HC6: Viruses and viral infections
1/40
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
41 Terms
Virus: onderdelen
type nucleïne zuur: DNA of RNA
proteine capside
zorgt voor de productie van nieuwe virusdeeltjes die genomisch materiaal transporten naar een andere gastheercel
vermenigvuldigt in de cel en met de hulp van de eiwit synthese van de cel
Virus: structuren
→ structuur van het virus wordt bepaald door de proteine capside
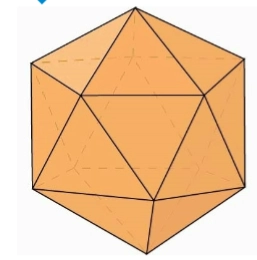
icosahedron
= 20 triangular faces, minimal 3 protein particles (capsomere) per face
helix
Virus: met/zonder envelop
Eigenschap | Virus ZONDER envelop (Naakt) | Virus MET envelop |
|---|---|---|
Buitenkant | Hard eiwitkapsel (capside) | Flexibele vetlaag (lipiden) met spikes |
Uitgang uit de cel | Laat de gastheercel exploderen (lysis) | Knoopt rustig naar buiten (budding) |
Overdracht | Kan makkelijk via oppervlakken of ontlasting (fecaal-orale route) | Vaak via direct contact of vochtruppeltjes (hoesten/bloed) |
Gevoelig voor alcohol/zeep | Nee, overleeft alcoholgel vaak wel | Ja, gaat direct dood door zeep of alcohol |
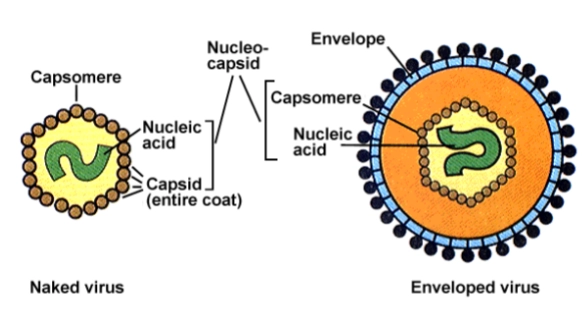
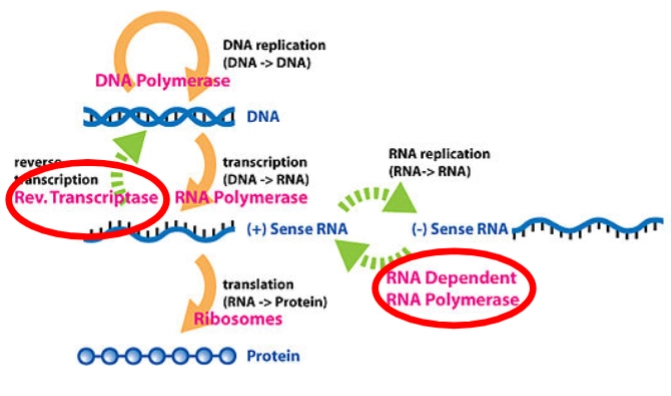
Genomisch materiaal: DNA of RNA
Als een virus DNA als genetisch materiaal heeft (zoals Herpes- of Adenovirussen), volgt het grotendeels de machines van onze eigen cel:
DNA replication (DNA → DNA): Het virus kopieert zijn eigen DNA met behulp van een DNA Polymerase (oranje pijl boven).
Transcription (DNA → RNA): Het virale DNA wordt overgeschreven naar mRNA door een RNA Polymerase (oranje pijl midden). Dit mRNA is gecodeerd als (+) Sense RNA, wat betekent dat het direct leesbaar is voor onze eiwitfabrieken.
Translation (RNA → Protein): Het ribosoom van de gastheercel (Ribosomes, oranje pijl onder) leest het mRNA af en bouwt de virale eiwitten (Protein, de blauwe kralenketting).
Sommige virussen hebben RNA als genoom, maar willen dit tijdelijk in het DNA van de gastheer bouwen (zoals HIV). Dit proces heet Reverse Transcription.
Het enzym: Rev. Transcriptase (Reverse Transcriptase): Dit is een uniek viraal enzym (gemarkeerd met de groene stippelpijl omhoog). Menselijke cellen bezitten dit enzym niet.
De actie: Het neemt het virale (+) Sense RNA en bouwt dit achterwaarts om naar viraal DNA. Dit gevormde DNA integreert vervolgens in het menselijke genoom, waarna de cel weer via de normale oranje pijlen viraal mRNA en eiwitten gaat maken.
Veel virussen hebben uitsluitend RNA bij zich (zoals het Coronavirus of Influenza) en komen in hun hele levenscyclus helemaal geen DNA tegen. Ze moeten hun RNA direct vermenigvuldigen via RNA replication (RNA → RNA).
Het probleem: Menselijke cellen kunnen alleen RNA maken op basis van een DNA-sjabloon. Onze cellen hebben geen enzymen die RNA kunnen kopiëren vanaf een RNA-streng.
Het enzym: RNA Dependent RNA Polymerase (RdRP) (De rode cirkel rechts): Het virus moet dus verplicht zelf de code meenemen voor dit specifieke enzym.
De actie (Groene stippelpijlen rechts):
Het RdRP leest het binnengekomen (+) Sense RNA af en bouwt een complementaire, omgekeerde spiegelstreng: het (-) Sense RNA.
Vervolgens gebruikt het RdRP die minstreng (-) weer als blauwdruk om duizenden nieuwe plusstrengen (+) te kopiëren.
Deze nieuwe (+) Sense RNA-strengen dienen direct als mRNA voor de Ribosomes om nieuwe virale eiwitten te maken óf worden verpakt als het genetische materiaal voor de nieuwe virusdeeltjes.
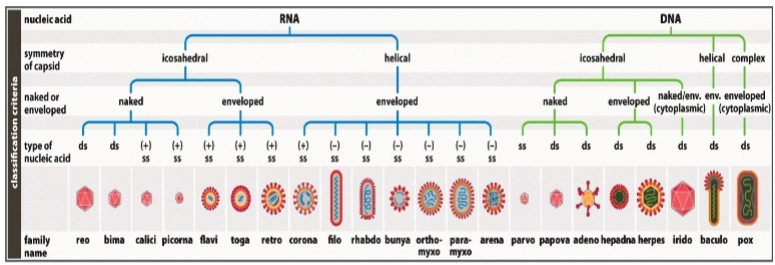
Virussen: classificatie
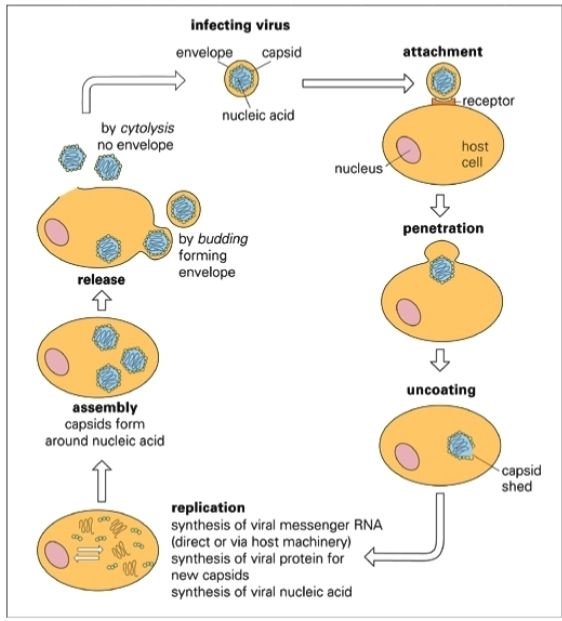
Infectiecyclus virussen
1. Attachment (Hechting)
Het virus (infecting virus), bestaande uit genetisch materiaal (nucleic acid), een eiwitjas (capsid) en een vetlaag (envelope), landt op de cel. De plakeiwitten van het virus binden heel specifiek aan een eiwitpoortje op het celmembraan: de receptor.
2. Penetration (Binnendringen)
Nadat het virus zich heeft vastgeankerd, dringt het de gastheercel binnen. Op de afbeelding zie je dat het celmembraan instulpt om het virusdeeltje in zijn geheel op te nemen in het cytoplasma.
