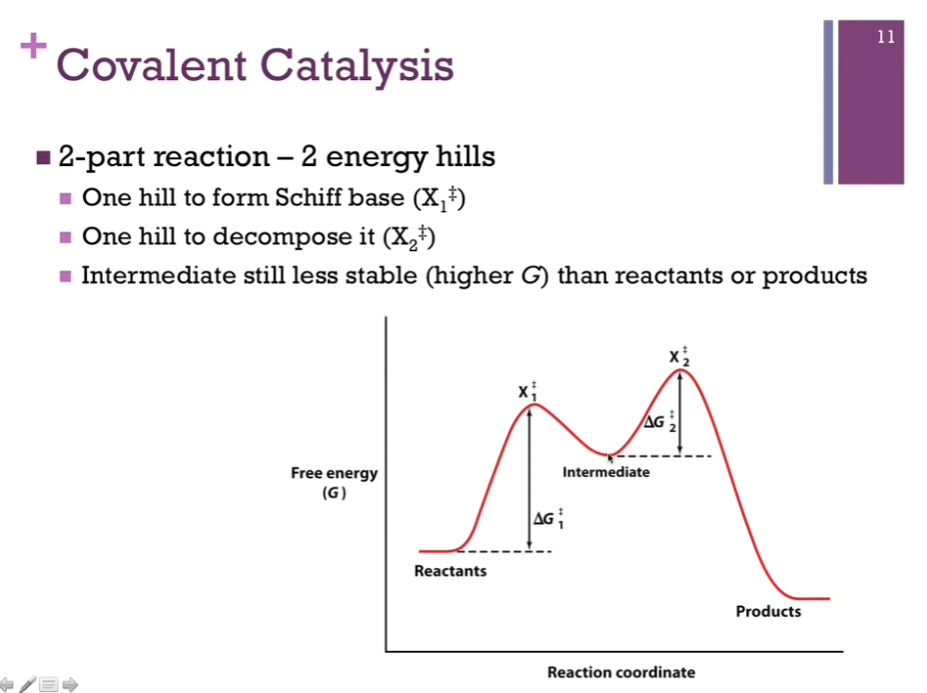
Metal Ion Catalysis
Slides: Metal Ion catalysts mediate oxidation-reduction (redox) reactions (movement of 1 or 2 electrons), may interact directly with substrate, or may stabilize charge of transition state!
Schematic: Have a positively charged zinc ion, and it’s stabilizing our carbanion intermediate.
Chymotrypsin Mechanism
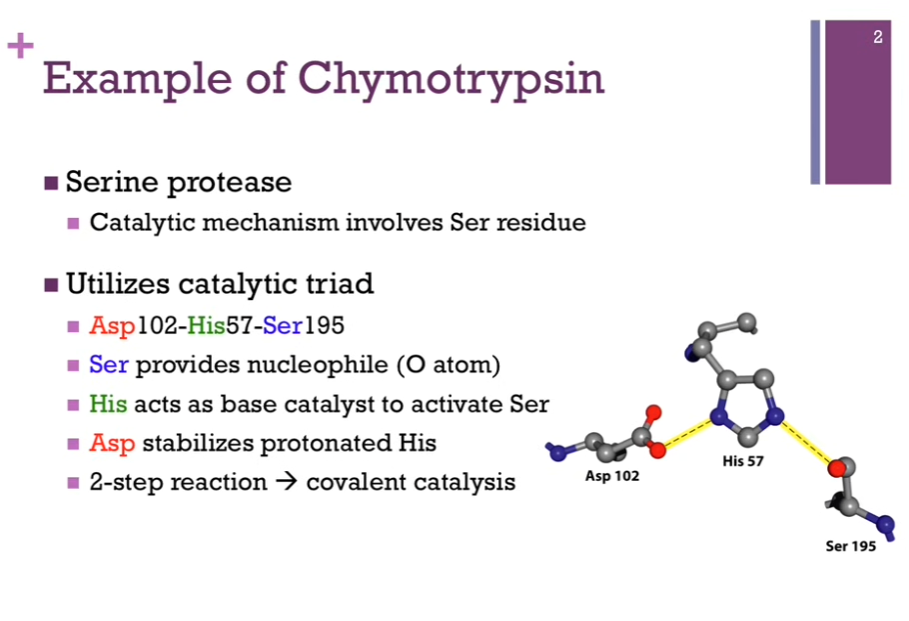
Mechanism of the digestive enzyme, chymotrypsin. Good example for us to consider since in the course of it’s reaction, we’ll see several examples of different types of catalysts. Chymotrypsin is part of a large family of Ser proteases (proteases = digest proteins AKA it breaks peptide bonds!).
Chymotrypsin always involves a Serine residue. On left = catalytic triad that’s part of the active site!
Catalytic triad = Asp 102, His 57, and Ser 195! (HAS!). Their numbers indicate their position within their primary structure. Can see they are widely separated from each other in the primary structure BUT when the enzyme adopts it final tertiary fold, these amino acids are perfectly positioned within the active site.
Ser provides the nucleophile (O atom).
His acts as base catalyst to ACTIVATE Ser.
Asp stabilizes protonated His.
2-step reaction => covalent catalysis (enzyme forms a covalent bond with its substrate).
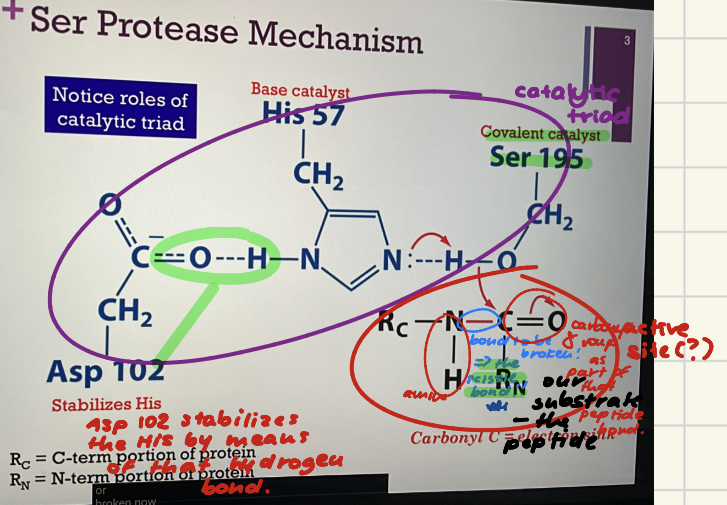
Ser Protease Mechanism
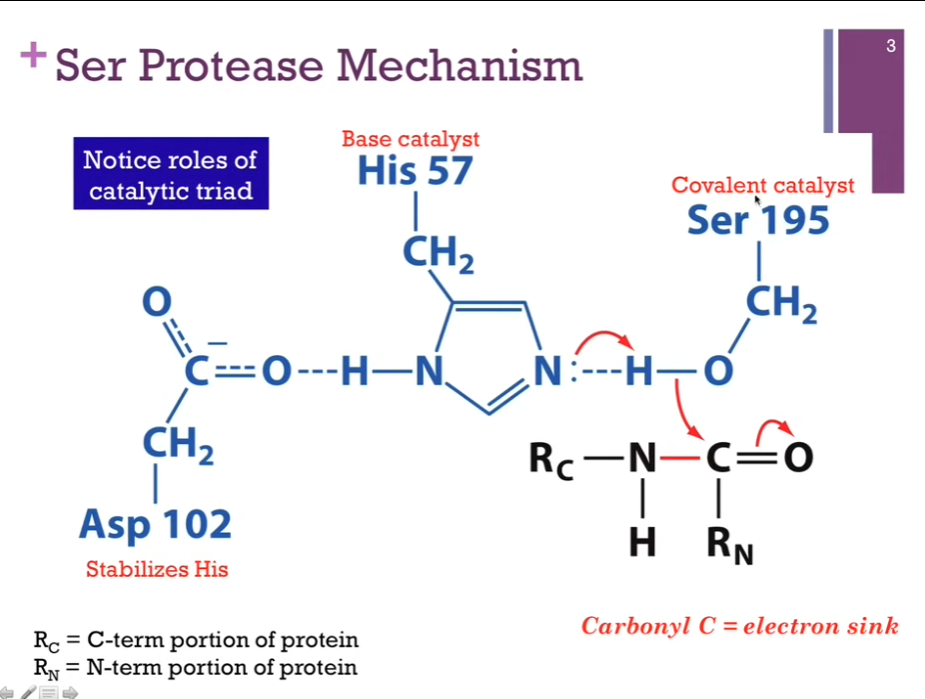
Catalytic triad in blue. Active site (?). Covalent catalyst = Ser 195. Base catalyst = His 57. Stabilizes His = Asp 102 (by hydrogen bond).
Substrate in black. Peptide bond to be broken is in red. Call the bond to be broken = the scissile (like scissors) bond. Interesting!!
R_C = The C terminal part of our peptide. R_N = the N terminal part of our peptide. Our peptide bond is somewhere within the peptide. Step 1.) His acts as a base catalyst to extract a proton from Serine. As it does so, that H will break its bond with oxygen!! and that allows the oxygen atom to form a NEW bond with carbon!! Slides: 1.) His EXTRACTS H+ from Serine to make it more nucleophilic; Ser-O- attacks carbonyl of peptide bond!!
Ser Protease Mechanism
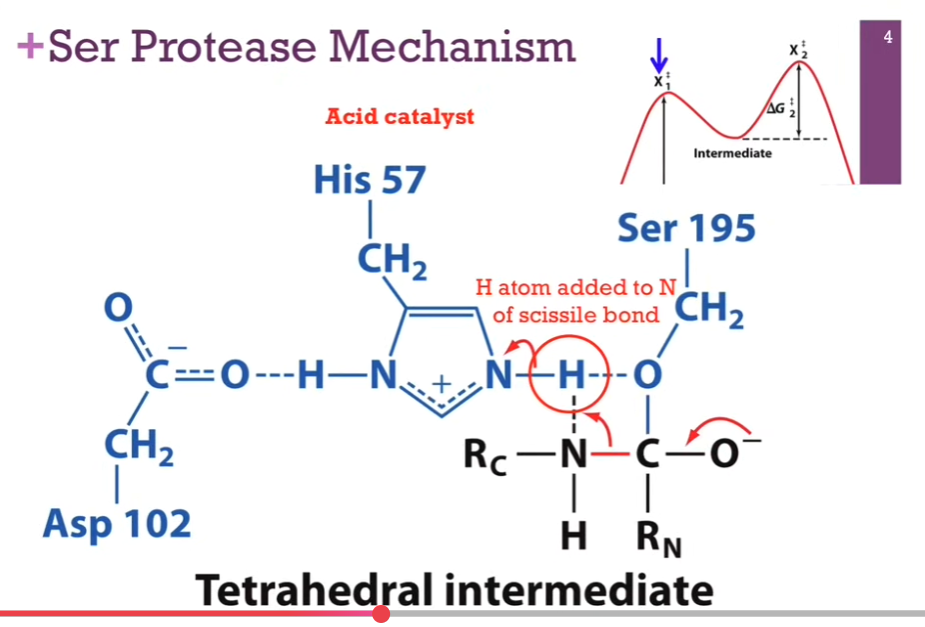
Resolved Structure from previous slide. N has formed a bond with H. His carries a POSITIVE charge because it was a proton! We have broken the bond between O and H, but it does form a hydrogen bond!! (THAT’S BETWEEN THE O and H; the dashed LINE!!). The oxygen on the serine has formed a new COVALENT bond with the carbonyl carbon! Notice that carbonyl carbon is now a tetrahedral intermediate. Also an unstable carb anion. Unstable tetrahedral intermediate (X1doubledagger).
