DNA structure and replication
1/31
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
32 Terms
Role of DNA
life depends on the ability of cells to store, retrieve and translate the instructions required to make and maintain a living organism
hereditary - passed on from a cell to its daughter cells at somatic cell division, and from one generation of an organism to the next through the organism’s gametes
stored within every living cell as its genes, information-containing elements that determine the characteristics of a species as a whole and of the individuals within it
Characteristic properties:
high capacity for information storage and be chemically stable to be able to encode information without fail
replicate accurately
be capable of variation (different traits)
DNA as hereditary material
chromosome analysis
metabolic stability of DNA
constancy of DNA within a cell
correlation btw mutagens and their effects on DNA
*Griffth’s Experiment
*Avery’s Experiments
*Hershey-Chase Experiment
Structure of DNA
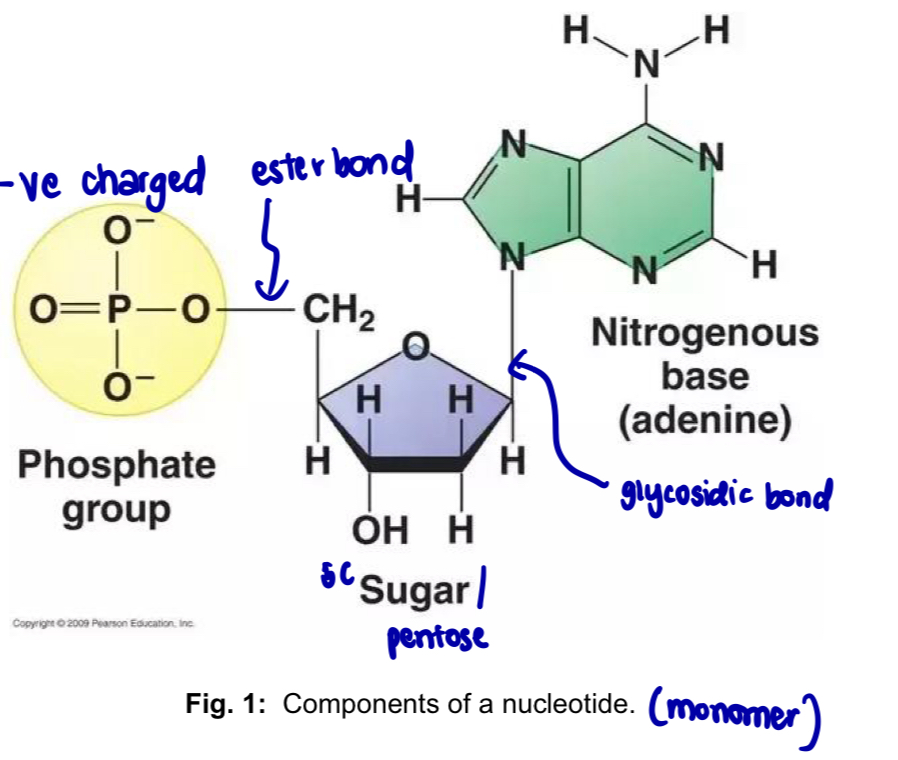
Nucleotides - building blocks of nucleic acids
Nucleic acids are macromolecules that exist as polymers called polynucleotides
each nucleotide is composed of
a. five-carbon sugar - pentose
b. nitrogenous base
c. phosphate group
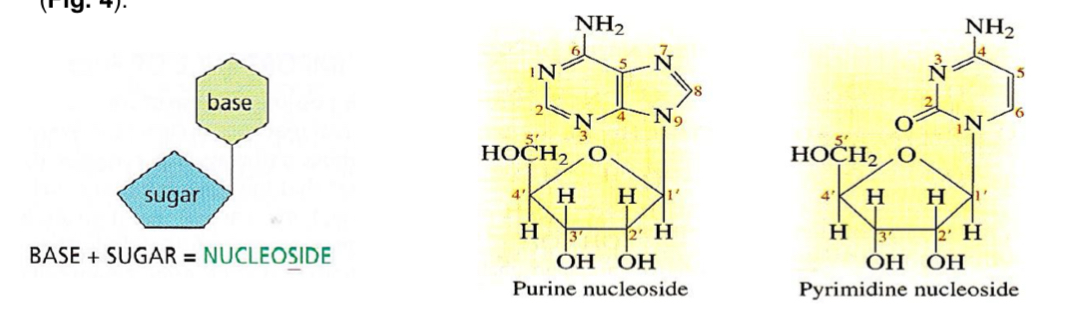
pentose + nitrogenous base = nucleoside
2 types of nucleic acids
a. Deoxyribonucleic acid (DNA) - pentose sugar is deoxyribose
—> deoxyribonucleotides are the monomers of DNA
b. Ribonucleic acid (RNA) - penrose sugar is ribose
—> ribonucleotides are the monomers of RNA
Pentose sugar
5-carbon sugars and occur as ring forms
in nucleic acids, the 5’ carbon is linked in an ester bond to the phosphate group
the 1’ carbon is linked in a glycosidic bond to the nitrogenous base
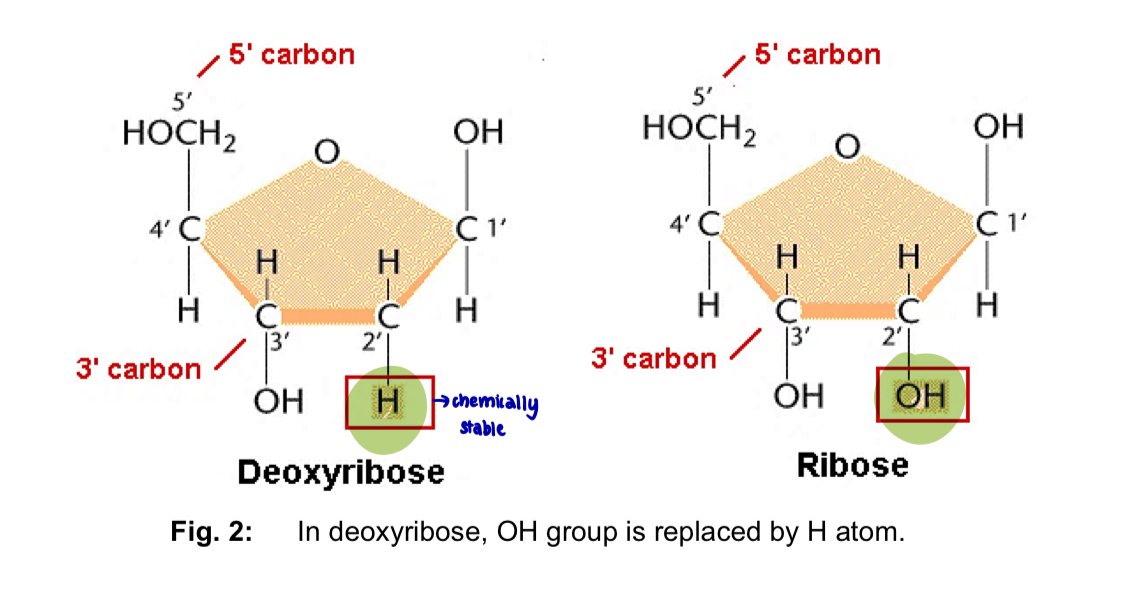
difference between ribose and deoxyribose sugars
(OH or H) at the 2’ carbon of deoxyribose, the hydroxyl group (-OH) is replaced by a hydrogen atom (H)
led to significant differences in structure and functions
—> (DNA coils more than RNA) partial negative charge of the -OH group in ribose repels the -ve charge of the phosphate, preventing the RNA chain from coiling in as tight as a helix as it does in DNA
—> RNA is more susceptible to chemical and enzyme degradation