3. Uncoating (Uitpakken)
Eenmaal binnen moet het genetische materiaal vrijkomen om afgelezen te worden. De harde eiwithand om het virus heen valt uit elkaar (capsid shed). De blauwe kronkellijn (het DNA of RNA) ligt nu los in de cel.
4. Replication (Kopiëren en Bouwen)
Dit is de actieve fabriekfase waarin de virusdeeltjes de cel kapen. Er vinden drie dingen tegelijk plaats:
Er wordt viraal mRNA gemaakt (synthesis of viral messenger RNA).
De celvloeistof bouwt op basis van dat mRNA nieuwe virale structuureiwitten (synthesis of viral protein for new capsids).
Het virale genetische materiaal wordt duizenden keren gekopieerd (synthesis of viral nucleic acid).
5. Assembly (In elkaar zetten)
Alle losse, nieuw gemaakte onderdelen komen samen. De nieuwe eiwitkapsels vouwen zich strak om de gekopieerde strengen genetisch materiaal heen (capsids form around nucleic acid). De cel zit nu vol met kant-en-klare, nieuwe virusdeeltjes.
6. Release (Vrijkomen / De uitgang)
De nieuwe virussen verlaten de cel om nieuwe cellen te infecteren. Dit kan visueel op twee manieren:
By cytolysis (Links): De cel barst volledig open en sterft. Dit gebeurt bij virussen zonder envelop (no envelope).
By budding (Rechts): De virussen duwen zichzelf door het celmembraan naar buiten en snoeren een stukje van dit membraan mee. Hierdoor krijgt het virus zijn vetlaag (forming envelope).
→ De cirkel is hiermee rond: het virus dat via budding of cytolysis vrijkomt, start bij een volgende cel weer direct bij stap 1 (attachment).
Influenzavirus: kenmerken
Orthomyxoviridae: Dit is de officiële biologische familienaam waartoe het influenzavirus behoort.
ssRNA virus: Dit betekent dat het virus een enkelstrengs (single-stranded) RNA-genoom heeft met een negatieve polariteit (minstreng). Zoals we eerder zagen bij de Baltimore-classificatie, betekent een minstreng dat het virus verplicht zijn eigen RNA-dependent RNA-polymerase moet meenemen om in onze cel eerst een plusstreng (mRNA) te maken voordat onze ribosomen het kunnen lezen.
Enveloped virus: Het is een virus met een envelop (een gestolen vetlaag). Zoals je nu weet, maakt dit het virus buiten de gastheer kwetsbaar voor zeep en alcoholgel.
Influenza A vs. Influenza B:
Influenza A: Kan zowel mensen, varkens als vogels infecteren. Omdat het tussen diersoorten kan springen, is type A berucht voor het veroorzaken van wereldwijde pandemieën (zoals de vogelgriep of de Mexicaanse griep).
Influenza B: Infecteert vrijwel uitsluitend mensen en muteert een stuk trager. Dit veroorzaakt de normale, jaarlijkse wintergriepgolven.
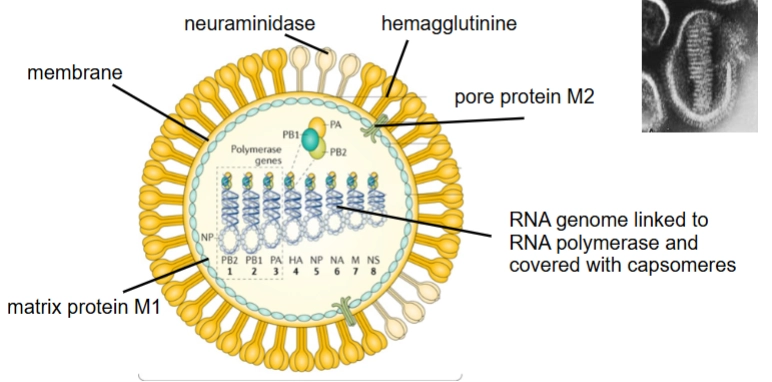
Influenzavirus: structuur
De buitenkant: De envelop en zijn spikes
In de vetlaag (membrane) zitten ongeveer 500 uitstekende plakeiwitten (spikes) die cruciaal zijn voor de infectie:Hemagglutinine (HA): De grote gele spikes. Dit is de 'sleutel' waarmee het virus zich vastankert aan de receptoren (siaalzuur) van onze slijmvliescellen om de cel binnen te dringen. Er zijn veel verschillende typen van (H1, H2, H3, etc.).
Neuraminidase (NA): De beige/witte spikes. Dit is een enzym dat de bacterie- of celreceptoren doorknipt wanneer nieuwe virusdeeltjes naar buiten willen knoppen (budding), zodat ze niet aan hun eigen cel blijven plakken. (Denk aan N1, N2, etc., wat de naamgeving van bijvoorbeeld H1N1 verklaart).
Pore protein M2: Een klein groen ionkanaaltje in het membraan. Dit kanaaltje laat zuren door zodra het virus in een endosoom zit, wat de uncoating (het uitpakken van het RNA) triggert.
Matrix protein M1: Een stevige eiwitlaag net onder het membraan die het virus zijn vorm en stevigheid geeft.
De binnenkant: Het gefragmenteerde genoom
Als je in de kern van de tekening kijkt, zie je de allergrootste bijzonderheid van influenza:8 RNA segments covered by a helical capsid: Het genoom van Influenza bestaat niet uit één lange streng, maar is opgeknipt in 8 losse RNA-fragmenten (genummerd van 1 tot 8 onderaan).Elk fragment is losjes omwikkeld met beschermende eiwitten (covered with capsomeres) tot een spiraalvorm (helical capsid).
Aan de top van elk fragment zie je drie gekleurde bolletjes (PB1, PB2, PA). Dit is het complete RNA-polymerase-complex dat het virus bij zich draagt om zijn eigen minstreng-RNA direct te kunnen kopiëren.
Influenzavirus: waarom is die structuur met 8 losse fragmenten gevaarlijk?
= als een varken of mens tegelijkertijd geïnfecteerd raakt met een vogelgriepvirus én een mensengriepvirus, kunnen deze 8 losse fragmenten tijdens de assembly (fase 5 van de levenscyclus) compleet gehusseld worden. Er rolt dan een gloednieuw virus naar buiten met bijvoorbeeld de binnenkant van een mensenvirus, maar de H-en-N-spikes van een vogelvirus. Dit biologische fenomeen noemen we Antigenic Shift en is de reden waarom ons immuunsysteem plotseling volledig blind is voor een nieuwe griepstam.
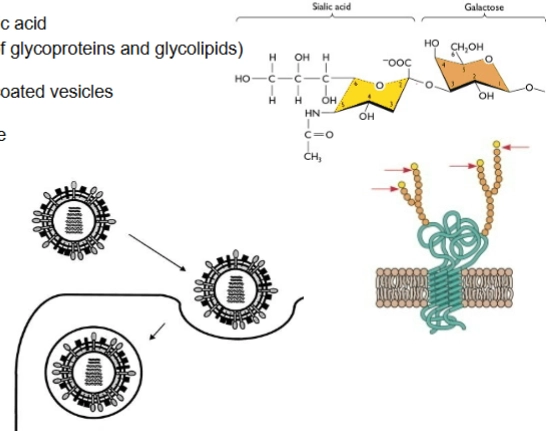
De 10 uur-durende infectiecyclus van het influenzavirus:
influenza binding and incorporation
= zien we hoe het virus de cel binnendringt:
De chemische landing (Rechtsboven): De grote gele spikes van het virus, Hemagglutinine (HA), binden aan siaalzuur (sialic acid). Siaalzuur is een specifiek suikermolecuul dat als het allerlaatste stationnetje (terminal sugar residue) vastzit aan de suikerketens van de menselijke membraaneiwitten.
Endocytosis: Zodra HA zich vastgrijpt aan het siaalzuur, activeert de cel clathrine-gemedieerde endocytose. De cel stulpt naar binnen en slikt het complete virus in, verpakt in een blaasje: het endosoom.
