Écologie des eaux douces : Introduction à la physico-chimie des eaux douces
1/22
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
23 Terms
Pourquoi les eaux douces sont une ressource rare mais essentielle, et pourquoi les eaux de surface comptent autant ?
À l’échelle de la planète, la majorité de l’eau est salée. Dans la fraction douce, une grande partie est immobilisée dans les glaces/pergélisol, et une autre part importante est stockée dans les eaux souterraines. Au final, les eaux de surface (lacs, étangs, rivières) représentent une portion infime de l’eau totale mondiale. Pourtant, ce sont elles qui soutiennent directement une grande partie de la biodiversité, des services écosystémiques et des usages humains (eau potable, agriculture, industrie, loisirs). Conclusion : une ressource très limitée, concentrée dans des milieux vulnérables et fortement exposés aux perturbations.
Cycle de l’eau et “rivières atmosphériques” : pourquoi les écosystèmes terrestres influencent la pluie et les eaux douces ?
Le cycle de l’eau relie évaporation → condensation → précipitations → ruissellement/infiltration. Un point fort est le rôle des rivières atmosphériques : de grands flux de vapeur d’eau transportés par l’atmosphère. Dans des régions comme l’Amazonie, l’évapotranspiration de la forêt produit d’immenses quantités d’humidité qui peuvent alimenter des régions éloignées (parfois en volumes comparables ou supérieurs à des fleuves). Cela montre que le fonctionnement des eaux douces est interconnecté au climat et aux systèmes terrestres : dégrader une forêt, c’est potentiellement modifier le régime de précipitations et donc les apports en eau des bassins versants.
Limnologie et types d’écosystèmes lentiques : mare, étang, lac — quelles différences écologiques majeures ?
La limnologie étudie les eaux continentales (équivalent de l’océanographie pour l’océan). Dans les systèmes lotiques (rivières), l’eau circule ; dans les systèmes lentiques (eaux stagnantes), la dynamique est dominée par la stratification et les échanges internes.
Mare : petite eau stagnante, souvent sans exutoire, parfois temporaire. Nombreuses et écologiquement importantes malgré leur taille.
Étang : plus grand, généralement alimenté par un cours d’eau avec un exutoire ; peu profond, souvent assez homogène physico-chimiquement.
Lac : plus grand et plus profond ; la profondeur permet une stratification verticale (température, oxygène, lumière, nutriments). Les grands lacs (Léman, Baïkal) illustrent l’importance de la profondeur : le Baïkal contient une part énorme de l’eau douce lacustre mondiale et abrite des environnements profonds très particuliers.
Pourquoi la lumière est le paramètre numéro 1 en physico-chimie des lacs, et comment on la mesure (Secchi, zone euphotique, compensation) ?
La base des écosystèmes aquatiques est la production primaire (phytoplancton), et elle dépend d’abord de la lumière. On estime la transparence avec le disque de Secchi : la profondeur de disparition donne un indicateur simple de turbidité et souvent d’eutrophisation.
Zone euphotique : couche où la lumière permet la photosynthèse (limite ≈ 1 % de la lumière incidente). Règle pratique : profondeur euphotique ≈ 3 × Secchi (variable selon conditions).
Profondeur de compensation : profondeur où photosynthèse = respiration ; au-dessus, production nette d’O₂, en dessous, consommation nette d’O₂.
La pénétration lumineuse dépend aussi de la saison (angle solaire), nuages, vent (réflexion), et particules en suspension.
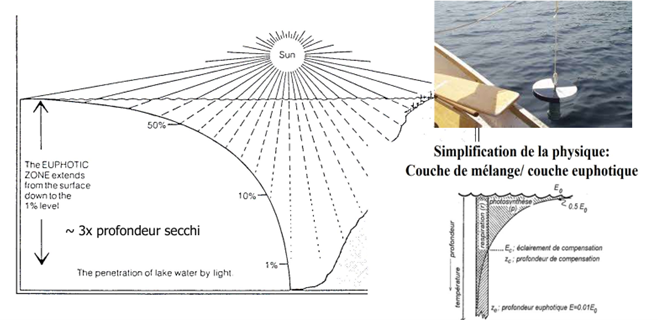
Qu’est-ce que le disque et la profondeur de Secchi ?
