T2 Licausi- Plant development I - plants are composed of repeated structural units
1/9
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
10 Terms
how do the directions of growth define plant shapes?
apical growth is elongation up and downwards
branching enables the lateral exploration of space
radial growth allows secondary thickening
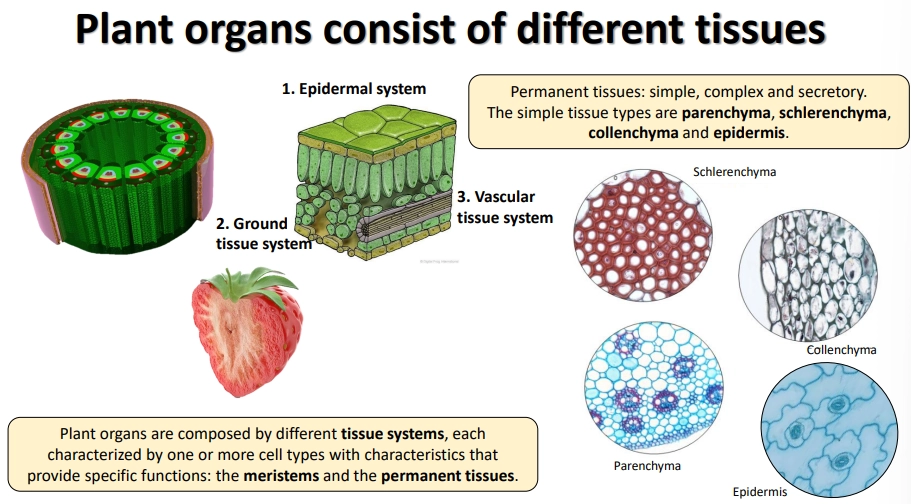
what are the different plant tissues and tissue systems?
the permanent tissue systems (non-meristem):
epidermal system
vascular tissue system
ground tissue system
these are made up of simple, complex and secretory tissues
the simple tissues are:
schlerenchyma- lignified cell walls + cell death eg. xylem
colenchyma- live cells modified with secondary thickening of the cell wall
parenchyma- thin cell walls
epidermis- thin cells + stomata
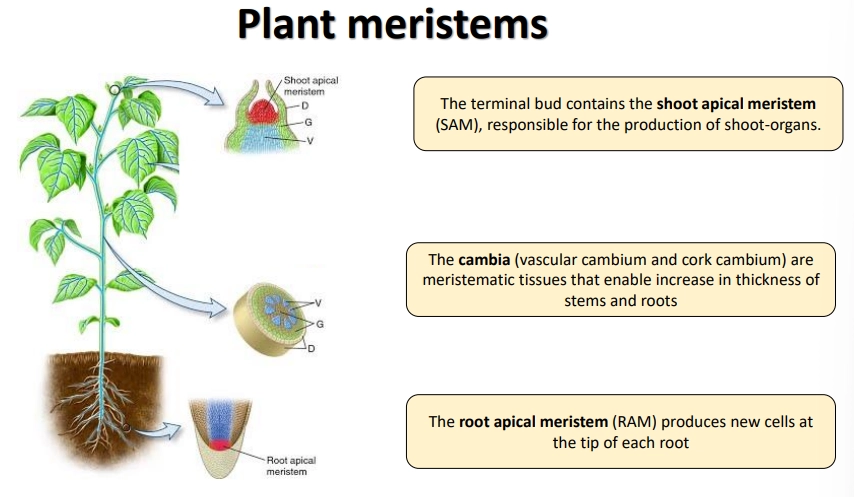
where are plant meristems found?
