W8 - Neuroplasticity
1/26
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
27 Terms
neuroplasticity
the capacity for neuronal change
happens in intact nervous system and after injuring it
types of neuroplastic changes
structural
anatomical changes
eg. to neuronal connections/size of cortical areas
functional
structures deviating from the initial function
phantom limb syndrome
Lotze et al. (2001)
participants:
phantom limb with pain
phantom limb without pain
healthy controls
participants imagined a movement:
patients with phantom limb pain showed activity in the face area (corresponding to the mouth) when imagining hand movements
participants pursed their lips:
patients with phantom limb pain showed activity patterns extending to the hand areas in S1 (and M1)
what does Lotze et al. (2001)’s phantom limb study show
there is selective coactivation of the cortical hand and mouth areas in patients with phantom limb pain
this reorganizational change may be the neural correlate of phantom limb pain
aka phantom limb pain is closely linked to cortical reorganization
what causes brain damage
tumours
strokes
infections
neurological diseases
injuries
neuroplastic responses to nervous system damage
degeneration/deterioration
regeneration/regrowth of damaged neurons
reorganisation
recovery
Pinel and Barnes (2017)
degeneration/deterioration
important aspect of brain development
observed in a healthy nervous system in early development
but also a characteristic of disease and neuron death
synaptic pruning during childhood
3 types:
anterograde
retrograde
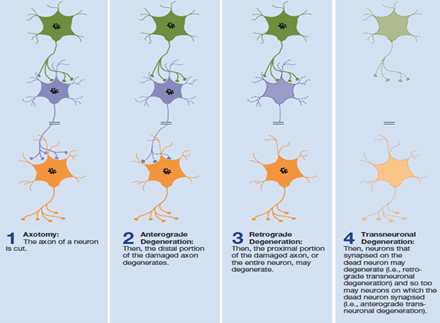
these 2 from axotomy (a cut in an axon)
transneuronal
synaptic pruning
process of removing weak/unused neural connections (synapses) to strengthen the most used ones
maximises the efficacy of mature neural circuits
occurs most at 4-6 years old
before this is just synapse formation
2 years has the highest amount of synapses
anterograde degeneration
distal segment swells up and degenerates
occurs within days following axotomy
retrograde degeneration
proximal segment degenerates
typically leads to entire neuron death
occurs after anterograde degeneration
transneuronal degeneration
when degeneration spreads from site of damage to adjacent neurons
anterograde transneuronal degeneration
spreads from damaged neuron to ones they’re synapsed onto
retrograde transneuronal degeneration
spreads to neurons synapsing on the damaged cell
regeneration
regrowth of damaged neurons
less accurate in mammals and higher vertebrates
but can take place during the early stages of development, before adulthood is reached
CNS neurons do not regenerate in adult mammals
but there is possibility for PNS neurons to regenerate
regeneration of PNS neurons
due to Schwann cells
these are glial cells that produce myelin sheath in the PNS
they produce neurotrophic factors that stimulate growth of new axons
cell-adhesion molecules provide a pathway for axonal growth
why can’t CNS neurons regenerate in adult mammals
due to oligodendroglia - Silver et al. (2015)
these are glial cells that produce myeline sheath in the CNS (remyelination)
which actively blocks axon regeneration, following injury
leads to permanent functional deficits like strokes and spinal cord injuries
instead of regenerating damaged CNS cells, the brain relies on plasticity/reorganisation to take over lost/damaged functions
reorganisation mechanisms
strengthening of existing connections through the release of inhibition of adjacent nerves
collateral sprouting
vacant synapses become occupied and establish new functional pathways and connections
synaptic plasticity
strengthening/weakening existing synapses based on activity
long-term potentiation/depression
differs to pruning, which is eliminating synapses to increase brain efficiency
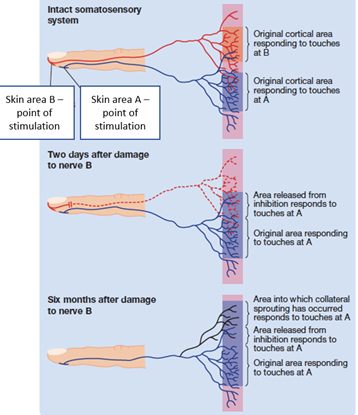
collateral sprouting
the formation of new axon branches in neighbouring neurons which then synapse at vacant sites left by degenerated axons
forms new functional pathways
evidence for cortical reorganisation in lab animals
Sanes et al. (1990)
cut the motor neurons which controlled muscles of rats’ whiskers
a few weeks later
stimulating regions of the motor cortex now activated muscles of the face rather than whisker movement
evidence that other areas of brain take over, potentially strengthening, their functions
there is rapid reorganisation of the mammalian motor cortex following peripheral nerve lesions
within 4 hours
the motor cortex is dynamically organised
evidence for cortical reorganisation in humans - Amedi et al. (2005)
visual areas of the brain activated when blind participants perform somatosensory discrimination tasks
where reorganisation can occur
after strokes
after nerve damage
in amputees
in blind people
recovery
recovery of functions following CNS damage is poorly understood due to:
biological limitations of regeneration
complexity of the nervous system
individual differences in injuries = hard to compare results
poor ability to distinguish recovery from compensatory mechanisms (plasticity, reorganisation)
treatment of nervous system damage
blocking neurodegeneration
neurotransplantation
rehabilitative training
blocking neurodegeneration - Xu et al. (1999) rat study
damaged hippocampus in rats
apoptosis of neurons in this area
programmed cell death
deficits in performing spatial learning task
introduction of a neuronal apoptosis inhibitor protein (NAIP) via a virus:
reduced neuron loss
better task performance
blocking neurodegeneration - Samantaray et al. (2011)
examined the effectiveness of low doses of estrogen following spinal cord injury in rats
estrogen administration found to reduce apoptosis/cell death and inflammation following injury
estrogen serves as a neuroprotective agent
study did not focus on behavioural outcomes however
suggests estrogen as potential factor in spinal cord injury treatment
neurotransplantation - Cheng et al. (1996)
transected the spinal cord of rats, making them paraplegic
implanted small sections of myelinated peripheral nerves (MPN) which bridged the gaps in the spinal cord
from Schwann cells
lead to regeneration of spinal cord neurons
this improved hind limb function in the rats
rehabilitative training - strokes
Nudo et al. (1996)
induced leisons in the hand area of monkeys’ motor cortex
those who went through intensive therapy on the affected limb showed greater functional recovery and reduced cortical damage in M1
shows neurotransplantation treatments might be more effective if accompanied by appropriate training
rehabilitative training - constraint-induced therapy
Kwakkel et al. (2015)
involves reducing the functioning of the intact limb and training the impaired one
leads to:
improved performance of the affected limb
cortical reorganisation favouring representation of the affected limb
rehabilitative training - facilitated walking
supporting spinal cord injury patients with a harness improves locomotion by producing greater speed and - coordination
results in patients being able to walk independently gradually