Leseliste - kapittel 4 og 5
1/83
Earn XP
Description and Tags
Enzymer, energi, fotosyntese og celleånding
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
84 Terms
Metabolisme
Inni alle celler er det kjemisk aktivitet, metabolisme, et stoffskifte som går ut på å bryte ned og bygge opp molekyler – enten eksoterme eller endoterme reaksjoner i metabolismen.
Eksoterme og endoterme reaksjoner
Eksoterme vil si at de avgir energi. Endoterm vil si at de tar opp energi. I en eksoterm reaksjon har altså produktene mindre energi enn reaktantene, mens i en endoterm reaksjon har produktene mer energi enn reaktantene. Eksoterme reaksjoner gir fra seg varmeenergi, endoterme reaksjoner tar opp varmeenergi. Mange eksoterme reaksjoner må tilføres en viss mengde aktiveringsenergi for å kunne komme i gang.
Anabolisme
Oppbygningsprosesser. I oppbygnigsprosesser skjer det en rekke kjemiske reaksjoner som krever energi. Hensikten er å sette sammen molekyler slik at de danner andre og gjerne større molekyler.
Katabolisme
Nedbrytningsprosesser. I nedbrytningsprosesser blir energi frigitt når molekylene reagerer og danner stadig nye og mindre forbindelser. Noen kjemiske reaksjoner som frigir energi, er svært langsomme og tar lang tid. Det er fordi de molekylene som deltar i reaksjonen, trenger en viss mengde energi før reaksjonen kan skje. Den energimengden som er nødvendig for at reaksjonen kan begynne, kaller vi aktiveringsenergien eller startenergien.
Aktiveringsenergi/startenergi
Den energimengden som trengs for at en reaksjon kan begynne. Vi kan sammenlikne aktiveringsenergien med energi vi tilfører ved som skal brenne og deretter frigi varmeenergi. Først må veden få tilført litt energi fra en fyrstikk, deretter avgir veden store mengder energi. Det må være slik at reaksjonen ikke kommer i gang før startenergien blir tilført.
Enzymer - bygning og funksjon
Enzymer er helt avgjørende for livet på jorda. Inni alle levende organismer skjer det forskjellige kjemiske reaksjoner, eksoterme og endoterme. Med enzymer til stede kan reaksjonene gå flere millioner ganger raskere fordi behovet for energi som får reaksjonen i gang, blir redusert med opptil 90 prosent.
Svært mange av de biokjemiske prosessene foregår i alle levende organismer, og dermed enzymene som trengs for å gjennomføre disse reaksjonene, like hos alle organismer – både de primitive og avanserte. Det tyder på en felles evolusjonær opprinnelse.
Reaksjoner er enzymspesifikke
For hver reaksjon er det nødvendig med ett bestemt spesialisert enzym. Vi sier at alle reaksjoner er enzymspesifikke.
Primitive og avanserte organismer
Organismer som er enkelt eller komplisert bygd.
Hvordan er proteiner bygd opp?
De aller fleste enzymer er proteiner. Proteiner er bygd opp av rekker av aminosyrer som er bundet sammen med peptidbindinger. Slike rekker kaller vi polypeptider. Et protein kan bestå av ett eller flere polypeptider som er bundet til hverandre. Polypeptidene blir foldet på kryss og tvers ved hjelp av bindinger mellom atomene og får på den måten en tredimensjonal struktur. Når polypeptidene har fått denne strukturen, kaller vi dem proteiner. Den tredimensjonale strukturen er avgjørende for proteinets funksjon.
De fleste enzymer er proteiner
Enzymene er viktige proteiner i levende organismer. Noen få enzymer består bare av proteiner, men de fleste enzymene har også en tilleggsfaktor, en kofaktor, koplet til proteinet.
Kofaktorer
Noen kofaktorer er metallioner, mens andre er organiske molekyler. Disse kaller vi koenzymer. Kofaktorene deltar når polypeptidet blir foldet til et tredimensjonalt protein, og den tredimensjonale fasongen avgjør hvilket stoff som kan sitte fast på enzymet.
Substrat
Plass hvor stoffene som skal reagere er.
Biologisk katalysator
En katalysator hjelper til i en reaksjon uten selv å bli brukt opp. Et enzym virker som en biologisk katalysator. De fleste enzymer påvirker bare en spesiell reaksjon. I en menneskecelle er det tusenvis av forskjellige enzymer.
Aktiveringsenergien som trengs for å starte en reaksjon, er lavere med enzym enn uten enzym, og reaksjonen går derfor raskere når det er enzym til stede. Enzymer er nødvendige for at reaksjoner skal kunne foregå tilstrekkelig raskt ved lav temperatur. De fleste kjemiske reaksjoner i levende organismer foregår ved temperaturer i området 0-45 grader C. Kroppstemperaturen vår er ca. 37,5, og enzymene våre virker derfor best ved den temperaturen. Andre organismer har enzymer som virker best ved helt andre temperaturer. For eksempel lever flere arter av arker i de varme kilder på siland i temperaturer opp mot kokepunktet for vann, og der kan enzymene trenge en temperatur på mer enn 80 for å begynne å virke.
Hvordan et enzym fungerer
Hvordan et enzym fungerer
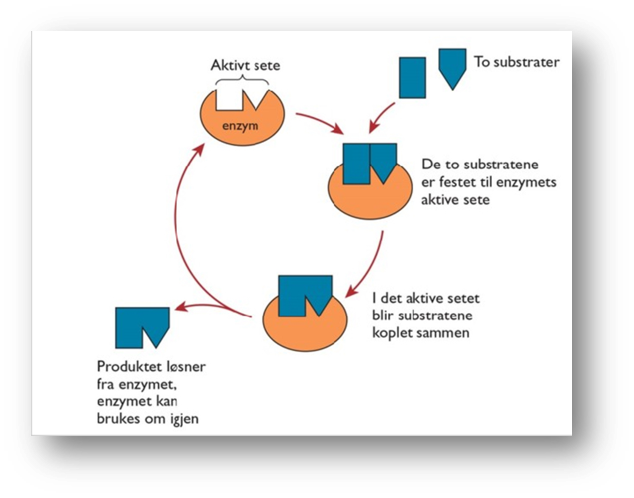
Når vi tegner en modell av et enzym, forenkler vi den kjemiske strukturformelen og tegner noe som nærmest ser ut som en klump. Klumpen består av proteindelen av enzymet og kofaktoren. Modellen av enzymet viser en svært forenklet fasong. Den må ha tomme rom – seter – for hvert av substratene som skal komme nør hverandre og reagere. Disse tomme setene symboliserer bindingsstedet der substratene kan reagere å danne produkt(er). Dette bindingsstedet kaller vi det aktive setet. Det er bare bestemte stoffer – substratene – som kan «sette seg» i det aktive setet. Det aktive setet har plass for ett eller flere substrater som inngår i en kjemisk reaksjon. Vi sier at reaksjonene er enzymspesifikke. Når vi skal forklare dette begrepet, sammenlikner vi det gjerne med en nøkkel i en lås. Akkurat som det bare er en bestemt nøkkel som kan låse opp en bestemt dør, er det bare ett enzym som er spesialisert til å delta i en spesifikk reaksjon.
Når en kjemisk reaksjon skal foregå ved hjelp av enzymer, fester substratmolekylene seg et øyeblikk i det aktive setet. Hensikten med det er at substratmolekylene skal få «sitte» ved siden av hverandre en kort stund, for da kommer de så nær hverandre at det kan skje en kjemisk reaksjon mellom dem. Plasseringen av substratene i det aktive setet gjør at aktiveringsenergien senkes og reaksjonen går raskere. Etter at substratene har reagert og dannet et produkt, løsner produktet fra det aktive sette. Det er denne mekanismen som gjør enzymet til en katalysator. Enzymet påvirker reaksjonen, men det lir ikke brukt opp. Det kan brukes om igjen og om igjen. Dermed trenger ikke cellen å bruke så mye ressurser på å lage nye enzymer.
Det aktive setet
Der substratene kan reagere å danne produkt(er). Det er bare bestemte stoffer – substratene – som kan «sette seg» i det aktive setet. Det aktive setet har plass for ett eller flere substrater som inngår i en kjemisk reaksjon. Vi sier at reaksjonene er enzymspesifikke. Når vi skal forklare dette begrepet, sammenlikner vi det gjerne med en nøkkel i en lås. Akkurat som det bare er en bestemt nøkkel som kan låse opp en bestemt dør, er det bare ett enzym som er spesialisert til å delta i en spesifikk reaksjon.
N
Karbonsyreanhydrase
Et enzym som finnes i alle dyreceller. Det er det mest effektive enzymet vi kjenner til. Enzymet er helt sentralt i reaksjonene som gjør at karbondioksidgassen som blir produsert ved den aerobe celleåndingen, fraktes ut av kroppen. Et enzymmolekyl karbonsyreanhydrase får 100 000 molekyler karbondioksid til å reagere per sekund. Reaksjonen går 10 millioner ganger raskere med enn uten enzymet. Enzymet katalyserer reaksjonen der gassen karbondioksid reagerer med vann og blir til karbonsyre og videre til hydrogenkarbonat o hydrogenioner. Denne reaksjonen skjer i blodet. Når det dannes hydrogenkarbonat, blir pH i blodet lavere, altså blir blodet for surt. I blodet er det derfor stoffer som virker som buffere og motvirker at pH blir for lav., Det er svært viktig, for de fleste reaksjonene i kroppen kan bare skje ved pH på rundt 7,4. En forandring i pH på mer enn 0,4 kan være livstruende.
Hydrogenkarbonat transporteres med blodet til gassutvekslingsorganene der den motsatte reaksjonen skjer. Også i denne omvendte reaksjonen er enzymet karbonsyreanhydrase helt avgjørende for at reaksjonen skal gå raskt og effektivt, og slik at hydrogenkarbonatet ikke hoper seg opp i kroppen. Karbondioksid går ut av blodårene og inn i gassutvekslingsorganene ved diffusjon fordi konsentrasjonen av CO2 er lavere i lungene enn i blodet. Karbondioksidgassen blir fjernet fra blodet, og vi puster ut og fjerner den fra lungene.
Buffer
En buffer er en løsning som gjør at pH verdien endrer seg svært lite selv om vi tilsetter syre eller base.
Diffusjon
Passiv transport av stoffer fra et sted med høy konsentrasjon til et sted med lav konsentrasjon.
Katalase
Enzymet katalase er viktig for å bryte ned hydrogenperoksid, H2O2. I cellene i alle levende organismer blir hydrogenperoksid dannet som et biprodukt ved metabolismen (stoffskiftet). Hydrogenperoksid er giftig og reagerer lett. Det må brytes ned umiddelbart. Derfor har alle levende organismer enzymet katalase. Det spalter hydrogenperoksid til vann og oksygengass.
Uten enzym trengs det en aktiveringsenergi på 75kJ/mol for at reaksjonen skal starte. Når katalase er til stede, er det nok med en aktiveringsenergi på bare 8kJ/mol. Katalase senker aktiveringsenergien med omtrent 90 prosent sammenlignet med aktiveringsenergien uten enzym og reaksjonen går enormt mye raskere. Når aktiveringsenergien går ned fordi enzym er til stede, øker reaksjonsfarten.
Faktorer som påvirker enzymaktiviteten
Det er mange faktorer som påvirker enzymaktiviteten. Reaksjonsfarten øker proporsjonalt med mengden av substrat og enzymer helt til alle de aktive setene i enzymmolekylene er fylt av substrat. Andre faktorer som kan gjøre at reaksjonen går raskere eller saktere er varierende temperatur eller varierende pH-verdi.
Denaturering
Proteiner forandrer struktur dersom de blir varmet opp til høye temperaturer, eller dersom de blir utsatt for svært høy eller svært lav pH-verdi. Da blir det oppkveilte proteinmolekylet først kveilet ut og deretter kveilet sammen igjen, men på en annen måte enn det først var kveilet. Dette kaller vi denaturering. Det er ikke sikkert at du har hørt selve ordet før, men alle har vært borti praktiske eksempler på hva dette er: koking eller steking av egg, fisk eller kjøtt. Også når vi legger kjøtt eller fisk i en marinade med litt eddik eller sitronsaft, blir proteinene denaturert. Vi legger kjøtt og fisk i sur marinade fordi den lave pH-verdien gjør at neon bindinger i de oppkveilte proteinmolekylene brytes. Da blir kjøttet og fisken mørnet. Enzymer eller andre proteiner lir varig ødelagt av denaturering. Denatureringen er irreversibel.
Irreversibel prosess
F.eks. denaturering. Det vil si at reaksjonen ikke kan snus o gå tilbake igjen. Et egg kan f.eks. ikke bli flytende igjen når du har stekt det.
Temperaturen påvirker reaksjonsfarten
Et enzym fungerer best ved en bestemt temperatur, den optimale temperaturen. Ved temperaturer som er lavere, går reaksjonen langsommere. Ved høy temperatur blir enzymet denaturert. Reaksjoner som foregår i menneskekroppen, har normalt en optimal temperatur på 37, mens reaksjoner som foregår i planter ofte har en optimal temperatur på 10-20.
Ph-verdien påvirker reaksjonsfarten
pH-verdien, surhetsgraden, er et mål for mengden av H3O+- ioner i løsningen. De ladde H3O+- ionene påvirker de bindingene som avgjør hvor polypeptidkjeden i enzymet er kveilet. Et enzym virker optimalt ved en spesiell pH-verdi. Enzymets aktivitet varierer med pH-verdien. Hvis pH-verdien blir for høy eller for lav, kan enzymet bli denaturert. I de fleste kroppscellene våre er pH-verdien 5,5-7 (omtrent nøytralt), og denne verdien er den optimale for enzymene inni cellene. Enzymet pepsin i magesekken virker utenfor cellene, og det har optimal virkning ved en pH-verdi på 1,7. Da spalter pepsin store proteiner til polypeptider og deretter til peptider på den mest effektive måten. Enzymet trypsin spalter så peptidene til aminosyrer i tynntarmen. Den optimale pH-verdien for trypsin er omtrent 8.
Enzymenes aktivitet blir regulert (reguleringsmekanismer)
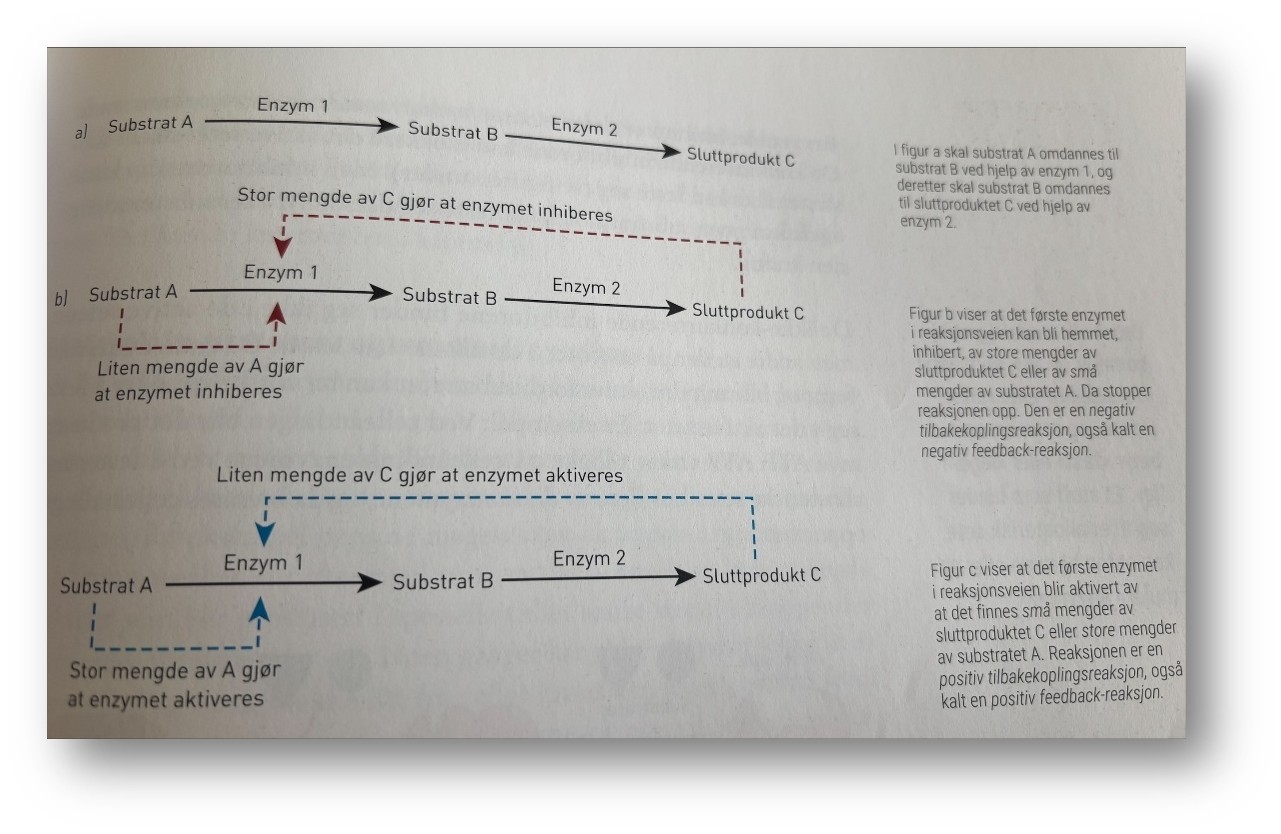
Med hundrevis av reaksjoner, substrater, enzymer og produkter i celler og vev er det lett å tenke at her kan det bli fullstendig kaos. Heldigvis blir det ikke det. Reaksjonsveien er nemlig regulert slik at sluttproduktet kan virke tilbake på det første enzymet i reaksjonsveien. Da blir det ikke laget mer av sluttproduktet enn nødvendig, eller det blir laget mer av sluttproduktet fordi det trengs. Substratet A, skal omdanne stil mellomproduktet B, og videre til sluttproduktet C ved hjelp av enzymene 1 og 2. Mengden av A, B, C reguleres ved inhibering og aktivering av enzymene 1 og 2. En stor mengde av sluttproduktet eller en liten mengde av substratet gjør at enzymet blir inhibert, mens en liten mengde av sluttproduktet eller en str mengde av substratet gjør at enzymet blir aktivert. Mengden av substrat og produkt kan altså påvirke og inhibere eller aktivere enzymet.