the shoot apical meristem (SAM)
the vascular cambium of the stem
the root apical meristem (RAM) + lateral root meristem
meristems are also produced at regular intervals in the stem that remain inactive- these allow regeneration following damage from herbivores, wildfires, etc, compensating for their inability to move, e.g., adventitious roots
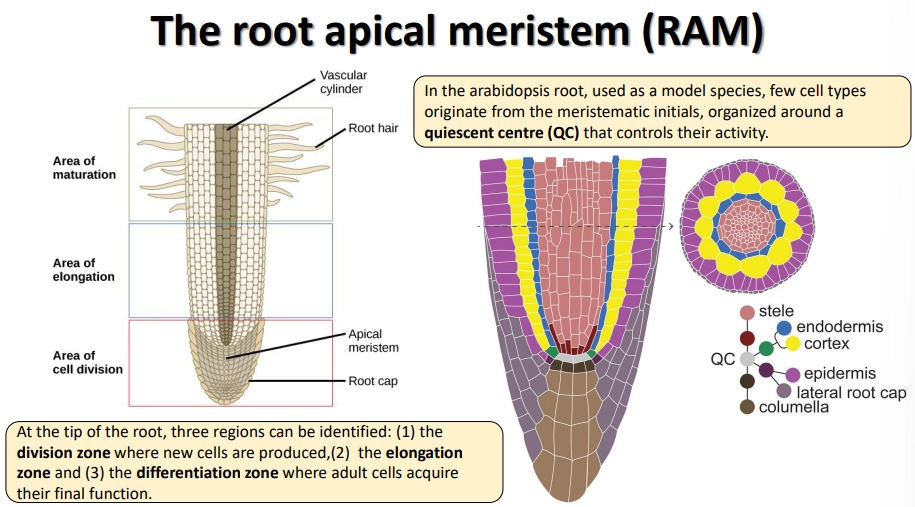
what is the structure of the root apical meristem?
at the tips of roots, there are three regions: the division zone, the elongation zone and the maturation zone (for the development of secondary features)
at the dividion zone of Arabidopsis, the rate of meristem cell division is controlled by a cluster of 4 cells called the quiescent centre
the root cap is made up of the lateral root cap itself and the columella, a cluster of cells at the tip that contain amyloplasts used to sense the direction of gravity