De 10 uur-durende infectiecyclus van het influenzavirus:
release of viral genome in host cell
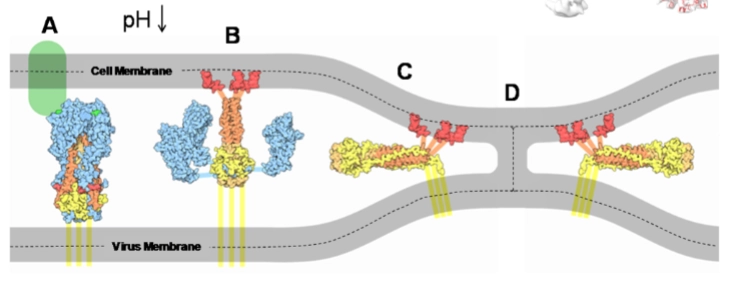
→ Eenmaal binnen in het endosoom moet het virus zijn 8 RNA-segmenten zien te lozen in het cytoplasma. Dit gebeurt via een spectaculaire pH-gestuurde ontsnapping:
Aanzuring via M2 (Dia 3): De gastheercel pompt protonen (H+) het endosoom in om de inhoud te verteren. Het virus gebruikt dit signaal juist tegen de cel: het kleine groene ionkanaaltje in de viruswand, het M2 pore protein, laat de protonen (H+) doorstromen tot diep in de kern van het virusdeeltje.
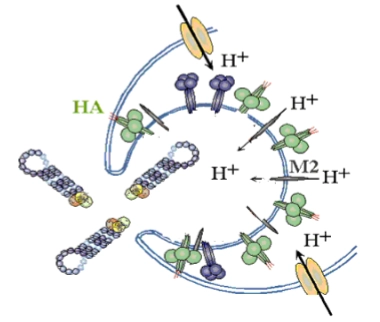
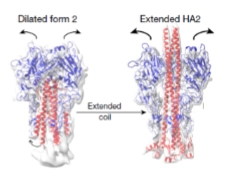
De moleculaire springveer (Dia 4): Door de aanzuring (Ph omlaag) klapt het Hemagglutinine (HA) eiwit volledig om van vorm (conformational change). Kijk naar de letters op dia 4:
A & B: HA staat in de ruststand (blauw complex). Bij de lage pH klappen de blauwe armen opzij (Dilated form 2) en schiet er een rode, verborgen spiraal naar buiten (Extended HA2).
C & D: De top van deze rode spiraal is extreem hydrofoob (vetminnend). Hij harpoeneert zichzelf rechtstreeks in het menselijke endosoom-membraan. De spiraal trekt vervolgens het virusmembraan en het celmembraan strak tegen elkaar aan tot ze volledig met elkaar versmelten (Fusion of virion with endosome).
Het resultaat: Er ontstaat een opening waardoor de 8 losse RNA-segmenten veilig het cytoplasma in glijden.
De 10 uur-durende infectiecyclus van het influenzavirus:
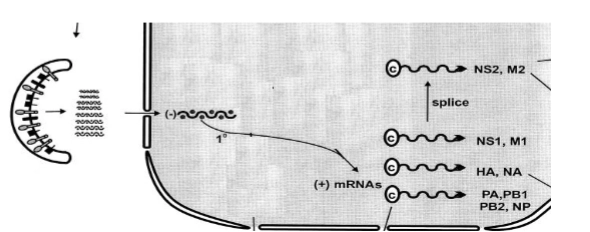
virus RNA transport and transcription
→ Nu liggen de 8 minstreng-RNA segmenten los in het cytoplasma, maar ze moeten naar de celkern om gekopieerd te worden.
De nucleaire postcode (NLS): Elk van de 8 RNA-segmenten zit vast aan het RNA-dependent RNA-polymerase complex (PA, PB1, PB2). Deze drie eiwitten dragen een ingebouwde moleculaire postcode: het Nucleus Localization Signal (NLS).
De reis naar binnen: De gastheercel herkent deze NLS-code, denkt dat het om eigen eiwitten gaat, en transporteert het virale RNA via de kernporiën actief de celkern in (Host transport system provides transport to the nucleus).
Transcriptie naar mRNA: Eenmaal in de kern gaat het meegeleverde polymerase direct aan het werk. Het leest het (-) sense vRNA af en bouwt de complementaire (+) mRNAs. Sommige van deze mRNA's worden daarna nog korter geknipt (splice) om coderend te worden voor specifieke eiwitten (zoals NS2 en M2).
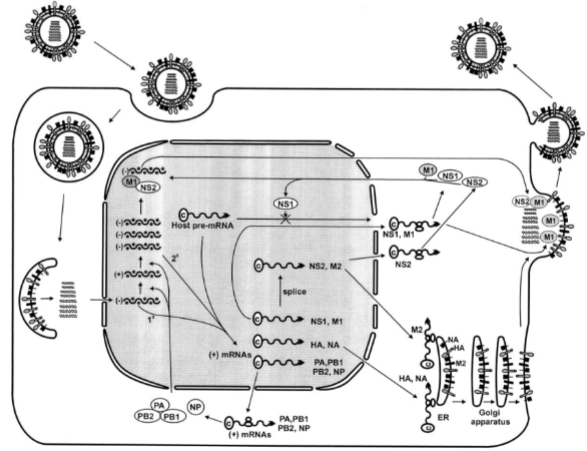
De 10 uur-durende infectiecyclus van het influenzavirus:
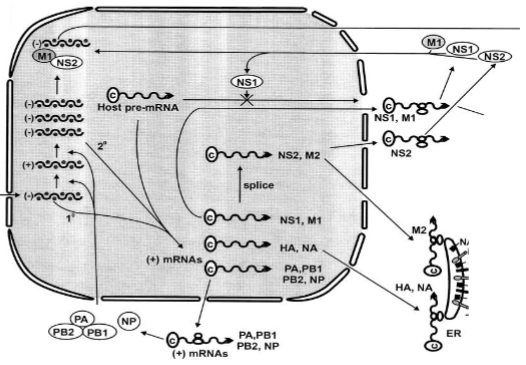
Translational viral proteins and production vRNA
→ In deze fase draait de celkern op volle toeren om twee verschillende stromen te produceren:
- Eiwitproductie: De nieuw gemaakte (+) mRNAs verlaten de kern via de poriën. In het cytoplasma haken ze aan bij de ribosomen om virale eiwitten te maken (zoals de polymerase-subunits PA, PB1, PB2 en het nucleoproteïne NP). Deze eiwitten reizen direct weer terug de kern in om te helpen.
- Genoomkopiëring: Het RNA-dependent RNA-polymerase switcht van stand. Het gaat nu de binnengekomen minstrengen (-) kopiëren naar een volledige plusstreng-bladdruk (+), om die vervolgens weer massaal te kopiëren naar duizenden gloednieuwe minstrengen (-) sense vRNA voor de nakomelingen.
- De diefstal (Host pre-mRNA): Influenza doet in de kern ook aan cap-snatching. Het knipt de kopjes van het menselijke pre-mRNA af (zie het kruis bij Host pre-mRNA) om te gebruiken voor zijn eigen mRNA's, waardoor de menselijke cel zelf acuut stopt met functioneren.
De 10 uur-durende infectiecyclus van het influenzavirus:
Transport viral membrane proteins to cell surface
→ De eiwitten die bestemd zijn voor de buitenkant van het nieuwe virus moeten correct worden gevouwen en getransporteerd:
De route via de eiwitfabriek: De membraaneiwitten Hemagglutinine (HA), Neuraminidase (NA) en het M2-kanaal worden door ribosomen direct in het ER (Endoplasmatisch Reticulum) gebouwd.
De post-trein: Vanuit het ER reizen deze eiwitten via transportblaasjes door het Golgi-apparaat (zie de getekende stap-pijlen onderaan). Tijdens deze reis worden er suikergroepen aan de eiwitten geplakt.
Lipid rafts: Uiteindelijk fuseren de blaasjes met het buitenste celmembraan. De eiwitten HA, NA en M2 clusteren heel gezellig samen op specifieke vette plekken op het celoppervlak: de lipid rafts.
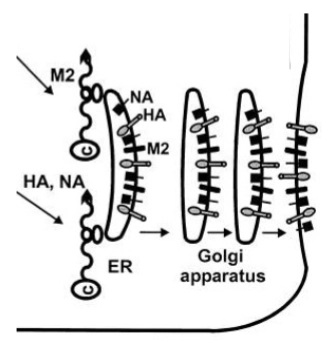
De 10 uur-durende infectiecyclus van het influenzavirus:
transport vRNA to cell surface
→ De vers gemaakte RNA-minstrengen liggen nog in de celkern en moeten naar het celmembraan.
