Comparative Animal Physiology Exam 3
1/104
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
105 Terms
What is our best current hypothesis for how musculature evolved?
Hypotheses about the evolution of muscle are based on characters of extant animals
• Debate about how major clades of animals are related to one another have complicated this, but most studies over the past 10 years suggest that, like neurons, muscles either have 2 independent evolutionary origins or have been lost twice
Muscle
tissue that consists of contractile cells
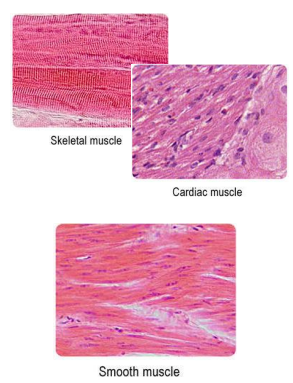
Types of muscle
• In muscle cells, forces of contraction are the result of interactions between contractile proteins myosin & actin
• Striated muscle fibers have bands of myosin & actin organized into repeating units called sarcomeres.
• Arthropods:
• Muscle attached to the exoskeleton, associated with the heart, & along the digestive tract are all striated
• Vertebrates:
• Skeletal muscle – striated muscle attached to bone
• Cardiac muscle – striated muscle of the heart
• Smooth (un-striated) muscle fibers have myosin and actin, but they are not arranged into sarcomeres.
• Vertebrates:
• Found in tubular organs (digestive tract, uterus, etc.)
Overview of vertebrate muscle cell types
2 types of filaments (polarized protein polymers) compose myofibrils:
• Thick filaments – (red) high in myosin & anchored to M lines
• Thin filaments – (teal) high in actin & anchored to Z discs
Z discs
dense areas that line-up across myofibrils within the same fiber, giving a striated appearance to skeletal muscle
Sarcomere
portion of the myofibril between two Z discs
Sarcolemma
membrane around the skeletal muscle fiber
Sarcoplasmic reticulum
branching tubules that surround all the myofibrils; membranes contain Ca2+ - ATPase activetransport pumps that keep [Ca2+] inside the SR > [Ca2+] in the cytoplasm
Transverse (T) tubule
runs transversely across myofibrils in an invagination, closely associated with the sarcoplasmic reticulum (SR)
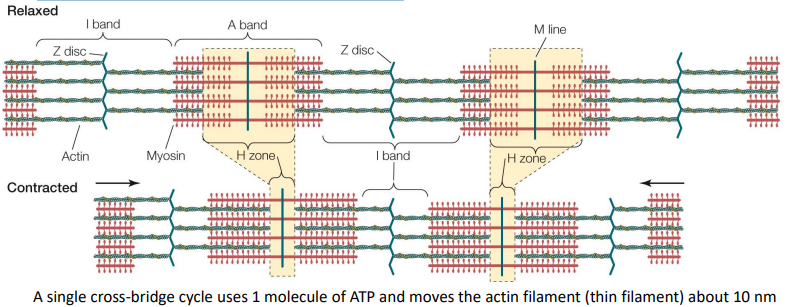
Sliding filament theory of muscle contraction
Thick (myosin) & thin (actin) filaments within muscle fibers pull past one another during muscle contraction and expansion
Vertebrate skeletal muscle contraction first 3 steps
Action potential in motor neuron stimulates the release of acetylcholine (ACh) into the neuromuscular synapse
ACh binds to ligand-gated Na+ channels in the sarcolemma
Action potential travels down into the t-tubule
Vertebrate skeletal muscle contraction steps 4-6
When the action potential reaches DHPRs in the t-tubules, it triggers the release of Ca2+
On the thin filaments (teal), Ca2+ molecules bind to troponin, allowing cross bridges to form between the thick and thin filaments, pulling the thin filaments in the opposite direction as the thick filaments
Each cross bridge goes through several cycles (costing 1 ATP each) during one contraction, if Ca2+ is present. - That moves the thin filament about 10 nm per ATP molecule
Vertebrate skeletal muscle contraction steps 7-9
Synaptic Ach is broken down, terminating the action potential