Nitrogenous bases
nitrogenous base has a nitrogen-containing ring structure
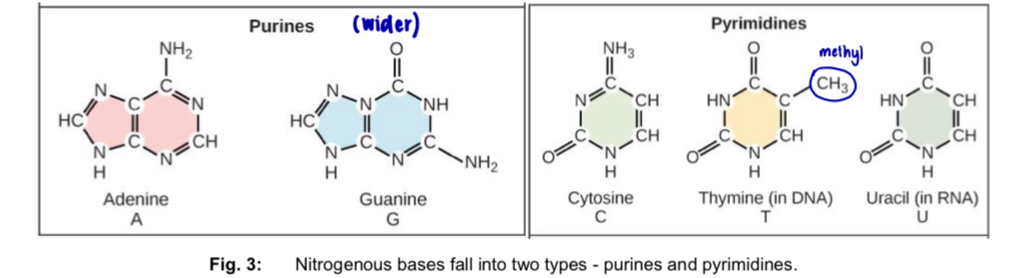
2 types: purines & pyrimidines
Purines
—> 6-membered ring fused to a 5-membered ring
DNA: Adenine (A) and Guanine (G)
RNA: A and G
Pyrimidines
—> 6-membered ring
DNA: Cytosine (C) and Thymine (T)
RNA: Cytosine (C) and Uracil (U)
*pyramid = 1 = single ring
*Pure As Gold
Cut The Py
difference between T and U is the presence of a methyl substituent at the 5-carbon
Nucleoside
nucleoside = pentose + nitrogenous base
occurs with the elimination of water (condensation)
1’ carbon of the pentose is linked in a glycosidic bond to the nitrogenous base
2 types: ribonucleoside, deoxyribonucleoside
Nucleotide
nucleotide = nucleoside + phosphate group
formed by further condensation between the nucleoside and phosphate group, forming a phosphoester bond btw the 5’ carbon of pentose and phosphate group
number of phosphate groups linked to pentose sugar varies from 1 to 3
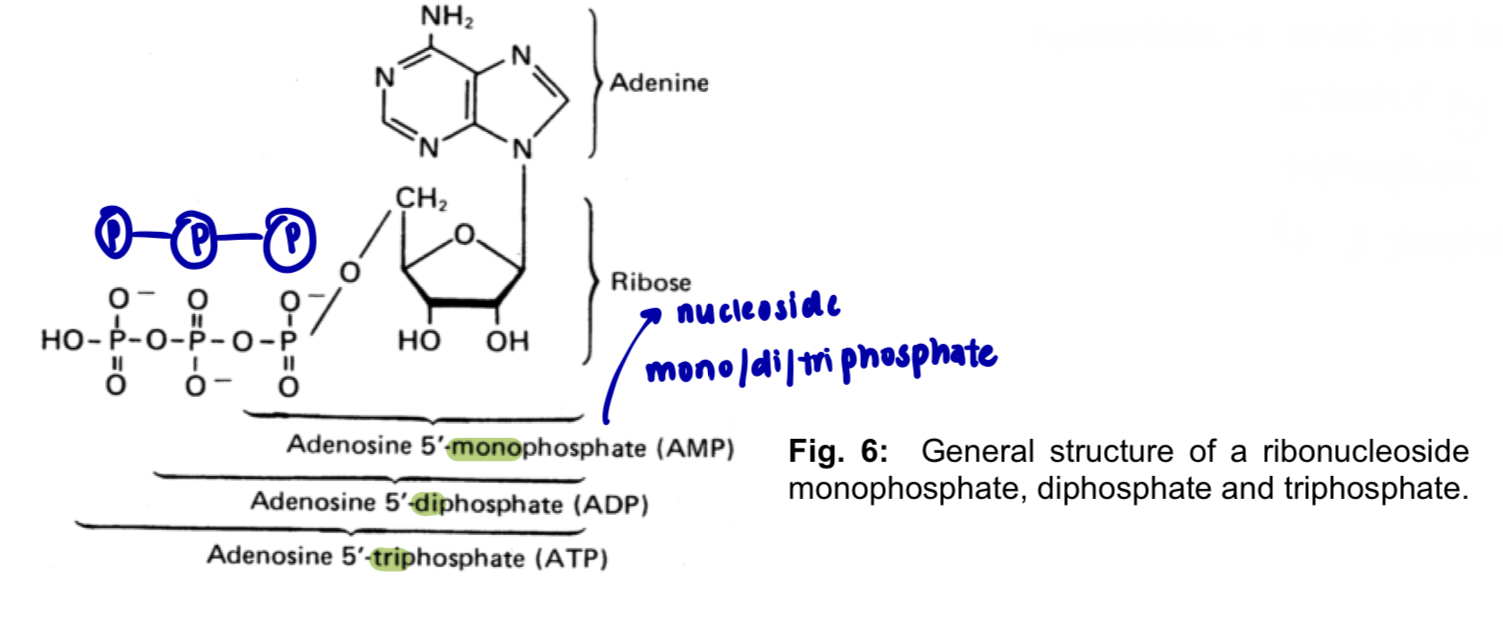
1 phosphate group —> nucleoside monophosphate e.g. AMP, adenosine monophosphate
2 phosphate groups —> nucleoside diphosphate e.g. ADP, adenosine diphosphate
3 phosphate groups —> nucleoside triphosphate
e.g. ATP, adenosine triphosphate
*nucleotides must first be activated by triphosphate (2 phosphates are released)
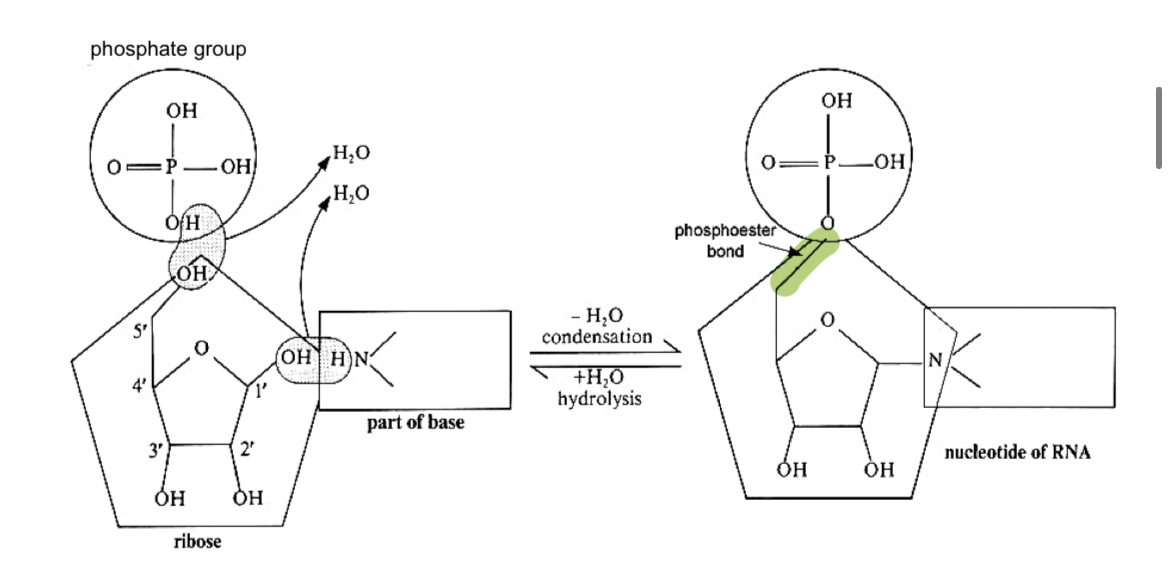
mono/di/triphosphate groups
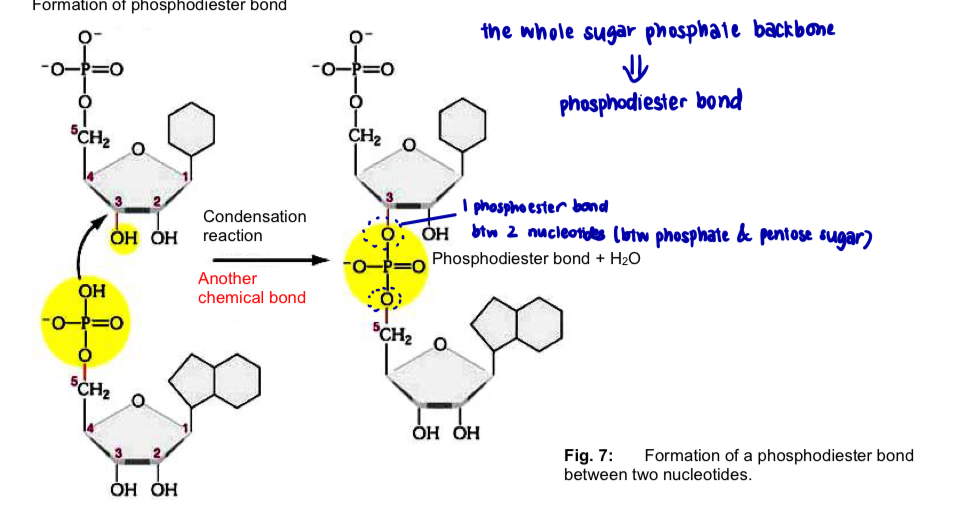
Formation of dinucleotides and polynucleotides
two nucleotides join to form a dinucleotide by condensation btw the 5’-phosphate group of one nucleotide and the 3’ hydroxyl group of the other to form a phosphodiester bond
*phosphoester bond btw 2 nucleotides
VS phosphodiester bond (the whole sugar phosphate backbone)
condensation rxn btw nucleotides is repeated several million times to form a polynucleotide
phosphodiester bonds btw 5’ phosphate and 3’ hydroxyl groups of nucleotides form a linear, unbranched sugar-phosphate backbone
phosphodiester bonds are strong and covalent bonds
—> confer strength and stability on the polynucleotide chain - prevents breakage of the chain during DNA replication
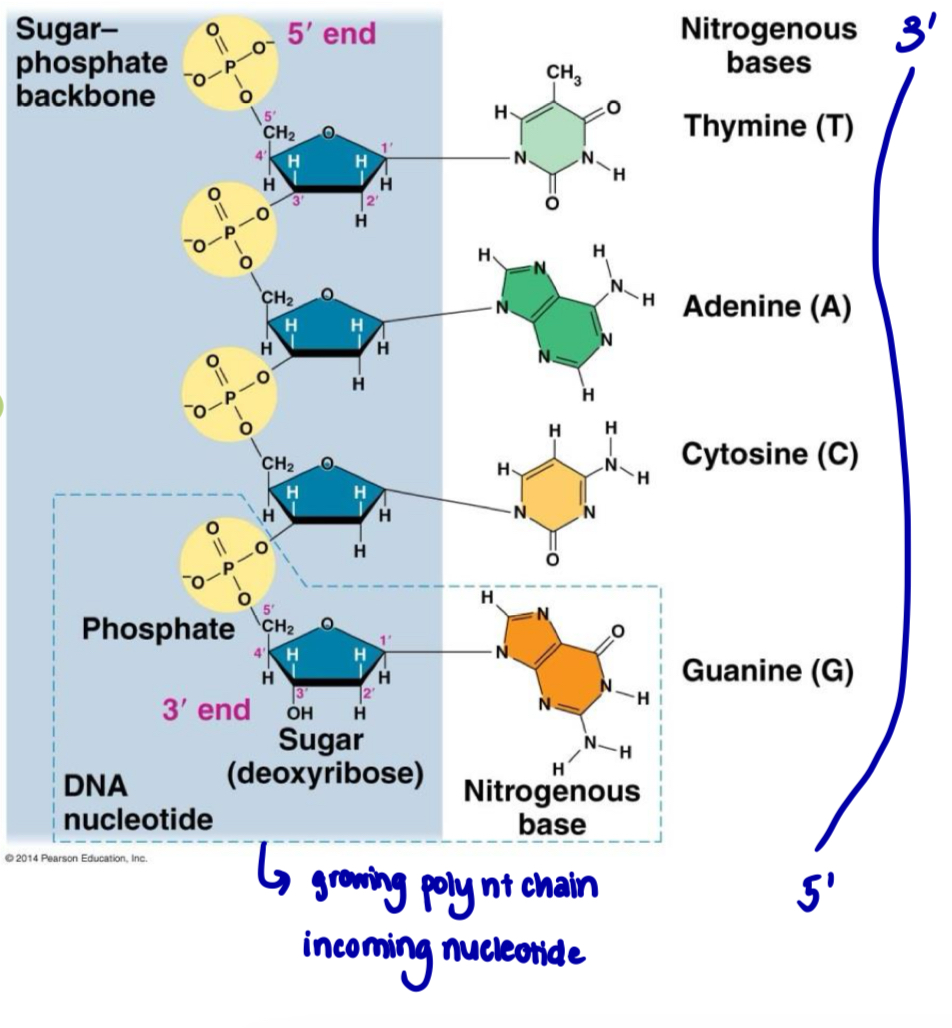
Polarity/directionality in a polynucleotide