Het nucleocapside-complex: De RNA-strengen (vRNA) wikkelen zich in de kern om eiwitten (NP - nucleoproteïne).
De nucleaire export-machine: Om de kernporie (Nuclear pore) te passeren, bindt het matrixeiwit M1 aan het vRNA. M1 rekruteert vervolgens NS2 (ook wel NEP - Nuclear Export Protein genoemd).
De poortwachter: NEP functioneert als de pasvorm die herkend wordt door het menselijke export-eiwit Exportin en de energieleverancier RanGTP. Samen trekken ze het hele virale RNA-pakket de kern uit richting het cytoplasma.
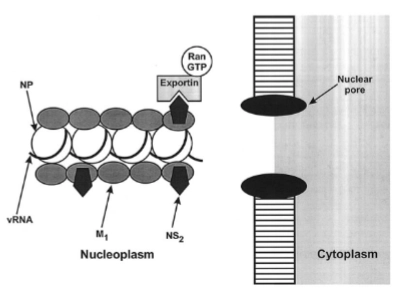
De 10 uur-durende infectiecyclus van het influenzavirus:
production of virions
→ Nu alle onderdelen bij de vette plekken van het celmembraan (lipid rafts) zijn aangekomen, start de montage.
M1 als de steiger (Dia 2): Het matrixeiwit M1 vormt grote ketens (higher order oligomers, zie de spiraalstructuur links). M1 functioneert als een dubbelzijdige brug: het bindt aan de onderkant aan het vRNA en aan de bovenkant aan de staarten van de membraanspikes (HA en NA).
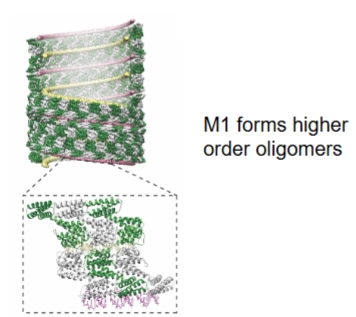
Budding (Dia 2 rechts): Door deze clustering stulpt het celmembraan naar buiten. De lipiden (vetten) van het virusmembraan worden zo rechtstreeks gestolen van de gastheercel.
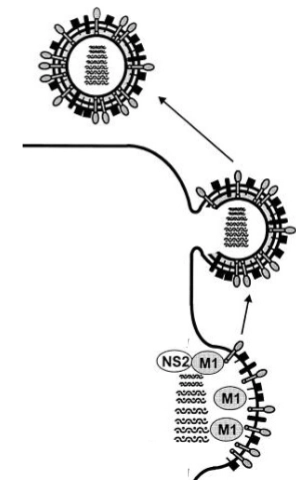
De 10 uur-durende infectiecyclus van het influenzavirus:
specific incorporation of vRNA segments
De 8 segmenten sorteren (Dia 3)
Het selectieprobleem: Een influenzavirus is pas functioneel als hij van álle 8 unieke RNA-fragmenten exact één kopie heeft.
Dual targeting: Elk segment heeft een universele regio (conserved region) én een unieke code (specific sequence). Hierdoor herkennen de 8 fragmenten elkaar, haken ze in een vaste cirkel van 1+7 in elkaar (zie de elektronenmicroscoopfoto en het gekleurde diagram), en worden ze als één complete set van 8 ingepakt.
Celsterfte: De gastheercel produceert een tijdlang non-stop virussen totdat de cel volledig uitgeput raakt en sterft (necrosis).
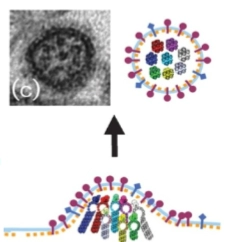
De 10 uur-durende infectiecyclus van het influenzavirus:
release of virions
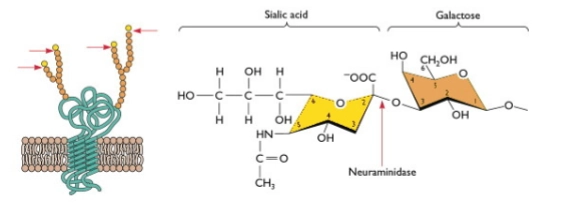
→ Wanneer het nieuwe virus naar buiten knopt, stuit het op een groot probleem: zijn eigen grijparmen (Hemagglutinine) plakken direct weer vast aan het siaalzuur op het membraan van de cel waar hij net uitkomt (virions cluster and bind to original producing cell). Het virus hangt vast.
De schaar (Neuraminidase): Hier komt de tweede grote spike in actie: Neuraminidase (NA).
De knip: NA functioneert als een biochemische schaar. Het knipt het siaalzuur (N-acetylneuraminic acid) koelbloedig weg van de menselijke membraaneiwitten (zie de rode pijl bij de chemische structuur).
Vrijheid: De binding wordt verbroken. Het virusdeeltje is definitief vrij in de omgeving (virion free in environment) en kan op weg naar de volgende gezonde cel in de luchtwegen om de infectie te verspreiden
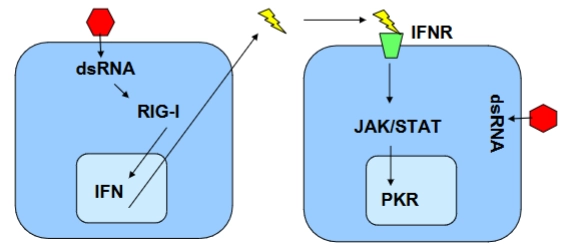
Influenza vs. het immuunsysteem
→ hoe een gezonde cel een naburige cel waarschuwt voor een virusinfectie
Cel 1 (Links - De geïnfecteerde cel): Wanneer Influenza zijn RNA kopieert, ontstaat er per ongeluk dubbelstrengs RNA (dsRNA, de rode zeshoek). Menselijke cellen hebben dit nooit los in het cytoplasma. De sensor RIG-I ontdekt dit dsRNA direct en activeert de productie van IFN (Type I Interferon). De cel loost dit interferon (de bliksemschicht) naar buiten.
Cel 2 (Rechts - De gezonde buurcel): Het interferon landt op de receptor (IFNR, het groene trechtertje) van de buurcel. Dit activeert de JAK/STAT-signaalroute.
De verdediging (PKR): JAK/STAT zet de productie aan van het antivirale eiwit PKR (Protein Kinase R). Zodra PKR geactiveerd wordt door binnendringend viraal dsRNA, legt het de complete eiwitproductie van de cel plat. De cel pleegt hiermee deels zelfmoord, waardoor het virus zich niet verder kan kopiëren.
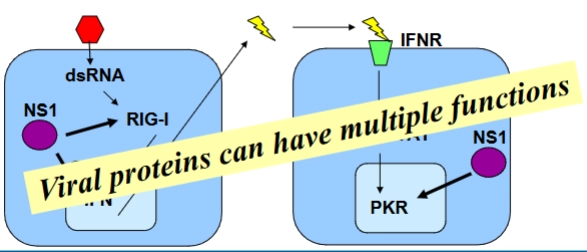
Influenza vs. het immuunsysteem
→ aanval van het virus
= het multifunctionele eiwit NS1 (de paarse cirkels) om deze verdedigingslinie op drie niveaus tegelijk te slopen:
Blokkade van de sensor: NS1 blokkeert direct de activatie van RIG-I (paarse cirkel links). De cel merkt het dsRNA niet meer op, waardoor de interferon-kraan (het alarm) dicht blijft.
Kaping in de kern: NS1 bindt aan de poly-adenylatiemachine in de celkern. Hierdoor kan de menselijke cel geen eigen mRNA's meer afmaken, wat de productie van Type I Interferonen extra hard platlegt.
Blokkade van het wapen: Mocht de buurcel via omwegen tóch PKR hebben aangemaakt, dan reist NS1 mee naar de buurcel (paarse cirkel rechts). NS1 bindt fysiek aan PKR en blokkeert de functie ervan. De cel kan zijn eiwitproductie niet meer stilzetten en het virus blijft zich ongestoord vermenigvuldigen.