Mengden av sluttproduktet C på figuren over påvirker aktiviteten til det første enzymet ved en tilbakekopling. C kan virke hemmende, slik at reaksjonen stopper, som i b, eller C kan virke stimulerende, slik at reaksjonsfarten øker, som i C. Dersom C virker hemmende, kaller vi det negativ tilbakekopling eller negativ feedback. Dersom C virker stimulerende, bruker vi begreper positiv tilbakekopling eller positiv feedback.
Enzymene i reaksjonsveien kan bli aktivert eller inhibert av substratene eller sluttproduktet.
Positiv tilbakekopling
At noe virker fremmende på en prosess.
Negativ tilbakekopling
At noe virker hemmende på en prosess-
Aktivert vs. inhibert
Inhibert går på å hemme, aktivering handler om det å aktivere noe.
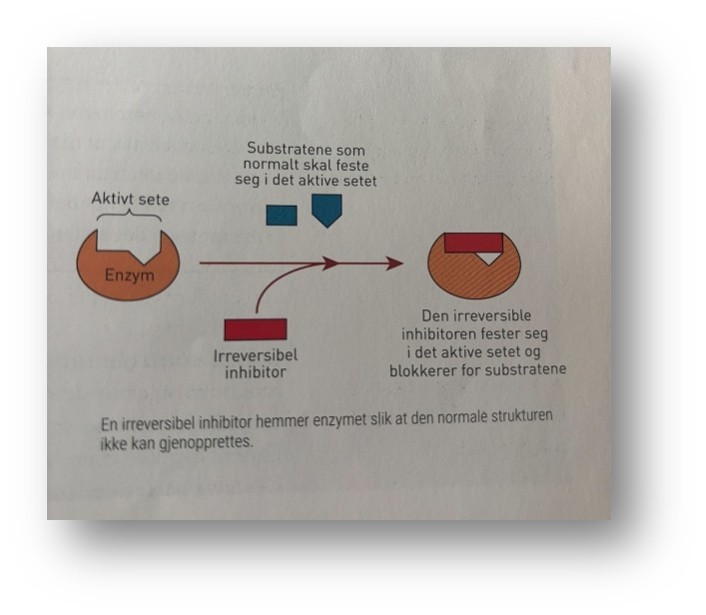
Inhibitorer påvirker enzymaktiviteten
Inhibitorer, hemmere, kan påvirke enzymer på to forskjellige måter: en irreversibel måte (kan ikke snus), og en reversibel måte (kan snus). Ved en irreversibel inhibering blir strukturen i enzymet ødelagt og kan ikke gjenopprettes. Eksempler på inhibitorer som virker irreversibelt, er plantevernmidler som sprøytes på matplanter for å drepe insekter, cyanid som gir livstruende forgiftning, og nervegasser som blir brukt ved biologisk krigføring.
Reversible inhibitorer deler vi inn i konkurrerende og ikke-konkurrerende
De konkurrerende inhibitorene kan blokkere det aktive setet slik at substratet ikke kan feste seg. Men inhibitorene kan løsne, og da kan noen substratmolekyler feste seg dersom vi øker substratmengden kraftig.
De ikke-konkureredene inhibitorene binder seg ikke i de aktive setene, men andre steder på enzymet, i de allo-steriske setene. Da endrer enzymet form og blir mindre aktivt fordi substratet kan ha vanskeligere f0rå feste seg i det aktive setet. Et eksempel: ved celleåndingen blir det produsert mye ATP. ATP virker tilbake på celleåndingsenzymene ved å feste se i allestoriske seter hos flere av disse enzymene, og da bremses celleåndingen opp.
Konkurrerende inhibitor
De konkurrerende inhibitorene kan blokkere det aktive setet slik at substratet ikke kan feste seg. Men inhibitorene kan løsne, og da kan noen substratmolekyler feste seg dersom vi øker substratmengden kraftig.
Ikke-konkurrerende inhibitor
De ikke-konkureredene inhibitorene binder seg ikke i de aktive setene, men andre steder på enzymet, i de allo-steriske setene. Da endrer enzymet form og blir mindre aktivt fordi substratet kan ha vanskeligere f0rå feste seg i det aktive setet. Et eksempel: ved celleåndingen blir det produsert mye ATP. ATP virker tilbake på celleåndingsenzymene ved å feste se i allestoriske seter hos flere av disse enzymene, og da bremses celleåndingen opp.
Allosterisk sete
Det allosteriske sete er området på et enzym som er forskjellig fra det aktive sete og som binder aktivatorer eller inhibitorer (hemmere). Enzymene kalles allosteriske fordi substratet og den allosteriske ligand (inhibitor eller aktivator) er sterisk forskjellige dvs. allosteriske.
Cellen kontrollerer enzymproduskjonen
Inni hver celle blir det produsert tusenvis av forskjellige enzymer. Noen enzymer blir produsert i alle cellene våre, mens andre bare blir produsert i noen få celle- eller vevstyper. Enzymene som deltar i kopieringen av DNA og i selve celleåndingen, må produseres i alle celler. Også reparasjonsenzymer som til enhver tid kan reparere småskader, små mutasjoner i DNA, må finnes i alle celler. Cellen kontrollerer hvilke enzymer som dannes, og mengden av hvert enzym. Cellen driver en «selvkontroll» ved kontroll av proteinproduksjonen, og dermed også enzymproduksjonen.
Metabolske prosesser og eventuell sykdom
Ut fra målinger av enzymmengden i blodplasma, urin og andre kroppsvæsker kan legen stille en diagnose. Metodene for å måle enzymer i kroppsvæsker er relativt enkle. Noen enzymer blir produsert i alle celler, for eksempel katalase. Andre enzymer blir bare produsert i enkelte vevstyper, for eksempel pepsin i magesekken og trypsin i tynntarmen.
I kroppsvæsker for eksempel urin, er det normalt å ha en liten mengde enzymer som blir produsert i spesielle celler andre steder i kroppen for å delta som katalysatorer der. Noen anger kan slike enzymer lekke ut av cellene. Særlig vil det lekke litt fra celler i muskler, lever og blod, fordi disse cellene har høy grad av metabolisme, og noe vil derfor lekke ut. Disse celletypene har mane forskjellige enzymer i aktivitet. Ved sykdom kan det lekke for mye av ett eller flere enzymer fra celler. Enzymer lekker ut blant annet i blodet og tas opp i nyrene. Derfor kan en undersøkelse av blodet eller urinen avsløre sykdom. Mengden av enzymer og hvilken tupe det er, kan si noe om hvor alvorlig sykdommen er.
Blodprøver kan også avsløre sykdom. Ved noen leversykdommer vil mengden av spesielle leverenzymer i blodet øke, og sykdommen kan da påvises med en enkel blodprøve. Det vil for eksempel lekke ut enzymer ved infeksjoner i gallegangene, leverbetennelse og begynnende skrumplever.
Ved noen hjertesykdommer som skyldes skader i muskelcellene i hjertet, øker mengden av et enzym som blir produsert i hjertemuskelcellene. Hvis dette enzymet opptrer i svært stor mengde, tyder det på hjerteinfarkt. Undersøkelser av blodets enzymer kan også si noe om personen har hatt hjerteinfarkt for en tid tilbake.
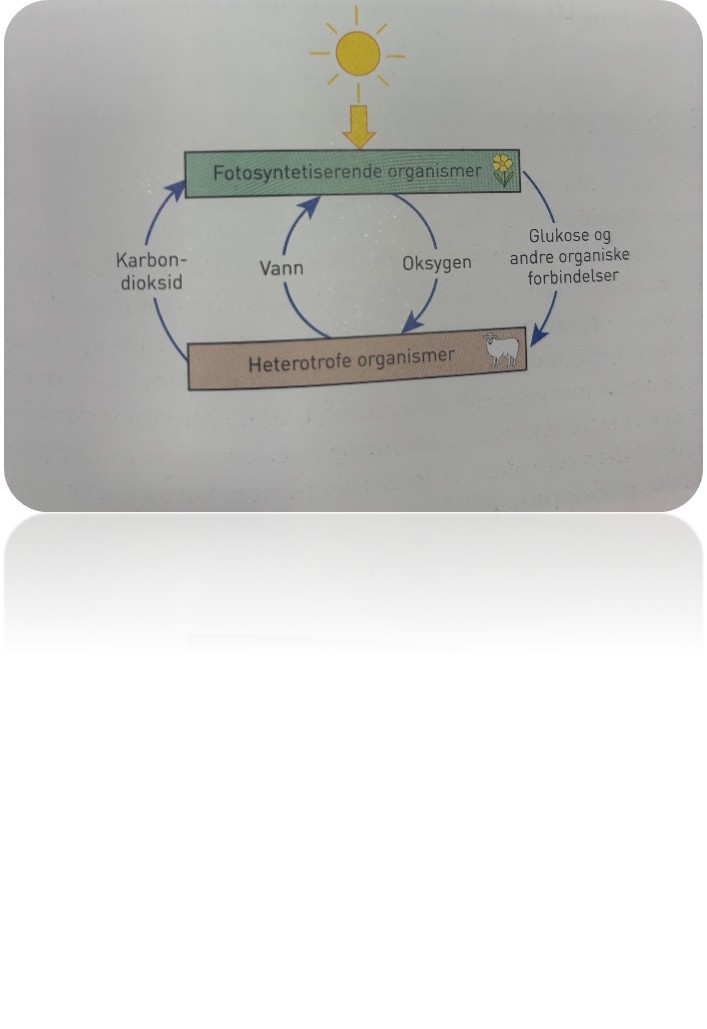
Energi blir omdannet
Sola er vår viktigste energikilde. På jorda begynner all energiomdanning med at autotrofe organismer tar opp energi og produserer organiske forbindelser. De aller fleste av de autotrofe organismene er fotoautotrofe, og de utfører fotosyntese. I fotosyntesen blir solenergi brukt til å bygge organiske forbindelser. Nesten alle planter, mange protister og noen bakterier og arker er fotoautotrofe.
Kjemoautotrofe organismer bruker ikke solenergi når de lager organiske forbindelser, men tar energi fra uorganiske forbindelser: de utfører en kjemosyntese. Eksempler på kjemoautotrofe organismer er en del bakterier og arker som lever i dyphavene, uten tilgang på lys. Både fotoautotrofe og kjemoautotrofe organismer er produsenter.
De organiske forbindelsene som dannes, blir overført fra ledd til ledd i en næringskjede – fra organisme til organisme – ved at den første organismen blir spist av en konsument, deretter blir denne konsumenten spist av en annen, og så videre til et stadig høyere trofisk nivå. Alle organismene driver med celleånding. Celleåndingen kan være uten oksygen, anaerob, eller med oksygen, aerob. Energi blir i den aerobe celleåndingen frigitt når glukose og andre organiske forbindelser reagerer med oksygen. Karbondioksid og vann dannes ved celleåndingen, forbrenningen.
De organismene som ikke produserer glukose selv, og som bare spiser andre organismer, er konsumenter (forbrukere). Vi sier at de er heterotrofe organismer. Inndelingen i autotrofe og heterotrofe organismer er gjort ut fra svarene på disse to spørsmålene: Hvem lager maten? Og hvem spiser hvem? Det er de autotrofe organismene som lager maten, og de heterotrofe organismene spiser de autotrofe organismene og andre heterotrofe organismer.
Både de autotrofe og de heterotrofe organismene har celleånding. Summen av alle disse prosessene der kjemiske reaksjoner tar opp eller frigir energi og byggesteiner, kaller vi metabolismen eller stoffskiftet.
Autotrofe organsimer
Autotrofe organismer er de som kan produsere sin egen mat fra uorganiske stoffer ved hjelp av energi fra lys eller kjemiske reaksjoner.
Heterotrofe organismer
Heterotrofe organismer er organismer som trenger organiske næringsstoffer, i motsetning til autotrofe organismer, som kan utvikle seg normalt med utelukkende uorganisk næring. Alle dyr, de fleste bakterier, alle sopp og noen høyere planter er heterotrofe.
Heterotrofe organismer er avhengige av organisk materiale for å skaffe seg energi og bygge opp cellemateriale. De kan ikke produsere sin egen mat som autotrofe organismer (som planter og alger) gjør gjennom fotosyntese. Heterotrofi er en viktig del av næringskjeden, der disse organismene fungerer som konsumenter.
- Heterotrofe organismer kan deles inn i flere kategorier basert på deres næringsstrategier:
- Herbivorer: Planteetere som lever av planter (f.eks. kuer, sauer).
- Karnivorer: Kjøttetere som lever av andre dyr (f.eks. løver, slanger).
- Omnivorer: Altetere som spiser både planter og dyr (f.eks. mennesker, bjørner).
- Saprofytter: Organismer som lever av dødt organisk materiale (f.eks. sopp, visse bakterier).
- Parasitter: Organismer som lever av levende vertskap uten å drepe dem (f.eks. bendelorm).
Fotoautotrofe
Fotoautotrofe organismer, også kjent som fototrofe organismer, er de som kan produsere sin egen mat ved hjelp av lysenergi. De omdanner karbondioksid (CO₂) og vann til organiske forbindelser, som glukose, ved hjelp av fotosyntese. Under denne prosessen frigjøres oksygen som et biprodukt.
Kjemoautotrofe
Kjemoautotrofe organismer er en type autotrofe organismer som lever av å omdanne uorganiske stoffer til organiske forbindelser ved hjelp av energi fra kjemiske reaksjoner, i stedet for lysenergi som i fotosyntese. De er i stand til å assimilere karbondioksid (CO₂) ved hjelp av energi som de får fra oksidasjon av ulike uorganiske materialer, som hydrogensulfid (H₂S), ammoniakk (NH₃) eller metan (CH₄).
Energibærere
Energi blir omdannet gjennom fotosyntesen ved at energi blir overført fra sola til kjemiske bindinger i molekyler. Videre i anabolismen blir energi bruk til å bygge andre kjemiske molekyler. Ved celleåndingen blir molekylene forbrent og energi frigitt, og energien brukt til transport, varme og arbeid i cellen. Noen av de kjemiske forbindelsene som en organisme laer, regner vi som kortvarige lagre fordi energien raskt blir overført til andre molekyler. En slik energibærer er ATP, adenintrifosfat, et kortvarig lager. I alle levende organismer finnes også mer langvarige lagre for energi, for eksempel glykogen i muskler og lever hos pattedyr, og fett hos både dyr og planter, som underhudsfett hos dyr, og for eksempel olivenolje eller rapsolje hos planter. Levende organismer har mange forskjellige energibærere, ikke bare ATP. Alle organismer har ATP.