the manner in which deoxyribonucleoside triphosphate are added to the 3’ end of a growing chain has resulted in a polynucleotide molecule that has polarity or directionality
each DNA/RNA strand/chain has 2 free ends that are chemically different from each other
5’ end with a free 5’ carbon carrying a phosphate group
3’ end with a free 3’ carbon carrying a hydroxyl group
DNA or RNA base sequence is read in a 5’ to 3’ direction —> directionality/polarity
Physical structure of DNA
a DNA molecule is double helix comprising 2 DNA strands
based on X-ray diffraction data, DNA is
long, thin molecule, diameter 2nm
2 strands
coiled in the form of a double helix, the helix making one complete twist every 3.4 nm
has 10 bases to each complete turn of the helix
ATGC base composition
base composition of DNA of an organism is constant throughout all the somatic cells and is characteristic for a given species
no. of A = no. of T; no. of G = no. of C
there’s always an equal proportion of purines (A+G) and pyrimidines (C+T)
specific complementary base pairing between A on one DNA strand and T on the other, btw G and C
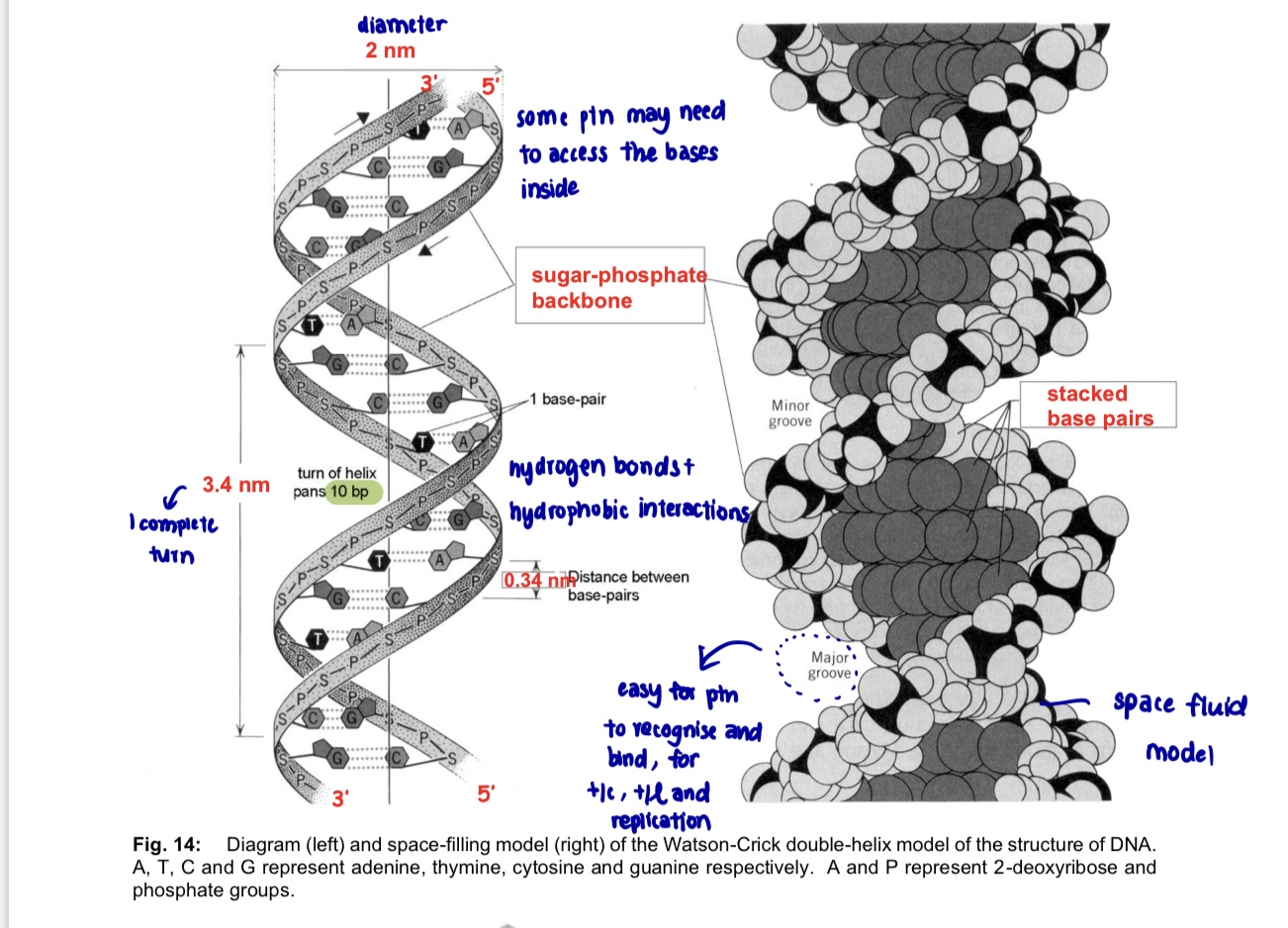
The Double helix of DNA
DNA consists of 2 polynucleotide strands/chains
each strand forms a right-handed helix and the two strands coil around each other to form a double helix
1 DNA molecule = 1 DNA double helix = 2 polynucleotide strands
the diameter of the helix is uniformly 2nm (Just enough space for 1 purine + 1 pyrimidine in the centre of the double-helix)
strands run in opposite directions i.e. antiparallel
5’ to 3’, the other is 3’ to 5’
each strand has a sugar-phosphate backbone with
phosphate groups that project outside the double helix since they are hydrophilic
nitrogenous bases that orientate inwards toward the central axis at almost right angles
bases of the opposite strands are bonded together by relatively weak hydrogen bonds
base pairs are stacked 0.34 nm apart along the central axis of the helix as a result of hydrophobic interactions contributing to overall stability of the molecule
double helix makes a complete turn every 10 base pairs, so each turn is 3.4 nm
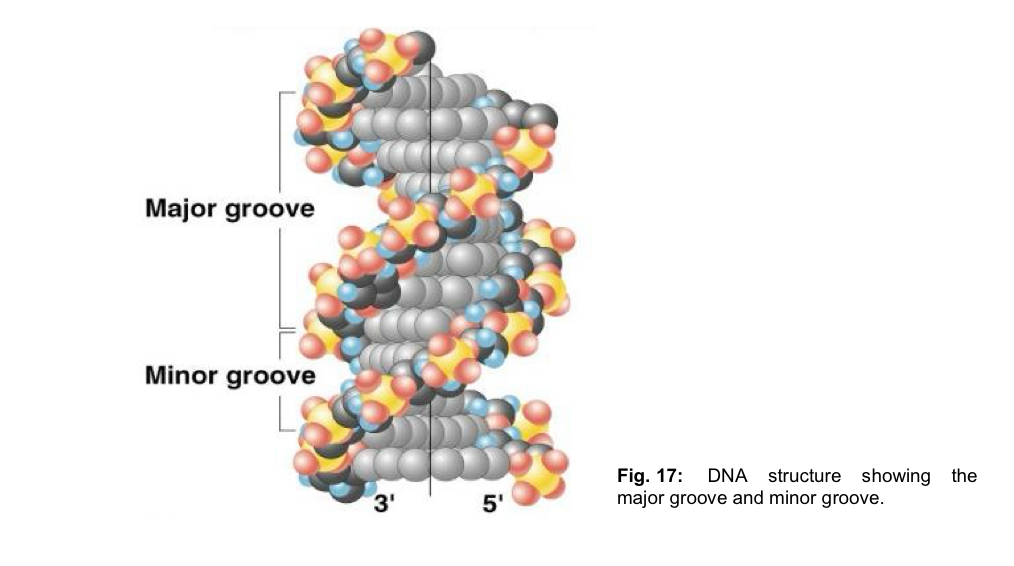
there are grooves of unequal sizes btw the sugar phosphate backbones - major grooves and minor grooves