the new cells then differentiate into the stele, endodermis, cortex and epidermis (including root hairs)
root hairs are produced at epidermal cells that are in contact with two inner cortex cells, due to receiving sufficient diffusible signals from the edges of these cells
lateral root primordia (LRP) are inactive meristems at regular intervals that don’t activate until the cells from the outermost layer of the stele (pericycle) begin to divide and break through, due to auxin peaks
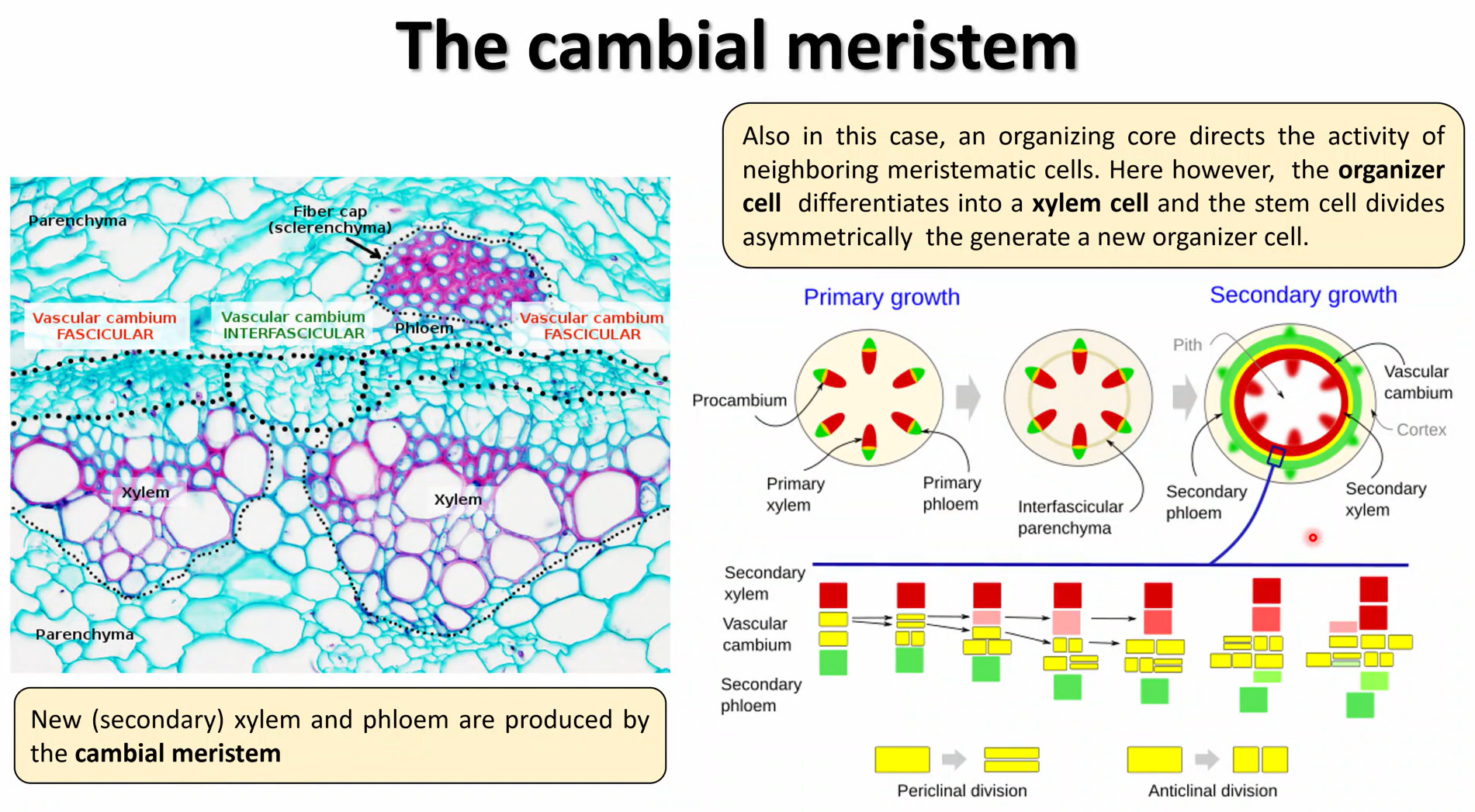
how does the cambium cause secondary thickening?
division is activated when the interfascicular parenchyma ring is formed
the cambium cells close to the xylem side divide periclinally (horizontally) and differentiate early
the cambium cells close to the phloem divide anticlinally (vertically) and do not differentiate as early, to increase the number of cells per layer
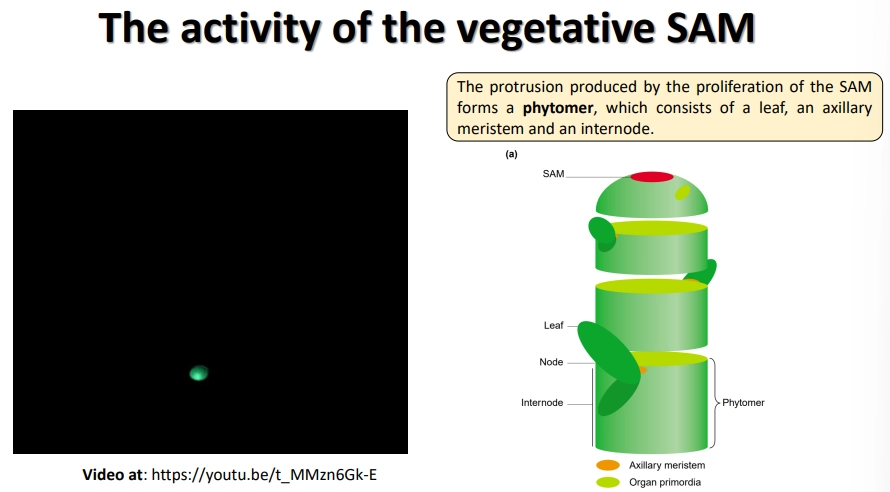
what is the structure of the shoot apical meristem?