I alle kjemiske forbindelser ligger det energi lagret i bindingene mellom atomer i form av kjemisk energi. Når det foregår kjemiske reaksjoner, kan de stoffene som reagerer, få fjernet atomer eller få tilført atomer eller funksjonelle grupper ved hjelp av en energibærer. På den måten blir de nye stoffene som bygges opp, mer energirike. Så kan de inngå i videre reaksjoner, og energien kan frigis igjen. Energibærere kan lagre denne energien, og bruke den til å drive andre reaksjoner som krever energi, blant annet til å bygge opp store forbindelser som fett og polysakkarider.
ATP
Energibærere eller energibærende molekyler inngår i reaksjoner der energi blir overført til et annet molekyl. I alle levende organismer finner vi forskjellige energibærere. Den viktigste energibæreren er ATP. Molekylet har tre fosfatgrupper. Bindingene mellom fosfatgruppene er ustabile. I energikrevende reaksjoner kan ATP overføre en fosfatgruppe til et annet molekyl, som slik får økt den kjemiske energien sin. Da kan molekylet delta i reaksjoner som eller sikke ville vært mulige.
Hos noen energibærende molekyler i levende organsimer kan evnen til å binde og overføre energi være knyttet til hydrogenatomer i form av elektroner (e-) og hydrogenioner (H+) som blir bundet til molekylet. Disse elektronene og ionenene gjør da molekylet mer energirikt.
Eksempler på andre energibærende molekyler er NADH, NADPH og FADH2. Hos disse tre molekylene er evnen til å bære og overføre energi knyttet til elektroner og hydrogenioner. ATP og NADPH er energibærere i fotosyntesen, mens ATP, NADPH og FADH2 er energibærere i celleåndingen. Både ATP, NADH, NADPH og FADH2 kaller vi korttidslagre for energi, eller hurtiglagre. De inngår hele tiden i kjemiske reaksjoner ettersom organismen trenger lett tilgjengelig energi.
ATP - bygning og virkemåte
Et ATP-molekyl er bygd opp av basen adenin, som er bundet til monosakkaridet ribose. Adenin er en base fordi den kan binde H+-ioner. Ribosen er bundet til en kjede av tre fosfatgrupper eller fosfationer. Adenin pluss ribose kaller vi adenosin og sammen med de fosfationene, får ATP navnet adenosintrifosfat. Fosfationet har kjemisk formel HPO42- men vi skriver dette ionet forkortet som bokstaven P.
ATP er et «vidundermolekyl» som alle levende organismer er avhengig av. Alle levende organismer kan lage ATP, og de bruker dette molekylet for å kunne fange opp og overføre energi. Ved hjelp av et ATP-ase-enzym kan ATP spaltes til ADP, adenosindifosfat, og en fosfatgruppe. Bindingen i ATP er ustabil og brytes derfor lett. Det er en vanlig misforståelse at energien lagres i selve bindingene mellom fosfatmolekylene i ATP. Det gjør den ikke. P løsner lett fra ATP, og resultatet blir ADP. Et fosfat blir overført til et annet molekyl, og dette molekylet får da økt kjemisk energi. Den økte energien er nødvendig for at molekylet skal kunne inngå i videre reaksjoner.
En menneskekropp på 75kg inneholder til enhver tid 40g ATP, som brytes ned til ADP og P og bygges opp igjen til ATP en kontinuerlig resirkulering der hvert ATP-molekyl blir resirkulert opptil 2500 ganger per døgn. Kroppen vår produserer og bryter ned til sammen mellom 75 og 100kg ATP per døgn.
Mengden kjemisk energi som blir overført til et annet molekyl mår en fosfatgruppe blir tilført, er ikke så stor, men siden hver celle inneholder enormt mange ATP-molekyler, lir summen av energien stor. Når en fosfatgruppe løsner fra ATP, har forbindelsen bare to fosfatgrupper igjen. Da kaller vi den ADP, der D står for di, som betyr to. Når P løsner fra ATP, blir altså fosfatgruppen overført til et annet molekyl, som dermed blir mer energirikt. Denne energien blir til transport gjennom cellemembraner, til reaksjoner der det blir dannet nye og gjerne større forbindelser, eller til bevegelse. Noe av energien fra ATP-spaltningen blir til slutt frigitt som varme.
Når P blir spaltet av fra ATP, er det nødvendig med et ATP-ase-enzym. ATP-ase virker også når reaksjonen går den andre veien, altså når ADP og P blir bygd sammen til ATP. I fordøyelsessystemet blir de store molekylene i maten som et dyr spiser, brutt ned til mindre molekyler. Dette gir byggesteiner til dyrets kropp og frigir energi som blir brukt blant annet til aktiv transport av stoffer, til varme eller til å bygge opp igjen ATP av ADP og P. Gjenbruk av ATP er helt avgjørende, for den mengden ATP som kroppen din trenger i løpet av en dag, tilsvarer kroppsvekten din.
Den andre fosfatgruppen i ADP kan også bli spaltet av, og da får vi AMP. M står for mono, som betyr en. Dette skjer sjeldnere, og det er bare en liten energimengde som blir overført når ADP spaltes til AMP.
Bruk av energien i ATP
ATP er den viktigste energibæreren i korttidslageret av energi. ATP finnes i alle levende celler hos alle organismer. ATP blir bygd opp og brutt ned kontinuerlig i metabolismen, i et høyt tempo. Energien fra dette korttidslageret blir raskt overført til andre energiformer. Den energiformen som blir overført når ATP inngår i reaksjoner, bruker organismene hovedsakelig til tre formål: mekanisk arbeid, transportarbeid, kjemisk arbeid og varme.
Også prokaryote organismer som bakterier og arker trenger enzymer til celleåndingen. Hos dem er disse enzymene festet til cellemembranen eller de flyter fritt i cytoplasmaet. Alle eukaryote organismer har organellen mitokondrier der celleånding foregår. Da blir det dannet mange ATP-molekyler i mitokondriene. ATP kan transporteres fra mitokondriene til andre organeller i cellen eller til andre celler som har behov for energi. Tabellen gir oversikt over hva energien fra ATP kan brukes til:
Energi overført til | Hva skjer? | Eksempler |
Mekanisk arbeid | - Innholdet i celler beveger seg, for eksempel blir organeller flyttet rundt - Celler beveger seg | - Kromosomer blir flyttet i forbindelse med kopiering og celledeling - Sædceller, arker, bakterier beveger seg - Proteinfibre i muskelceller trekker seg sammen |
Transportarbeid | - Aktiv transport | - Ioner, molekyler og partikler blir pumpet gjennom membraner, mot en konsentrasjonsgradient |
Kjemisk arbeid | - Noen reaksjoner trenger aktiveringsenergi før de kommer i gang og energien blir frigitt - Når små byggesteiner inngår i reaksjoner der det blir dannet større og mer sammensatte forbindelser, må det tilføres energi for at oppbygningsreaksjonene skal begynne | - All metabolisme |
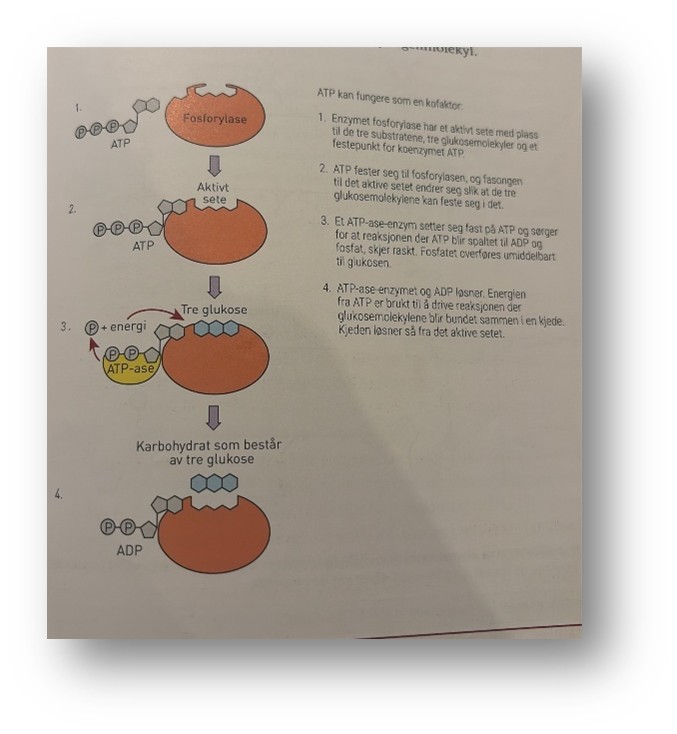
ATP som kofaktor
I tillegg til at ATP er en energibærer, er molekylet ATP også en kofaktor hos en gruppe enzymer, fosforylasene. Fosforylaser finnes i alle celler. Fosforylase har et aktivt sete der det er plass til substrater, og for at det aktive setet skal ha en fasong som passer for substratet, må fosforylasen ha koenzymet ATP festet til seg. Fosforylasene deltar når små molekyler skal bygges sammen til store molekyler, for eksempel når monosakkaridet glukose skal bygge til polysakkaridet glykogen, eller når store molekyler skal brytes ned til mindre, for eksempel glykogen til glukose.
Reaksjonen som skjer under oppbygginger av glykogen, krever energi, og denne energien blir tilført i reaksjoner der ATP inngår og blir spaltet til ADP og P. Ved nedbrytingen av glykogen til glukose skjer det omvendte – da blir det frigjort energi som blant annet kan brukes til at ADP og P igjen danner ATP. For hvert glukosemolekyl som blir festet til et annet, trengs energi fra tre ATP-er. Når glykogen blir spaltet til glukose molekyler, blir det frigitt to ATP-er for hvert glukosemolekyl som blir spaltet av fra glykogenmolekylet.
Figuren under viser hvordan glukosemolekylene blir bygd sammen. Fosforylasen har et aktivt sete med plass til tre glukosemolekyler. Ved hjelp av energi fra ATP blir det dannet bindinger mellom glukosemolekylene, slik at vi får et karbohydrat av tre glukosemolekyler som er festet ammen. Etterpå deltar to andre enzymer i to delreaksjoner som binder sammen flere av glukosekjedene til et stort glukosemolekyl
Andre energibærere: NADH, NADPH, FADH2
NADH, nikotinamid.adenin,dinukeloitid, NADPH, nikotinamid-adenin-dinukleotid-fosfat, og FADH2, flavin.adenin-dinukleotid, er tre energibærere som også spiller en viktig rolle i energiomdanningen i levende organismer samen med ATP.
Energibærere deltar i fotosyntese og celleånding
NADH OG NADPH er ganske likt bygd. Begge er bygd opp av ribose, adenin og to fosfatgrupper, det vil si ADP, og begge inneholder et molekyl B3-vitamin (niacin). Forskjellen mellom NADH og NADPH består i at NADPH har en ekstra fosfatgrupper (P) som NADH ikke har. Fosfatgruppen i NADPH er festet til ribosen.
Planter produserer selv B-vitaminer de trener til NADPH, NADH og FADH2. Dyr og mennesker må ha tilførsel av B-vitaminer gjennom maten vi spiser, men siden vi bare trenger svært få mengder, regnes B-vitaminer til mikronæringsstoffene. Av slike mikronæringsstoffer trenger vi vanligvis mellom 2 ug og 2 mg hver dag. Vi trener så små mengder fordi NADH og NADPH blir gjenbrukt.
NADH og NADPH er kofaktorer i flere enzymer som finnes i organeller inni cellene. NADH deltar i mitokondrienes celleånding og NADPH i kloroplastenes fotosyntese. Uten kofaktorene fungerer ikke disse enzymene. Hos pattedyr kan mangel på celleåndingsenzymene gi dødelige stoffskiftesykdommer.
FADH2 er også bygd opp av ribose, adenin og to fosfatgrupper, det vil si ADP. I tillegg inneholder FADH2 et molekyl B2-vitamin (riboflavin). FADH2 er en nødvendig kofaktor i neon spesielle enzymer som hovedsakelig finnes i mitokondriene. Der spiller den en avgjørende rolle i den delen av energiomsetningen som er celleåndingen.
Slik virker energibærere på celleåndingen
I celleåndingen skjer det en rekke energikrevende reaksjoner NADH dannes ved å ta opp ett H+-ion og to elektroner, og i en påfølgende reaksjon blir NADH spaltet igjen. I celleåndingen blir også FADH2 dannet gjennom reaksjoner der energi blir tilført molekylet, som tar opp to H+-ioner og to elektroner, og deretter blir FADH2 splittet igjen- Energien som blir tilført i reaksjonene som danner NADH og FADH2 kommer fra nedbrytningen av næringsstoffer, og denne energien bli brukt til å danne ATP.
Lagre av energi
ATP, NADH, NADPH og FADH2 er kortidslagre av energi. Energi blir lagret og brukt i et høyt tempo. Alle levende organismer har også langtidslagre av energi. Langtidslagrene er energireserver som vi kan bruke når kortidslagrene er tomme. Langtidslagrene er ikke så raskt tilgjengelige som kortidsslagrene. Det er nødvendig med nedbrytningsprosesser i flere trinn for å få frigitt energi fra langtidslagrene. Langtidslagrene i levende organismer består hovedsakelig av makromolekyler av karbohydrater, fett og noe protein. Planter lagrer ulike forbindelser: soya lagrer proteiner, sukkerroer lagrer karbohydrater, og i mange typer frø finnes stivelse og fett. Langtidslagrene i frø er ment for nye planter som trenger mer næring når frøene skal spire.
Hos dyr lagres langtidslagre vanligvis som karbohydratet glykogen, fett eller muskelproteiner.
Hvis du fyller en vedovn med energirike vedkubber, vil alt brenne opp, ovnen bli glovarm, og temperaturen stiger i rommet der ovnen står, Heldigvis skjer ikke dette med levende oransimer som spiser energirik mat. Kroppstemperaturen vår er regulert ved homeostasen og temperaturen holder seg nokså konstant. Dyr kan forbrenne nok til å holde seg i aktivitet, men aktiviteten varierer avhengig av om dyret er vekselvarmt eller likevarmt. Dyr har reguleringsmekanismer som sørger for at overskuddsenergien kan lagres i et langtidslaer. Dyrenes reguleringsmekanismer består i et homeostatisk samspill og et samarbeid mellom arvestoffet DNA, enzymer, hormoner og nerveceller.
Også planter har energilagre. Når vårsola sinner på snoen, vil hestehovspirene under snøen registrere lyset, og i blomstenes celler blir energien i langtidslagrene overført først til ATP, deretter dannes varme. Snøen rundt blomsterhodet smelter, og blomsten får tilgang på solas lys og varme.
Langtidslagre hos mennesker
I leverne og i musklene har vi mennesker langtidslagre av energi som består av omtrent 250 gram av karbohydratet glykogen. Når glykogenlagrene er fulle, øker ikke kroppen forbrenningen, men bruker overskuddsenergien i reaksjoner der det i spesielle celler dannes fettmolekyler eller proteinmolekyler. Hos en person som ikke trener, blir overskuddsenergien brukt til å legge opp et langtidslager av fett. Hos en person som trener en del, danner kroppen også energilagre av proteiner, men mye av energioverskuddet blir brukt i reaksjoner der det dannes proteiner, og da øker proteinmengde i musklene – og dermed også muskelmassen. Også en person som trener mye, kan øke fettmengden i kroppen hvis energiinntaket er for stort.
En person som får i seg mat med mer energi enn det kroppen trenger for å opprettholde evnen til å arbeide å holde kroppstemperaturen konstant, bygger opp et langtidslager- Da øker vekten. En person som får is seg mindre mat enn det kroppen trenger, vil først begynne å forbrenne av glykogenlageret, deretter av fettet og til slutt av proteinene, alt dette for å få nok energi ved celleåndingen. Da går de ned i vekt.
Når store organiske forbindelser som karbohydrater, fett og proteiner blir spaltet i celleåndingen, gir reaksjonen energi. Varedeklarasjonen på matvarer, angir hvor mye energi maten inneholder. Energimengden er gitt i en internasjonal standardenhet (SI-enhet) som heter joule, forkortet J. Likevel er det fremdeles mange som bruker den gamle målenheten kalori når de snakke rom energiinnholdet i mat. Energiinnholdet i mat er et mål på hvor mye energi som frigjøres når stoffene i maten inngår i kjemiske reaksjoner der store organiske forbindelser brytes ned til mindre og enklere sluttprodukter.
Fotosyntesen
Vi regner med at fotosyntesen slik den foregår i eukaryote celler, oppstod for mer enn 2 milliarder år siden, men vi vet ikke hvordan eller hvorfor dette skjedde. Vi har funnet fossiler av encellede, eukaryote fotosyntetiserende organismer som levde for 1,8 milliarder år siden. Vi antar at fotosyntesen hos eukaryote organismer oppstod ved at de tok opp encellede prokaryote blågrønnbakterier som levde videre inni de eukaryote cellene. De prokaryote cellene hadde klorofyll og de drev fotosyntese. De eukaryote cellene fikk evnen til fotosyntese etter opptak av blågrønnbakterier, og de utviklet seg videre til flercellede planter.