both these grooves are large enough to allow protein molecules to gain access and make contact with the bases
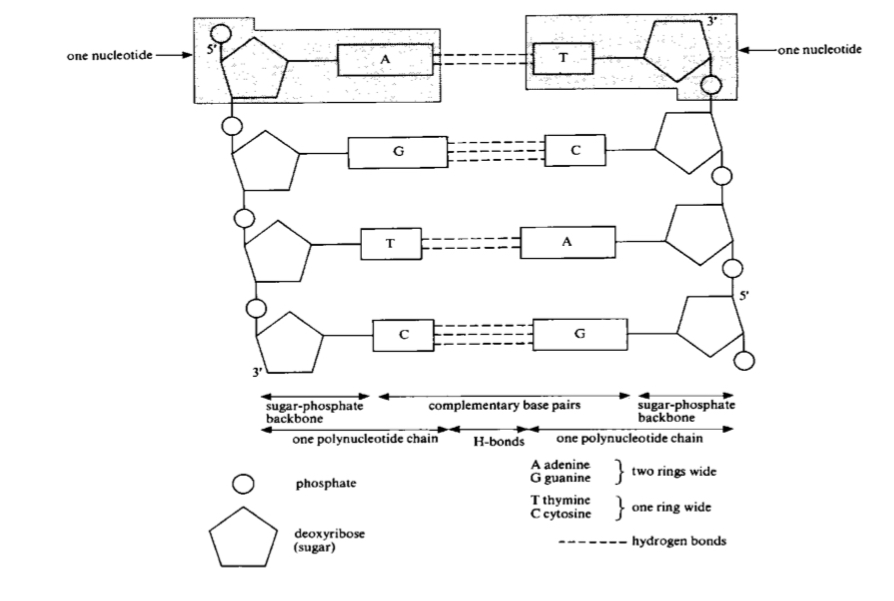
Complementary base pairing between DNA strands
complementary base pairing occurs btw A and T (2 hydrogen bonds) and btw C and G (3 hydrogen bonds)
base sequence in one strand determines the base sequence in the complementary strand
weak hydrogen bonds make it relatively easy to separate the two strands of the DNA e.g. by heating
separating the A-T pair is easier than separating the G-C pair, since A-T pair only involves 2 H bonds while G-C involves 3 H bonds
Grooves
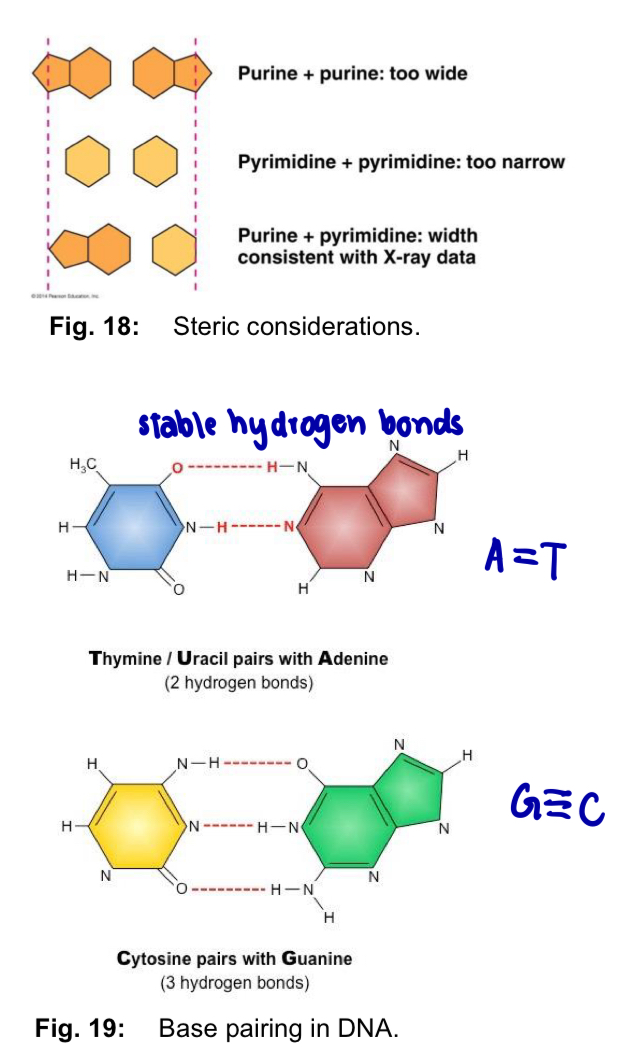
Why form complementary base pairs?
Steric restrictions
sugar-phosphate backbone of each polynucleotide chain has a regular helical structure
DNA double-helix has a uniform diameter of 2nm
T and C are pyrimidines - single ring
A and G are purines - twice as wide as pyrimidines
pair purine with pyrimidine = fits diameter
purine + purine = too wide
pyrimidine + pyrimidine = too narrow
Hydrogen bond factors
each nitrogenous base has chemical side groups (H, N and O) - hydrogen bonds with its appropriate partner
such chemical side groups in purines and pyrimidines have well defined positions
A is capable of forming 2 hydrogen bonds with T, while G is capable of forming 3 hydrogen bonds with C
Important notes on CBP
significance of CBP - since the 3D structure of DNA is only stable when the base pairs are complementary = base sequence of one strand could determine the base sequence of its complementary strand —> **necessary in DNA replication and transmission of the genetic information stored
variation of linear base sequence - linear sequence of the 4 bases can be varied
1 nucleotide —> 4 combinations
10 nucleotides —> 4^10 combinations
humans have 3 × 10^9 nucleotide pairs —> 1 048 576 combinations
each gene has a unique base sequence
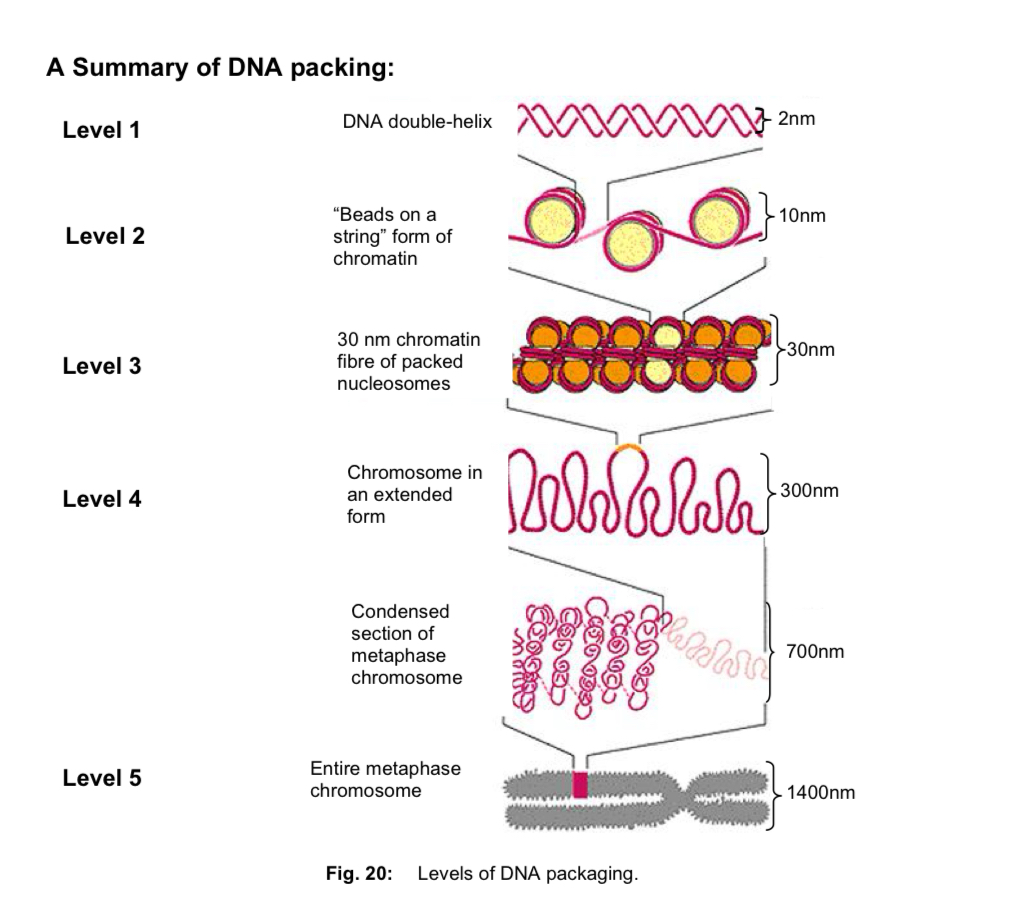
Packing of DNA in eukaryotic chromosomes
human cell contains 46 chromosomes, each of which is a single DNA molecule bundled up with various ptn
inherit 23 chromosomes from each parent
each set of 23 chromosomes encodes a complete copy of our genome, made up of 6 × 10^9 nucleotides
chromosomes must condense intro structures that are more easily managed = properly distributed to each daughter cell during cell and nuclear division