Ser Protease Mechanism
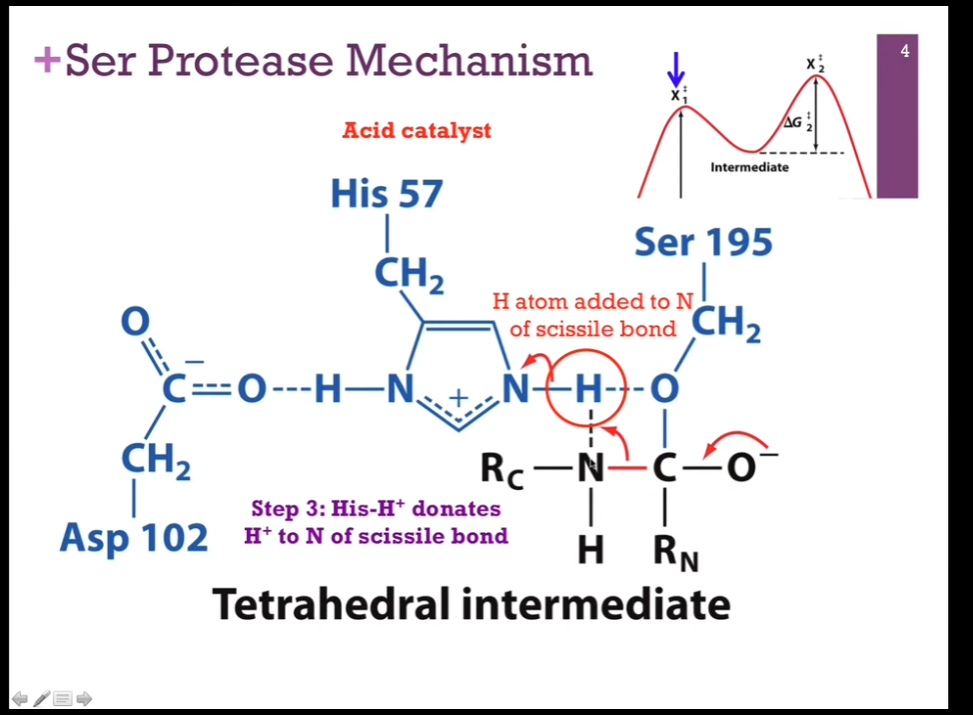
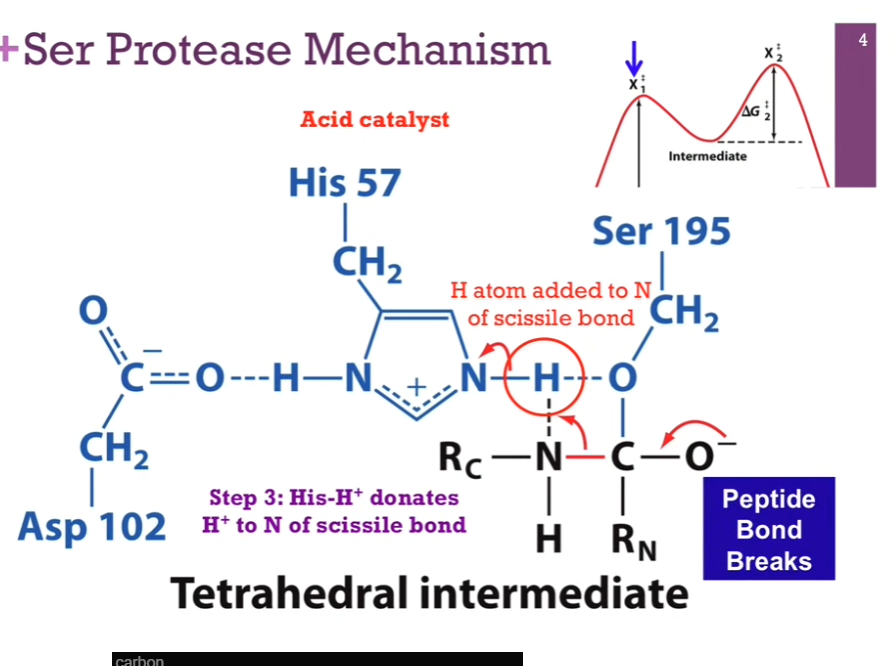
Slides: Step 3: His-H+ DONATES H+ to N of the SCISSILE BOND!
NEXT STEP => His is going to act as an acid catalyst!! It has that proton it can donate, but INSTEAD of giving it back to Serine, it is going to donate it to the NITROGEN THAT’S PART of that SCISSILE BOND that we’re going to break (LOOK at schematic!). As it does so, that N-H bond will break (check schematic!!), it will receive back its electron and again be neutral. Will form a NEW bond between the N in the PEPTIDE bond (careful and look at schematic!!!) and that hydrogen atom (LOOK at schematic!). For that N to form that BOND with hydrogen, it must BREAK its bond with CARBON!!!!!!! (LOOK AT SCHEMATIC!!).
Ser Protease Mechanism
Slide: Peptide bond breaks!
Once we have done so, we will BREAK that peptide bond! Note once this reaction sequence ends, we will have added a H atom TO THAT peptide bond! So when we break that bond, we will release that C-terminal part of our peptide AND form a new AMINE terminus on that portion of the peptide (interesting wording!!).
Ser Protease Mechanism
Slide: [PEPTIDE] Bond cleaved. R-C portion released. Rest of peptide covalently bonded to Ser.
Slide: Acyl-enzyme intermediate. Relatively stable!
Formed that covalent bond. Histamine back in its original form. Now, we have a covalent intermediate, which we need to resolve.
Valley between our 2 hills.
![<p>Slide: <strong>[PEPTIDE] Bond cleaved.</strong> R-C portion released. Rest of peptide covalently bonded to Ser.</p><p>Slide: <strong>Acyl-enzyme intermediate.</strong> Relatively stable!</p><p>Formed that covalent bond. Histamine back in its original form. Now, we have a covalent intermediate, which we need to resolve.</p><p>Valley between our 2 hills.</p>](https://assets.knowt.com/user-attachments/5f8a151c-e586-443a-85e9-472e623e0eb6.png)
Ser Protease Mechanism
Next series of steps is a REPEAT of what we have just seen JUST that the nucleophile is different!
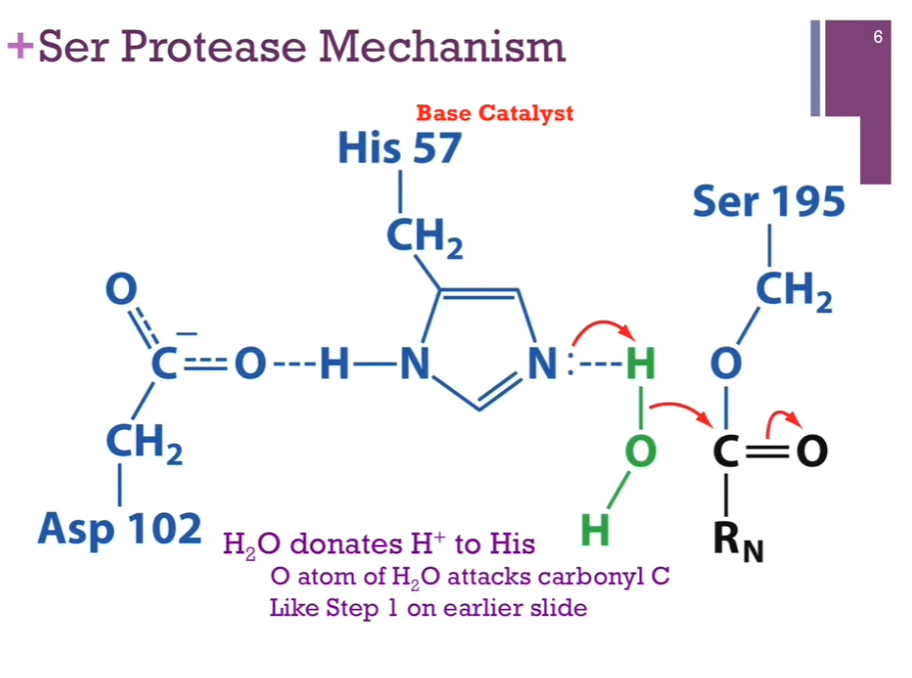
Slide: H2O donates H+ to His. O atom of H2O attacks carbonyl C. Like Step 1 on earlier slide.
Have His acting as a base catalyst but NOW our nucleophile is WATER! His extracts a H+ from water. Form a new N-H bond. Allow oxygen in WATER to act as a NUCLEOPHILE to attack our carbonyl carbon. Like step one in earlier slide.