Antigenic drift vs. antigenic shift
Antigenic drift (Puntmutaties): Dit proces draait om kleine, spontane foutjes van het virale kopieerenzym. De slide vermeldt de statistiek: RNA polymerase mistakes: 1 in 10^4. Omdat het viraal RNA-polymerase geen ingebouwde foutcorrectie (proofreading) heeft, ontstaat er bij zowat elke gekopieerde virusstreng wel één willekeurige letterstoring. Dit leidt tot langzame, subtiele veranderingen in de H- en N-spikes.
→ virus krijgt een andere kleur jas, waardoor je immuunsysteem hem moeilijker herkent (oorzaak van de jaarlijkse seizoensgriep).
Antigenic shift (Genoom-husseling): Dit is de grote, plotselinge sprong waarbij complete vRNA-segmenten worden uitgewisseld.
→ een plotselinge, gigantische verandering. Dit gebeurt wanneer twee verschillende stammen van een virus tegelijkertijd dezelfde gastheercel infecteren en hun genetisch materiaal volledig door elkaar husselen (reassortment)
→ antigenic shifts gedurende de geschiedenis
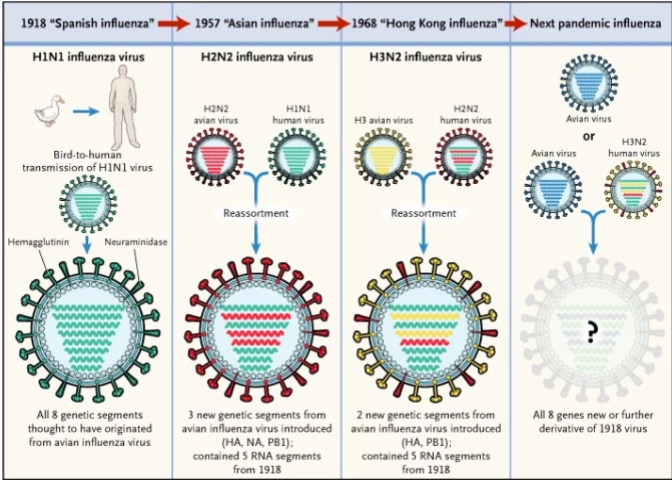
Waarom is influenza zo veranderlijk?
= het heeft een enorm vogel- en dierenreservoir.
Native Host (Het centrum): Wilde watervogels en eenden (wild water fowl, duck, etc.) zijn de natuurlijke gastheer. Zij dragen alle bekende varianten van de spikes bij zich: H1 t/m H15 en N1 t/m N9.
De oversteek naar de mens: Vanaf de watervogels kunnen virussen muteren en overspringen naar kippen (chicken), varkens (swine), paarden (equine), zeehonden (seal) en uiteindelijk de mens (Human).
Host range restriction: Normaal gesproken kan een puur vogelvirus niet zomaar direct een mens infecteren (de barrière). Maar zoals de rode pijlen laten zien, kan een virus via kippen of kwartels (quail) muteren en alsnog de overstap naar de mens maken (zoals H5N1 of H7N7).
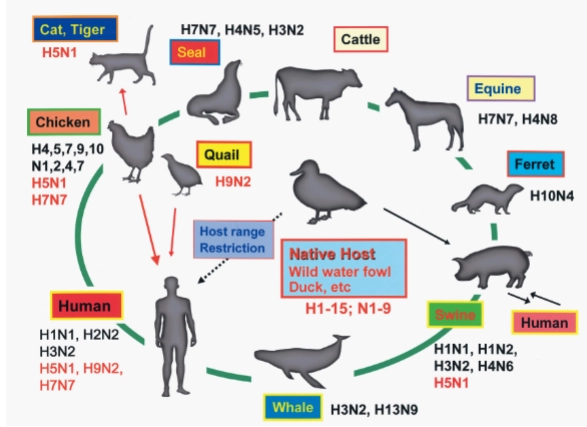
Wat is het voordeel van reassortment voor het virus (influenza)?
Ontsnappen aan de afweer (Immune evasion): Ons immuunsysteem maakt antistoffen die heel specifiek de buitenkant van de HA- en NA-spikes herkennen.
Door Antigenic Drift: Veranderen de spikes elke winter een heel klein beetje. Je oude antistoffen passen net niet goed meer op de nieuwe variant. Je wordt weer een beetje verkouden of krijgt een milde griep. Dit dwingt ons om elk jaar een nieuwe griepprik te halen.
Door Antigenic Shift: Krijgt het virus in één klap een compleet nieuwe 'jas' (nieuwe HA of NA). Niemand op de wereld heeft hier antistoffen tegen. Het virus kan zich daardoor razendsnel verspreiden via een wereldwijde pandemie omdat de complete populatie weer vatbaar is.
Coronavirussen
→ zijn er al heel lang
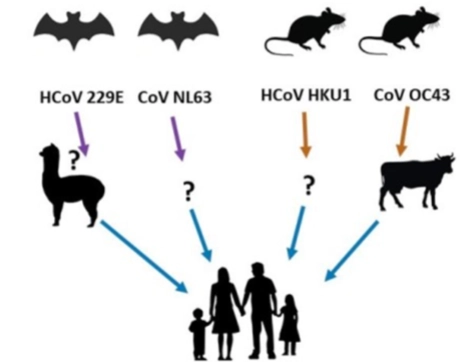
Er zijn 4 milde coronavirussen die de gewone winterverkoudheid (common cold symptoms) veroorzaken. Ze stammen oorspronkelijk allemaal af van wilde dieren:
HCoV-229E & CoV-NL63: Stammen oorspronkelijk af van vleermuizen (bats). 229E is via de alpaca/kameelachtige overgesprongen op de mens. Van NL63 is de tussenstap nog onbekend (?).
HCoV-HKU1 & CoV-OC43: Stammen oorspronkelijk af van knaagdieren/ratten (rodents). OC43 is via rundvee (cow) overgesprongen op de mens.
→ uiteindelijk na 2000, 3 variaties die wel echt schadelijke symptomen veroorzaakten:
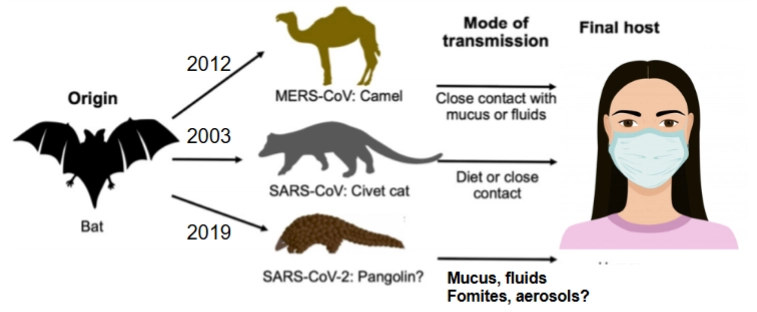
Coronavirussen: de 3 variaties die wel echt schadelijke symptomen veroorzaakten
→ Visueel zie je dat ze alle drie hun absolute oorsprong (Origin) in de vleermuis hebben, maar via een ander tussenwijd dier zijn overgestapt:
SARS-CoV (2003): Sprong van de vleermuis naar de civetkat (civet cat), en via dieet of nauw contact naar de mens.
MERS-CoV (2012): Sprong van de vleermuis naar de dromedaris/kameel (camel), en via nauw contact met slijm of vloeistoffen naar de mens.
SARS-CoV-2 (2019): De veroorzaker van de COVID-19 pandemie. Sprong vermoedelijk over via het schubdier (Pangolin?) en verspreidde zich razendsnel via ademhalingsdruppels, aerosolen en besmette oppervlakken (fomites).
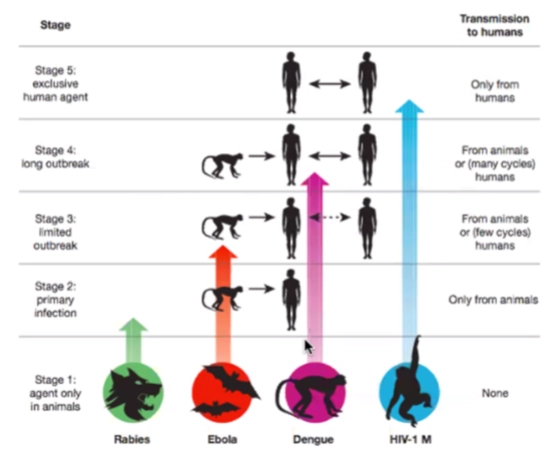
De 5 stadia van virusadaptatie
Stage 1 (Agent only in animals): Het virus zit uitsluitend in dieren (geen overdracht naar de mens).