DNA packing —> proteins helping in the folding and condensation of DNA (into highly compact chromosome)
Level 1: DNA double helix
Level 2: beads on a string form chromatin
Level 3: 30nm chromatin fibre of packed nucleosomes
Level 4: chromosome in an extended form
Level 5: metaphase chromosome
Stable, Invariant storage of genetic information
genetic information that must be stored/preserved lies in the specific order of the base pairs i.e. base sequence must be stable and invariant
DNA is relatively resistant to spontaneous changes (mutations) = stable storage
structural features that stabilise the DNA double-helix
extensive hydrogen bonds btw base pairs
hydrophobic interactions btw the stacked base pairs
exposure to outside influences of only the sugar-phosphate backbone
nitrogenous bases being safely tucked inside the double-helix
eukaryotes only - DNA double helix being tightly wound around histones to form a repeating array of nucleosomes —> folded into chromosome, in which DNA is prevented from thermal and physical damage
structural features —> invariant base sequence
specific, complementary base pairing btw DNA strands
genetic info is redundant (present more than once) in the DNA
if base sequence in one of the two strands is accidentally altered, cell discards the damaged strand, makes a perfectly good strand by using remaining intact strand as a template
redundancy of genetic info helps to maintain its integrity
Replication of DNA
WHY
to ensure that the genetic information is passed down the progeny faithfully, there must be a high degree of accuracy during the copying process
daughter DNA molecules must be exactly the same as parental DNA molecule in terms of sequence information
DNA strand templates for replication
two strands of DNA are complementary
when a cell copies a DNA molecule, each strand serves as a template for ordering nucleotides into a new, complementary strand
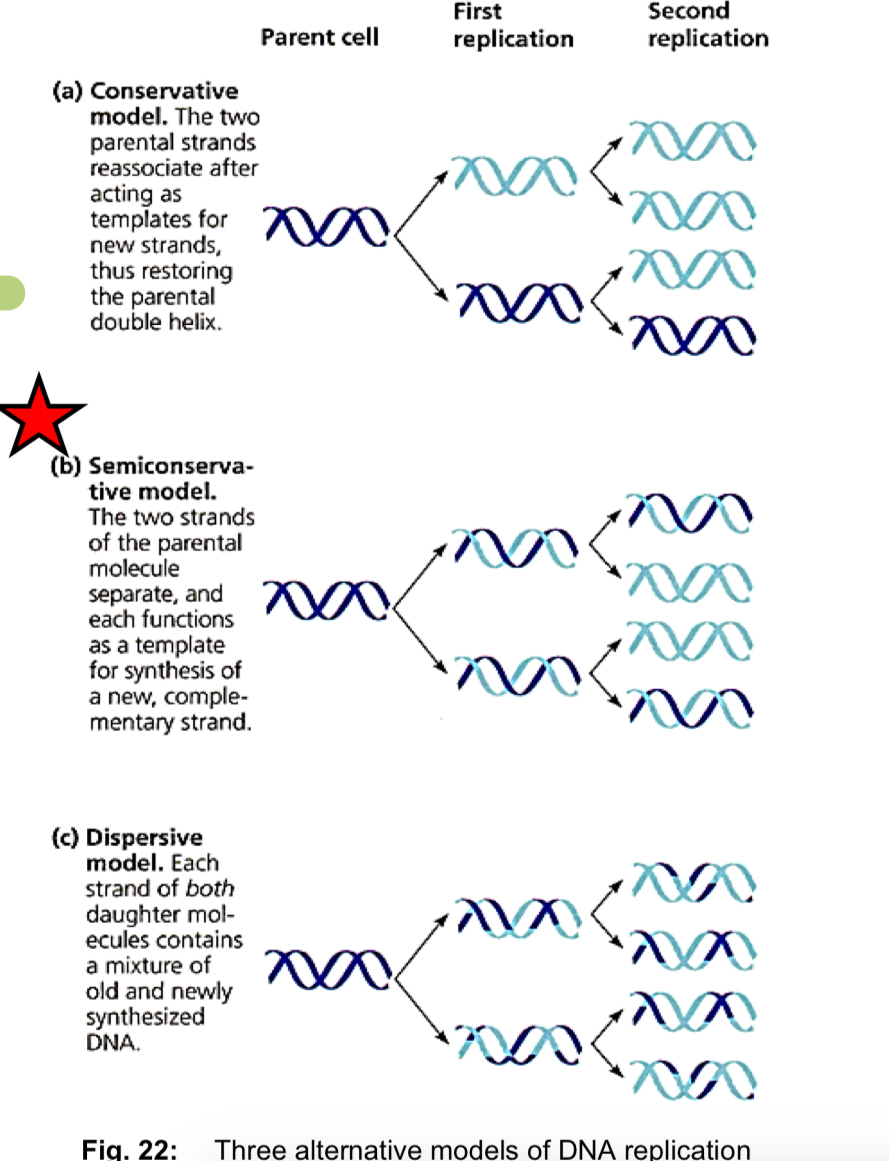
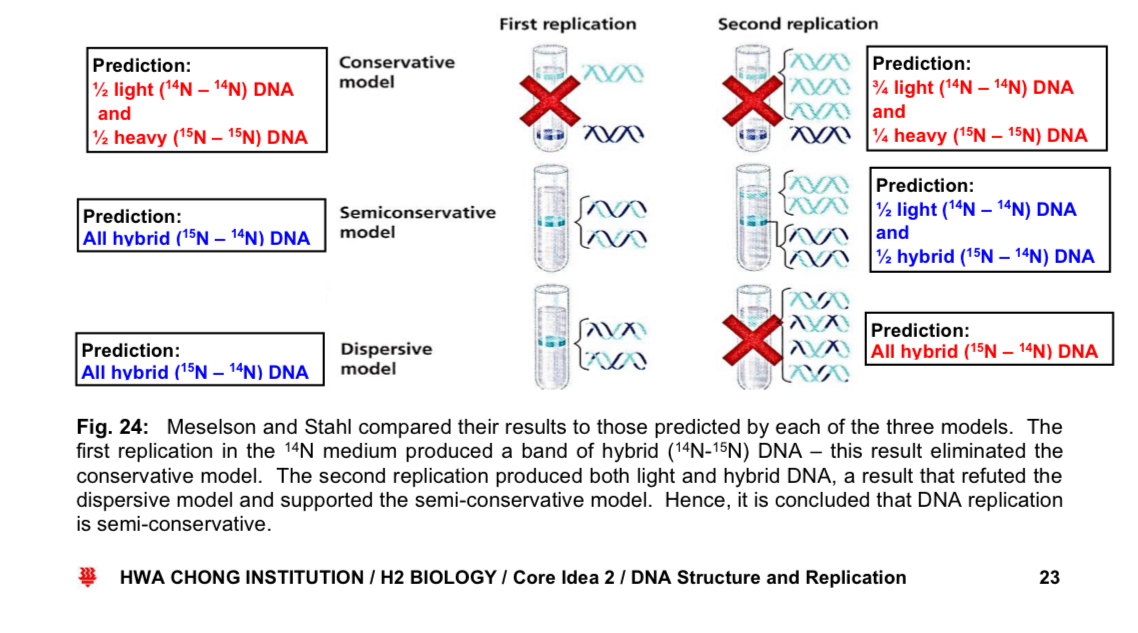
Models of DNA replication - semi-conservative model
*Semi-conservative model
the two DNA strands unwind and separate from each other, hydrogen bonds btw CBP are broken
each DNA strand then acts as a template for the assembly of a complementary strand