Før fotosyntesen oppstod, hadde ingen organsimer tilgang på oksugengass. De levde anaerobt, det vil si at de hadde anaerob celleånding uten oksygen. I fptosyntesen blir det dannet glukose, og samtidi blir oksygengass dannet som et biprodukt. En følge av fotosyntesen var at organismer utviklet den aerobe celleåndingen, altså celleånding medoksygen.
Mikroskopiske protister som planteplankton lever i avnn sammen med større protister som grønn, brun og røde alger. Disse små og store protistene er fotosyntiserende og produserer store mender oksygen og energirike organiske forbindelser. De vannlevende fotosyntetiske protistene står for mer enn halvparten av fotosyntesen på jorda, mens de landlevende plantene står for den andre halvparyen.
Nesten alle planter, mange protister og noen bakterier og arker er fotosyntetiserende organismer. Det vil si at de tar opp solenergi og bruker den i reaksjoner der det dannes glukose (druesukker) og oksygen. Fordi de lager produktene glukose og vann, karbondioksid og solenergi, kalles de produsenter. Slike organismer er autotrofe, de produsererer oganiske forbindelser og uorganiske forbinelser. Autotrofe organismer som driver fotosyntese er fotoautotrofe. Fotosyntiserende organismer har celler som inneholder fargestoffer, pigmenter. Klorofyll er ett eksmepel på et slikt fargestoff og er det viktigste pigmentet som deltar i fotosyntesen. Fargestoffene er molekyler som klarer å farge energi i sollyset og omdanne den til kjemisk energi. Hos prokaryote organismer (bakterier ig arker) er klorofyllet bundet til cellemembranen som omgir cellen. Hos eukaryote organismer med fotosyntese (planter og protister) er klorofyllet samlet i egne organeller, kloroplaster.
En enkel reakjsonslikning for fotosyntese er:
vann + karondioksid + solenergi – glukose + oksygen
Alt liv på jorda er avhengig av fotosyntesen. Fotosyntesen er den eneste prosessen som lager oksygen til atmosfæren og produserer mat til alle primærkonsumenter i alle nøringskjeder.
Solenergien omdannes
Når lys treffer et objekt, blir det eneten absorbert, reflektert eller transmitert. Absorbert lys kan varme opp objekter eller sette i gang kjemiske og biologiske prosessser. Reføektert lys sendes tilbake, altså ut fra objektet igjen, mens transmimtert lys bare passerer gjennom objkett uten at objektet blirpåvirket. Reflektert og transmietrt lys treffer øynenen våre og gir objekt fargge.
Absorbert lys
Absorbert lys kan varme opp objekter eller sette i gang kjemiske og biologiske prosessser.
Reflektert lys
Reføektert lys sendes tilbake, altså ut fra objektet igjen
Reflektert og transmietrt lys treffer øynenen våre og gir objekt fargge.
Transmittert lys
transmimtert lys bare passerer gjennom objekt uten at objektet blir påvirket. Reflektert og transmitert lys treffer øynene våre og gir objekt farge.
Pigmenter i bakterier og arker
Bakterier og arker er prokaryote, encellede organismer med få organeller. De har verken kloroplaster eller mitokondrier, men noen av dem har likevel fotosyntese. Fotosyntesen foregår ved hjelp av forskjellige pigmenter som er festet til cellemembranen og i foldede innbuktninger i cellemembranen. For eksmepel har blågrønne bakterier pigmententene røde og blå fykobiliner, gule og oransje karotenoider og flere grønne og blålige fomrer for klorofyll. Purpurbakterier har purpurfargede (lilla og fiolette) karotenioder og grønt klorodyll. Noen arker har et pigment av typen rhodopsin, som liker det rhodopsinpigmentet vi har i øyet. Alle disse pigmentene fanger solenergi.
Pigmenter i planter
Også plantene har forskjellige pigmenter som kan absorbere lys. De fleste blader er grønne fordi de inneholder de grønne pigmentet klorofyll. Klorofyll absorberer rødoransje og blålilla lys, mens grønt lys blir reflektert. Det er derfor klorofyllet ser grønt ut. I bladcellene er det også andre pigmenter som samler solenergi og overfører denne energien til klorofylletm som er det pigmentet som deltar i fotosyntesereaksjonene. Eksempler på slike pigmenter er karotenoider som karoten, som absorberer blått og grønt lys og reflekterer gult, oransje og rødt lys, og xantofyll som absorberer blågrøntlys og reflekterer brunt lys. Blålilla fykocyanin absorberer grønt og gult lys. Jo flere typer pigmenter en plantecelle har, desto flere bølgelengder av fargespekteret kan den ta opp energi fra. Energien utnytter den til fotosyntese.
Når du ser på bladene om våren og sommeren, er de fleste grønne. Om høsten blir det grønneklorofyllet omdannet til et brunt fargestogg, faeofyntin, som planten lagrer i stengel og rot. Slik sparer og resirkulerer den verdifulle byggesteiner til neste vår. Når planten trekker faeofyntin bort fra bladene, kan vi se de gule, oransje og rød4 fargestoffene – de vanlige høstfargene. Disse fargene finnes altså i blandene hele tiden, men om våren og sommeren er de dekket av grønt klorofyll, og da kan vi ikke se dem. Når klorofyllet blir omdannet og trukket bort fra bladene om høsten, blir de andre pigmentene synlige.
Bladcellene inneholder kloroplaster
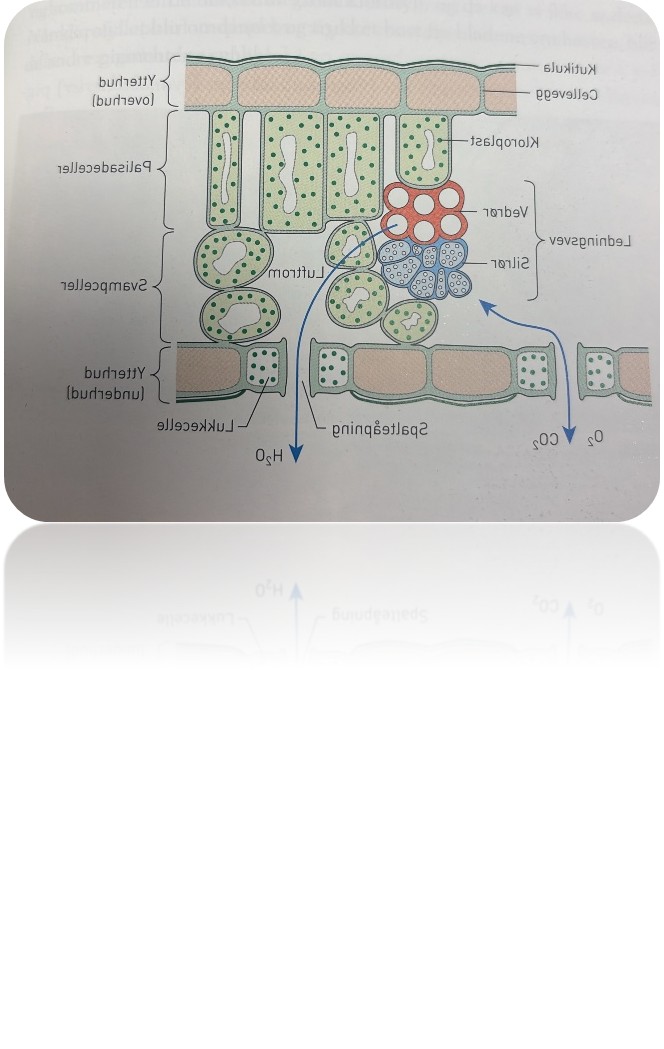
Hos de fleste planter er bladene bare noen få cellelag tykke, og sollyset slipper inn i alle cellene. Grunnvevet i bladene består som regel av to ulike typer celler: palisade celler og svampeceller.
Begge celletypene har kloroplaster, og i disse celletypene foregår det meste av fotosyntesen. Palisadevveet, som liggger like under overhuden. Det består av langstrakte, boks-fprmede celler som ligger tett i tett. Inni palisadecellene sirkulerer cytoplasmaet slik at organeller går rundt og rundt. Da kommer alle kloroplastene nær overflaten, og dermed kan de fange mest mulig av solenergien. Under palisadevevet danner svampeceller svampevevet. Her er cellene rundere og mer uregelmessige i formen, med store luftrom mellom dem med luft som inneholde roksygenass, karondioksid og vanndamp. Under svampevevet finner vi underhuden med mange spalteåpneinger som kan åpnes og lukkes ved hjelp av lukkeceller. Spalteåpningene reulere hvor mye oksygengass, karbondioskkif og vanndamp som kan slippe inn og ut av bladet.
Plantens totale bladarela er stort slik at lysabsorbsjonen kan bli stor. Noen planter kan vende bladene mot lyset etter som sola beveger seg på himmelen i løpet av dagen. Dette kaler vi heliotropisme. Andre planter vokser mot lyset. Dette kaller vi fototropisme. Dermed kan de fange større mengder av solenergien og driver mer fotosyntese. Også andre plantedeler enn bladene kan ha grlnn farge av klorofyll, men det foregår ikke noe vesentlig i for eksmepel rønne stengler sammenliknet ,med i bladnee. Kaktus har en pesiell bygningg og er et unntak. Kaktuskroppen er stengelen, og de rksjer fotosyntesen. Tornene er bladene, og i dem skjer det omtrent ingen fotosyntese.
Heliotropisme
Noen planter kan vende bladene mot lyset etter som sola beveger seg på himmelen i løpet av dagen.
Fototropisme
Planter vokser mot lyset.
Kloroplastenes oppbygning
I grønne planteceller kan vi finne opptil flere hundre kloroplaster per celle. Kloroplastene er stedet der selve fotosyntesen foregår. Der blir energien i sollys pluss vann og karbondioksid omdannet til glukose og oksygengass. Kloroplastene virker en viss tid før de går til grunne. Planteceller produserer nye kloroplaster hele tiden. Kloroplastene inneholder sitt eget arvestoff og kan til en viss grad styre kopieringen av seg selv. Samtidig blir prosessen overordnet kontrollert av arvestoffet i cellekjernen, slik at cellen ikke får dor få eller for mange kloroplaster.
Kloroplastene har doble ytre membraner rundt det væskefylte rommet, stroma. I stroma ligger stabler av blæreformede sekker. En slik stabel kalles et granum, flere stabler kalles for grana. En blæreformet sekk, en tylakoid, er en membrandekket sekk med væske. Membranen rundt en tylakoid er tylakoidmembran, og delreaksjonene i fotosyntesen foregår både i tylakoidmembranen og i stroma. I tylakoidmembranene ligger alle pigmentene som deltar i fotosyntesen, både klorofyll og de andre pigmentene. Væsken både i granum og stroma består blant annet av en mengde enzymer som katalyserer reaksjonene i fotosyntesen. Da kan reaksjonene gå tilstrekkelig raskt slev ved lav temperatur.
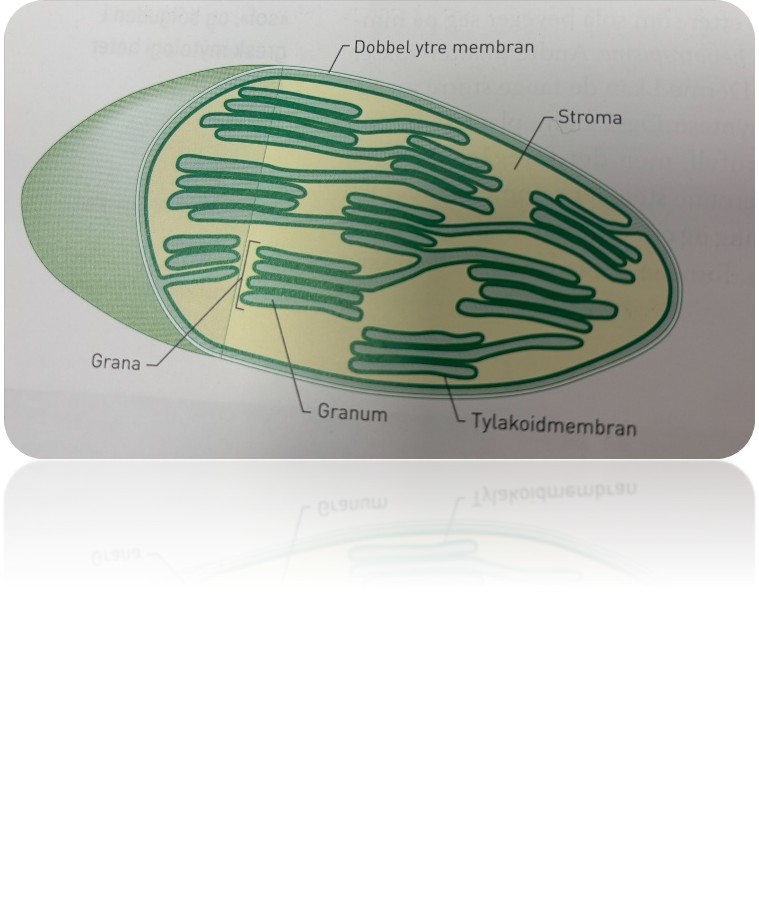
Tylakoider
En blæreformet sekk, en tylakoid, er en menmbrandekket sekk med vøske. Membranen rundt en tylakoid er tylakoidmembran, og delreaksjonene i fotosyntesen foregår både i tylakoidmembranne og i stroma. I tylakoidmembranene ligger alle pigmentene som deltar i fotosyntesen, både klorofyll og de andre pigmentene
Stroma
Kloroplastrene har doble ytre embraner rundt det vøskefylyte rommet, stroma. I stroma ligger stabler av blærefomrede sekker. En slik stabel kalles et granum, flere stabler kalles for grana. . Væsken både i granum og stroma består blant annet av en mengde enzymer som katalyserer reakjsonene i fotsyntesen. Da kan reaksjonene gå tilstrekkelig easkt slev ved lav temperatur.
Fotosyntesens lysavhengige reaksjon
Fotosyntesne består av to deler: den lysavhengige reaksjonen og den lysuavhengige reaksonen.
Den lysavhengige reaksjonen omdanner lysenergi til kjemisk bundet energi. Reaksjonen er lusavhengig og foregår ikke i mørket. Den kjemiske energien blir lagret i to eneribærende forbindelser. ATP og NADPH, og det blir også dannet o2
Den lysuavhengige reaksonen kan foregå båd ei lys og mørke. Reaksjonen er lysuavhengig. Den kjemiske enerien fra ATP og NADPH blir poverført i reaksoner med vann og karbondioksif, og vi får glukose.
Antennekompleks
Evnen til å fange energien i sollyset er et resultat av et samspill mellom flere molekyler i tylakoidmembranene: pigmenter, proteiner og enzymer. En slik gruppe av samarbeidende molekyler kaller vi et antennekompleks. I et antennekompleks samarbeider pigmentene med proteiner og enzymer for å kunne ta opp mest mulig solenerg, det vil si for å kunne fange opp solenergi fra forskjellige bølgelengder av lyset. Antennekompleksene gjør at mer energi i sollyset blir tatt opp enn om bare klorofyll hadde tatt opp sollys. Jo flere forskjellige pigmenter kloroplastene inneholder, desto mer av energien i sollyset kan de ta opp, og desto mer effektiv blir fotosyntesen.
De forskjellige pigmentene som tar opp energien fra sola, sender energien videre til klorofyllet. Figuren nedenfor viser hvordan solenergien blir tatt opp av gule, røde og oransje pigmenter før den sendes videre til klorofyllet.
Når sollys treffer elektroner i atomene i pigmenter, kan energien gjøre at noen av elektronene hopper fra sin naturlige bane til en høyere bane som ligger lenger ut fra kjernen. Vi sier både at atomet er eksitert, og at det har fått eksiterte elektroner. Eksiterte atomer er brakt til en høyere energitilstand; de er altså brakt opp i energinivå enn ikke-eksiterte atomer. De eksiterte elektronene er ustabile og kan ikke være i denne posisjonen særlig lenge. Da har de to muligheter: De kan enten falle tilbake til sin opprinnelige bane og samtidig frigjøre energi, eller de kan overføre energien til et annet atom, primært elektronakseptor, som trekker til seg et til seg elektronet.
De eksiterte elektronene i antennekompleksene kan falle tilbake til sin bane og frigjøre lys, eller energien kan overføres til et annet atom i antennekomplekset. Når energien overføres til et annet atom, vil elektronakseptoren overføre sitt elektron videre, og går da fra å være elektronakseptor til å bli elektron donor. De eksiterte elektronene blir overført til atomer i klorofyllet, som er neste elektronakseptor.