Stage 2 (Primary infection): Het virus kan overspringen van dier naar mens, maar verspreidt zich niet tussen mensen onderling.
Voorbeeld: Hondsdolheid (Rabies); je krijgt het alleen via een beet van een dier.
Stage 3 (Limited outbreak): Het virus kan van dier naar mens, en er ontstaan een paar korte, lokale infectiecirkels tussen mensen, maar de uitbraak dooft snel uit (few cycles).
Voorbeeld: Ebola.
Stage 4 (Long outbreak): Het virus springt over van dier naar mens en kan daarna langdurige, grote epidemieën veroorzaken via mens-op-mens transmissie (many cycles).
Voorbeeld: Dengue (Knokkelkoorts) en de zware coronavirussen (SARS/MERS) ten tijde van hun uitbraak.
Stage 5 (Exclusive human agent): Het virus heeft zich genetisch volledig aangepast aan de mens. Het heeft het dierenrijk niet meer nodig om te overleven en wordt alleen nog maar van mens op mens overgedragen.
Voorbeeld: HIV-1 M (ooit begonnen bij chimpansees, nu puur menselijk). De 4 milde verkoudheidscoronavirussen uit dia 1 bevinden zich inmiddels ook in dit eindstadium.
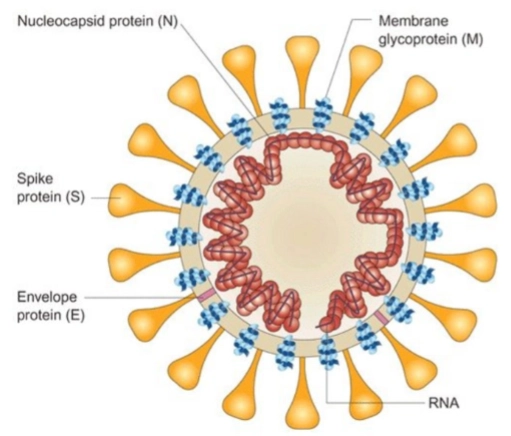
Coronavirussen: anatomie van het virusdeeltje SARS CoV2 virion
S: Spike protein (Het grote oranje 'knots'-eiwit): Dit eiwit steekt als een kroon (corona) naar buiten. Het is verantwoordelijk voor de binding aan de gastheer-receptor (ACE2) en zorgt ervoor dat het virusmembraan versmelt met het celmembraan (receptor binding and virion fusion).
N: Nucleocapsid protein (De rode spiraal binnenin): Dit eiwit functioneert als een beschermend omhulsel (capsomere) dat zich strak om het kwetsbare virale RNA-genoom heen wikkelt.
M: Membrane glycoprotein (De blauwe eiwitblokjes in de wand): Dit is het meest voorkomende structurele eiwit in de viruswand. Het geeft het virus zijn stevigheid en vorm, en regisseert de assembly in de cel (required for virion production).
E: Envelope protein (Het roze streepje in de wand): Een heel klein, functioneel envelop-eiwit (small envelope protein). Het helpt bij het in elkaar zetten van het virus en functioneert als een ionkanaal tijdens het infectieproces.
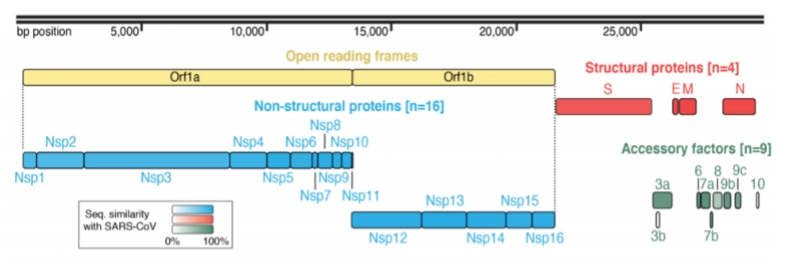
SARS-CoV2: het genoom
→ bezit een +ssRNA virus genoom (enkelstrengs plus-RNA)
Grootte: Het genoom is maar liefst 29,9 kilobasen (kb) lang en codeert voor 25 verschillende eiwitten. Daarmee behoort het tot de absolute reuzen onder de RNA-virussen (among the largest RNA viruses).
De verdeling (Open reading frames): * Orf1a / Orf1b (Geel - De eerste 20.000 letters): Dit gigantische stuk codeert voor de 16 niet-structurele eiwitten (Nsp1 t/m Nsp16, blauw). Dit zijn de functionele gereedschappen van het virus, zoals het kopieerenzym (Nsp12 - RdRP) en het enzym dat RNA-fouten corrigeert (Nsp14).
Structural proteins (Rood - De laatste 10.000 letters): Hier liggen de blauwdrukken voor de 4 eiwitten van dia 1 en 2 (S, E, M, N).
Accessory factors (Groen): Codeert voor 9 extra hulpfactoren die het immuunsysteem van de gastheer saboteren.
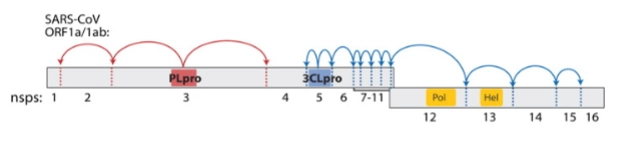
→ Omdat SARS-CoV2 zo groot is, heeft het een slimme logistieke truc om efficiënt met zijn genetische ruimte om te gaan (higher efficiency with genomic information)
= Wanneer onze menselijke ribosomen het grote gele stuk (Orf1a/1b) aflezen, maken ze niet direct 16 losse eiwitten. In plaats daarvan rollen er twee gigantische, aan elkaar geplakte ketens uit: de polyproteïnes. Om hier losse, werkende machines van te maken, bezit het virus 2 ingebouwde scharen (proteases) die zichzelf en de rest van de keten lossnijden:
PLpro (Papain-like protease - Het rode blokje op Nsp3): Knipt de keten door op de plekken van de rode pijlen (tussen Nsp1, 2, 3 en 4).
3CLpro (3Chymotrypsin-like protease / Main protease - Het blauwe blokje op Nsp5): Knipt de rest van de keten los op de plekken van de blauwe pijltjes (tussen Nsp4 t/m Nsp16). Hierdoor komen onder andere het polymerase-kopieerenzym (Pol / Nsp12) en het helicase-eiwit (Hel / Nsp13) vrij om hun werk te gaan doen.
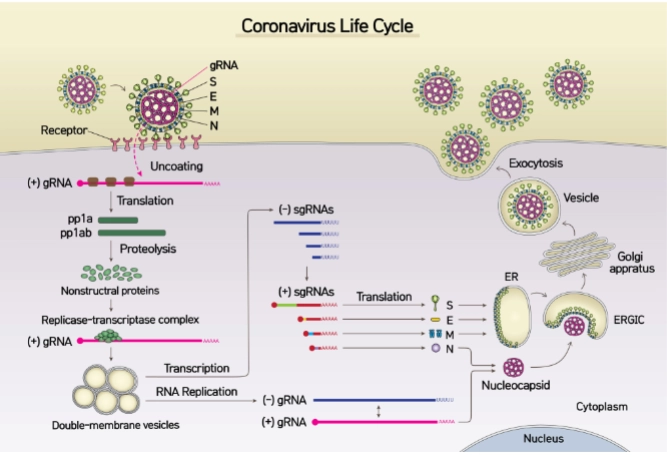
De levenscyclus van het Coronavirus
→ Je ziet het virus links binden aan de receptor. Het vRNA komt vrij, wordt direct vertaald naar vroege eiwitten, en start replicatie in speciale blaasjes. De nieuwe virusonderdelen assembleren via het ER/Golgi-netwerk en verlaten de cel rechts via transportblaasjes (exocytosis).