nucleotides line up singly along the template DNA strand according to CBP
DNA polymerases join the nucleotides together at their sugar-phosphate moieties
upon completion of DNA replication, 2 identical daughter DNA molecules are produced from a single* parental DNA molecule
***each of the two daughter DNA molecules consists of 1 parental DNA strand + 1 newly-synthesised daughter DNA strand
other proposed models
—> Conservative model: parental DNA molecule emerges from replication process intact (it is conserved) and generates DNA copies consisting of entirely new molecules
—> Dispersive model: all 4 strands of DNA following replication have a mixture of old and new DNA
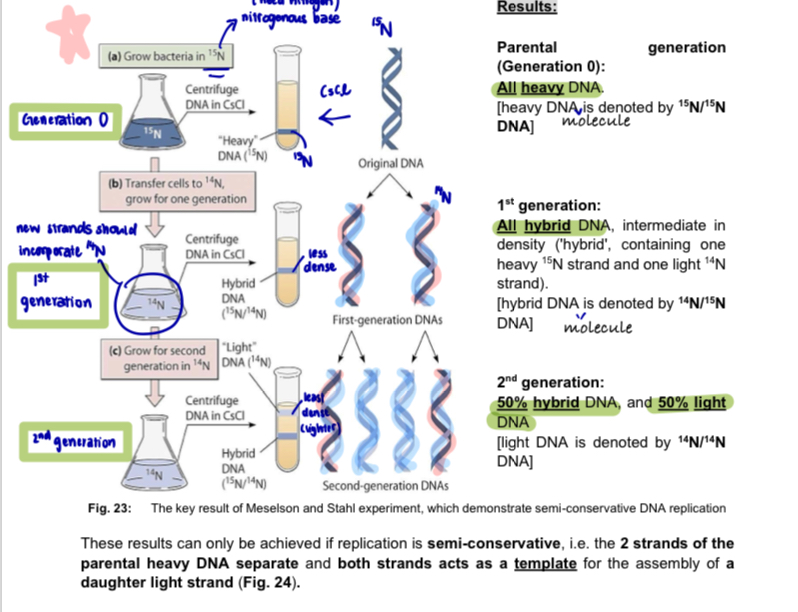
Meselson-Stahl Experiment: evidence for semi-conservative replication
used isotopes of nitrogen 14N & 15N
for many generations, cells of the bacterium (Escherichia coli) were grown on medium containing only the heavy isotope of nitrogen 15N - 15N was incorporated into all the nitrogenous bases —> heavy DNA
bacteria were then transferred to medium containing only the light isotope of nitrogen 14N and allowed to divide just once (1st generation of bacteria)
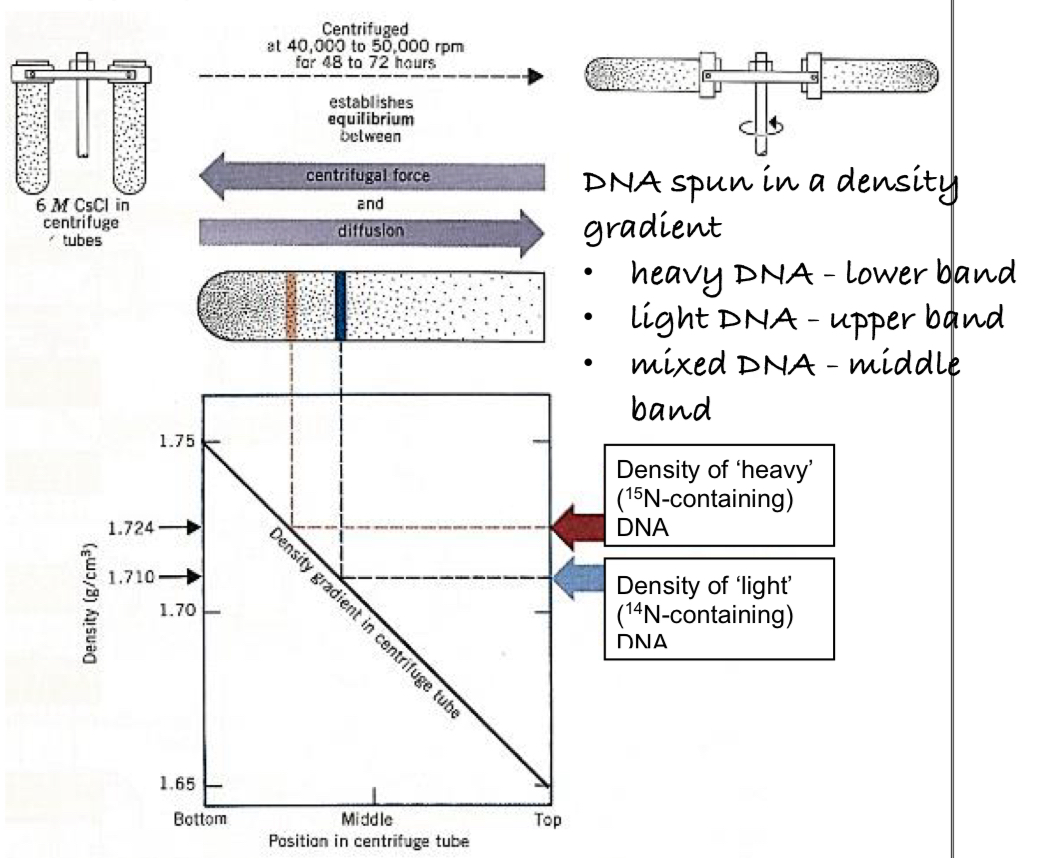
density-gradient centrifugation was performed on a DNA extract from the bacteria
DNA is separated on the basis of density
heavier 15N DNA molecules are denser than the 14N DNA molecules, hence 15N DNA molecules are spun further down in the centrifuge tube where CsCl were denser
lighter 14N DNA molecules were closer to the top where CsCl was less dense
density-gradient centrifugation:
rapid rotations to separate particles
a CsCl density gradient with increasing density towards the bottom of the centrifugation tube is formed because of the equilibrium btw
sedimentation of the CsCl to the bottom of the spinning tube as a result of centrifugal forces
diffusion of CsCl towards the top of the tube
DNA molecules move to the position where their density equals that of CsCl and floats at that position
these bacteria were allowed to undergo a second round of replication and binary fission (in the presence of 14N) producing the 2nd generation
DNA was again separated by density centrifugation
Semi-conservative model illustration
prediction of the DNA strands with the different models