Ser Protease Mechanism
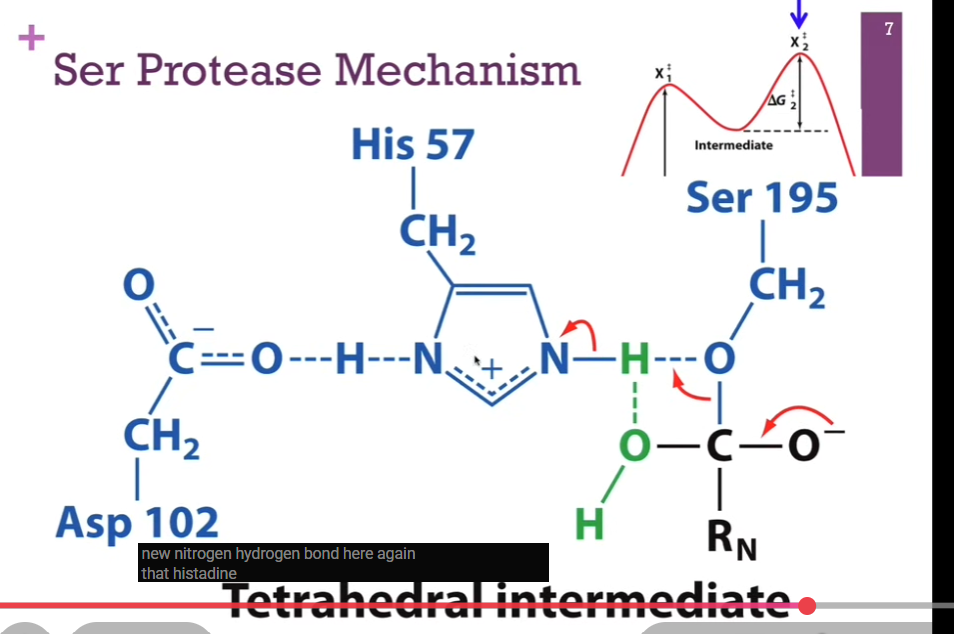
Have our new N-H bond. His carrying a positive charge. Remember, it’s stabilized by this His hydrogen bond! (look at schematic!). Now, we have a C-O bond with out PEPTIDE here! SO, now we have a new tetrahedral intermediate. Carbonyl carbon is now again a tetrahedral intermediate, an unstable carb anion, our SECOND energy hill, intermediate = X2doubledagger. Must resolve the structure.
Ser Protease Mechanism
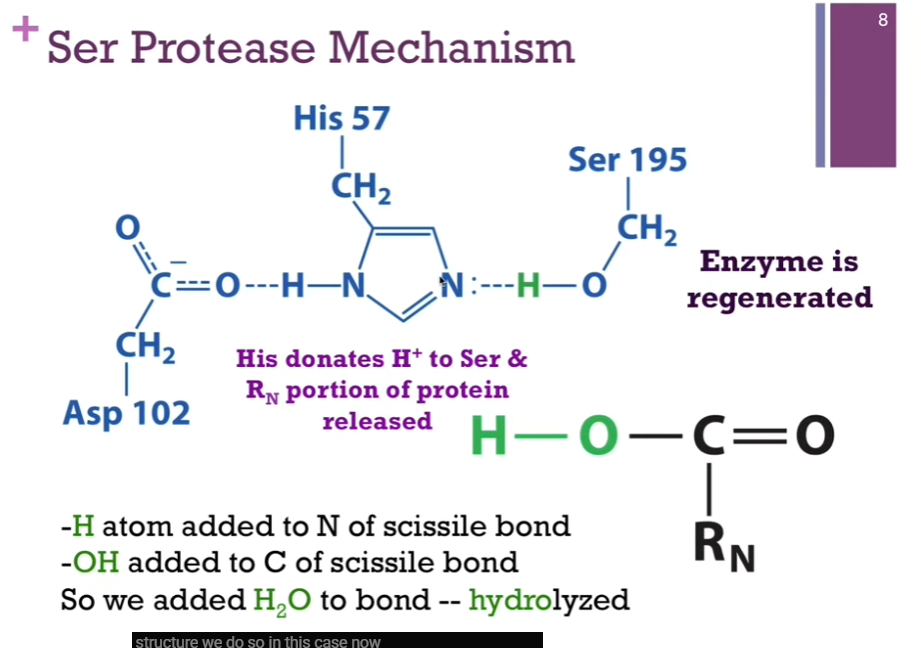
His can now DONATE that proton BACK to SERINE. We have accomplished the goal of breaking the peptide bond, and the active site looks just like it did when we began. SO, the enzyme has been regenerated! Here is our N-terminal of our peptide that has been released (look at schematic!). Note: In the first case, we had an H atom added to N of the scissile bond! IN THIS CASE, we added an -OH to the CARBON of the scissile bond! SO, overall, we simply added water to the bond. It was hydrolyzed; THAT is, LYSED by adding water.
Ser Protease Mechanism
Covalent catalyst = Ser 195.
Base catalyst = His 57.
Stabilizes His = Asp 102.
Scissile bond
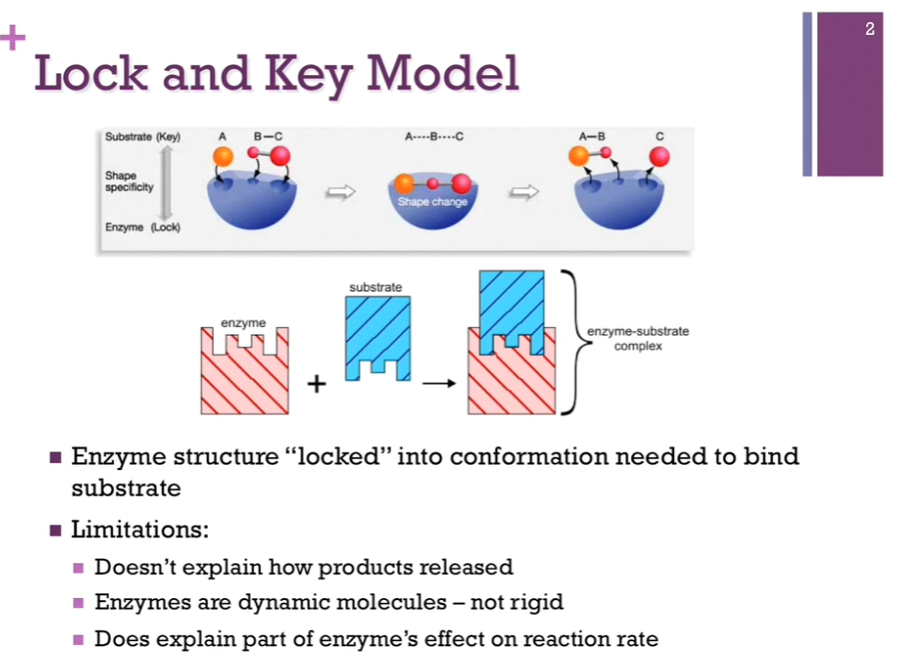
Lock and Key Model
Lock and key model of substrate binding. Enzymes were locked into the conformation they wanted to be in order to bind substrate. Top schematic: enzyme = in blue. It’s in perfect conformation in which it can bind substrate. In this model, the enzyme is the lock, and the substrates are the key that so perfectly fits that lock!
Limitations: Doesn’t explain how products are released. Also, we know enzymes are NOT rigid structures; they often change conformation in the course of a reaction! It also doesn’t explain how the enzyme affects the reaction rate!
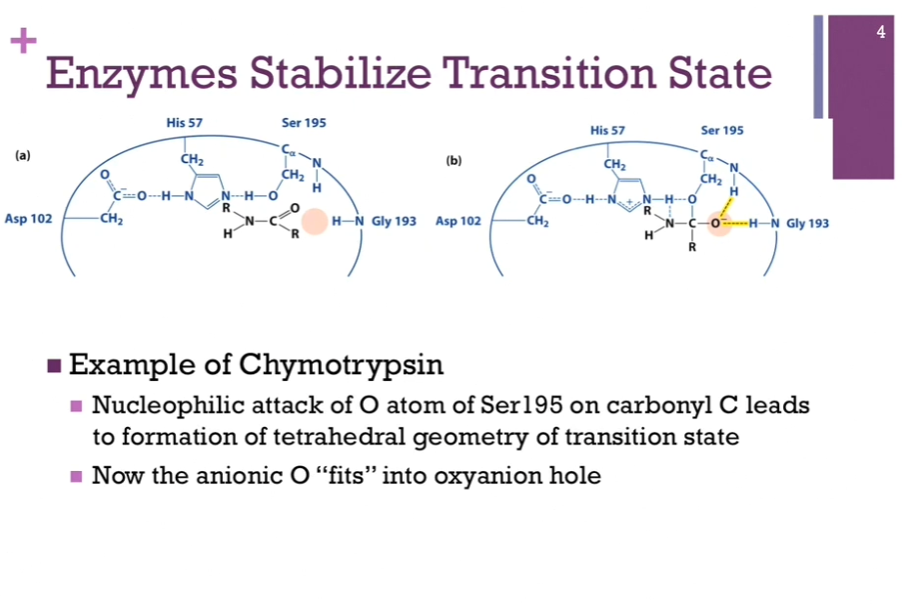
Enzymes Stabilize Transition State
Proposed by Linus Pauling.
Suggests that enzymes help to STABILIZE the transition state. AKA enzymes increase the reaction rate by binding tightly to and thereby stabilizing the transition state! Remember: our transition state is the midpoint between our substrates and products (Clarification: The structure of the transition state has characteristics of both the substrate and the product. For example, suppose a reaction breaks a bond: Substrate: A-B. Product: A + B.). SO, at that point, we are STRAINING the substrates to form products.
In this model, the enzyme is still the lock, but the key that fits the lock perfectly is the TRANSITION.
Example of Chymotrypsin:
HERE we have the catalytic triad in the active site. OUR peptide = in black!! (look at schematic!). Peptide bond between the N and C being the scissile bond that we’re going to break. At this point, the enzyme has BOUND the substrate, but no reaction has occurred! Notice the carbonyl carbon has trigonal geometry. In this model of SUBSTRATE BINDING, we see that the carbonyl oxygen (in black) is NOT close enough to the amine group of this glycine residue to have ANY contact whatsoever! Peach color oval area = referred to as the oxyanion hole!

Enzymes Stabilize Transition State
Left = our bound substrate. No reaction occurred.
Right = once we form that tetrahedral intermediate, our carbonyl carbon is now TETRAHEDRAL! It now carries that oxyanion! Now it FITS perfectly into that area called the oxyanion hole! It is also NOW able to form H-bonds with some of those peptide backbones!