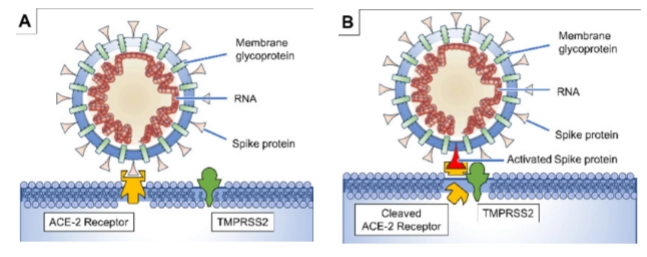
Verschillen in de levenscyclus tussen Corona en Influenza:
specifieke receptor & uncoating bij het celmembraan
De binding (A): Het coronavirus gebruikt zijn Spike protein om te binden aan de ACE-2 receptor op de gastheercel (zoals long- of darmepitheelcellen).
De activatie (B): Een menselijk knipenzym op het celmembraan, TMPRSS2, knipt een stukje van de ACE-2/Spike-verbinding af (Cleaved ACE-2 Receptor).
Het gevolg: Dit activeert het spike-eiwit, waardoor het virusmembraan direct versmelt met het celmembraan van de menselijke cel (uncoating at cell membrane). Het RNA glijdt direct de cel in zonder dat er per se een endosoom nodig is.
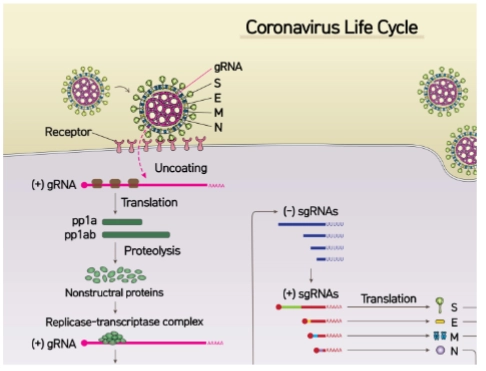
Verschillen in de levenscyclus tussen Corona en Influenza:
vRNA (+ssRNA) is direct vertaalbaar
Direct aan de slag: Omdat het genoom uit (+)ssRNA bestaat, herkent het menselijke ribosoom dit direct als een bruikbaar mRNA.
De eerste eiwitten: De cel gaat het binnengekomen RNA meteen vertalen (Translation) naar de eiwitketens pp1a en pp1ab.
Geen polymerase in het virion: Het virus hoeft dus—in tegenstelling tot Influenza—geen kant-en-klaar kopieerenzym (RdRp) mee te smokkelen binnenin het virusdeeltje. Het virus laat de gastheercel dit enzym simpelweg na binnenkomst direct zelf maken.
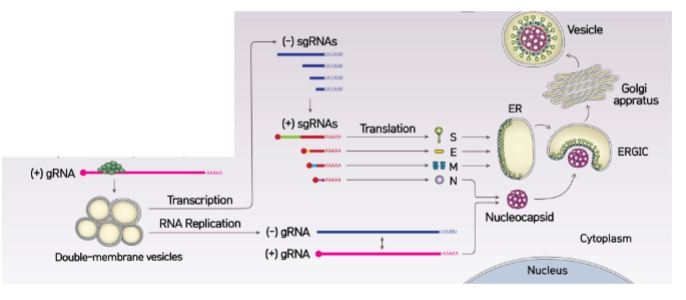
Verschillen in de levenscyclus tussen Corona en Influenza:
Intracellular viral replication compartment
→ Wanneer het virus zijn RNA gaat kopiëren, bouwt het via gemodificeerde ER-membranen speciale kamers: Double-membrane vesicles (DMVs) (getekend als de gele bollen op dia 4).
Volgens dia 5 heeft dit twee vitale redenen:
Concentratie: Het verhoogt lokaal de concentratie van alle bouwstoffen die nodig zijn voor de replicatie.
Camouflage (Cruciaal): Het houdt het gevormde dubbelstrengs RNA (dsRNA) fysiek verborgen voor de menselijke immuunsensoren (zoals RIG-I uit je eerdere slides), waardoor er geen interferon-alarmsignaal afgaat.
Verschillen in de levenscyclus tussen Corona en Influenza:
proofreading by RNA-dependent RNA polymerase
RNA-virussen muteren normaal gesproken gigantisch snel omdat hun kopieerenzym veel fouten maakt. Coronavirussen zijn echter zo reusachtig groot dat ze stabiel moeten blijven. Daarom hebben ze een unieke foutcorrectie (proofreading):
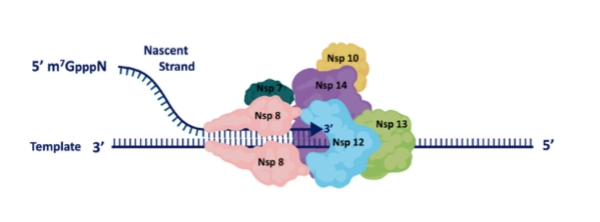
Nsp7-8-12: Vormen samen het centrale kopieercomplex (RdRp complex).
Nsp13: Windt het dubbelstrengs RNA uit elkaar (dsRNA unwinding).
Nsp14 (ExoN): Dit is de unieke kwaliteitscontroleur. Als het RdRp-complex per ongeluk een verkeerde letter inbouwt, reist Nsp14 erachteraan, knipt de fout eruit (excision of misincorporated nucleotides) en herstelt de streng. Hierdoor muteert corona een stuk trager dan influenza.
Verschillen in de levenscyclus tussen Corona en Influenza:
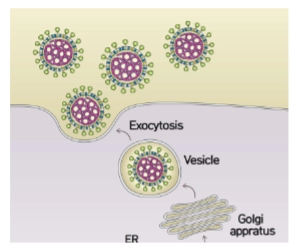
Corona virions are exocytosed from the ER/Golgi
→ Dit is een fundamenteel verschil in waar de vetlaag (lipiden) vandaan komt:
Influenza: Knoopt naar buiten via het buitenste celmembraan en steelt vetten uit de lipid rafts. De vetwand bestaat daarom vooral uit phosphatidylethanolamine (PE).
Coronavirus: Knoopt al diep in de cel naar buiten, in het ER/Golgi-netwerk (ERGIC). Het virus wordt verpakt in een menselijk transportblaasje (Vesicle) en verlaat de cel via exocytose. De vetwand van corona bevat hierdoor typische ER/Golgi-vetten: veel sphingomyeline (SM), phosphatidylinositol (PI) en phosphatidylserine (PS).
De gouden regels voor het ontwerpen van virusremmers (antiviraalmiddelen)
→ legt uit waar een goed medicijn aan moet voldoen, waarom 'breed-spectrum' virusremmers bijna niet bestaan, en welke specifieke virale eiwitten we momenteel platleggen met medicatie.
Criteria:
Target present in pathogen, absent in host (Selectieve toxiciteit):
Dit is de belangrijkste regel in de toxicologie en geneeskunde. Een medicijn mag alleen een structuur aanvallen die uniek is voor het virus, en die niet voorkomt in onze eigen menselijke cellen (absent in host). Omdat virussen onze eigen celmachines kapen om te overleven, is dit heel lastig. Als je een menselijk enzym remt, word je zelf doodziek.Viruses much more unique than bacteria → no equally ‘broad-spectrum’ drugs:
Bij bacteriën kun je makkelijk één type antibioticum (zoals penicilline) geven dat honderden verschillende bacteriesoorten tegelijk doodt (broad-spectrum), omdat ze biologisch erg op elkaar lijken. Virussen zijn onderling echter extreem verschillend. Een coronavirus werkt moleculair totaal anders dan HIV of het griepvirus. Daarom werkt een virusremmer bijna altijd maar tegen één specifieke virusfamilie; een universele "virus-killer" bestaat nagenoeg niet.
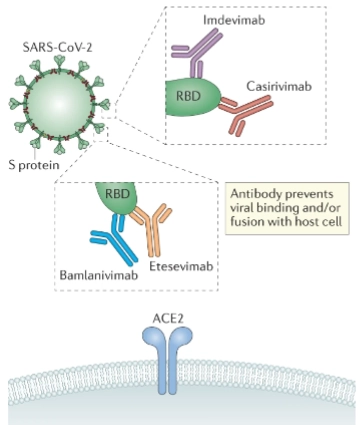
Monoclonal antibodies (& werking)
= In plaats van de interne enzymen (zoals polymerases of proteases) te remmen, kun je het virus ook aan de buitenkant blokkeren met kunstmatige, in het lab gemaakte antistoffen (recombinant monoclonal Human antibodies).