Mechanism of DNA replication
actual process
is very complex, timing of steps is very precise —> double helix must unwind while the replication machinery copies the two anti parallel strands simultaneously
is very fast - each human cell has approximately 3 × 10^9 base pairs, takes the cell just a few hours to copy all these DNA
is very accurate
requires the cooperation of a large team of enzymes and other proteins, as well as ATP
Diagram of DNA replication
Locations of origins of replication
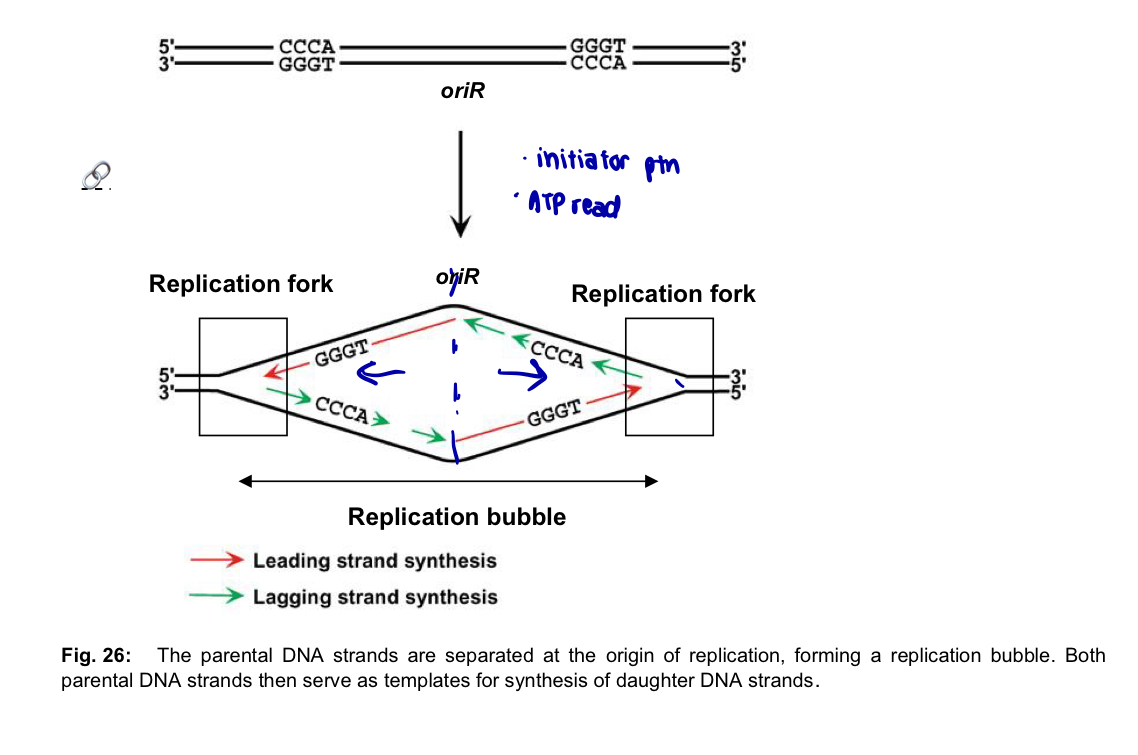
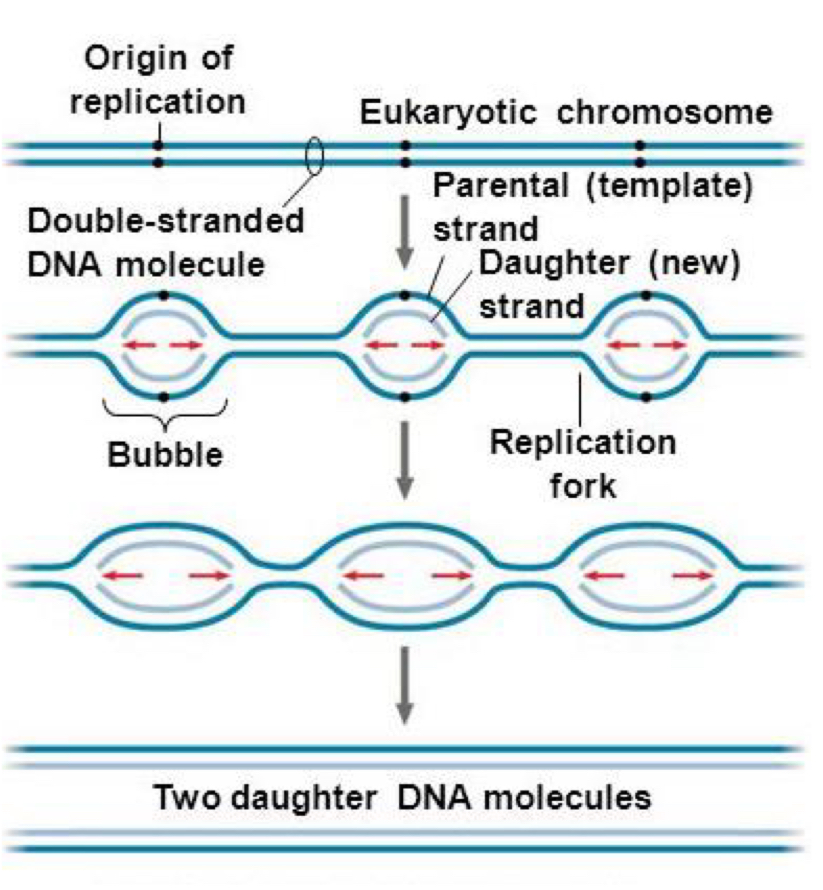
DNA replication begins at one or more sites on the DNA molecule called origins of replication (oriR)
each oriR is a specific sequence of nucleotides, which is generally A-T rich (easier to disrupt the bonds cos only 2 hydrogen bonds btw A and T)
proteins that initiate DNA replication (initiator proteins) recognise this sequence and bind to the oriR sequence - DNA double helix is separated into two strands —> replication bubble
length of DNA unwound to initiate replication is typically ~ 50bp, ATP required
at each end of a replication bubble Y-shaped structure called a replication fork, where the new strands of DNA are synthesised
2 replication forks move away from the oriR as replication proceeds bidirectionally, until the entire DNA molecule is separated
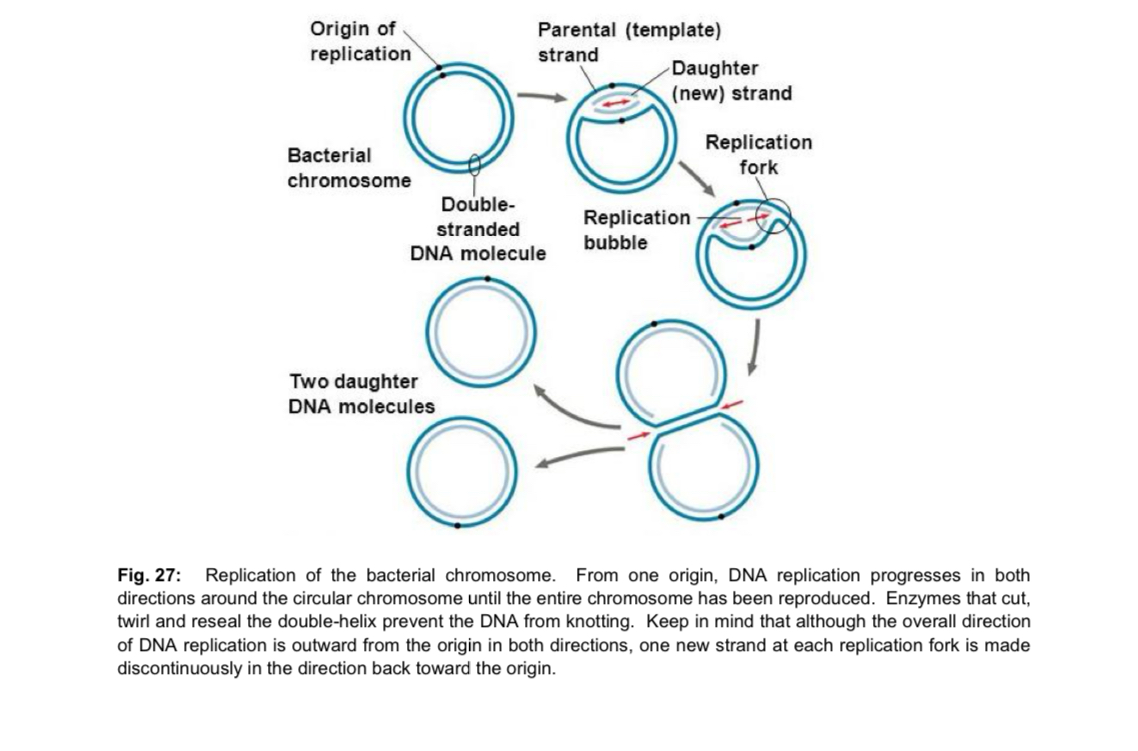
Prokaryotes (bacteria)
prokaryotic chromosome is a small circular DNA molecule, with a single origin of replication
DNA replication proceeds bidirectionally from the origin of replication to a termination site located approximately halfway around the circular chromosome —> synthesis of 2 daughter DNA molecules
Eukaryotes
eukaryotic chromosome is much larger and consists of a linear DNA molecule, with multiple origins of replication
advantage of having multiple oriR: multiple replication bubbles form and eventually fuse = speed up the copying of very long DNA molecules
replication takes approximately 8 hours in human cells with multiple oriR; if only 1 oriR, 100 times longer
impt given the much larger size of eukaryotic chromosome
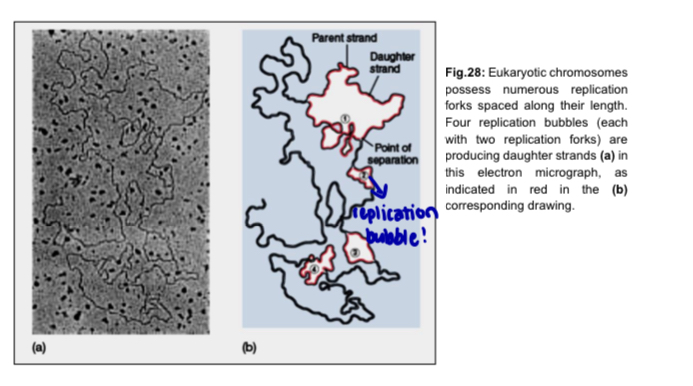
Replication of eukaryotic chromosome
Replication begins at multiple origins of replication, where the two parental strands separate to form replication bubbles