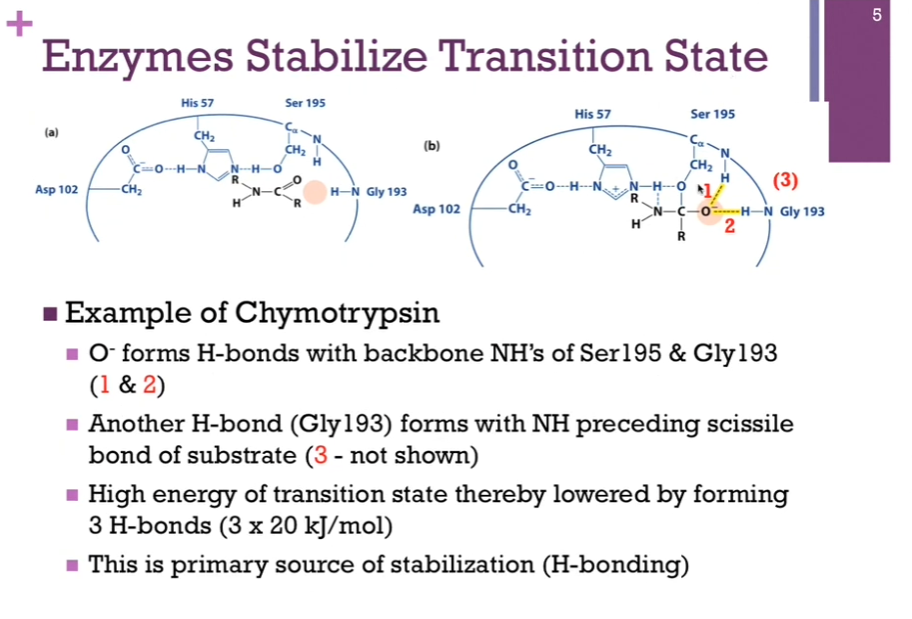
Enzymes Stabilize Transition State
1.) can form a H-bond with the peptide amine group in our active Ser195 residue.
2.) can form a H-bond with the amine group in the peptide bond of this Gly194 residue.
3.) can form a H-bond with carbonyl group of Gly193 AND the amine group preceding the scissile bond of the SUBSTRATE (not shown in the illustration here!).
Suffice to say there are three hydrogen bonds that formed in the process of INTERACTING with this transition state. Each of those H-bonds average ~20 kJ/mol. 60 kJ total. AKA our HIGH ENERGY transition state has been reduced by 60 kJ/mol in order to form those hydrogen bonds. This is the PRIMARY source of stabilization! These THREE hydrogen bonds!
Enzymes Stabilize Transition State
Our best SUPPORTING actor => Asp102. Role in the catalytic triad is to stabilize that His residue by FORMING a hydrogen bond.
Remember the HYDROGEN bond (top molecule), we have a hydrogen atom between two EN element (H is between O and N!!). It’s COVALENTLY bound to one (paying attention to top molecule is confusing? I would have thought the O and H bond would have a covalent bond and is closer together/stronger according to clarification below?). Dotted, highlighted bond = the hydrogen bond with the other EN atom. In a typical hydrogen bond, the covalent bond being stronger means that THAT would represent a shorter bond length, and a further distance away from the other EN element.
This H bond that forms between Asp and His, however, is a LOW barrier hydrogen bond! AKA (bottom molecule), the H bond is equidistant between the O and the N. So, the distance between that H atom and its hydrogen bonding partner oxygen is CLOSER than it otherwise would be (?) (Clarification: For a TYPICAL H bond between an O and a N: O-HdashdashN. The H is much closer to the oxygen it is covalently attached to [makes sense!]. The O-H bond is short and strong. The HdashdashN interaction is longer and weaker. BUT in a low barrier hydrogen bond: OdashdashHdashdashN. The O-H and H-N distances become mote similar. The bond is stronger than a typical hydrogen bond!). Since the bond length is SHORTER, the bond strength is STRONGER (makes sense) (stronger by 3-4X). By forming this low barrier hydrogen bond, we have decreased our energy hill transition state even further! => this is the SECONDARY source of stabilization!
![<p>Our best SUPPORTING actor => Asp102. Role in the catalytic triad is to stabilize that His residue by FORMING a hydrogen bond. </p><p>Remember the HYDROGEN bond (top molecule), we have a hydrogen atom between two EN element (H is between O and N!!). It’s COVALENTLY bound to one (paying attention to top molecule is confusing? I would have thought the O and H bond would have a covalent bond and is closer together/stronger according to clarification below?). Dotted, highlighted bond = the hydrogen bond with the other EN atom. In a typical hydrogen bond, the covalent bond being stronger means that THAT would represent a shorter bond length, and a further distance away from the other EN element. </p><p>This H bond that forms between Asp and His, however, is a LOW barrier hydrogen bond! AKA (bottom molecule), the H bond is equidistant between the O and the N. <strong>So, the distance between that H atom and its hydrogen bonding partner oxygen is CLOSER than it otherwise would be</strong> (?) (Clarification: For a TYPICAL H bond between an O and a N: O-HdashdashN. The H is much closer to the oxygen it is covalently attached to [makes sense!]. The O-H bond is short and strong. The HdashdashN interaction is longer and weaker. BUT in a low barrier hydrogen bond: OdashdashHdashdashN. The O-H and H-N distances become mote similar. The bond is stronger than a typical hydrogen bond!). Since the bond length is SHORTER, the bond strength is STRONGER (makes sense) (stronger by 3-4X). By forming this low barrier hydrogen bond, we have decreased our energy hill transition state even further! => this is the SECONDARY source of stabilization! </p>](https://assets.knowt.com/user-attachments/2b914dd8-69c3-4030-8e44-154c29cc896e.png)
Enzymes Stabilize Transition State
Energy level for our uncatalyzed transition state (Gdoublecrossuncat!!).
Start at some energy level, very unstable intermediate AND NOW, we are going to use some of that ENERGY to form a H bond! By doing so, we decrease the energy of that transition state! Form another H bond, and we reduce it even further! And third H bond, and we reduce energy level even further! Reduced energy state considerably by using some of the energy OF that transition state to form those hydrogen bonds! AND now, we have our LOW BARRIER hydrogen bond! So => Gdoublecrosscat = much lower than it would be!
Primary Stabilization: 3 H-bonds.
Secondary Stabilization: Low barrier H-bond.
Good example of how enzymes stabilize the transition state!!

Substrate Specificity
How do enzymes with SIMILAR CATALYTIC mechanisms recognize different substrates?
Chymotrypsin, trypsin, elastase (divergent evolution!). They all catalyze the same reaction, hydrolysis of a peptide bond and in fact, similar structures (look at backbone trace of slide!). Active site residues are highlighted by the black oval. Practically overlap! Practically identical tertiary structure and the same catalytic mechanism AND YET they recognize different substrates!!
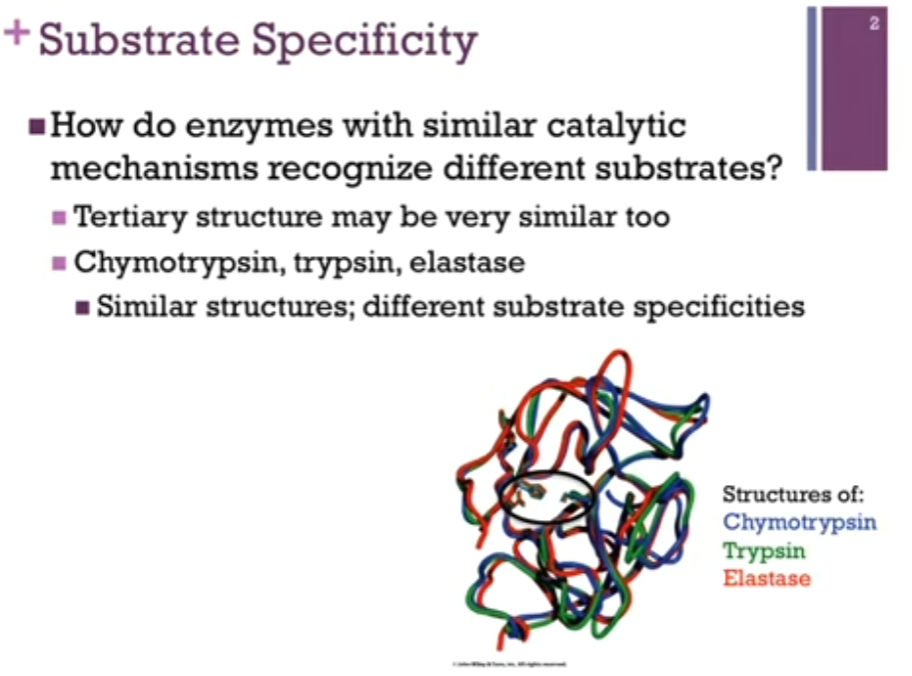
Substrate Specificity
Also find enzymes with different tertiary structures and yet active site residues are the SAME!!!
True for => subtilisin (in grey!) (different structure from chymotrypsin, trypsin, and elastase) yet the active site residues are the same (look at ovaled on slide!). Note: convergent evolution!
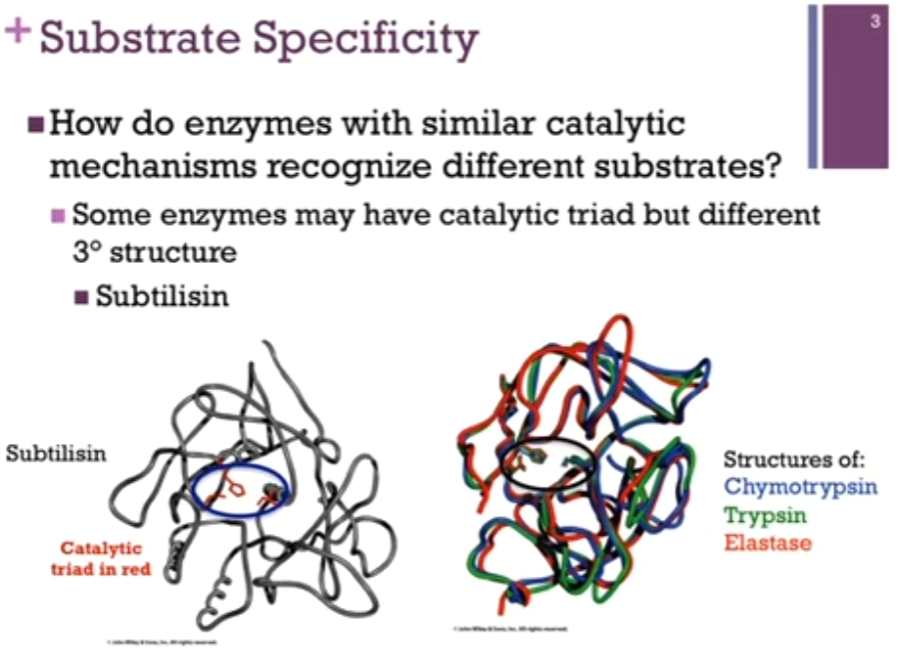
Substrate Specificity
Enzymes vary in the specificity pocket!