Når sollys treffer klorofyllmolekylet, eller når klorofyllmolekylet får tilført energi fra andre pigmenter i antennekompleksene, blir klorofyllet atomet eksitert. De eksiterte elektronene blir tatt fra klorofyllet — som da er en elektrondonor — og fanget av et protein som trekker elektronene til seg og fungerer som en elektronakseptor. I neste omgang vil et annet protein ta imot elektroner fra det første proteinet, som dermed går fra å være en elektronakseptor til å bli en elektrondonor. Slik skifter proteiner mellom å akseptere (ta imot) elektroner og donere (gi bort) elektroner i en elektrontransportkjede.
Fotosystemene
Et fotosystem består av et antennekompleks med pigmenter, proteiner og enzymer som danner en elektrontransportkjede. Et fotosystem er altså et område i tylakoidmembranene der solenergien blir fanget opp og der elektroner i pigmenter først blir eksitert og deretter sendt videre gjennom ei elektrontransportkjede bestående av proteiner. Hver kloroplast har hundrevis av slike områder i hvert grannum.
I tylakoidmembranene finner vi to ulike fotosystemer: fotosystem II og fotosystem I. Fotosystem II kommer før fotosystem I. Årsaken til denne omvendte nummereringen er historisk: Fotosystem I ble nemlig oppdaget før fotosystem II. De to fotosystemene har en serie av delreaksjoner, og de samarbeider om å omdanne solenergi til kjemisk energi.
Når vi nå tar utgangspunkt i pigmentet klorofyll videre, er det fordi klorofyll er det pigmentet som direkte deltar i fotosyntesen. Husk at også andre pigmenter i antennekompleksene fanger solenergi.
Fotosystem II
Fotosystem II består av klorofyll og en elektrontransportkjede i tylakoidmembranene. I fotosystem II treffer sollyset et klorofyllmolekyl (K) og gjør at to elektroner i et av atomene i klorofyll hopper ut i en bane som ligger lenger ut fra kjernen. Vi skriver K*. Stjernen står her for eksitert klorofyll. De to eksiterte og energirike elektronene er ustabile og blir raskt fanget av en elektronakseptor og ført videre i en elektrontransportkjede, slik figuren ved siden av viser. Klorofyllet, som har mistet to elektroner, blir positivt ladd, og vi skriver derfor K²⁺ for å angi den positive ladingen.
For at K²⁺ igjen skal kunne ta opp solenergi, må det ta opp elektroner og gjendannes til et nøytralt klorofyllmolekyl, K. Det skjer ved at molekylet tar to elektroner fra et vannmolekyl, som da blir spaltet til oksygengass og H⁺-ioner. Det er i denne prosessen oksygengass blir dannet. En fotosyntetiserende organisme har ikke bruk for oksygengassen til fotosyntese.
I denne sammenhengen er oksygengass derfor et avfallsprodukt. Oksygenet som blir produsert i fotosystem II, slippes ut til omgivelsene og blir dermed tilgjengelig for alle organismer som har celleånding, inkludert de fotosyntetiserende organismene.
H⁺-ionene (protoner) som blir dannet ved spaltingen av vann, gjør at innsiden av tylakoidmembranen blir positiv i forhold til utsiden. Ladningsforskjellen mellom innsiden og utsiden gir en spenning. Når elektronene blir overført fra protein til protein i elektrontransportkjeden, bruker proteinene samtidig energien fra de eksiterte elektronene til å pumpe flere H⁺-ioner fra stroma, altså utsiden av tylakoidmembranen, til innsiden og det indre av tylakoiden. Store mengder H⁺-ioner hoper seg opp i tylakoiden, og slik øker spenningen ytterligere på grunn av ladningsforskjellen mellom innsiden og utsiden av tylakoidmembranen.
Ladningsforskjellen kaller vi en protongradient. Den kan brukes til å danne ATP. Spesielle porter i tylakoidmembranene består av et ATP-ase-enzym. Bare gjennom disse portene kan H⁺-ionene passere for å komme ut i stroma og utjevne ladningsforskjellen. ATP-ase-enzymet katalyserer reaksjonen der det blir dannet ATP. Når H⁺-ionene passerer portene, frigjøres det energi som blir brukt i reaksjonen der det dannes ATP av ADP og fosfat, P:
ADP + P + energi → ATP
Den opprinnelige solenergien som eksiterte elektroner i klorofyll, blir fraktet via elektrontransportkjeden i fotosystem II. Den blir først brukt til å pumpe H⁺-ioner inn i tylakoiden, deretter til å slippe H⁺-ioner ut gjennom ATP-ase-porter i tylakoidmembranen, og til sist blir den omdannet til kjemisk energi i det energibærende molekylet ATP. Energi fra ATP blir overført senere i reaksjonene der det dannes glukose i den lysuavhengige reaksjonen.
Fotosystem I
Fotosystem I består også av klorofyll og en elektrontransportkjede i tylakoidmembranene. Det begynner med at et atom i klorofyllmolekylet blir eksitert av solenergi. Da gir klorofyllet fra seg to energirike elektroner til en elektronakseptor, slik som i fotosystem II. Elektronene som klorofyllet ga fra seg, blir ikke erstattet av elektroner som kommer fra spalting av vann – i motsetning til i fotosystem III. Klorofyllet i fotosystem I får derimot donert to elektroner fra det siste proteinet i elektrontransportkjedne i fotsystem II.
I fotosystem I blir elektronene overført fra protein til protein i elektrontransportkjeden og ender i stroma. Der blir de to elektronene bundet sammen med NADP+ og et H+-ion til et molekyl NADPH:
NADPH+ + 2E- + H+ - NADPH
H+- ionene er de som opprinnelig stammer fra spaltningen av vann, og som i fotosystem II passerte gjennom membranen i ATÅ-ase-portene. I fotosystem I blir altså solenergi overført til kjemisk energi i det energibærende molekylet NADPH.
Vi antar at utviklingen av de to fotosystemene begynte hos en enkelt organisme for om lag 3 milliarder år siden. Grønne svovelbakterier er en gruppe bakterier som bar har fotosyste I med hydrogensulfid som elektrondonor til klorofyllet, etter eksitasjonen, ikke vann. Fotosyntiserende blågrønnebakterier, alger o planter har både fotosystem II og I. Blågrønnebakyerierne var kanskje forløpere for de eukaryote cellene i planter, for de har indre membranliknende strukturer som likner tylakoidmembraner.
Alle fotosyntiserende organismer får det samme resultatet av den lysavhengige reaksjonen: to typer energirike molekyler, nemlig ATP og NADPH.
Fotosyntesens lysuavhengige reaksjon
I den lysuavhengige reaksjonen blir den kjemiske energien fra energibærerne ATP og NADPH₂, som blir dannet i lysreaksjonen, brukt i det begrensende aktivitetene der karbondioksid blir til glukose. Produksjonen av glukose skjer så lenge det finnes CO₂, ATP og NADPH₂, og skjer altså både i lys og i mørke inntil den kjemiske energien i ATP og NADPH₂ er brukt opp. Det er egentlig galt å si lysuavhengige reaksjon eller mørkereaksjonen, men det er et misvisende navn, siden den foregår både i lys og også en stund i mørke.
Den lysuavhengige reaksjonen skjer i stroma, og delreaksjonene blir katalysert av mange enzymer som flyter fritt der. I den første delreaksjonen blir karbondioksid bundet. Binding av karbondioksid kaller vi karbonfiksering.
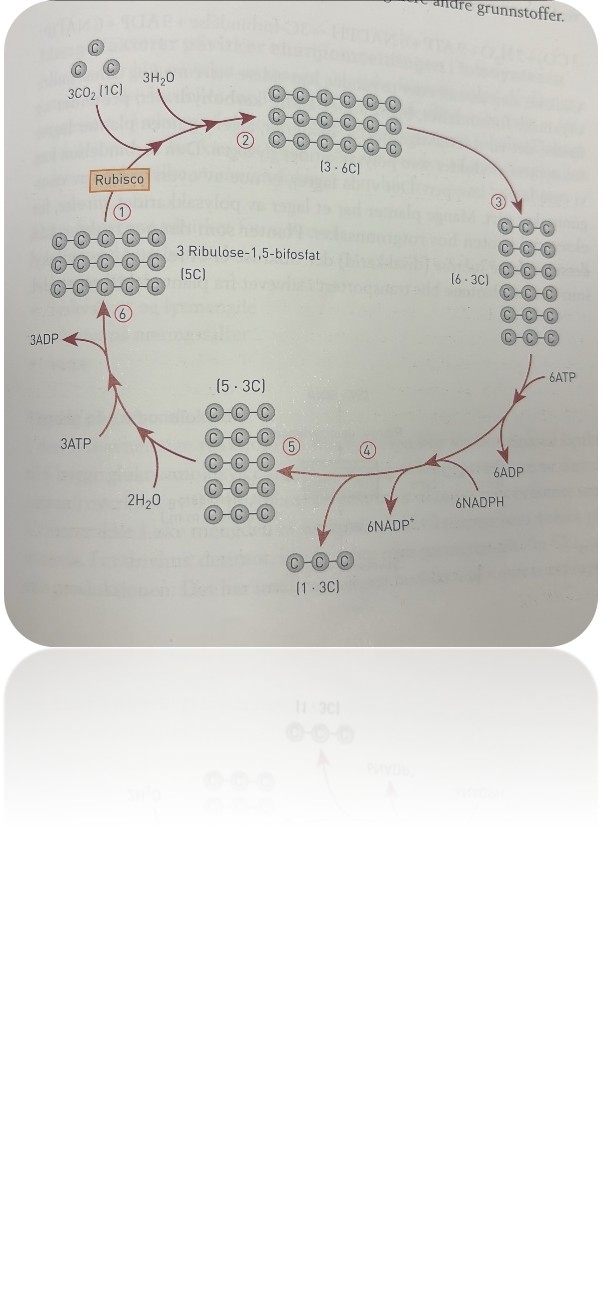
Calvin-syklusen
Spalteåpninger i underhuden i bladene kan åpnes og slippe inn karbondioksid. Se figuren på side 170 som viser et snitt av et grønt blad. Dette karbondioksidet går inn i en lysuavhengig syklisk reaksjon der karbondioksid og vann blir tatt opp gjennom flere runder slik at sluttproduktet blir glukose. Syklusen har navn etter én eller to av dem som i 1950-årene beskrev detaljene i det som skjedde: Calvin-syklusen eller Calvin-Benson-syklusen. Den fungerer som en «sukkerfabrikk».
Det første enzymet som deltar, er rubisco, det vil si ribulose-bifosfat-karboksylase. Rubisco består av 16 polypeptidkjeder som er kveilet sammen til et protein. De har metallionet magnesium som kofaktor. Rubisco fikserer tre molekyler karbondioksid per sekund. Det er ekstremt lavt sammenliknet med reaksjonsfarten i de fleste andre enzymkatalyserte reaksjoner. Plantene trenger store mengder av dette enzymet for å sikre tilstrekkelig med karbon. Bladene inneholder derfor store mengder rubisco: 50 prosent av proteininnholdet er rubisco. Rubisco er antakelig det enzymet som samlet sett finnes i størst mengde i levende celler. Noen forskere kaller rubisco for «verdens viktigste enzym».
Figuren nedenfor viser den lysuavhengige Calvin-syklusen. Legg merke til at vi på figuren bare tegner karbonatomene (grå kuler), selv om molekylene også inneholder hydrogen, oksygen og flere andre grunnstoffer.
De ulike stegene:
1. Tre molekyler ribulose 1,5 bifosfat binder seg til hvert karbondioskidmolekyl og vann. Rubisco er katalysator.
2. Det blir dannet tre molekyler av en ustabil sekskarbonforbindelse
3. sekskarbonforbindelsen spaltet i seks molekyler med tre karboner i hver
4. Trelarbonforbindelseme blir bygd om til andre trekarbonforbindelser, og energien fra ATP og NADPH blir brukt til å flytte på atomer. En av de skes trekarbonforbindelsene går ut av syklusen, og de fem andre fortsetter
5. De fem trekjarbonforbidnelsene blir bygd gjennom flere delreaksjoner der vann og ATP blir brukt.
6. De fem trekarbonforbindelsene blir omdannet til tre molekyler med rbulose 1,5 bifosfat.
Starten på Calvin-syklusen er at tre molekyler ribulose-1,5-bifosfat (femkarbon, 5C) binder seg til hvert sitt karbondioksidmolekyl (1C) og vannmolekyl, med enzymet rubisco som katalysator. Hvert av de tre molekylene ribulose-1,5-bifosfat har fem karbonatomer, og hvert av molekylene kopler seg sammen med et karbonatom som kommer fra karbondioksid. Da blir det dannet tre molekyler av en sekskarbonforbindelse (6C). Sekskarbonforbindelsen er ustabil, og de tre molekylene blir raskt spaltet i seks molekyler med tre karbon (3C) i hver. Disse første reaksjonene blir raskt fulgt av flere der trekarbonforbindelsene blir bygd om til andre trekarbonforbindelser, og energien fra 6 ATP og 6 NADPH blir brukt i disse reaksjonene. Da blir ATP spaltet til ADP og P, mens hvert NADPH blir spaltet til blant annet NADP⁺. Én av de seks trekarbonforbindelsene går ut av syklusen. Resten – fem trekarbonforbindelser – fortsetter i syklusen og blir omdannet gjennom flere delreaksjoner der to molekyler vann og tre molekyler ATP blir brukt. Til slutt blir de fem trekarbonforbindelsene omdannet til tre molekyler ribulose-1,5-bifosfat (5C), og syklusen gjentar seg. Den ene trekarbonforbindelsen som forlot syklusen, kan binde seg til enda en trekarbonforbindelse når syklusen har gått en runde til, og da får vi dannet glukose, som er en sekskarbonforbindelse, C₆H₁₂O₆.
Forenklet kan vi si at resultatet av en runde av den lysuavhengige reaksjonen som vi kaller Calvin-syklusen, er
3 CO₂ + 5 H₂O + 9 ATP + 6 NADPH → 3C-forbindelse + 9 ADP + 6 NADP⁺
Glukose, som dannes etter to runder, kan videre være med og danne andre organiske forbindelser, blant annet andre karbohydrater, proteiner og lipider, det vil si fettaktige stoffer. Bakterier, alger og noen planter lagrer fotosynteseprodukter som polysakkaridet glykogen. Den forbindelsen kan vi også lagre i kroppen. Det vil da lagres, når det er mye av overskuddet av energiinntaket vårt. Mange planter har et lager av polysakkaridet stivelse, for eksempel i roten hos rotgrønnsaker. Planter som danner frukter, kan dessuten lagre sukrose (disakkarid) der. Sukrose er et relativt lite molekyl som hos karplantene blir transportert i silvevet fra plantedel til plantedel, og som også kan brukes av plantecellene i celleåndingen når de trenger energi. Sukrose fungerer som en energikilde som lett kan flytte sfra ett stedtil et annet. Hos mennesker fungerer glukose på samme måte. Den blir transportert viua blodårene til kropsceller som trenger energi.
Faktorer som påvirker energiomsetningen i fotosyntesen
Alle planter blir påvirket av mange abiotiske faktorer enten de står inne i stua, i et drivhus eller ute i naturen. En bonde eller en gartner er ofte interessert i å øke produksjonen, altså at fotosyntesen skal øke. Dersom produksjonen er lav, kan det være viktig for bonden å forstå hvorfor den er lav – altså hvilke minimumsfaktorer som hindrer større fotosyntese.
Disse faktorene påvirker fotosyntesen:
· tilgang på karbondioksid
· temperatur
· lyskvalitet og lysmengde
· tilgang på næringssalter
· vann
Tilgang på karbondioksid
I den lysuavhengige reaksjonen i fotosyntesen blir karbondioksid brukt til å bygge glukosemolekyler. I lufta i naturen er konsentrasjonen av denne gassen i overkant av 0,04 prosent, uansett hvor økosystemet befinner seg. Vi klarer ikke å øke mengden av karbondioksid til planter som vokser på et jorde. I et drivhus, derimot, kan bonden eller gartneren tilsette CO₂ og øke produksjonen. Det har imidlertid ingen hensikt å øke den svært mye, for når alle kloroplastene arbeider maksimalt, kan ikke mer karbondioksid få aktiviteten til å øke, og ved en konsentrasjon på 0,5 prosent virker gassen som en gift. Da kan plantene dø.