Le disque de Secchi est un disque circulaire blanc (ou blanc et noir) utilisé pour mesurer la transparence de l’eau. Il est descendu verticalement dans l’eau à l’aide d’un câble gradué jusqu’au moment où il devient invisible depuis la surface. La profondeur à laquelle il disparaît correspond à la profondeur de Secchi. Cet outil permet d’évaluer la turbidité de l’eau, c’est-à-dire la quantité de particules, de plancton ou de matière en suspension présentes dans le milieu aquatique. La profondeur de Secchi correspond à la profondeur maximale à laquelle le disque de Secchi reste visible depuis la surface. Plus cette profondeur est grande, plus l’eau est transparente. À l’inverse, une faible profondeur de Secchi indique une eau trouble, riche en particules, sédiments ou phytoplancton. Elle permet d’estimer jusqu’où la lumière pénètre dans l’eau. Cette information est essentielle car la lumière conditionne la photosynthèse des algues et des plantes aquatiques. Une faible transparence limite donc la production primaire. La profondeur de Secchi est également utilisée comme indicateur de pollution, d’eutrophisation ou de prolifération phytoplanctonique dans les milieux aquatiques.
Qualité du spectre lumineux dans l’eau : pourquoi la couleur de la lumière change avec la profondeur et ce que ça implique pour le phytoplancton ?
La photosynthèse utilise la lumière PAR (400–700 nm). Dans l’eau, l’infrarouge est absorbé très vite (chauffe la surface), les grandes longueurs d’onde (rouge) disparaissent progressivement, et le bleu pénètre le plus profondément. Donc le spectre change avec la profondeur, et les organismes s’adaptent via leurs pigments (chlorophylles + pigments accessoires). Résultat : la lumière structure la répartition verticale du phytoplancton, influence la stratification thermique et conditionne l’architecture des zones oxygénées/anoxiques.
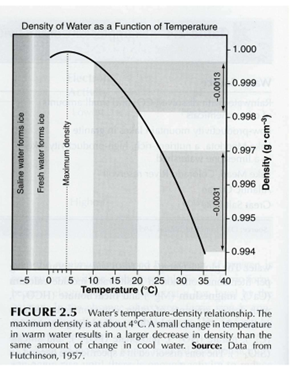
Température de l’eau : inertie thermique, densité maximale à 4°C, et pourquoi ça crée une stratification
L’eau a une forte chaleur spécifique : elle se réchauffe et se refroidit lentement, ce qui tamponne les variations thermiques (moins de stress “brutal” qu’à l’air). Particularité clé : l’eau a une densité maximale à 4°C ; au-dessus, elle devient moins dense quand elle chauffe, et en dessous de 4°C elle redevient moins dense (glace moins dense car occupe un moins grand volume que l’eau chaude → flotte). Dire que l’eau est plus ou moins dense selon la température signifie que la masse d’eau occupe un volume plus ou moins grand quand elle chauffe ou refroidit. Cette relation fait que de faibles écarts de température peuvent suffire à créer des différences de densité qui empêchent le mélange vertical en été.
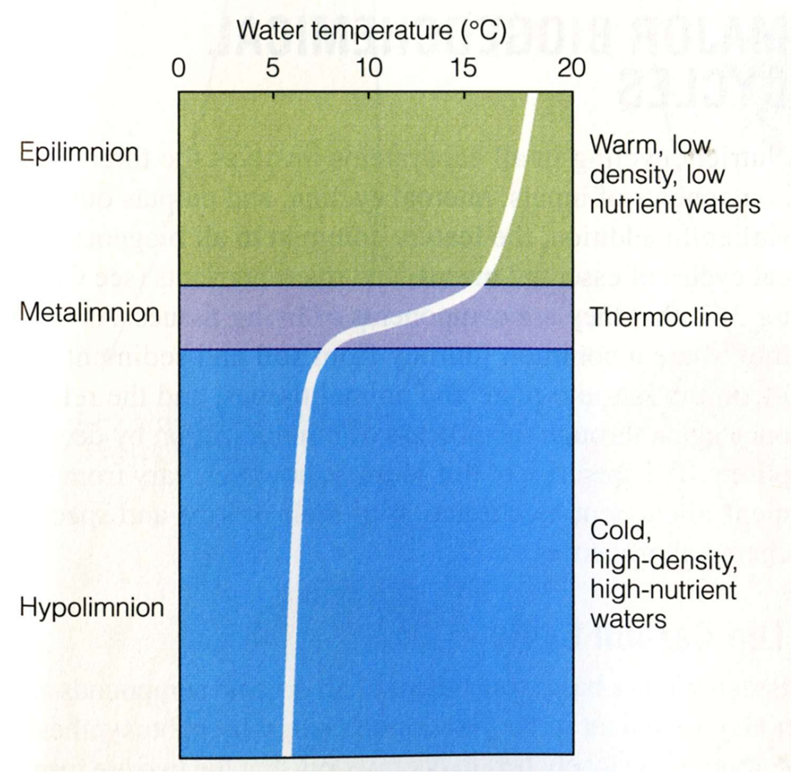
Stratification thermique : épilimnion, métalimnion/thermocline, hypolimnion — pourquoi un lac devient “deux systèmes” en été ?