At least in the case of these 3 enzymes.
AKA they vary in the amino acids that form that pocket: controls the chemical environment and dimensions of that pocket (that is, the amino acid residues that comprise that pocket!).
First example = Chymotrypsin! Specificity pocket in red! Can see Gly residues on the sides of that pocket. Gly = smallest amino acid; gives us the largest dimensions on the pocket => allows us to ACCOMMODATE large AR rings! (like Phe!). Note the Ser residue at the bottom of the pocket (Ser = polar but no charge there so as to repel the very nonpolar Phe sidechain!).
Let’s look at the RELATIONSHIP between the specificity and the active site. Have Phe sidechain fitting nicely in our SPECIFICITY pocket. Here is SCISSILE bond, the ONE that is going to FIT IN THE active site to be hydrolyzed!!). SO the sidechain sits in the pocket AND that perfectly positions that scissile bond IN THE ACTIVE SITE. AKA the SPECIFICITY pocket is DIFFERENT than the active site but adjacent to it.
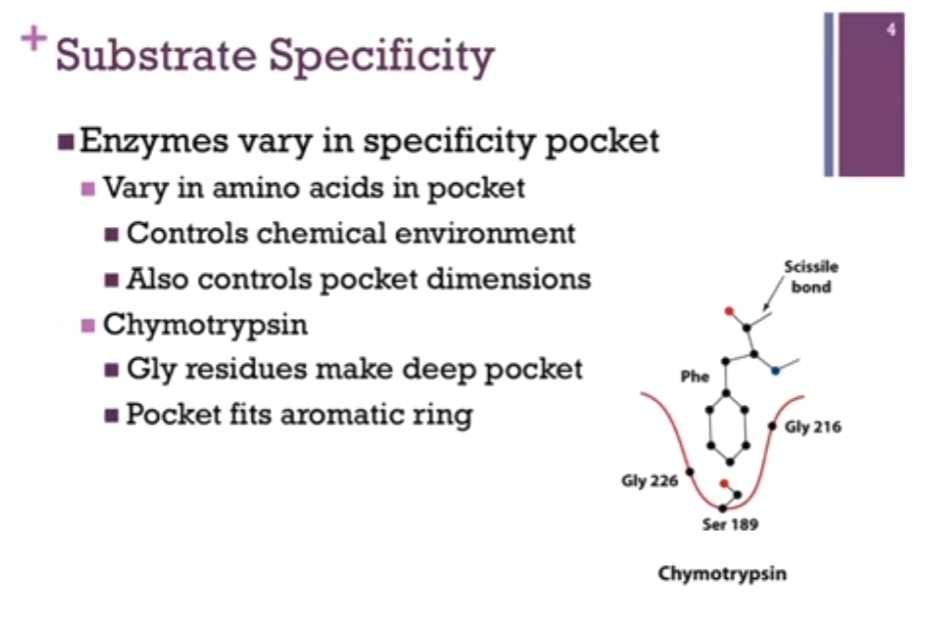
Substrate Specificity
Enzymes vary in the specificity pocket!
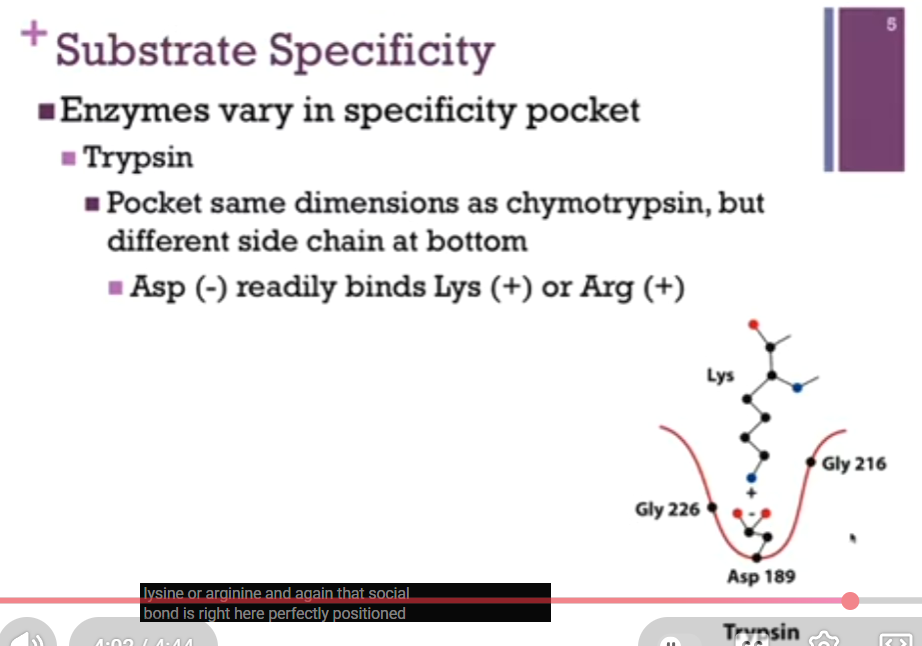
Second example = Trypsin!
In this case, the pocket has the SAME dimensions as chymotrypsin because there are glycine side chains that outline that pocket, but there is a different side chain at the bottom of the pocket, Asp! At physiological pH, that Asp carries a negative charge! SO, the dimensions of that pocket can accommodate a BULKY group BUT with that negative charge, it will no longer bind a nonpolar group (like Phe in chymotrypsin!). Instead, it’s more likely to bind a bulky basic residue such as Lys or Arg. That scissile (note spelling!) bond (second image!) is perfectly positioned within that active site.
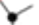
Substrate Specificity
Enzymes vary in the specificity pocket!
Third example = Elastase!
THERE are bulkier hydrophobic (Great Ambitions Vanish Limitations Tired Inspired Future Medical Professionals; Glycine, Alanine, Valine, Leucine, Threonine, Isoleucine, Phenylalanine, Methionine, Proline) groups on the walls of the pocket and that minimizes the dimensions, the pocket is smaller! SO, it accommodates smaller nonpolar side chains!
So, it is the specificity in those specificity pockets that determines the side chains that fit into the pocket AND therefore the peptide bonds that are hydrolyzed WITHIN the active site!!
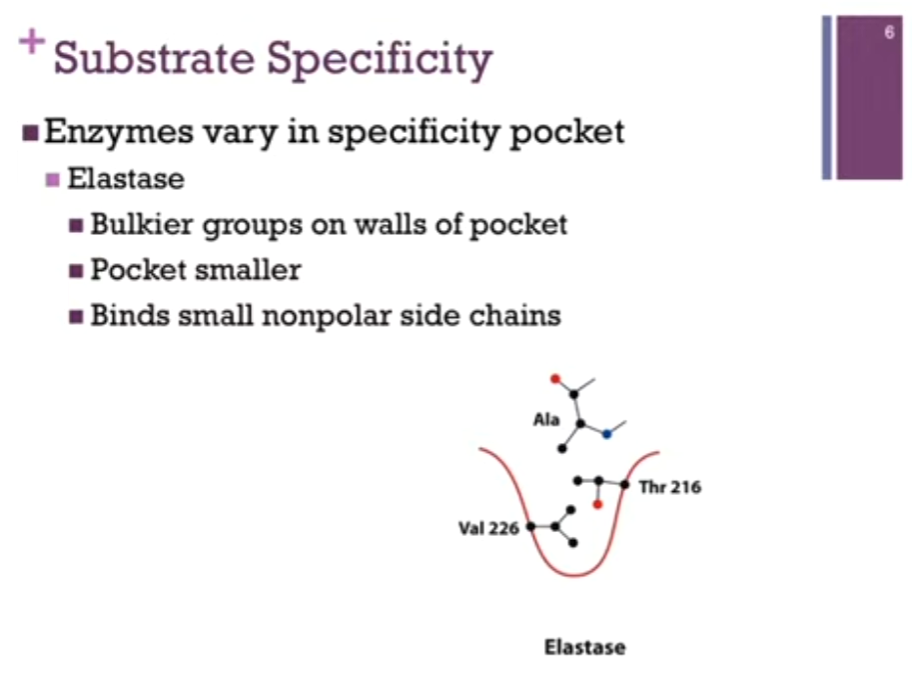
Other Enzyme Effects
All enzymes work by lowering the activation energy and making it MORE likely that we will form MORE product per unit time. AKA => they lower the energy of the transition state; they stabilize the intermediate.
(a) Reactants have to bump into each other through simple diffusion for the reaction to occur in the absence of the enzyme.
(b) In the presence of the enzyme, the enzyme provides a platform that we can bring these reactants or SUBSTRATES together; that’s the PROXIMITY effect! They’re close to one another.
There is also the orientation effects! We’re going to position them with respect to one another so that we can facilitate the chemistry that’s going to occur and so enzymes help with this process; it often involves conformational changes.
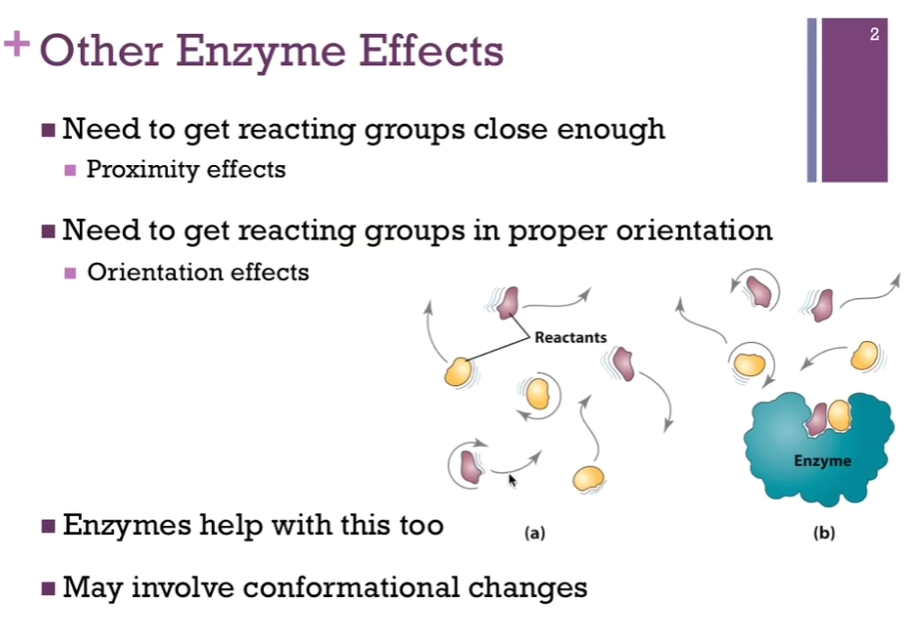
Induced Fit Model
Hexokinase; this is the enzyme that catalyzes the transfer of a phosphate from ATP to glucose to form Glucose-6-phosphate (upper left schematic). This is the first reaction in glycolysis! In the right schematic, we have the space filling model of hexokinase in blue. Left portion of right schematic, hexokinase in blue, active site (hole area) as if it has its mouth open and ready to receive the substrate, which is glucose in red. Once it binds that substrate (right portion of the panel), there’s a conformational change in the enzyme, and it closes down over that substrate. Called the induced model!! because of the binding of substrate induces this conformational change. This helps to position the ATP for easy transfer of that phosphate group to glucose.