I dag er det ingen tvil om at menneskeskapte utslipp av karbondioksid til atmosfæren gir en økning i den globale oppvarmingen, det vil si økt drivhuseffekt. En del forskere mener at det kan være en løsning å sette i gang skogplantingsprosjekter, fordi flere trær på jorda vil gjøre at det blir tatt opp mer CO₂. Men det forutsetter at trærne får stå, at de verken blir brent eller får råtne opp, for i så fall blir CO₂ frigjort igjen. Også å dyrke trær som skal brukes til byggematerialer, innebærer lagring av CO₂.
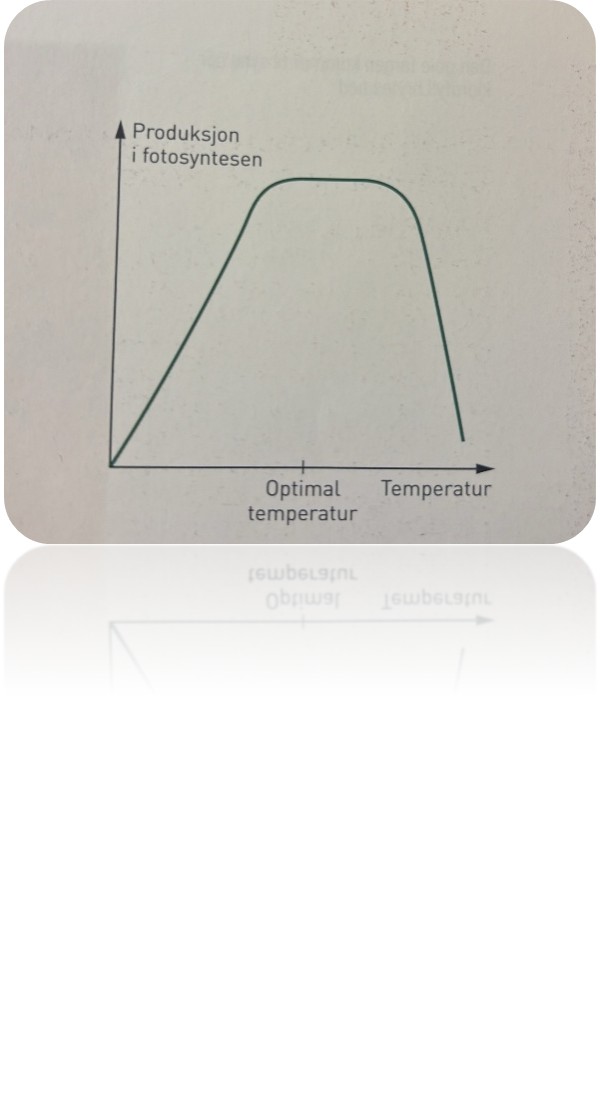
Temperatur
Alle enzymer virker best ved en bestemt temperatur, den optimale temperaturen for reaksjonen. Den optimale temperaturen varierer fra art til art. Hos noen av høyfjellsplantene i Norge kan den ligge på rundt 15 °C, mens den hos regnskogsplanter i Sør-Amerika kanskje er 35 °C. Planters enzymer er tilpasset sin optimale temperatur, og dersom temperaturen øker mye, kan enzymene bli denaturert.
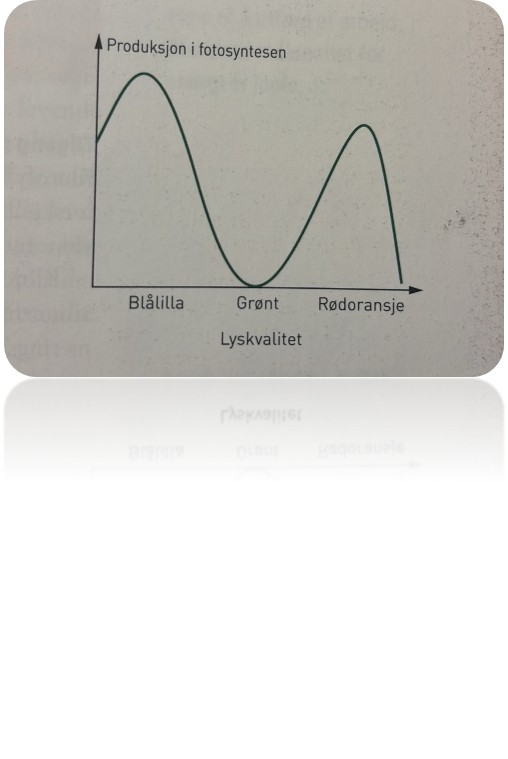
Lyskvalitet og lyskvantitet
I avsnittet om pigmentene som deltar i den lysavhengige delen av fotosyntesen, viste figuren på side 169 fargene i lysspekteret, og hvordan forskjellige pigmenter absorberer ulike bølgelengder av lys. Det er opplagt at dersom en plante er plassert i ren, grønt lys, vil klorofyll og flere an
dre pigmenter i rent, grønt lys, reflektere det grønne lyset og absorbere svært lite. Det vil komme lite fra klorofyllmolekylene til å skaffe energi, og fotosystem II kommer ikke i gang. Derfor er lyskvaliteten viktig for den lysavhengige reaksjonen i fotosyntesen. I noen gartnerier kan du se at gartneren bruker lysrør som bare gir rødfarget lys, fordi klorofyll utnytter dette lyset så godt.
Det finnes en grense for hvor lite lys planter kan leve i; det vil si hvor svakt lyset kan være hvis planter skal kunne overleve og vokse. Årsaken er den samme som vi nevnte ovenfor: Lite lys gir få eksiterte klorofyllmolekyler, det blir lite fotosyntese og derfor liten produksjon. Omvendt kunne vi tenke oss at en gartner satte på et svært sterkt lys i drivhuset for å øke produksjonen maksimalt. Når alle kloroplastene og klorofyllet i dem er «i arbeid», har planten en maksimal produksjon. Da sier vi at produksjonen er lysmettet. For sterkt lys kan faktisk svi pigmentene, med den følge at kloroplastene blir ødelagt og går til grunne.
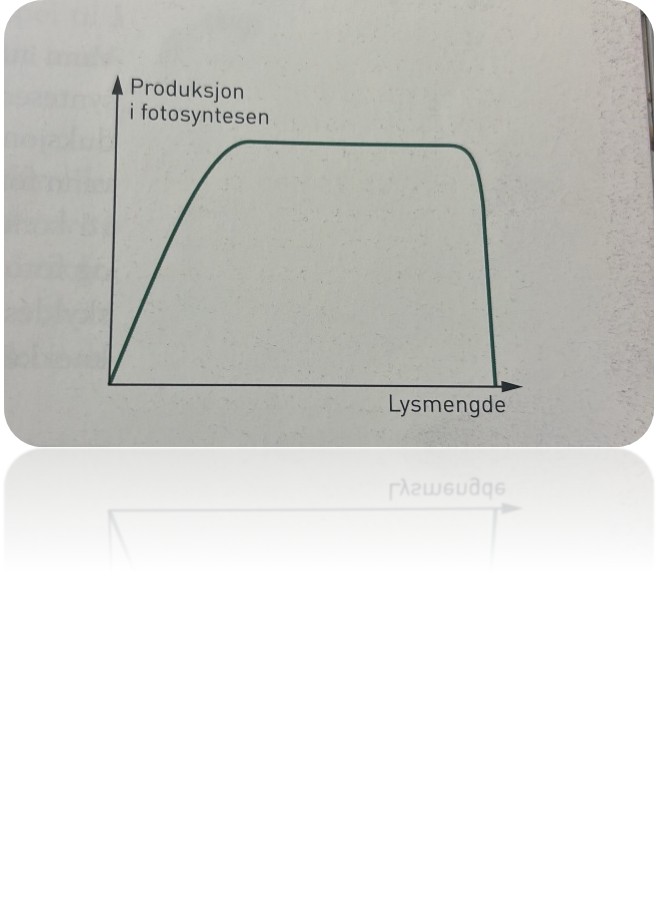
Tilgang på næringssalter
Klorofyll, andre pigmenter, enzymer o andre proteiner inneholder mange forskjellige grunnstoffer. De får i seg disse stoffene når planten suger opp vann med oppløste næringssalter fra jorda.
Klorofyllmolekylet inneholder magnesium, mens det i proteinene blant annet finnes nitrogen, fosfor og svovel. Planter som får for lite av disse nøringssaltene, lager ikke nok klorofyll, og de lager ikke nok proteiner som kan delta i elektrontransportkjeden. Da kan de få klorose, som er gulfarging av bladene på grunn av klorofyllmangel, eller nekrose, som er brune og døde områder på bladene fordi det ikke foregår noen fotosyntese der. Slike planter har svært liten fotosyntese.
Vann inngår både i den lysavhengie og den lysuavhengige delen av fotosyntesen, men vannmangel alene er sjelden en minimumsfaktor for produksjon. Planter på tørre steder lukker spalteåpningene for å unngå at vann fordamper fra bladene, men da blir samtidig karbondioksd hindret i å komme inn gjennom spalteåpmingene. CO2 er minimumsfaktoren, og fotosyntesen stopper opp. Planten kan dø av vannmangel, men det skyldes at de på grunn av uttørkede celler faller sammen slik at stilken knekker og vevet går i stykker.
Celleåndingen (metabolismen =stoffskiftet)
I en bilmotor blir bensin og oksygen omdannet til karbondioksid og vann gjennom en rekke kjemiske reaksjoner. Samtidig blir det frigitt energi. Noe av energien frigjøres som varme, resten bidrar til at bilen beveger seg. I alle levende organismer skjer det samme som i bilmotoren: organiske forbindelser blir brutt ned ved katabolisme, men ikke alle levende organismer bruker oksygen til nedbrytningen. Noen organismer bryter ned ved anaerob celleånding. Hos organismer med aerobe celleånding, som hos alle dyr og planter, blir mat og oksygen omdannet til karbondioksid, vann og energi. Enzymer sørger for at reaksjoner der store molekyler blir brutt ned til små molekyler, går raskt. I nedbrytningsprosesser blir det frigitt energi når molekylene reagerer og danner stadig nye og mindre forbindelser.
Mange dyr har sirkulasjonssystem som hjelper til med transport av ulike stoffer. Små molekyler kan, i motsetning til større molekyler, transporteres med blodet rundt i kroppen. I cellene i kroppen kan de små molekylene settes sammen til store molekyler i anabolismen, eller de kan brytes videre ned og frigjøre energi som kan utnyttes for eksempel til å transportere molekyler og ioner gjennom membraner (såkalt aktiv transport) eller til å gi organismen kroppsvarme.
Summen av alle prosessene der energi og byggesteiner blir bundet eller frigjort, kaller vi metabolismen, stoffskiftet. Metabolismen består av to hoveddeler: anabolismen og katabolismen. Oppbyggingen av små molekyler til store molekyler kalles anabolisme, mens nedbrytningen av store molekyler til små molekyler kalles katabolisme. Hver eneste av delreaksjonene i metabolismen blir katalysert av ett spesifikt enzym.
I katabolismen kan store organiske molekyler bli brutt delvis ned og brukes til byggesteiner i nye forbindelser, eller de kan bli brutt videre ned i celleåndingen. Celleåndingen kan foregå aerobt, altså med oksygen, eller anaerobt, uten oksygen. Ordet forbrenning blir ofte brukt om både aerob og anaerob celleånding. Noen organismer har bare aerob celleånding, mens andre ikke trenger oksygen og har altså anaerob celleånding, mens andre kan veksle mellom aerob og anaerob celleånding, avhengig av oksygeninnholdet i miljøet.
En enkel reaksjonslikning for den aerobe celleåndingen er:
glukose + oksygen → vann + karbondioksid + energi
C₆H₁₂O₆ + 6 O₂ → 6 H₂O + 6 CO₂ + energi
I likhet med fotosyntesereaksjonen, består også reaksjonslikningen over av mange delreaksjoner. En sammenlikning av fotosyntese og aerob celleånding viser at den totale reaksjonslikningen likner en «omvendt» fotosyntese, men reaksjonen er ikke det. Fotosyntesen og aerob celleånding foregår i helt ulike organeller (hos eukaryote) og med helt forskjellige enzymer.
Prokaryote organismer har enzymer for celleåndingen festet til den indre delen av cellemembranen som omgir cellen, eller enzymene flyter fritt i cytosol. Hos eukaryote organismer foregår celleåndingen i mitokondriene. Mitokondriene er cellenes «kraftverk». Der blir organiske molekyler spaltet og energi frigitt. Jo høyere energibehov en celle har, desto flere mitokondrier har den. En muskelcelle kan ha opptil 10 000 mitokondrier, mens en levercelle kan ha 2500. Mitokondrier inneholder eget DNA, og vi antar at eukaryote celler med mitokondrier har oppstått ved endosymbiose.
Mat gir byggesteiner og energi
Det finnes byggesteiner i all mat. Maten vår kommer både fra autotrofe og heterotrofe organismer, og i et balansert kosthold finner vi vanligvis så vel planter som dyr. Både i grønnsaker, frukt og kjøtt er det proteiner, fett og karbohydrater.
Hvis kostholdet vårt skal bestå av bare planter, er det svært viktig at vi har nok kunnskap til å sette sammen en variert og riktig mengde av forskjellige planter.
Mat gir også energi. Når de store organiske molekylene i maten brytes ned, blir det frigjort store mengder energi. I celleåndingen blir energien enten frigjort som varme, overført til andre molekyler eller brukt til å utføre arbeid. I alle organismer kan proteiner brytes ned til aminosyrer, fett kan brytes ned til fettsyrer og glyserol, og store karbohydrater brytes ned til monosakkarider, for eksempel glukose. Alle disse stoffene kan være med i delreaksjonene både i aerob og anaerob celleånding. Når celleåndingen foregår, blir energien «lastet over» i korttidslagre før den eventuelt blir frigitt som varme, overført til andre molekyler eller brukt til arbeid. De vanligste korttidslagrene i celleåndingen er:
• ATP (adenosintrifosfat)
• NADH (nikotinamid-adenin-dinukleotid)
• FADH₂ (flavin-adenin-dinukleotid)
Aerob celleånding
I aerob celleånding er oksygen nødvendig. Mange dyr har gassutvekslingsorganer og et sirkulasjonssystem som sørger for at oksygen kan transporteres inn i o rundt omkring i kroppen, og at noe av avfallet etter celleåndingen – karbondioksidet – blir fraktet ut. Planter har også aerob celleånding. Hos planter diffunderer luft med oksygengass og karbondioksidgass ut og inn gjennom spalteåpningene o bladene. Fåcellede dyr og planter kan ta opp nok oksygen og kvitte seg med karbondioksid ved diffusjon direkte gjennom cellemembranen o hver celle.
Vi starter med å se nærmere på hvordan glukose brytes ned. Den aerobe nedbrytningen av glukose består av mange delreaksjoner. Vi grupperer dem i tre hoveddeler:
- Glykolysen
- Krebssyklusen
- Oksidativ fosforylering
I de to første hoveddelene spaltes molekyler flere ganger. Dette frigir energi, som blir overført til energibærere i form av ATP, NADH og FADH2. I den tredje hoveddelen blir energi fra NADH OG FADH2, som ble dannet i de to første hoveddelene, overført til ATP. Vi regner ATP for den mest anvendelige formen for kortidslagre energi.