the shoot apical meristem is highly protected in layers of leaf primordia
the inner meristem dome can be separated into three layers of meristem cells
the organising centre is a pool of up to 100 cells, which determines the speed of division + identitiy of neighbouring cells
the SAM differentiates the stem in regular units called phytomers, which each contain a primordial leaf at a node, an inactive axillary meristem and an internode region
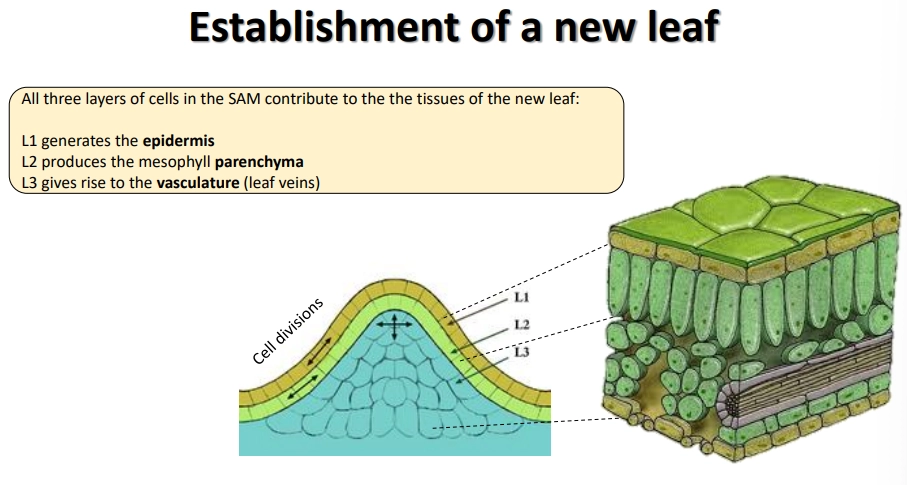
how are leaves produced at the shoot apical meristem?
the inner meristem dome of the SAM can be separated into three layers of meristem cells
to produce leaves, the 1st layer generates the epidermis, the 2nd produces the mesophyll parenchyma, and the 3rd produces the vascular tissue
the petiole contains the midrib/vein, and the flat layer of cells is the blade- leaf shape is genetically determined and defined by serrations, lobes and leaflets (compound leaves)
the adaxial (upper) side is optimised for light capture and the reduction of water loss, while the abaxial (lower) side is optimised for gas exchange- the polarity of leaf primordia is established early, defined by which side is closer to the SAM
once the leaf cells have stopped dividing, they normally continue to replicate their DNA, because polyploidy supports more active metabolism
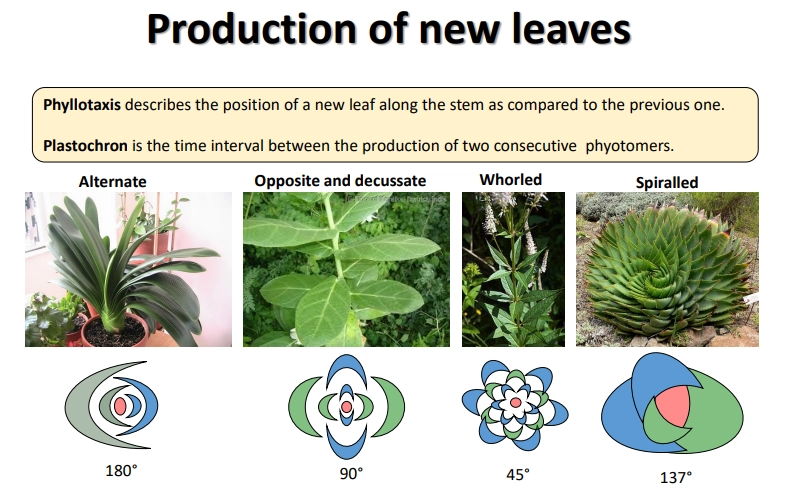
what is phyllotaxis and what are the different forms?