Au printemps/été, la surface se réchauffe, devient moins dense et reste au-dessus : le lac se stratifie en :
Épilimnion : chaud, brassé par le vent, bien mélangé.
Métalimnion (thermocline) : zone où la température chute fortement sur peu de profondeur ; barrière au mélange.
Hypolimnion : froid, dense, peu brassé.
Conséquence majeure : les échanges verticaux sont limités, donc l’oxygène et les nutriments ne circulent plus librement ; les couches fonctionnent presque comme deux systèmes séparés, même si la matière (particules) peut sédimenter vers le fond.
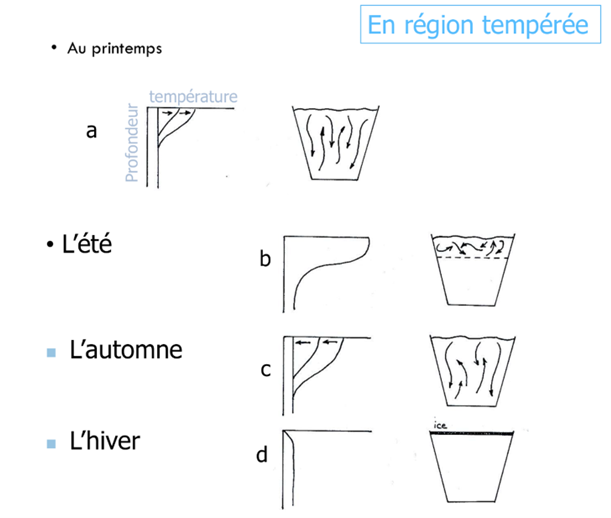
Cycle saisonnier des lacs tempérés et types de lacs selon le mélange (dimictique, monomictique, polymictique… meromictique)
Un lac tempéré suit souvent :
Printemps : colonne d’eau homogène → mélange complet.
Été : stratification installée → échanges bloqués.
Automne : refroidissement → densité se rééquilibre → second mélange.
Hiver : possible glace, brassage réduit.
Classification :Holomictique : mélange toute la colonne.
Méromictique : mélange incomplet ; couche profonde isolée (souvent densité accrue par salinité).
Amictique : pas de mélange (souvent gel durable).
Fréquence : dimictique (2 mélanges/an), monomictique (1), polymictique (mélanges fréquents), oligomictique (irrégulier). Ces régimes dépendent de latitude, altitude, profondeur, exposition au vent, morphologie du bassin.
Zonation d’un lac : verticale (euphotique/thermocline/hypolimnion) et horizontale (pélagique, littorale, benthique)
Un lac est structuré :
Verticalement : lumière, température, O₂, nutriments varient avec la profondeur (zone euphotique, thermocline, hypolimnion).
Horizontalement :
Pélagique : “au large”, domaine planctonique (phyto/zooplancton).
Littorale : proche des berges, plus de végétation, forte influence du substrat.
Benthique : fond/sédiments, organismes fixés ou vivant dans le sédiment.
Cette double structuration organise la distribution des organismes et des processus biogéochimiques.
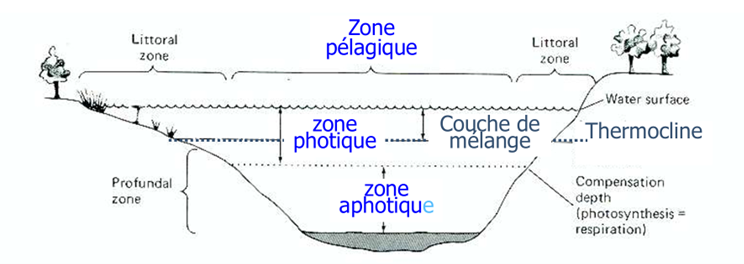
Oxygène dissous : mg/L vs % saturation, sous-saturation/sursaturation, et loi de Henry
Deux façons de décrire l’O₂ :
Concentration (mg/L) : ce que “voit” réellement le poisson ; seuils critiques pour stress/asphyxie.