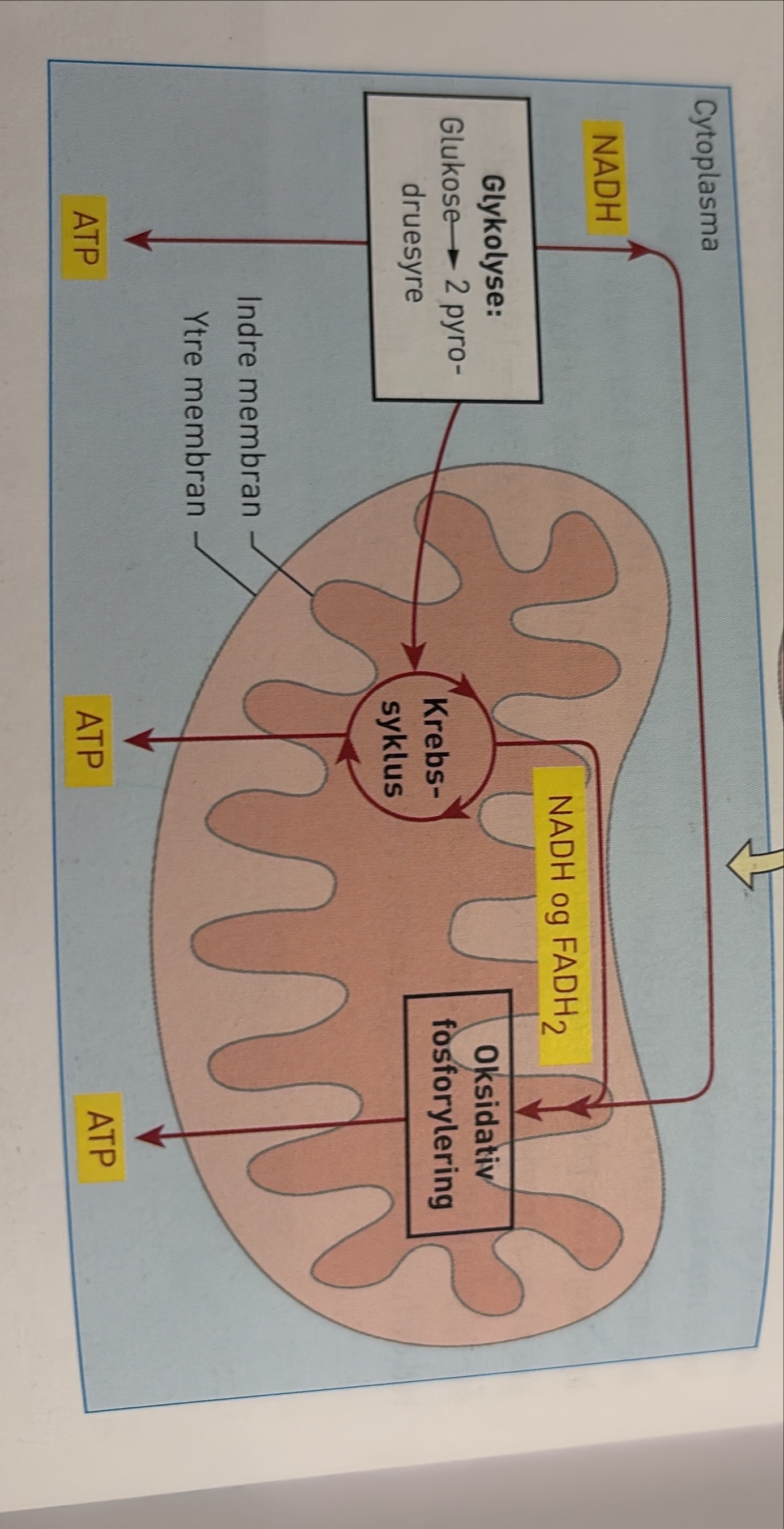
Glykolysen: nedbrytning av glukose
Glukose (sekskarbon, 6C) er den første forbindelsen i den hoveddelen vi kaller glykolysen. Glukose er et monosakkarid. Det blir dannet i fotosyntesen og når større karbohydrater, som glykogen, stivelse eller sukrose, blir spaltet. Alle levende organismer har fellestrekk i måten den biokjemiske nedbrytningen av glukose skjer på. Glykolysen skjer uten bruk av oksygen og foregår dermed på samme måte uansett om det er oksygen til stede eller ikke. I alle celler i alle levende organismer skjer glykolysen i cytosol, cellevæsken.
Nedbrytningen i glykolysen skjer gjennom mange delreaksjoner, og for hver delreaksjon er det et bestemt molekyl med spesifikke enzymer. Her tar vi bare med hovedtrekkene. Når ATP gir fra seg en fosfatgruppe til et annet molekyl, får det andre molekylet mer kjemisk energi. To ATP blir spaltet, og to energirike fosfatgrupper overføres til glukosemolekylet. Vi får et ustabilt energirikt molekyl, fruktose 1,6-bifosfat (6C). Fruktose 1,6-bifosfat blir raskt spaltet i to molekyler triosefosfat (3C). I neste delreaksjon blir triosefosfat omdannet, og fosfatgruppene og energien blir frigjort. Noe av energien blir da brukt i reaksjoner der fire elektroner og to H⁺-ioner blir bundet og danner to molekyler energirikt NADH. Samtidig blir fire ADP og fire ATP. To triosefosfat blir til to molekyler pyrodruesyre (3C).
Energibærerne som blir dannet ved nedbrytning av ett glukosemolekyl i glykolysen, er:
• 4 ATP (blir dannet) – 2 ATP (blir brukt) = 2 ATP
• 2 NADH
Nedbrytningen av glukose til pyrodruesyre blir regulert av ATP, som dermed oså fungerer som en enzymhemmer. Mye ATP hemmmer enzymene som skal virke i glykolysen, og ga stopper glykoslysen opp. Når ATP-mengden avtar, kommer glykolysen i gang igjen.
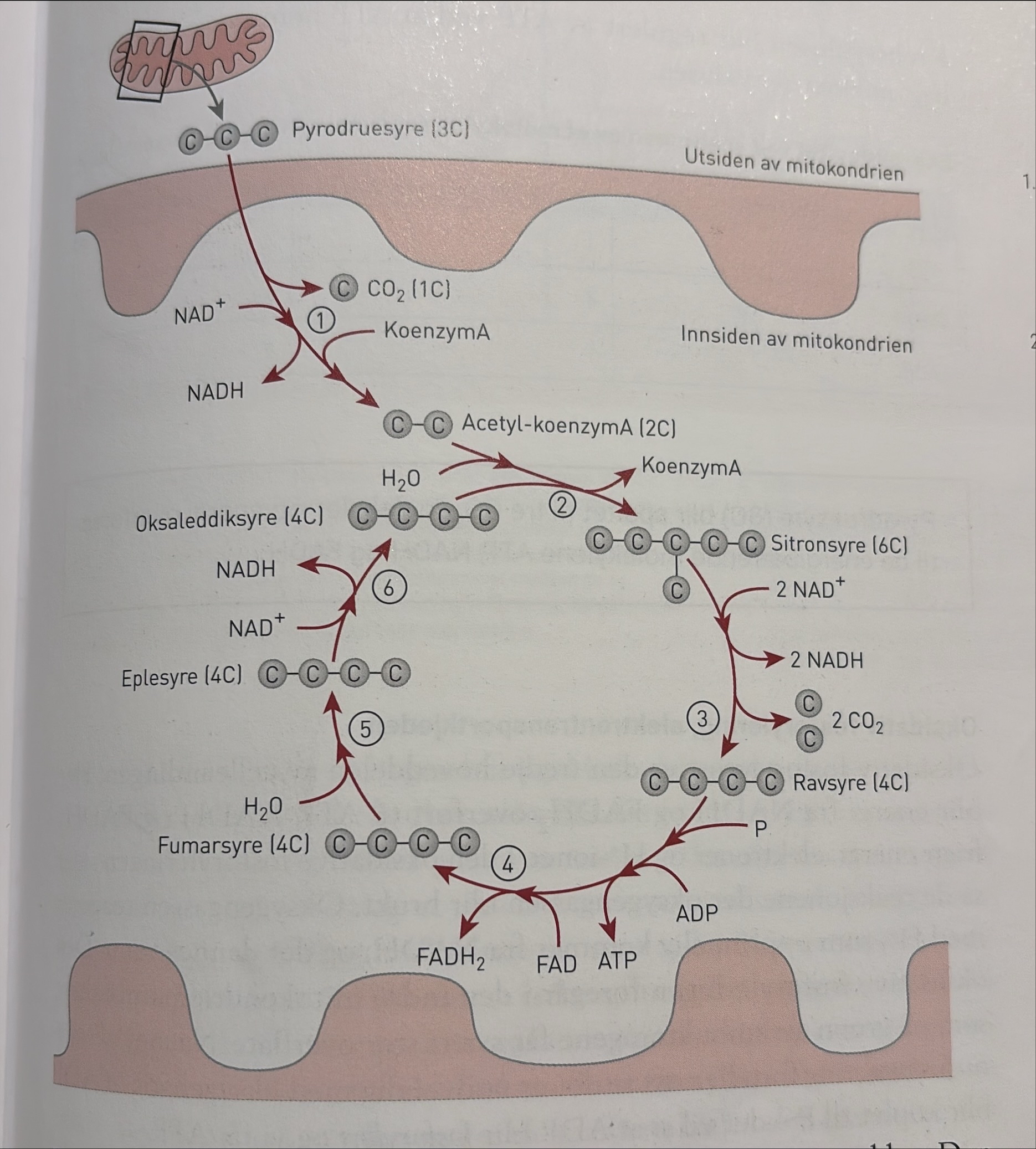
Krebssyklusen, sitronsyresyklus
Den videre spaltingen av pyrodruesyre skjer på forskjellige steder i prokaryote og eukaryote celler. Spaltingen skjer på forskjellig måte avhengig av om organismene har tilgang på oksygen, aerobt, eller ikke, altså anaerobt. Spaltingen varierer også fordi organismene produserer ulike enzymer. Dermed tar reaksjonene forskjellige veier. Figuren nedenfor viser en forenkling av glykolysen sammen med aerob og anaerob spalting av pyrodruesyre. Senere i delkapittelet kan du lese om den anaerobe spaltingen av pyrodruesyre.
Krebssyklusen er den andre hoveddelen av den aerobe celleåndingen. Vi følger ett pyrodruesyremolekyl videre, selv om hvert glukosemolekyl gir to pyrodruesyremolekyler. Når vi ser på energiutbyttet videre, må det derfor dobles når vi skal summere det totale energiutbyttet ved nedbryting av ett glukosemolekyl. Her skal vi se på spaltingen av pyrodruesyre i eukaryote organismer, som alle har mitokondrier i cellene. I mitokondriene skjer de kjemiske reaksjonene som frigir den energien som blir brukt når ADP og P danner ATP. Mitokondrien er omgitt av en dobbelt membran. Den ytre membranen er glatt, og den indre er buklet og får ved det en stor overflate.
Pyrodruesyremolekylet er så lite at det kan passere gjennom mitokondriemembranene ved diffusjon og komme inn i det innerste rommet i mitokondriene. Deretter går det inn i delreaksjonene i krebssyklusen.
Før pyrodruesyremolekylet (3C) går inn i krebssyklusen, blir det spaltet av ett karbondioksidmolekyl (1C). Denne reaksjonen frigir noe energi til dannelsen av ett NADH. «Resten», en tokarbonforbindelse (2C) som blir kalt acetyl, blir koplet til et koenzym, koenzymA. Da får vi acetyl-koenzymA (2C). Acetyl-koenzymA går inn i syklusen der acetyl (2C) reagerer. Da løsner koenzymA og kan så brukes på nytt. Vi kan si at koenzymA fungerer som en bærer for tokarbonforbindelsen acetyl inn i krebssyklusen.
Krebssyklusen består av mange delreaksjoner som går i en syklus. Den første delreaksjonen begynner med at oksaleddiksyre (4C), som var en rest etter forrige runde, kopler seg til acetyl (2C) og vann og blir med inn i en ny runde av delreaksjoner. Produktet i den første delreaksjonen er sitronsyre (6C), og derfor blir krebssyklusen også kalt sitronsyresyklusen.
I de to neste delreaksjonenene spatltes to karbondioskidmolekyler av sitronsyre, og vi får ravsyre. Strukturen i firekarbonforbindelsensen blir byg om flere anger, fra ravsyre, til funarsyre ogg videre til eplesyre, uten at detblir spaltet av karbon. Ved ombygging fra fumarsyre til eplesyre bindes ett vannmolekyl. Strukturendringene fra ravsyre til oksaleddiksyre gjlr at elekjtroner, H+-ioner, fosfat og frigitt energi blir brukt til å danne enrgibærede forbindelser: ett molekyl ATP, ett molekyl FADH2 og ett molekyl NADH.
Krebssyklusen blir regylert av ATP ved at ATP hemmer enzymene i begynnelsen av syklusen.
Steg for steg:
1. Pyrodruesyremolekylet (3C) inngår i en reaksjon som avgir ett karbondioksidmolekyl, ett H⁺-ion, to elektroner og nok energi til at NAD⁺ kan danne NADH. «Resten» (acetyl, 2C) blir koplet til et koenzym, og vi får acetyl-koenzymA (2C).
2. KoenzymA løsner fra acetyl-koenzymA. Acetyl går inn i krebssyklusen og binder seg til et molekyl oksaleddiksyre (4C). Et molekyl vann blir også bundet. Vi får sitronsyre (6C). KoenzymA kan brukes på nytt.
3. I to nye delreaksjoner spaltes to karbondioksidmolekyler av fra sitronsyren, og vi får ravsyre (4C). Samtidig med at karbondioksid går ut, frigjøres nok energi til at to NAD⁺ danner to NADH.
4. Strukturen i firekarbonforbindelsen blir bygd om flere ganger, først fra ravsyre til fumarsyre (4C). Strukturendringene frigjør energi som blir lagret i energibærende forbindelser: ett molekyl ATP og ett molekyl FADH₂.
5. Videre omdannes fumarsyre til eplesyre (4C) uten at det blir spaltet av karbon. Ved ombyggingen fra fumarsyre til eplesyre bindes et vannmolekyl.
6. Når eplesyre blir omdannet til oksaleddiksyre (4C), blir det frigjort nok energi til at NAD⁺ kan danne NADH.
Oksidativ fosforylering, elektrontransportkjeden
Oksidativ fosforylering er den tredje hoveddelen av celleåndingen. Her blir energi fra NADH og FADH₂ overført til ATP. NADH og FADH₂ frigir energi, elektroner og H⁺-ioner. I den oksidative fosforyleringen skjer så de reaksjonene der oksygengassen blir brukt. Oksygengassen reagerer med H⁺ som opprinnelig kommer fra NADH, og det dannes vann.
Den oksidative fosforyleringen foregår i den indre mitokondriemembranen, som på grunn av innbuktningene får svært stor overflate. Navnet oksidativ fosforylering forteller oss at det er nødvendig med oksygen, og at ADP blir koplet til P – det vil si at ADP blir fosforylert og vi får ATP.
Den reaksjonen vi nå skal beskrive for NADH, skjer på tilsvarende måte for FADH₂. Elektronene fra NADH blir fanget av spesielle proteiner i mitokondriemembranen og blir transportert fra protein til protein. Proteinene fungerer vekselvis som elektronakseptorer og elektrondonorer.
Reaksjonen ikner den som foregår i tylakoidmembranen i klroplastene i fotosystem II ig i, dne lysavhengige reakjsonen.
Den oksidative fosforyleringen blir også kalt elektrontransportkjeden. Når elektronene går fra protein til protein, blir det frigitt energi. Energien blir brukt til å pumpe H⁺-ioner gjennom transportkanaler fra det innerste rommet i mitokondriene og ut i mellomrommet mellom de to mitokondriemembranene. Denne reaksjonen gir et overskudd av positive ladninger i mellomrommet. Da blir det en ladningsforskjell – en spenning – fordi mellomrommet blir mer positivt og det innerste rommet i mitokondriene mer negativt. Ladningsforskjellen mellom mengden av H⁺-ioner utenfor og innenfor membranen kalles en protongradient både i den oksidative fosforyleringen i aerob celleånindg og i fotosyntesens fotosystemer.
I dne indre membranen ligger ogsp transportkanaler med ATP-ase enzymer. Ladningsforksjellen mellom det indre rommet og mellomrommet i mitokondrien gjør at H+-ionene blir slupet gjennom transportknalene og går fra mellomrommet og tilbaktil det innerste rommet.
Samtidig blir det frigjkort energo. Denne energien blir brukt i reakjsonen der ADP o fosfatgruper danner ATP. ATP-ase katalyserer reakjsonen. H+-ionene, som nå er i overskudd i det innerste rommet, blir koplet til oksygengass, og danner vannmolekyer. Oksygen blir altså brukt først når vi kommer til den siste delreakjsonen i den aerobe nedbryntingen. Det er tilgangg på oksygen som driver reakjonen videre. Fordi H+ fra spaltningen av NADH blir fjernet og ATP blir prodsert. Mangel på oksygen stopper den oksitatice fosforyleringen, og derretter glykolysen, slik at ikke noe pyrodruesyre går inn i krebssyklusen.