Induced Fit Model
Also provides a protected environment for this phosphorylation! As if that hexokinase closes its arms so tightly around that glucose and ATP substrate (?) that there’s no room for anything else! NOT EVEN water can get in. Important. ATP CAN be spontaneously hydrolyzed to release Pi and ADP. If this occurs, we have not been successful. We have not transferred that PHOSPHATE to glucose; we would’ve just broken a bond without actually carrying out the chemistry we’re interested in. SO, it becomes important that we exclude water so that we don’t have this competition for the actual catalytic reaction we want to carry out.

Induced Fit Model
Also find true for many SUBSTRATES and that is certainly true for all CARBOHYDRATES like glucose, they have many -OH groups and so, they are very highly hydrated. If, in the absence of this conformational change, to carry out the chemistry, we would have to rearrange all of those water molecules so that could get to the atoms and perform the chemistry we want to perform but by causing this induced fit, by closing down over the SUBSTRATE, we are essentially forcing OUT the water; now we no longer have the energy barrier that we need to rearrange those solvent molecules, so it has eliminated that problem for us!
Slides: Eliminates energy barrier caused by ordered solvent molecules: Substrate molecules most likely hydrated; Must shed water to FIT IN active site; Eliminates energy barrier of rearranging solvent molecules.
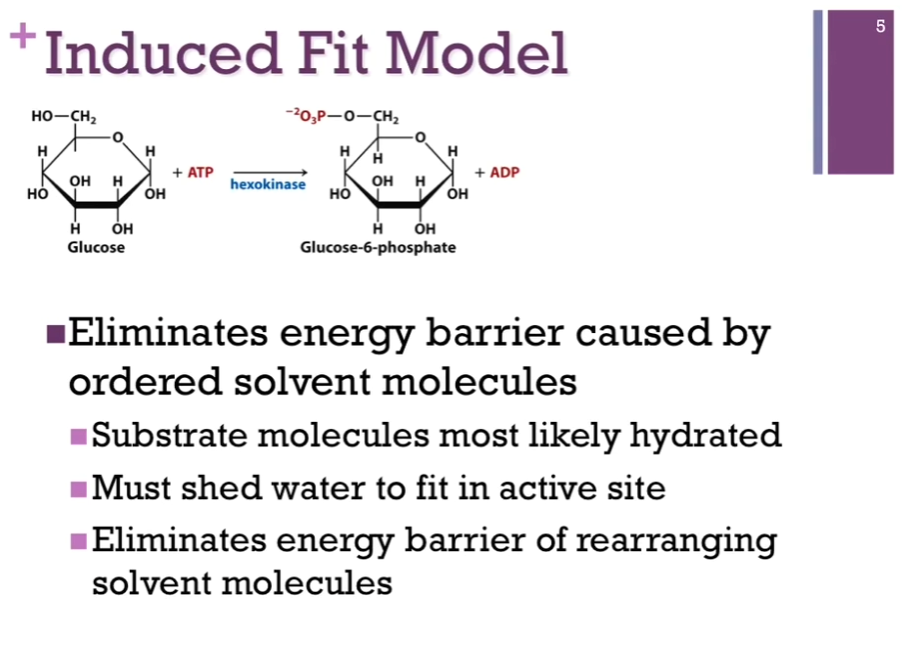
Induced Fit Model
This is referred to as ELECTROSTATIC CATALYSIS. AKA: the electrostatic interactions are more pronounced in the absence of water. Want to remember that water has a high dielectric constant; that is, it’s a charged insulator. To carry out the reaction, you have to those electrostatic interactions, you can’t AFFORD to have water present because it will INSULATE those charges and make the reaction less likely to happen => what referred to as electrostatic catalysis.
Thus, enzymes might also eliminate other substances that might interfere.

Activation of Chymotrypsin
We find that chymotrypsin is a good example of proteases that are synthesized as inactive precursors (Clarification: Chymotrypsin is made by the body in an inactive form first, and only later is it converted into the active enzyme. Why make it inactive? Chymotrypsin is a protease, meaning it cuts peptide bonds in proteins. If it were active immediately after being synthesized in the pancreas, it could start digesting the pancreas’s own proteins and damage the tissue. To prevent this, the enzyme is produced as in inactive precursor, called a zymogen [or proenzyme]. Specific example: The inactive precursor is: Chymotrypsin => (activation) => Chymotrypsin).
In other words, as it’s synthesized off the ribosome, it is inactive and requires to become activated as an enzyme. These are called zymogens (Chat: Zymogens, also called proenzymes, are inactive forms of enzymes) because they CAN be generated as active enzymes (?). AKA they are activated only when and where they are needed. Some activate themselves (autoactivation); active form of the enzyme converts an inactive to an active form.
![<p>We find that chymotrypsin is a good example of proteases that are synthesized as inactive precursors (Clarification: Chymotrypsin is made by the body in an inactive form first, and only later is it converted into the active enzyme. Why make it inactive? Chymotrypsin is a protease, meaning it cuts peptide bonds in proteins. If it were active immediately after being synthesized in the pancreas, it could start digesting the pancreas’s own proteins and damage the tissue. To prevent this, the enzyme is produced as in inactive precursor, called a zymogen [or proenzyme]. Specific example: The inactive precursor is: Chymotrypsin => (activation) => Chymotrypsin). </p><p>In other words, as it’s synthesized off the ribosome, it is inactive and requires to become activated as an enzyme. These are called <strong>zymogens </strong>(Chat: Zymogens, also called proenzymes, are inactive forms of enzymes) because they CAN be generated as active enzymes (?). AKA they are activated only when and where they are needed. Some activate themselves (autoactivation); active form of the enzyme converts an inactive to an active form. </p>](https://assets.knowt.com/user-attachments/222a7684-5da8-46c3-b85d-7a3566e0f463.png)
Activation of Chymotrypsin
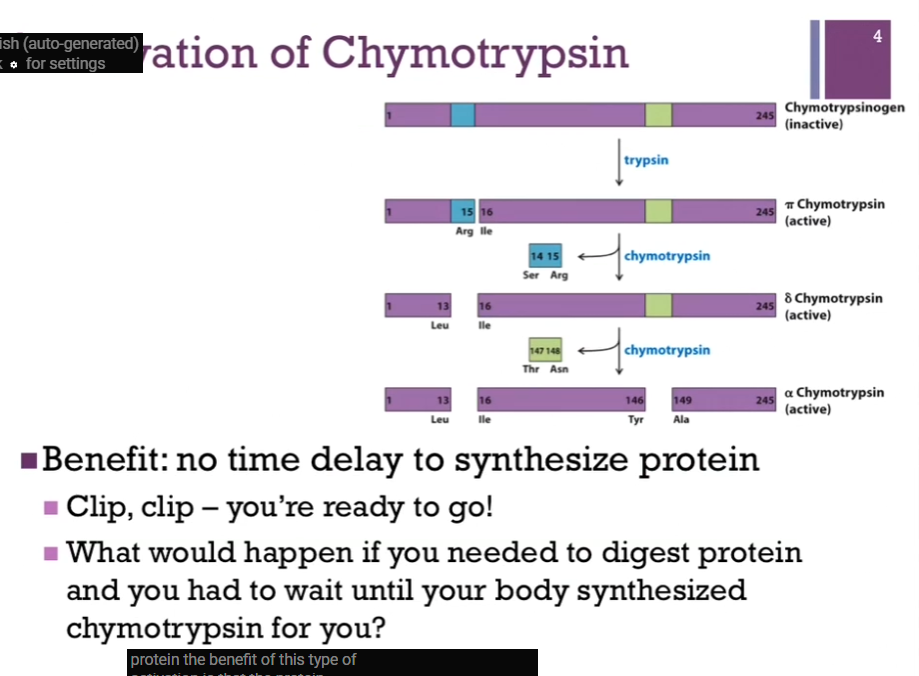
As synthesized from the ribosome, it is chymotrypsinOGEN. This is the zymogen form (yes!). As can be seen from the top of the screen, it contains 245 amino acid residues. First event to occur is hydrolysis by trypsin (look at schematic!). That breaks the peptide bond between residues 15 and 16 (Arg and Ile residues) and that converts inactive Chymotrypsinogen to active PI chymotrypsin. At this point then, chymotrypsin takes over the activation and autoactivates itself. In the first step, it’s going to clip out a two amino acid dipeptide (Ser and Arg residues, 14 and 15) and that converts from PI to DELTA chymotrypsin. Note: both of these are active forms. In the final step, Chymotrypsin does one more cut (in lime green), it’s going to CUT out the dipeptide (Thr and Asn, residues 147 and 148). AND here we have the final form alpha Chymotrypsin.
Also note: (in schematic) although [alpha] Chymotrypsin here looks like there are THREE separate polypeptides, they are all peptide fragments that are connected by DISULFIDE bonds, so it’s all still part of the same protein!
Slides: Trypsin converts inactive to active (pi) chymotrypsin. Chymotrypsin further activates itself: All forms active (pi, delta, alpha).