% saturation : compare à la valeur théorique attendue selon température/pression/salinité ; très informatif pour l’écologue car il révèle l’activité biologique.
Sous-saturation = consommation d’O₂ (souvent par bactéries décomposeuses de matière organique).
Sursaturation (120–140% ou plus) = photosynthèse intense (bulles possibles).
Physico-chimie : loi de Henry → la solubilité dépend surtout de la température (eau chaude = moins d’O₂ soluble), aussi de la pression et salinité. S’ajoutent photosynthèse, respiration/décomposition, et ré-aération (vent/agitation ; lacs encaissés ré-oxygènent mal).
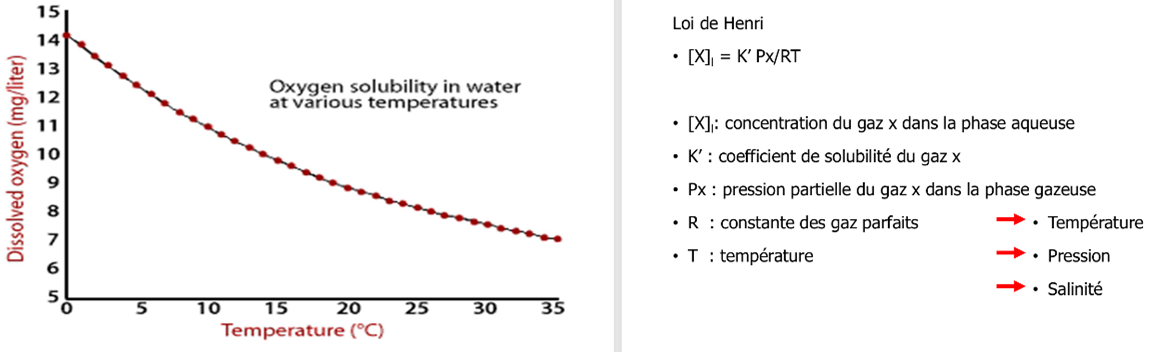
Quels facteurs influencent la quantité d’oxygène dissous dans l’eau ?
La quantité d’oxygène dissous dépend principalement de la température, de la pression et de la salinité. Lorsque la température augmente, la solubilité de l’oxygène diminue, donc l’eau chaude contient moins d’oxygène que l’eau froide. Une augmentation de la pression favorise au contraire la dissolution de l’oxygène. Enfin, plus la salinité augmente, moins l’oxygène se dissout dans l’eau.
Qu’est-ce que la saturation en oxygène ?
La saturation correspond à la concentration maximale d’oxygène que l’eau peut contenir à une température et une pression données lorsqu’elle est en équilibre avec l’atmosphère.
Le taux de saturation correspond au rapport entre la concentration réelle d’oxygène mesurée dans l’eau et la concentration théorique à saturation, exprimé en pourcentage.
La réaération correspond au transfert d’oxygène de l’atmosphère vers l’eau. Elle dépend de la différence entre la concentration réelle en oxygène et la concentration à saturation.
La sursaturation correspond à une concentration d’oxygène supérieure à la concentration de saturation. Elle apparaît souvent lors d’une forte photosynthèse des végétaux aquatiques. La déplétion correspond au contraire à une diminution importante de l’oxygène dissous dans l’eau. Elle peut être causée par la respiration des organismes, la décomposition de matière organique ou encore la pollution.
Profils verticaux d’oxygène en été : orthograde, clinograde, hétérograde — et ce que ça dit sur la trophie
En période de stratification, différents profils apparaissent :
Orthograde : O₂ assez uniforme, parfois plus élevé en profondeur (eau froide) → lac oligotrophe (peu de nutriments, faible production, peu de décomposition).
Clinograde : sursaturation en surface, chute forte, hypolimnion pouvant atteindre 0 → lac eutrophe (forte production → beaucoup de matière organique qui sédimente → décomposition bactérienne → consommation d’O₂, sans renouvellement à cause de la thermocline).
Hétérograde : pic d’O₂ à profondeur intermédiaire → maximum de photosynthèse là où lumière suffisante mais moins de photoinhibition, conditions optimales pour certaines algues.