Werking:
Het mechanisme: Kijk naar de getekende uitvergrote vakjes. De antistoffen (zoals Imdevimab, Casirivimab, Bamlanivimab en Etesevimab) richten zich allemaal specifiek op het groene bolletje: de RBD (Receptor Binding Domain) van het spike-eiwit.
De blokkade: Doordat de antistof als een kapje over de RBD heen klikt, kan het virus fysiek niet meer binden aan de blauwe menselijke ACE2-receptor (antibody prevents viral binding and/or fusion with host cell).
De cocktail-strategie: Waarom zie je steeds twee verschillende antistoffen per vakje? RNA-virussen muteren snel (antigenic drift). Als het virus één letter verandert in zijn RBD, past antistof A (bijv. Bamlanivimab) misschien niet meer. Maar omdat antistof B (Etesevimab) net op een ander hoekje (different binding motif) grijpt, blijft het virus alsnog gevangen. Dit voorkomt dat het virus ontsnapt via een enkele mutatie (prevent early escape by single mutation).
Herpesviridae (herpesvirus → alpha, beta, gamma)
→ is (in tegenstelling tot influenza en corona) een DNA virus
→ De familie is opgedeeld in drie subfamilies (alpha, beta, gamma) en kent 8 varianten die de mens infecteren:
alpha (Alfa-herpesvirussen): Groeien snel en trekken zich latent (in slaapstand) terug in zenuwknopen.
HSV-1: Veroorzaakt de klassieke koortslip (herpes facialis/labialis, zie de foto rechtsboven).
HSV-2: Veroorzaakt herpes aan de geslachtsorganen (herpes genitalis).
VZV: Veroorzaakt waterpokken (chicken pox) bij kinderen en kan later als gordelroos (shingles) terugkeren.
beta (Bèta-herpesvirussen): Groeien traag en slapen in immuuncellen of organen.
CMV: Veroorzaakt milde pfeiffer-achtige klachten, maar is gevaarlijk tijdens de zwangerschap vanwege het risico op aangeboren afwijkingen (foetale misvormingen).
HHV-6 & HHV-7: Veroorzaken de zesde ziekte (roseola infantum) bij jonge kinderen.
gamma (Gamma-herpesvirussen): Infectieren lymfecellen en kunnen cellen aanzetten tot ongecontroleerde deling (oncogeen).
EBV: Veroorzaaker van de ziekte van Pfeiffer (mononucleosis) en geassocieerd met bepaalde kankervormen (Burkitt-lymphoma).
HHV-8: Veroorzaakt het Kaposi's sarcoma (een bloedvattumor, veel gezien bij aids-patiënten).
Herpesviridae: genoom
dsDNA virus (125-250 kb): Het genoom bestaat uit dubbelstrengs DNA (double-stranded). Vergelijk dit eens met SARS-CoV-2 (30 kb) of Influenza (13,5 kb); het herpesgenoom is tot wel acht keer groter.
More than 80 genes: Het virus codeert voor een gigantische hoeveelheid (meer dan 80) eigen eiwitten.
In vitro vs. In vivo: Ongeveer de helft van die genen is strikt noodzakelijk om simpelweg in een laboratorium-kweekschaaltje te kunnen kopiëren (half required for in vitro replicatie). Maar om in een echt levend organisme ziekte te kunnen veroorzaken, heeft het virus ze állemaal nodig (all essential for in vivo virulence).
Genetische diefstal (vIL-10, MHC): Omdat het genoom zo groot is, heeft het virus tijdens de evolutie complete genen gestolen van de gastheer (originally from host). Het heeft bijvoorbeeld een eigen variant van het menselijke remmende cytokine IL-10 (genaamd vIL-10) en MHC-achtige structuren ingebouwd om ons immuunsysteem doelbewust te misleiden en te onderdrukken.
Varicella Zoster Virus (VZV):
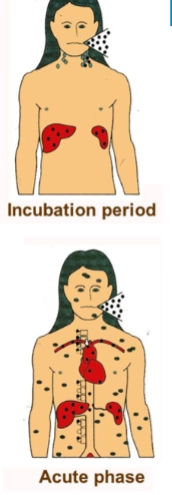
→ primaire infectie (waterpokken)
Infection of airways: Je ademt het virus in via druppeltjes in de lucht. Het virus infecteert als eerste de slijmvliezen van de bovenste luchtwegen.
Viral replication in local lymph node: Vanaf de luchtwegen reist het virus naar de lokale lymfeklieren in de hals. Hier vermeerdert het virus zich stilletjes.
Primary viremia: Het virus barst uit de lymfeklieren en komt in de bloedbaan terecht (viremie). Via het bloed reist het direct naar dieper gelegen organen: de lever en de milt (liver, spleen, te zien aan de rode organen in het bovenste poppetje).
Secondary viremia: In de lever en milt vermeerdert het virus zich opnieuw massaal. Het kaapt specifieke immuuncellen (T-lymfocyten / mononucleaire cellen) om zich via een tweede, veel grotere viremie golf door het hele lichaam te verspreiden. Deze T-cellen brengen het virus specifiek naar de huid en slijmvliezen (transport to skin and mucosa).
Local replication and shedding: Eenmaal in de huid nestelt het virus zich in de cellen. Dit leidt tot lokale ontsteking, blaasjes en intense jeuk over het hele lichaam, zoals te zien op de foto van de baby. Uit deze blaasjes wordt het virus ook weer uitgescheiden (shedding) naar de omgeving.
Varicella Zoster Virus (VZV):
→ heractivatie (gordelroos)
→ Nadat je bent genezen van de waterpokken, verdwijnt het herpesvirus nooit meer uit je lichaam. Het reist via de zenuwbanen omhoog en trekt zich levenslang terug in een slaapstand (latentie) in de sensibele zenuwknopen vlak bij het ruggenmerg.
Wanneer je afweer op latere leeftijd daalt, kan het virus ontwaken (Reactivation). De diagrammen laten precies zien waarom gordelroos er zo specifiek uitziet:
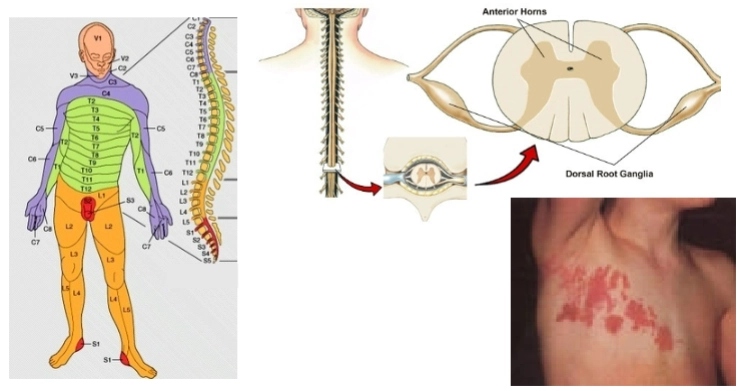
De anatomische slaapplek (Midden & Rechtsboven): Op de dwarsdoorsnede van het ruggenmerg zie je de Dorsal Root Ganglia (Dorsale wortelganglia) aangeduid. Dit zijn de zenuwknopen aan de rugzijde waar het virus al die jaren heeft geslapen.
Het dermatoom (Links): Elk dorsaal ganglion is verantwoordelijk voor het gevoel in één specifieke strook huid op je lichaam. Zo'n huidstrook heet een dermatoom (op de linkerkaart aangegeven met de letters en nummers zoals T1 t/m T12 voor de borstkas).
De heractivatie (Rechtsonder): Als het virus in één specifieke zenuwknoop (bijvoorbeeld T5) wakker wordt, reist het via die ene zenuwbaan weer naar beneden, terug naar de huid.
Het klinische beeld: Hierdoor ontstaan de pijnlijke blaasjes en rode uitslag strikt binnen de grenzen van dat ene dermatoom (skin area innervated by the dorsal root of a single spinal cord nerve). Zoals je op de foto rechtsonder ziet, stopt de uitslag heel karakteristiek exact in het midden van de borstkas, omdat de zenuw van de linker- of rechterkant daar ophoudt.