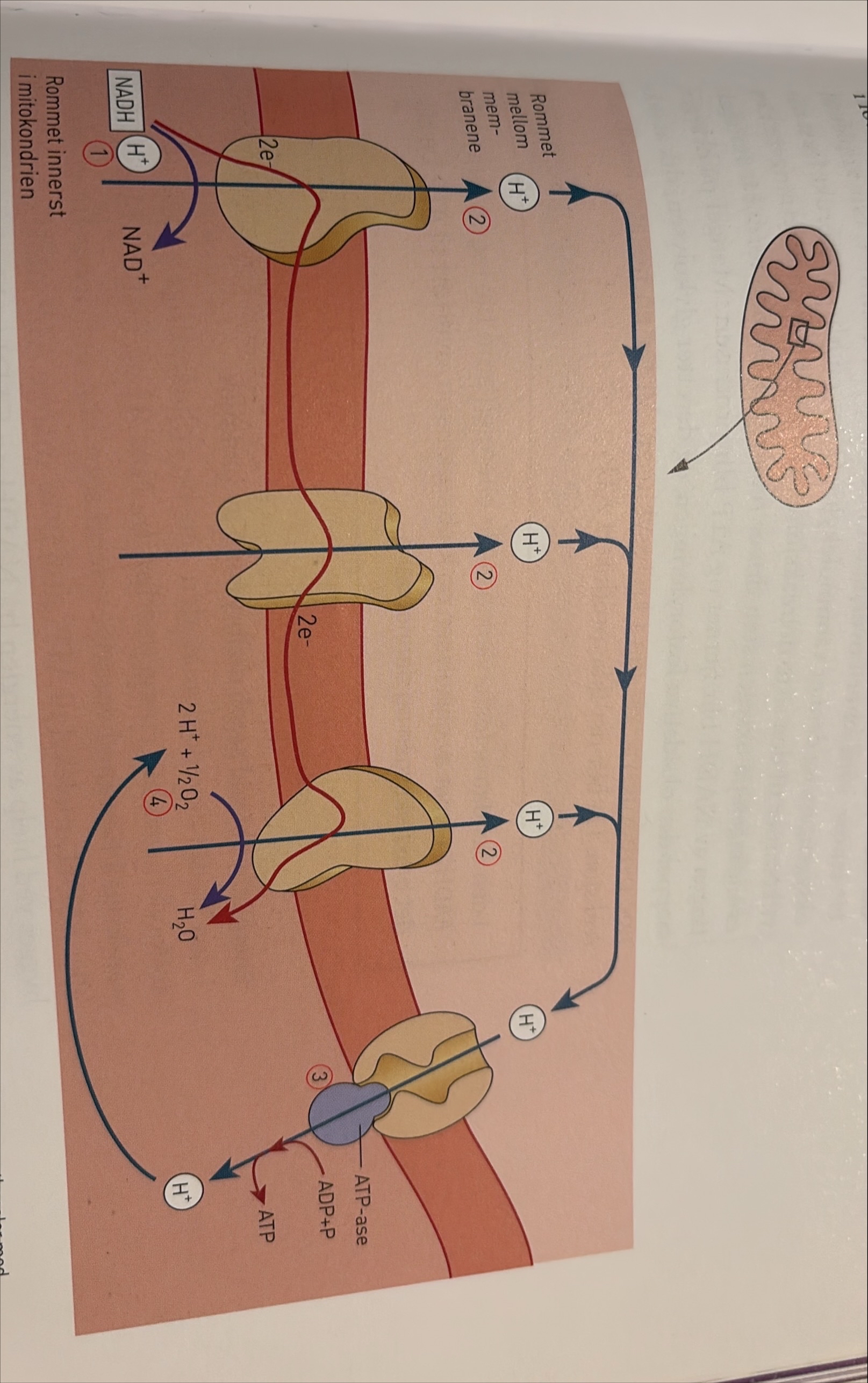
Energiutbyttet ved aerob nedbrytning av glukose
I den tredje hoveddelen av den aerobe celleåndingen, den oksidative fosforyleringen, blir energi overført fra NADH og FADH₂ til ATP. Det er vanskelig å beregne hvor mye energi som kan overføres fra et NADH- eller FADH₂-molekyl til ATP, altså hvor mange ATP-molekyler som kan bygges ved hjelp av energien fra NADH og FADH₂. Vanligvis antar vi at energien i to NADH kan danne fem ATP, mens energien i to FADH₂ er nok til å danne tre ATP. De to NADH-molekylene som ble dannet i cytosol ved glykolysen, må fraktes inn gjennom mitokondriemembranene, der energien fra NADH kan overføres til ATP. Transporten er aktiv, det vil si at den er energikrevende.
Tidligere antok man at det “kostet” ett ATP per to NADH å få dem gjennom membranen, men det har vist seg at den energimengden som kreves, er svært liten. Derfor blir det totale energiutbyttet ca. 32 ATP. Før ble energiutbyttet ved spalting av et glukosemolekyl målt til 36 ATP, men nyere beregninger angir et litt lavere utbytte. Det er vanskelig å si sikkert om det riktige antallet er 36 eller 32.
Figuren ved siden av viser den aerobe nedbrytningen av glukose til karbondioksid, vann og energibæreren ATP. ATP blir dannet i de katabolske reaksjonene som skjer når glukose blir brutt ned i glykolysen, krebssyklusen og den oksidative fosforyleringen. Også energien i FADH₂ og NADH blir overført til ATP.
NADH fra glykoluysen mp inn i mitokondriee ved aktiv transort for å få overført energien til ATP, men den energiemngden som kreves, er svørt liten. Totalt blir det dannes ca 32 ATP per gluoksemoleyl.
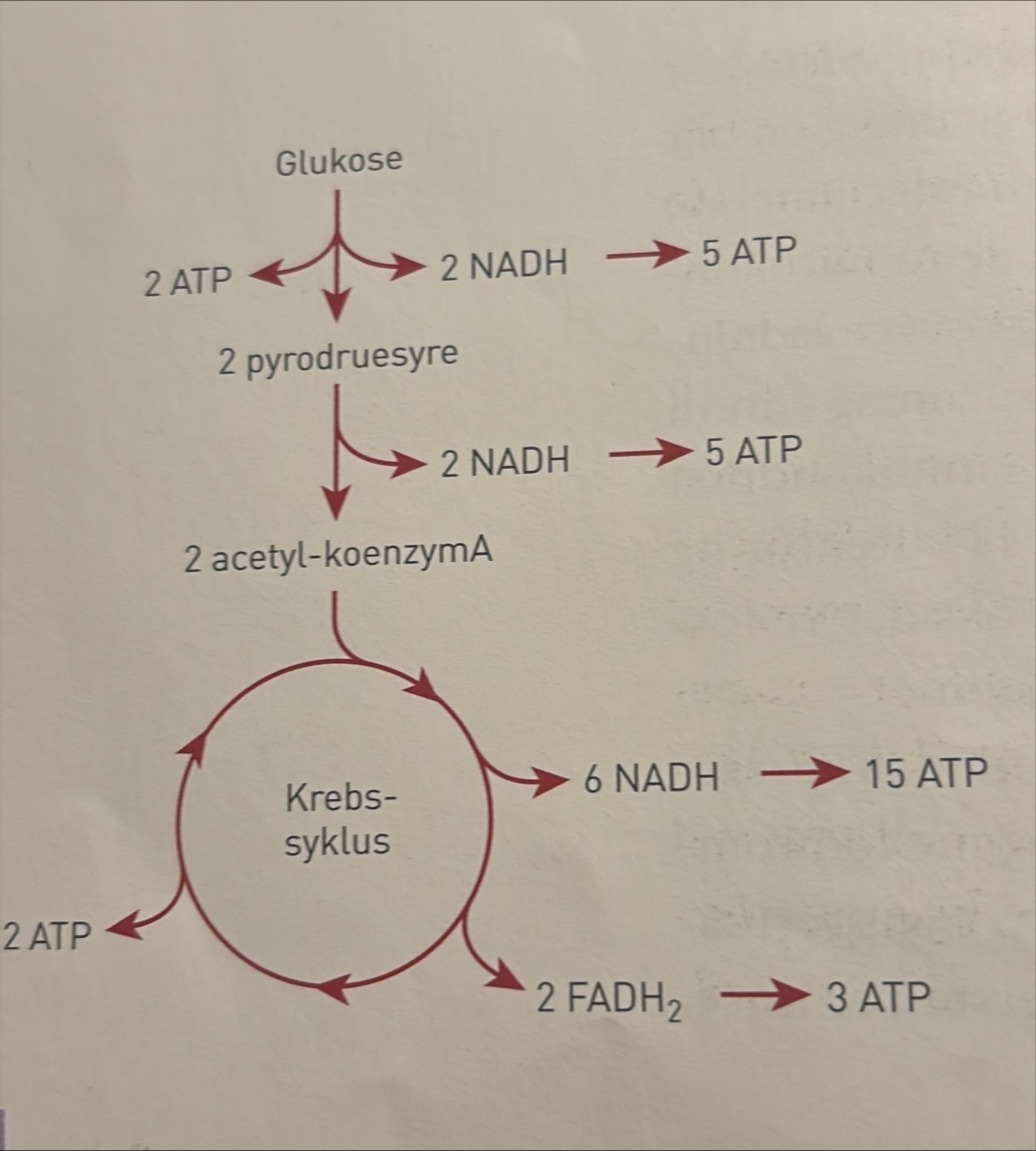
Anaerob celleånding
Den anaerobe nedbrytningen av glukose gir mye mindre energi enn den aerobe nedbrytningen, men organismene som driver med anaerob nedbrytning, får likevel et visst energiutbytte. Mange organismer dør dersom de kommer i et aerobt livsmiljø fordi oksygen er giftig for dem. De kan bare leve i et anaerobt miljø og ha anaerob nedbrytning.
De fleste prokaryote og eukaryote organismer har aerob celleånding som gir mer energi enn den anaerobe. Dersom det er dårlig tilgang på oksygen, kan de likevel ha anaerob nedbrytning i kort periode som en midlertidig løsning. Pattedyr vil kunne ha en slik løsning bare i noen få minutter. Når vi mangler oksygen, får cellene for lite ATP. Vi merker raskt ubehag, og hvis det varer for lenge, vil vi besvime og til slutt dø.
Når en organisme som normalt har aerob celleånding, går over til anaerob celleånding, vil oksygenmangelen gjøre at den oksidative fosforyleringen stopper opp. Da vil ikke pyrodruesyren gå inn i krebssyklusen, men må spaltes videre anaerobt. Produktene varierer hos de forskjellige organismene. Noen produserer karbondioksid samtidig, andre ikke. Hos melkesyrebakterier og gjærsopp finnes det enzymer som er tilpasset en anaerob nedbrytning av pyrodruesyre. Enzymene i dem er forskjellige, og nedbrytningsproduktene blir ulike: for eksempel melkesyre eller etanol.
All anaerob nedbrytning kaller vi gjæring eller fermentering, også den melkesyreproduksjonen som foregår i musklene våre når vi får for lite oksygen. I dagligtalen sier vi gjæring om det som skjer i brøddeigen når den hever, når korn gjærer til øl, eller når druer gjærer til vin, mens ølprodusent eller en vinbonde gjerne vil bruke begrepet fermentering. Uansett om vi kaller det gjæring, fermentering eller anaerob nedbrytning, er det snakk om den samme prosessen.
Prokaryote organismer, for eksempel melkesyrebakterier, har ikke mitokondrier, men de kan ha innbuktninger i cellemembranen som gjør membranoverflaten større. Enzymene som er nødvendige for nedbrytningen av glukose og den videre nedbrytningen av pyrodruesyre, er festet til cellemembranen.
Hos organismer som bryter ned glukose anaerobt, foregår glykolysen akkurat slik som hos organismer med aerob nedbrytning. Vi forklarer derfor ikke denne prosessen her.
Fra pyrodruesyre til melkesyre
Når du trener hardt, trenger du mye energi raskt. Store karbohydratmolekyler blir brutt ned til glukose, og den videre nedbrytningen av glukose gir deg energi. Når du trener, puster du ekstra dypt. Du utvider lungevolumet seg slik at du får mye oksygen ned i lungene. Likevel kan det hende at du ikke får nok oksygen til bare aerob celleånding, og da må muskelcellene gå over til anaerob nedbrytning av glukosen. Mennesker og andre pattedyr har enzymer for å kunne ha anaerob nedbrytning av glukose når tilgangen på oksygen er for liten til å drive aerob. Da blir pyrodruesyre til melkesyre. Den anaerobe nedbrytningen til melkesyre er reversibel. Reaksjonen kan altså snu og gå tilbake. Når vi hviler og puster inn nok oksygen, blir melkesyren omdannet til pyrodruesyre som går inn i krebssyklusen og blir til oksidativ fosforylering, se figur 190. Melkesyreopphoping i musklene gjør deg sliten. Toleransen for melkesyre i musklene varierer fra person til person, og vi kan trene oss opp til å tåle mer. Når musklene får lite nok ATP, kan de ikke strekkes eller trekke seg sammen. Da snakker vi om å stivne. Hvis du beveger deg litt og tøyer ut etter litt hviling, får du lettere transportert oksygenrikt blod til musklene, og da blir melkesyren omdannet. Noe fortsatt er det kr opphopingen av melkesyre fra anaerob nedbrytning. Cellene i øyet er svært følsomme for økt varr, for eksempel cellene på baksiden av øyet som danner blodårer, så mye at de klager når de har stri. Derfor har øyet høy toleranse for melkesyre, og blodsårene trekker seg sammen når de får for lite oksygen, blant annet under hard fysisk aktivitet. Da går cellene over til anaerob nedbrytning av glukose til melkesyre. Resten av kroppen må ha aerob nedbrytning av glukose for å få nok energi til blant annet bevegelse, transport gjennom membranene og kjemisk arbeid.
Når vi lager melkeprodukter som drikkeyoghurt og rømme, bruker vi vanlig søt melk og tilsetter melkesyrebakterier, for eksempel Lactobacillus. I glykolysen og den anaerobe nedbrytningen av pyrodruesyre bruker Lactobacillus de samme enzymene som vi gjør. Først gjør bakteriene melkesukkeret om til glukose, deretter gjør de glukosen om til pyrodruesyre, og så gjør de pyrodruesyren om til melkesyre. Melkesyre fra Lactobacillus gjør at melkeproteinene blir denaturert. Det får melken til å tykne. Vi kan bruke den tykkende melken til å lage blant annet rømme, kefirmelk og yoghurt. Ved anaerob nedbrytning til melkesyre blir det ikke dannet karbondioksid:
1 glukose + 2 P + 2 ADP → 2 melkesyre + 2 ATP + 2 vann
Energiutbyttet per glukosemolekyl er bare 2 ATP mot 32 i den aerobe nedbrytningen. De to molekylene NADH som ble dannet i glykolysen, blir brukt når pyrodruesyre blir til melkesyre, og vi får igjen NAD⁺.
Fra pyrodruesyre til etanol/eddiksyre
Gjærsopp kan ha både aerob og anaerob nedbrytning av pyrodruesyre. Ved anaerob nedbrytning blir pyrodruesyre omdannet til alkoholen etanol og karbondioksid. Eksempler på gjærsopper er brødgjær, vingjær og ølgjær. Brødgjær og ølgjær bryter ned stivelsen i kornet til glukose. Vingjær bryter ned sukrose og fruktose i frukten til glukose. Under nedbrytningen dannes det samtidig smaks- og aromastoffer.
Når vi lager en brøddeig, bør væsken ha en temperatur på omtrent 37 °C, for da deler soppcellene seg raskest og driver med mye anaerob ånding. Karbondioksidet de produserer, hever brøddeigen. Deig som står og hever, lukter alkohol. Når deigen blir stekt, fordamper alkoholen.
Energien fra glykolysen frigjøres som varme, og temperaturen i deigen stiger. Nedbrytningen foregår slik:
1 glukose + 2 P + 2 ADP + 2 H⁺ →
etanol + 2 karbondioksid + 2 ATP + 1 vann
Også gjærsopp bruker opp de to NADH-molekylene fra glykolysen når den danner etanol, slik at netto energiutbytte fra ett glukosemolekyl er 2 ATP. Alle gjærsoppceller inneholder de spesielle enzymene som er nødvendige for å danne etanol. Etanoldannelsen er irreversibel. Etanol kan altså ikke «gå tilbake» og bli til pyrodruesyre igjen. Gjærsoppcellene dør når alkoholmengden i brøddeigen, vinen eller ølet er ca. 15 prosent.
Eddiksyrebakterier inneholder enzymer som kan bryte ned pyrodruesyre til eddiksyre. Dette skjer anaerobt. Eddiksyrebakterier kan også omdanne etanol i en alkoholsvek løsning som vin eller øl til eddiksyre. Denne reaksjonen skjer imidlertid aerob. Eddiksyrebakterier finnes normalt i lufta. En åpen flaske øl eller vin har tilgang til oksygen. Når flasken blir stående framme, vil etanolen i ølet eller vinen omdannes til sur eddik. Eddiksyrebakteriene tåler en svært lav pH og blir ikke drept under dannelsen av eddik. Før benyttet man seg av denne reaksjonen når man produserte vineddik, men nå foregår produksjonen vanligvis på raskere måter uten bruk av eddiksyrebakterier.