Un simple profil O₂ + température permet d’inférer productivité, intensité de décomposition et risque d’anoxie.
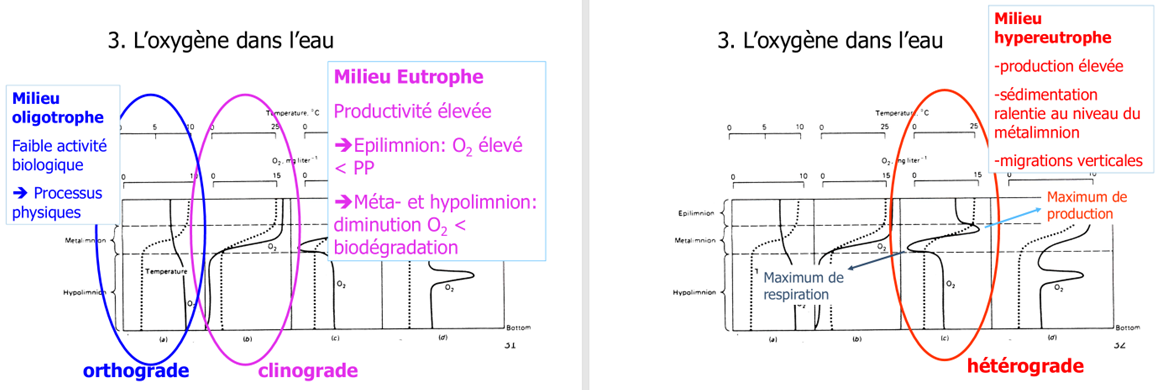
Pourquoi un pic d’O₂ apparaît parfois près du métalimnion, et comment un bloom peut conduire à l’anoxie (auto-ombrage + respiration bactérienne)
Deux mécanismes :
Accumulation près du changement de densité : le phytoplancton qui coule ralentit au niveau de la thermocline/métalimnion → zone d’accumulation de biomasse ; si la lumière y reste suffisante et la photoinhibition moindre, la photosynthèse peut y être maximale → pic d’O₂.
Cyanobactéries : elles peuvent ajuster leur flottabilité et rester dans la couche optimale, contrairement à beaucoup d’algues qui sédimentent.
Lors d’un bloom, l’auto-ombrage réduit la lumière → la production chute, mais la biomasse morte nourrit les bactéries → respiration bactérienne massive qui consomme l’O₂ et peut provoquer une déplétion rapide en profondeur, jusqu’à l’anoxie.
Carbone, CO₂ et pH : pourquoi le CO₂ est souvent limitant malgré un “carbone total” élevé, et quel rôle de tampon jouent les carbonates ?
Le CO₂ atmosphérique est peu abondant. Il se dissout bien dans l’eau, mais une fois dissous, il change de formes :
CO₂ + H₂O ⇄ H₂CO₃ ⇄ HCO₃⁻ + H⁺ ⇄ CO₃²⁻ + H⁺.
Or beaucoup de producteurs utilisent surtout le CO₂ dissous, alors que dans l’eau une grande part du carbone inorganique devient bicarbonate (et carbonate en pH alcalin). Donc le pool de CO₂ libre peut être petit → limitation possible de la photosynthèse. Les cyanobactéries peuvent souvent exploiter HCO₃⁻, ce qui les avantage en eaux alcalines pauvres en CO₂ libre.
Le système carbonate participe aussi au pouvoir tampon : bassins versants riches en CaCO₃ → pH plus stable ; roches non carbonatées → acidification plus facile (sensibilité aux pluies acides).
Qu’est-ce que la salinité, les résidus secs et la conductivité ?
La salinité correspond à la concentration totale des éléments majeurs dissous dans l’eau, généralement exprimée en milligrammes par litre. Elle représente donc la quantité globale de sels minéraux contenus dans une eau naturelle.
Les résidus secs, aussi appelés TDS (Total Dissolved Solids), correspondent à la masse totale des substances dissoutes qui restent après évaporation de l’eau et séchage à 105°C. Ils permettent d’estimer la charge minérale totale d’une eau.
La conductivité mesure la capacité de l’eau à conduire le courant électrique entre deux électrodes. Plus une eau contient d’ions dissous comme le calcium, le sodium ou les chlorures, plus sa conductivité est élevée. Elle est généralement exprimée en microsiemens (µS) et standardisée à une température de référence.