spiralled is the most common- less likely to shade previous leaf
how are reproductive structures produced at the shoot apical meristem?
the SAM transitions from vegetative to reproductive (it becomes the inflorescence meristem)
the infloresence meristem loses phyllotaxis and produces many determinate flower meristems, that produce concentric whorls of new organs
these whorls are the sepals (together form the calyx), the petals (together form the corolla), the stamens, the carpel (multiple carpels form a pistil)- (the sepals + petals are the perianth)
shoot architecture is dependent on: if the vegetative meristems along the stem also turn into reproductive meristems, if the reproductive meristems terminate with flowers or continue producing more meristems, and how the terminal flower meristems branch
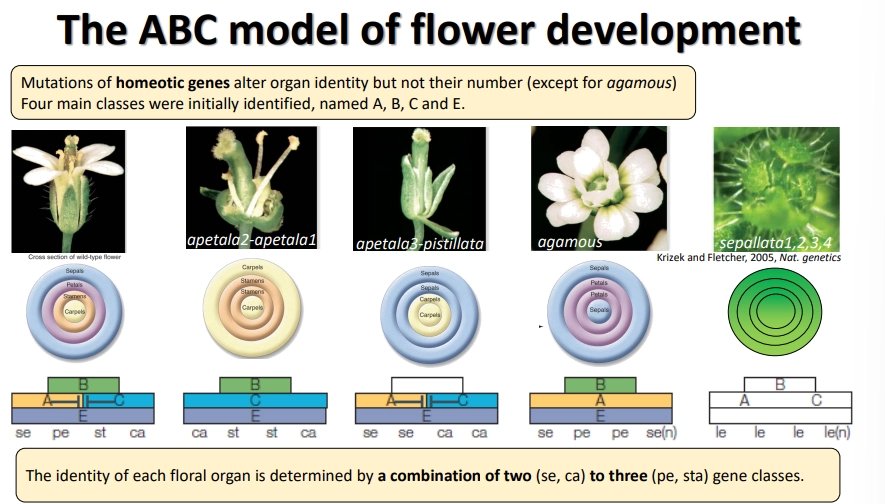
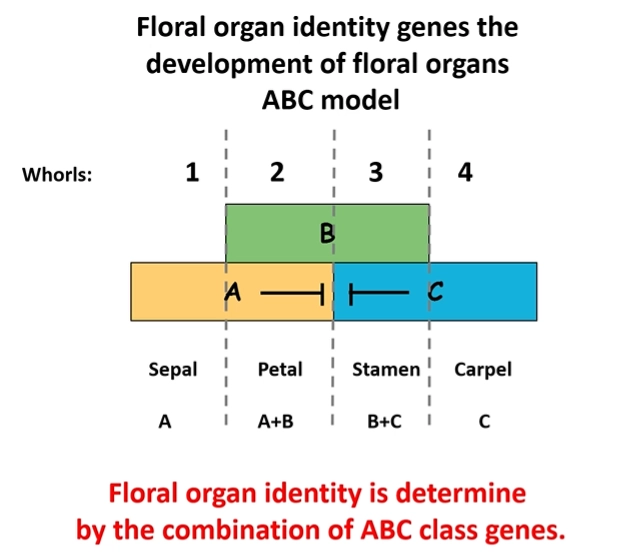
how is flower development controlled by genes?
five main classes of homeotic flower genes were found in Arabidopsis: A, B, C, D and E
the development of each floral organ in the whorl is dependent on a combination of two or three gene classes that get expressed
A and C mutually repress each other’s expression
all organs need E class genes to be present
eg. sepals need A and E genes- if A is lost, they will have C and E genes and become carpels
the D class was not identified for a long time due to high gene redundancy- redundancy is also present within the other classes in different species
class C genes are also responsible for terminating the meristem, so C gene homeotic mutants produce an indeterminate number of organs