the bubbles expand laterally as DNA replication proceeds bidirectionally
eventually, the replication bubbles fuse and synthesis of the daughter strands is complete
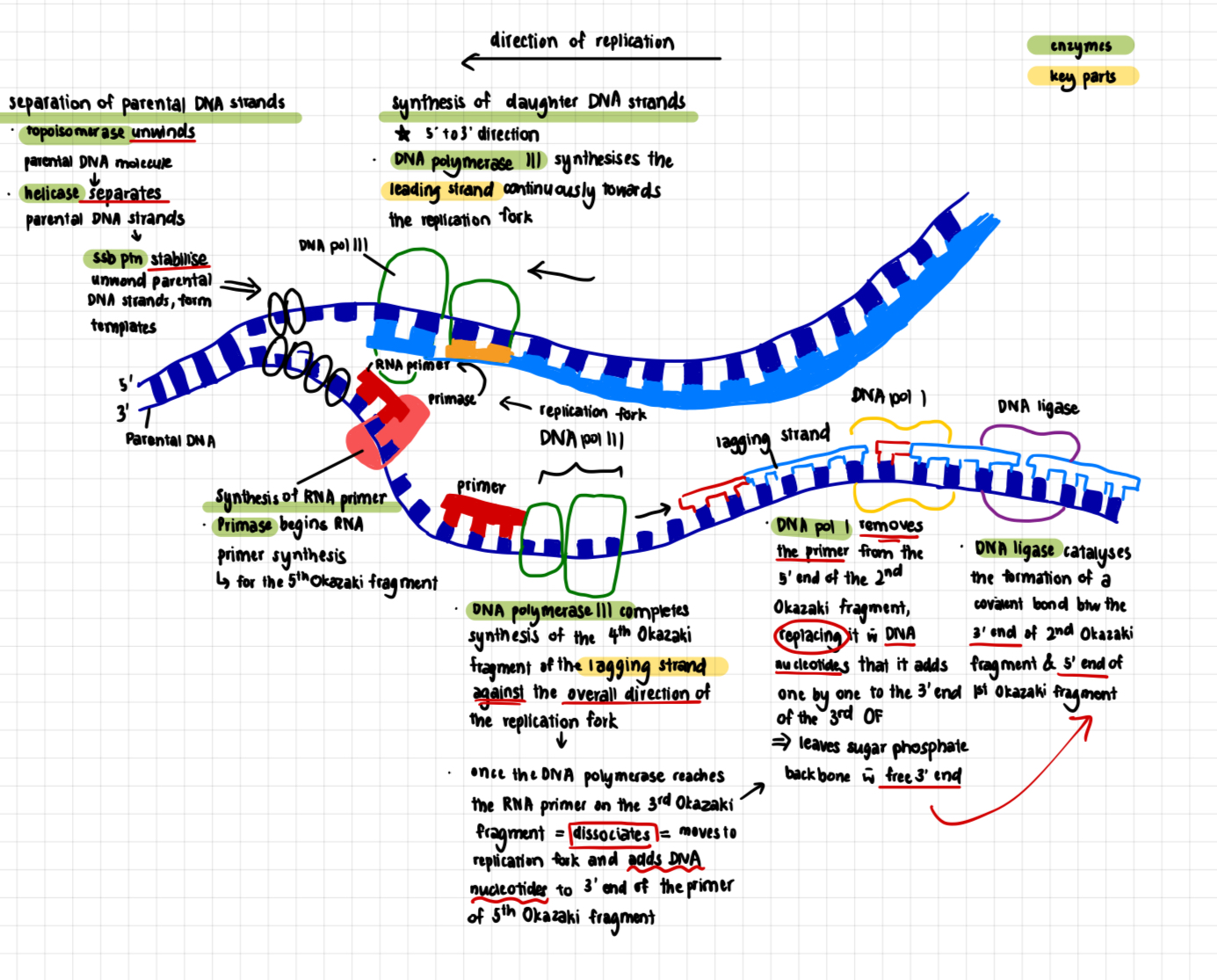
Separation of parental DNA strands
separate base pairs of parental DNA molecules so that both DNA strands can act as templates for the synthesis of daughter DNA strands
origin of replication - where separation initially occurs
3 proteins involved
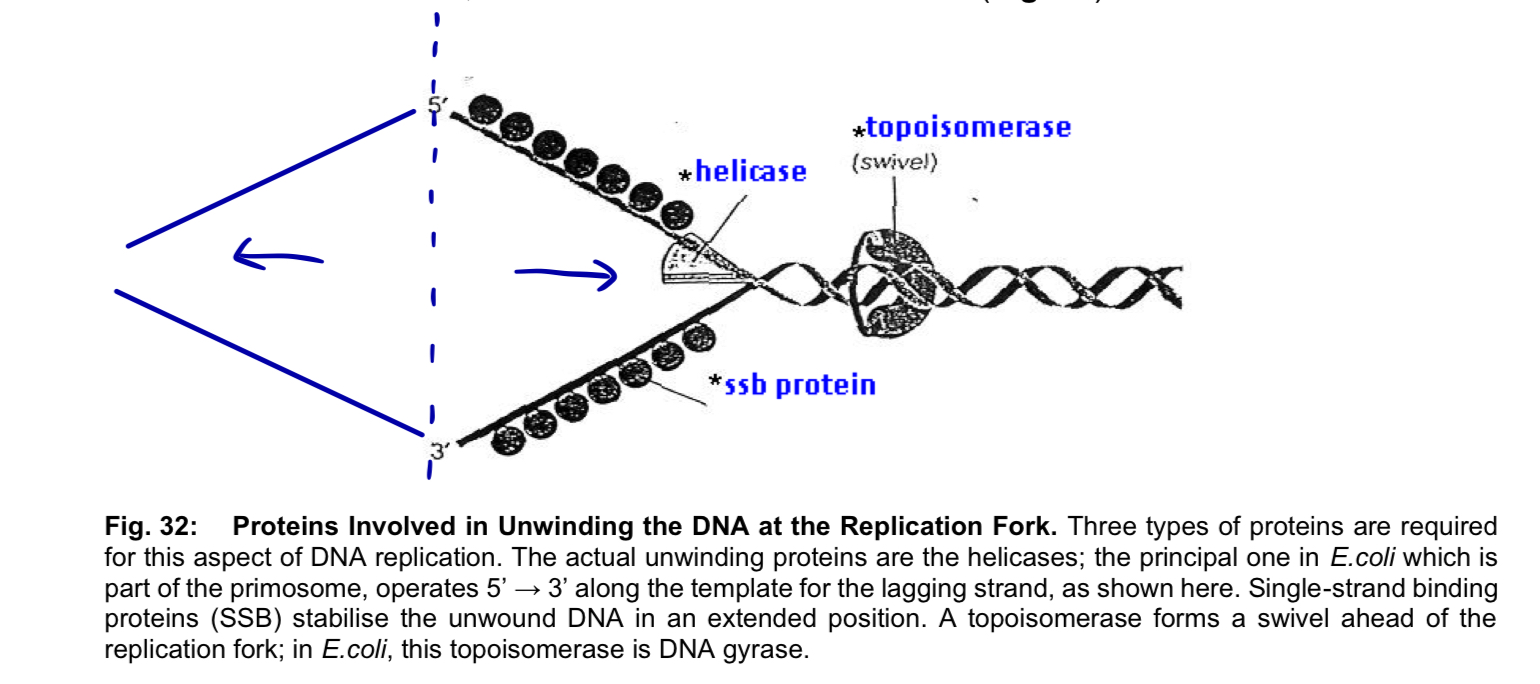
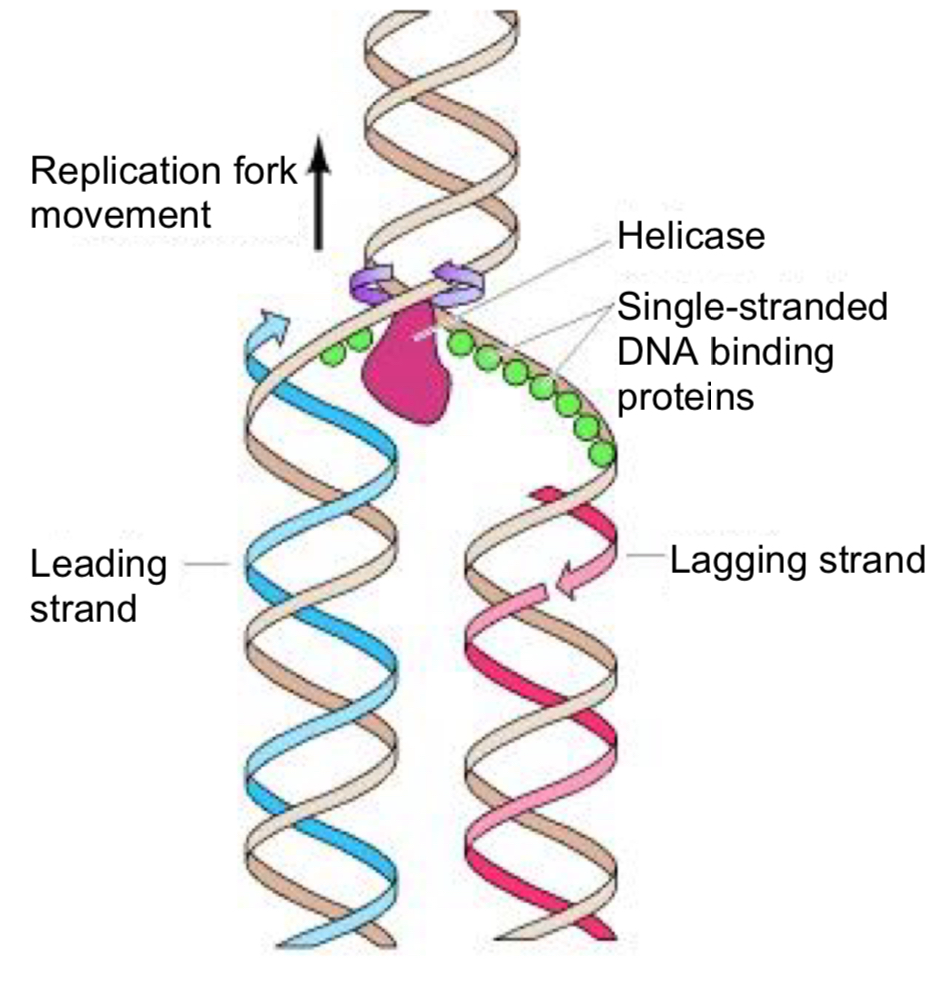
helicases (unwind and separate)
after initiation, helicase bind to one strand of the DNA molecule
using ATP as energy source, helicases break the hydrogen bonds holding the two strands of DNA together = unwind the DNA double-helix and separates the parental DNA strands at the region of the replication fork
each of the 2 parental DNA strands serve as the template for the synthesis of a new DNA strand
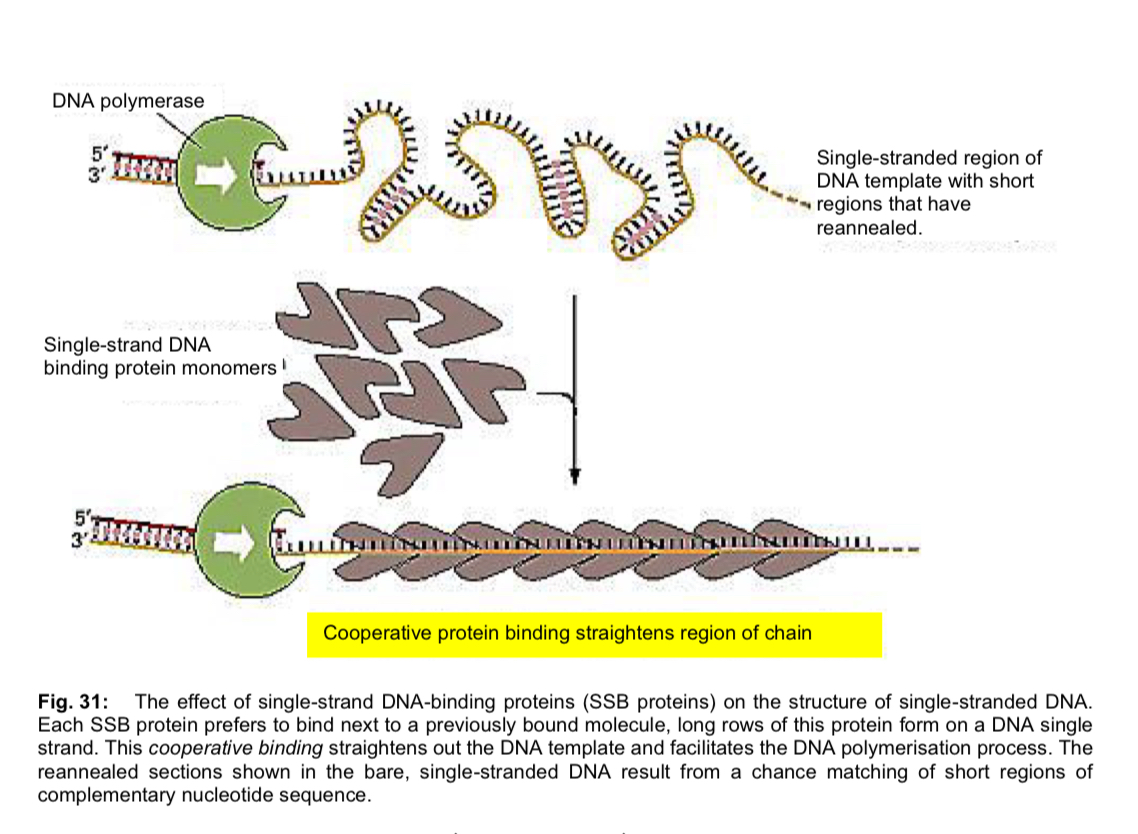
single-strand DNA binding proteins (ssb proteins)
unwound single-stranded portion of the DNA double-helix is temporarily stabilised by the binding of single-strand DNA-binding proteins
prevents the single-stranded (ss) DNA from re-annealing to reform the duplex
keeps the two parental strands in the appropriate single-stranded condition to act as template
protects the ssDNA, which is very unstable, from being degraded
topoisomerases
unwinding causes tighter twisting/supercoiling ahead of the replication fork, resulting in tension/torque
topoisomerases cleave a strand of the helix to create a transient single-stranded nick
relieves strain on the DNA molecule by allowing free rotation (swiveling) around the intact strand, and then reseal the broken strand
Helicase diagram
SSB proteins
Synthesis of RNA primers
limitation 1 of DNA polymerases: none of the DNA polymerases can initiate the synthesis of a DNA strand on its own. DNA synthesis cannot occur de novo.
solution: RNA primer to initiate synthesis of a DNA strand
a portion of the parental DNA strand serves as template for making the RNA primer with the complementary base sequence
an enzyme called primate joins the ribonucleotides to make the primer.
primer is about 10 nucleotides losing in eukaryotes