Quels sont les principaux minéraux présents dans l’eau et quelle est leur origine ?
Les eaux naturelles contiennent différents ions minéraux dissous. Les principaux cations sont le calcium (Ca2+), le magnésium (Mg2+), le sodium (Na+) et le potassium (K+). Les principaux anions sont les carbonates (CO3 2−), les bicarbonates (HCO3−), les sulfates (SO4 2−) et les chlorures (Cl−). Certains ions, comme le sodium, le magnésium ou les chlorures, sont appelés éléments conservatifs car leur proportion reste relativement constante avec la salinité de l’eau.
Ces minéraux proviennent principalement de l’érosion et de la dissolution des roches lorsque l’eau circule dans les sols et les formations géologiques. Les cycles biogéochimiques participent également au recyclage d’éléments importants comme le soufre, l’azote, le phosphore et le silicium.
Cycle du soufre : sources, formes (SO₄²⁻ vs H₂S), et ce que change l’anoxie (précipitation avec le fer)
Les sulfates (SO4 2−) présents dans l’eau proviennent principalement de la dissolution des roches contenant du sulfate, notamment le gypse (CaSO4). Ils peuvent aussi provenir des épandages de fertilisants agricoles ainsi que des dépôts et précipitations atmosphériques. Ces apports atmosphériques peuvent être naturels ou liés aux activités humaines, notamment à la combustion des énergies fossiles.
Les sulfates peuvent également être produits par l’oxydation des sulfures présents dans certaines roches. Par exemple, l’oxydation de la pyrite (FeS2) en présence d’oxygène et d’eau produit du sulfate de fer et libère des ions H+, ce qui acidifie le milieu : FeS2 + 3/2 O2 + H2O → FeSO4 + H+
Cette réaction entraîne une diminution du pH et contribue au phénomène de drainage minier acide.
Le trioxyde de soufre (SO3) peut réagir avec l’eau atmosphérique pour former des composés acides. Cette hydratation participe à la formation des pluies acides et provoque une diminution du pH des eaux et des sols.
Dans les milieux aquatiques, les sulfates peuvent être utilisés par certaines bactéries en conditions anaérobies. Lors de la décomposition de la matière organique en absence d’oxygène, les bactéries réduisent les sulfates en sulfures ou en sulfure d’hydrogène (H2S).
Le sulfure formé peut ensuite réagir avec le fer ferreux (Fe2+) pour produire des sulfures métalliques insolubles comme le sulfure de fer (FeS), qui se déposent dans les sédiments.
Le cycle du soufre est qualifié de cycle ouvert car une partie du soufre quitte définitivement le système aquatique sous forme de sulfures métalliques déposés dans les sédiments. Ces pertes empêchent le recyclage complet du soufre dans le lac.
Quelles sont les principales sources d’azote dans les milieux aquatiques ?
L’azote présent dans les lacs et les eaux de surface provient de sources naturelles et anthropiques. Parmi les sources naturelles, la plus importante est la fixation de l’azote atmosphérique (N2) par les cyanobactéries. Ces organismes possèdent des hétérocystes, des cellules spécialisées permettant l’action d’enzymes fonctionnant uniquement en absence d’oxygène. Cette fixation peut représenter jusqu’à 50 % des nitrates (NO3−) entrant dans un lac.
L’azote peut également provenir des dépôts atmosphériques liés aux éclairs, qui produisent des oxydes d’azote (NOx−) et de l’acide nitrique. Les apports du bassin versant jouent aussi un rôle important grâce à l’érosion des roches et au transport de matière organique azotée. Les sources anthropiques proviennent principalement des engrais agricoles et des effluents urbains, qui enrichissent les eaux en nitrates et autres composés azotés.
L’azote est rarement limitant dans les lacs, sauf parfois dans l’épilimnion ou dans certains lacs tropicaux. Cela signifie qu’il est généralement disponible en quantité suffisante pour permettre la croissance des organismes producteurs primaires.
Les producteurs primaires utilisent préférentiellement l’azote sous forme d’ammonium (NH4+). Cet ammonium est principalement produit lors de la dégradation de la matière organique et, dans une moindre mesure, par l’excrétion des animaux. Les nitrates (NO3−) peuvent également être utilisés, mais cette utilisation est moins avantageuse énergétiquement car les nitrates doivent d’abord être réduits grâce à l’enzyme nitrate réductase avant d’être assimilés.