![<p>As synthesized from the ribosome, it is chymotrypsinOGEN. This is the zymogen form (yes!). As can be seen from the top of the screen, it contains 245 amino acid residues. First event to occur is hydrolysis by trypsin (look at schematic!). That breaks the peptide bond between residues 15 and 16 (Arg and Ile residues) and that converts inactive Chymotrypsinogen to active PI chymotrypsin. At this point then, chymotrypsin takes over the activation and autoactivates itself. In the first step, it’s going to clip out a two amino acid dipeptide (Ser and Arg residues, 14 and 15) and that converts from PI to DELTA chymotrypsin. Note: both of these are active forms. In the final step, Chymotrypsin does one more cut (in lime green), it’s going to CUT out the dipeptide (Thr and Asn, residues 147 and 148). AND here we have the final form alpha Chymotrypsin. </p><p>Also note: (in schematic) although [alpha] Chymotrypsin here looks like there are THREE separate polypeptides, they are all peptide fragments that are connected by DISULFIDE bonds, so it’s all still part of the same protein! </p><p>Slides: Trypsin converts inactive to active (pi) chymotrypsin. Chymotrypsin further activates itself: All forms active (pi, delta, alpha).</p>](https://assets.knowt.com/user-attachments/57fb69c2-325d-4e86-973d-0aab0e4e81da.png)
Activation of Chymotrypsin
The benefit of this type of activation is THAT the protein has already been synthesized, and it’s ready to go. It only has to be activated! Benefit cause imagine in order to have active chymotrypsin, we had to actually synthesize the protein from scratch. Ate a big meal. Remember chymotrypsin digest peptide bond, SO we are going to need that enzyme. Eaten meal. Now we have to send signal to nucleus so that we can convert DNA into mRNA. Message needs to be transported to the cytoplasm so that it can be translated into protein and then the protein has to be sent where it needs to go. Well now that hamburger is sitting in the bottom of your stomach; now you have indigestion cause you had to wait to make the protein. Benefit of inactive form of protein or enzyme is that it’s there when you need it; all you need to do is activate it (clip here here and there!). Then have active chymotrypsin and can digest that protein, that hamburger we ate!
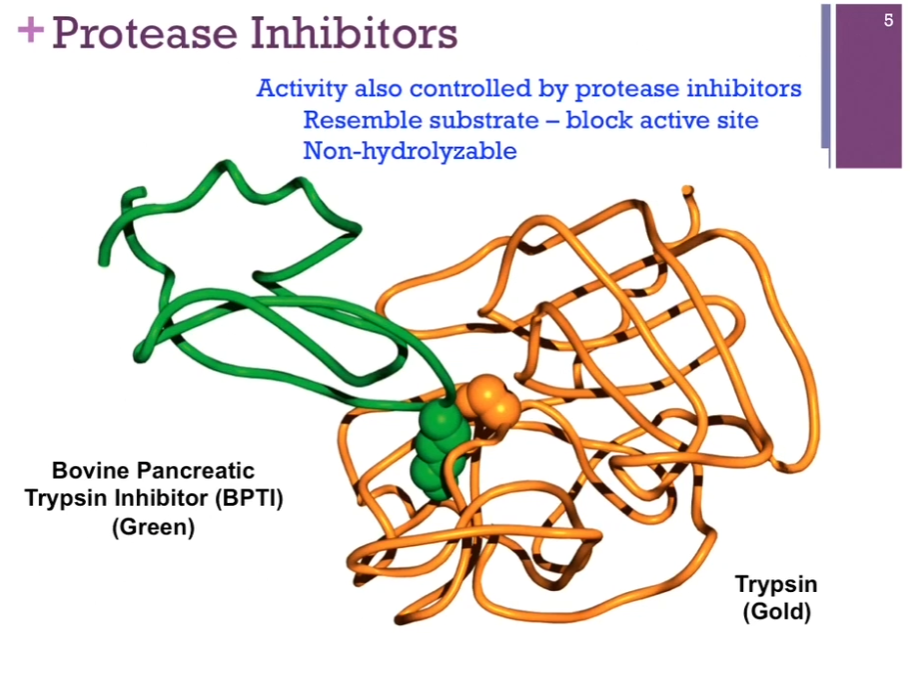
Protease Inhibitors
Reminder: Chymotrypsin is a PROTEASE, meaning it is an enzyme that breaks peptide bonds in proteins.
Another way PROTEASES (enzymes that break peptide bonds in proteins!) are controlled is by protease inhibitors. Good example = enzyme trypsin in gold here. Trypsin inhibitor in green; resembles SUBSTRATE closely so it can bind to the active site, but there is no reaction. AKA it blocks the activity of the enzyme.
Biochemical Cascades
Chymotrypsin activation cascade is a very common theme in biological systems! Another good example = the clotting cascade (eventually lead to formation of fibrin clots [in electron micrograph]).
Thrombin initiates the sequence of events. Converts inactive XI to active XIa and so forth down the line until we get to the final molecule fibrin. 10 molecules of thrombin => activates 10 molecules to form XIa; by now, we have amplified our effect ten fold! If we do that for every step of the cascade; because there are SO many steps; one molecule, thrombin, may be responsible for producing ONE MILLION molecules of fibrin (interesting?). By having this cascade, several steps, this cascade of events, we can produce a very large effect in a very small time frame.
Don’t need to know steps of clotting cascade.
![<p>Chymotrypsin activation cascade is a very common theme in biological systems! Another good example = the clotting cascade (eventually lead to formation of fibrin clots [in electron micrograph]). </p><p>Thrombin initiates the sequence of events. Converts inactive XI to active XIa and so forth down the line until we get to the final molecule fibrin. 10 molecules of thrombin => activates 10 molecules to form XIa; by now, we have amplified our effect ten fold! If we do that for every step of the cascade; because there are SO many steps; one molecule, thrombin, may be responsible for producing ONE MILLION molecules of fibrin (interesting?). By having this cascade, several steps, this cascade of events, we can produce a very large effect in a very small time frame. </p><p>Don’t need to know steps of clotting cascade. </p>](https://assets.knowt.com/user-attachments/c8ae7aa3-3c7c-4841-aa6b-e6767f840755.png)
Biochemical Cascades
There’s an amplifying effect in the biochemical cascades.

Lipids (fats)
Lipids are insoluble in water due to the hydrophobic effect (Review: The exclusion of nonpolar substances from an aqueous solution is known as the HYDROPHOBIC EFFECT. The hydrophobic effect REDUCES the solubility of NONPOLAR molecules because water prefers to hydrogen-bond with itself rather than organize around substances it cannot interact with!!! Chat: The hydrophobic effect is the tendency of nonpolar (“water-fearing”) molecules or parts of molecules to cluster together in water rather than mix with it).
Lipids have NO defining functional groups!
SOME!! but not all lipids are amphipathic (both polar and nonpolar regions!!).

Lipids (fats)
Functions: Cellular barrier (Ch. 8): Control movement into and out of cell; Control of membrane fluidity (Ch. 8); [Cellular?] barrier are also places for proteins to dock and travel; Can also serve as waterproofing agents (wax that might be found in plant leaves) (Ch. 8-10); Can also serve as signaling agents (Ch. 10).
![<p>Functions: Cellular barrier (Ch. 8): Control movement into and out of cell; Control of membrane fluidity (Ch. 8); [Cellular?] barrier are also places for proteins to dock and travel; Can also serve as waterproofing agents (wax that might be found in plant leaves) (Ch. 8-10); Can also serve as signaling agents (Ch. 10). </p>](https://assets.knowt.com/user-attachments/8aa260a9-854d-4af0-b570-785a8c9375f9.png)
Families of Lipids
Here are the general families or classes of lipids. Waxes on the far left. Have Triacylglycerols and Glycerophospholipids (both built on glycerol backbones!). Have sphingolipids, similar in many respects to glycerophospholipids. (different backbone of sphingosine). Steroids like cholesterol are built from isoprene (a small hydrocarbon molecule with the chemical formula C5H8; it’s a colorless, volatile liquid and is one of the most important building blocks). Finally, we have eicosanoids.
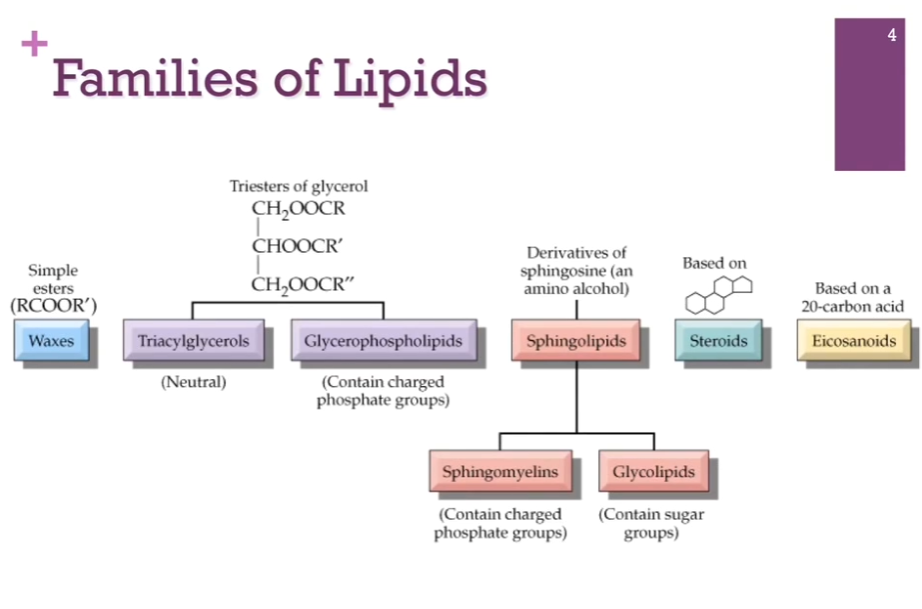
Types of Lipids
Start with the SIMPLEST type of lipid: a fatty acid! They are long chain carboxylic acids, so they have carboxylic acid groups at the end (polar region!) (look at schematic!), but they have these LONG, hydrocarbon chains at the end that are mostly nonpolar/hydrophobic. Even-numbered chains are the most common!
PALMITATE on left of schematic! 16 carbon atoms in its chain. C_16. Stearate on the right of schematic has 18 carbon atoms. FOR palmitate and STERATE, you’ll notice these are SATURATED (think Simplest case of lipids = fatty acids = all hydrogen atoms, no double bonds, all single bonds!) fatty acids. THAT is the TAILS are saturated with hydrogen atoms. They’re all single bonds, no double bonds. Has to do with how they aggregate or pack together. AND that saturation means at room temperature, these tend to be solid like butter (SIMPLEST CASE, SATURATED, SINGLE BONDS, palmitate, Stearate, SOLID).