La dénitrification est un processus bactérien anaérobie qui transforme les nitrates (NO3−) en nitrites (NO2−), puis en azote gazeux (N2) rejeté dans l’atmosphère : NO3− → NO2− → N2
Ce processus retire donc de l’azote des milieux aquatiques.
L’ANAMMOX correspond à l’oxydation anaérobie de l’ammonium. Certaines bactéries utilisent simultanément l’ammonium (NH4+) et les nitrites (NO2−) pour produire de l’azote gazeux : NH4+ + NO2− → N2
L’ANAMMOX est une voie métabolique microbienne importante du cycle de l’azote. L’énergie chimique libérée par cette réaction est utilisée par les bactéries pour fixer le dioxyde de carbone.
Sous quelles formes le phosphore est-il présent dans l’eau ?
Le phosphore existe sous plusieurs formes. Le phosphore inorganique assimilable est principalement présent sous forme de phosphate : PO4 3−, HPO4 2− et H2PO4−. Ces formes peuvent être directement utilisées par les organismes aquatiques.
Le phosphore peut aussi exister sous forme de phosphore organique dissous (DOP : Dissolved Organic Phosphorus) ou de phosphore organique particulaire (POP : Particulate Organic Phosphorus), contenus dans la matière organique vivante ou morte.
Le phosphore inorganique peut réagir avec certains cations comme le calcium (Ca2+) ou le fer (Fe3+) en conditions oxydantes. Ces réactions forment des composés insolubles qui précipitent dans les sédiments, rendant le phosphore moins disponible pour les organismes.
Le phosphore organique particulaire (POP) et le phosphore organique dissous (DOP) proviennent principalement de la matière organique. Lors de la décomposition et de l’excrétion des organismes, une partie de ce phosphore est reminéralisée et redevient disponible sous forme inorganique assimilable.
Les plantes et les microorganismes peuvent aussi utiliser le phosphore organique dissous grâce à des enzymes appelées phosphatases alcalines, capables de libérer les phosphates assimilables.
Le SRP (Soluble Reactive Phosphorus) correspond au phosphore réactif soluble. Il représente la fraction de phosphore directement disponible pour les producteurs primaires et constitue donc une mesure importante de la fertilité des eaux.
Lorsque la concentration en oxygène est supérieure à 1 mg O2/L, le phosphore reste plus facilement piégé dans les sédiments sous forme de composés insolubles liés notamment au fer. Lorsque la concentration en oxygène devient inférieure à 1 mg O2/L, ces composés peuvent se dissoudre et relarguer du phosphore dans l’eau. Ce phénomène augmente la disponibilité du phosphore et peut favoriser l’eutrophisation des milieux aquatiques.
Quel est le rôle de la silice dans les milieux aquatiques ?
La silice (Si) est un élément important pour certaines algues, en particulier les diatomées. Les diatomées utilisent la silice pour fabriquer leur frustule, c’est-à-dire leur coque rigide siliceuse.
Au printemps, les conditions de lumière et de nutriments favorisent souvent un bloom de diatomées. Durant cette prolifération, les diatomées consomment rapidement la silice dissoute présente dans l’eau pour construire leurs frustules. Lorsque les diatomées meurent, leurs frustules siliceux sédimentent rapidement vers le fond des lacs. Le recyclage de cette silice est relativement lent, ce qui entraîne progressivement une diminution de la silice disponible dans les eaux de surface. La silice devient alors un facteur limitant pour les diatomées.
Quand la silice devient insuffisante, la croissance des diatomées diminue. D’autres groupes d’algues qui n’ont pas besoin de silice prennent alors le relais durant l’été, comme certaines algues vertes ou cyanobactéries.
Quels sont les différents états trophiques des lacs ?
Les lacs sont classés selon leur richesse en nutriments et leur productivité biologique :
oligotrophe : eau pauvre en nutriments, claire et bien oxygénée ;
mésotrophe : état intermédiaire avec une productivité modérée ;
eutrophe : eau riche en nutriments avec forte production d’algues ;
hypertrophe : enrichissement extrême avec blooms algaux importants.
Lorsque les apports en nutriments augmentent, notamment en phosphore et en azote, la production d’algues devient excessive et provoque des blooms algaux. Après leur mort, la décomposition de cette matière organique consomme beaucoup d’oxygène. Cela peut entraîner une diminution importante de l’oxygène dissous, voire une anoxie dans les couches profondes des lacs.