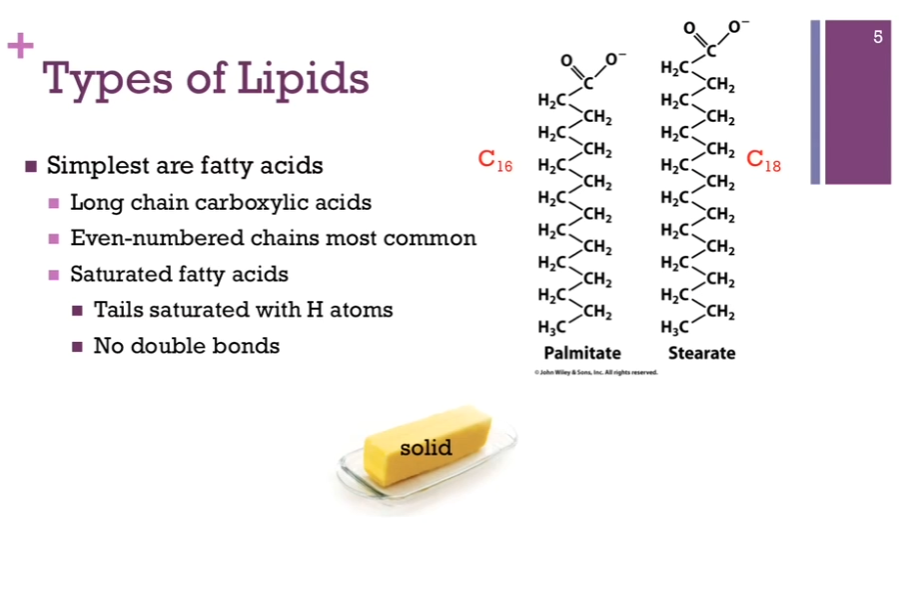
Types of Lipids
Might also have an unsaturated fatty acid. Hydrocarbon chain has one OR more double bonds! Pictured on left = oleate (Oleate = One double bond) and linoleate (has 2 double bonds). In each case, you will notice that the double bond has a CIS configuration; the H atoms are on the same side of that bond.
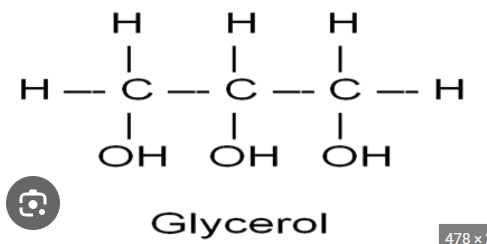
In unsaturated chains, they pack together differently that saturated chains; they tend to more loosely associated and that makes them LIQUID at room temperature! like olive oil the top here! Fatty acids because they are so very hydrophobic are not found in free form in an aqueous environment (Chat: Not entirely true for all fatty acids. Statement is mostly true for long-chain fatty acids). They ARE USUALLY bound to protein, and many times, they are usually ESTERIFIED to GLYCEROL!!! (a small 3-carbon alcohol with the formula, C3H8O3), as we’ll see!

Glycerol Structure Review
Acyl Chain Saturation
Saturated vs. Unsaturated.
Carbon diagram on the far left = GLYCEROL phospholipid! Look at chains! Have a saturated (simplest = solid = all single bonds = straight structure) chain in gray on the left; very straight structure because they are all single bonds. Have an unsaturated chain on the right, which has ONE double bond. Because of that, there’s a kink that’s introduced. What controls how they pack together.
Top right = see Saturated fatty acids = closely pack together (makes sense!). Very ordered and rigid structure, which is what makes them Solid at room temperature (butter).
Middle right = if unsaturated, they [fatty acids?] can’t pack together very well. The tend to be more disordered. These are fluid at room temperature (oils).
Bottom right = (sometimes we’ll see in lipid bilayers), there are ordered microdomains; these are referred to as LIPID RAFTS (later).
![<p>Saturated vs. Unsaturated.</p><p>Carbon diagram on the far left = <strong>GLYCEROL phospholipid</strong>! Look at chains! Have a saturated (simplest = solid = all single bonds = straight structure) chain in gray on the left; very straight structure because they are all single bonds. Have an unsaturated chain on the right, which has ONE double bond. Because of that, there’s a kink that’s introduced. What controls how they pack together. </p><p>Top right = see Saturated fatty acids = closely pack together (makes sense!). Very ordered and rigid structure, which is what makes them Solid <strong>at room temperature</strong> (butter). </p><p>Middle right = if unsaturated, they [fatty acids?] can’t pack together very well. The tend to be more disordered. These are fluid at room temperature (oils). </p><p>Bottom right = (sometimes we’ll see in lipid bilayers), there are <strong>ordered microdomains</strong>; these are referred to as LIPID RAFTS (later). </p>](https://assets.knowt.com/user-attachments/4b21c7b9-d9d1-4a11-a115-ae98db35d50f.png)
Types of Lipids
A CLASS of lipids: TRIacylglycerols (note spelling!!) (triglycerides). They are built on a glycerol backbone!
Glycerol is the top schematic! Three carbon compound, 3 hydroxyl groups, EACH hydroxyl group may be ESTERIFIED to a fatty acid chain (that is pictured on the bottom!!!). So on the bottom schematic, the glycerol backbone is in black and our 3 fatty acid chains are in blue! (btw, acyl means [likely acyl group]: acyl group = a carbonyl-containing group derived from a carboxylic acid by removing the -OH portion) (general structure: R - C(=O) -). You’ll notice there’s an ester link HERE TO that glycerol backbone!!! (saw this esterified to glycerol part in unsaturated fatty acids section!). These three fatty acid chains may be all different in terms of the length of the carbon chain and the DEGREE of saturation and that provides a lot of variety in terms of structure and function.
![<p>A CLASS of lipids: TRIacyl<strong>glycerols </strong>(note spelling!!) (triglycerides). They are built on a <strong>glycerol backbone</strong>!</p><p><strong>Glycerol is the top schematic! Three carbon compound, 3 hydroxyl groups, EACH hydroxyl group may be ESTERIFIED to a fatty acid chain (that is pictured on the bottom!!!). </strong>So on the bottom schematic, the glycerol backbone is in black and our 3 fatty acid chains are in blue! (btw, acyl means [likely acyl group]: acyl group = <strong>a carbonyl-containing group derived from a carboxylic acid by removing the -OH portion</strong>) (general structure: R - C(=O) -). You’ll notice there’s an ester link HERE TO that glycerol backbone!!! (saw this esterified to glycerol part in unsaturated fatty acids section!). These three fatty acid chains may be all different in terms of the length of the carbon chain and the DEGREE of saturation and that provides a lot of variety in terms of structure and function. </p>](https://assets.knowt.com/user-attachments/ec99e521-2c36-4680-91e4-109a8efee32e.png)
Acyl Group Structure Review
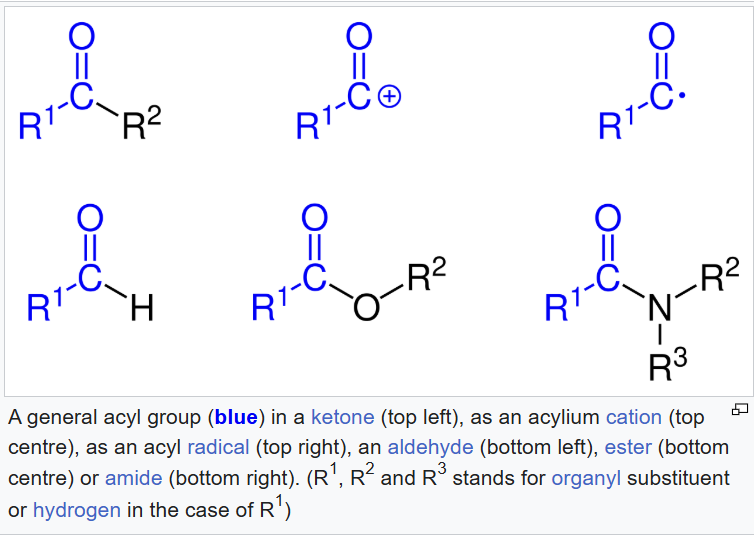
Types of Lipids
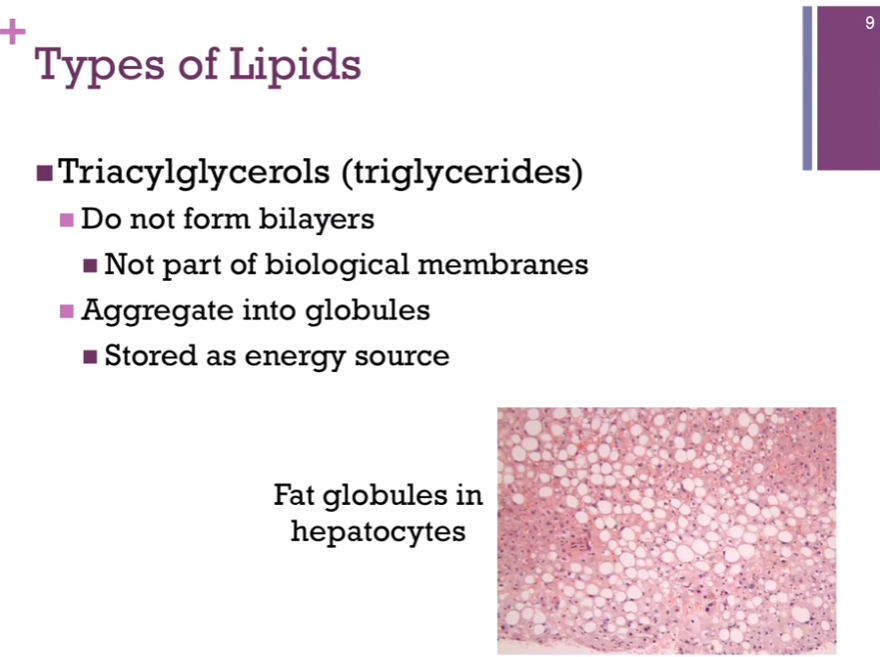
A CLASS of lipids: TRIacylglycerols (triglycerides) (continued).
Triglycerides DON’T form bilayers!! It has to do with the GEOMETRY of their structure, so they are not part of biological membranes. Triglycerides’ role: to be stored as an energy source! They AGGREGATE into globules; THAT is illustrated in the electron micrograph of hepatocytes. Those flat fat globules = white portions! Not a healthy liver. Tend to store fat long term as triglycerides. (Clarification: Hepatocytes = main cells of the liver. Adipocytes are FAT cells that STORE triglycerides. Hepatocytes store some glycogen, synthesize and break down fats, produce bile, detoxify substances, and regulate many metabolic pathways). THAT’S how it’s stored in our adipocytes.