DHPR returns to previous state & Ca2+ is pumped back into the SR
This ends the contraction
Whole skeletal muscles
• Skeletal muscles have intramuscular connective tissues composed of collagen
• Serve 2 main functions:
• structural organization
• transmit contraction force to the skeletal system
• Intramuscular connective tissues weave themselves into tendons, which connect muscles to skeletal elements
• Skeletal muscles can only exert pulling force
• They cannot lengthen under their own power
• They often function in antagonistic pairs that, (for vertebrates) pull on opposite sides of skeletal joints
• When one muscle shortens, its antagonist partner lengthens & visa versa
• This ensures that muscles can be relengthened after being contracted
• The same thing is true for arthropod muscles, except that they deform the jointed exoskeleton by pulling from the inside
Isometric contraction
cross-bridge cycling occurs, but the whole muscle stays the same length as it contracts
• Help animals maintain steady postures as they stand still
• Muscles actively spend energy, even though they are not shortening (ex. muscles keeping your head up right now)
concentric contractions
whole muscle shortens as it contracts
eccentric contractions
whole muscle resists lengthening as it contracts
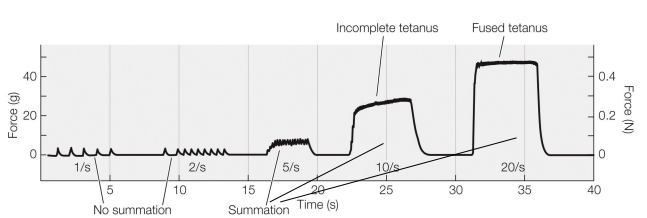
Muscle twitches on a graph (memorize for final)
• Twitch - the mechanical response of a muscle to a single action potential
• Very little force is transmitted to the skeletal structure from a single twitch
• Action potentials take 2 ms, but twitches are >100 ms in duration
• Summation – the fusion of multiple twitches into a single larger contractile force
• A tetanic contraction (tetanus) is a strong, sustained muscle contraction that occurs as a result of summation
Slow oxidative (SO) fibers
small crosssectional area, many mitochondria, many capillaries
• specialize on aerobic respiration
Fast glycolytic (FG) fibers
large crosssectional area, few mitochondria, few capillaries
• specialize on anaerobic respiration
Fast oxidative glycolytic (FOG) fibers
intermediate diameter, many mitochondria, many capillaries
Characteristics of mammalian skeletal twitch muscle fibers
Duration of twitches:
Slow oxidative (SO) - long
Fast oxidative glycolytic (FOG) - short
Fast glycolytic (FG) - short
Diameter of fiber:
Slow oxidative (SO) - small
Fast oxidative glycolytic (FOG) - intermediate
Fast glycolytic (FG) - large
Levels of glycolytic enzymes:
Slow oxidative (SO) - low
Fast oxidative glycolytic (FOG) - intermediate
Fast glycolytic (FG) - high
Ability to produce ATP using oxidative phosphorylation:
Slow oxidative (SO) - high
Fast oxidative glycolytic (FOG) - high
Fast glycolytic (FG) - low
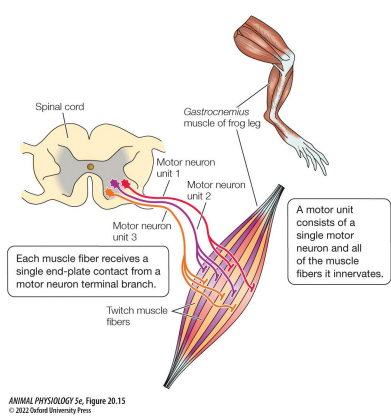
Motor unit
A motor neuron and all the muscle fibers it innervates
Vertebrates neural control
• Each muscle fiber is innervated by 100s – 1000s of motor neuron dendrites
• Each muscle fiber is innervated by 1 motor neuron
• Motor units are distinct
• Muscle fibers are only innervated by excitatory motor neurons
• Excitatory neurotransmitter is ACh
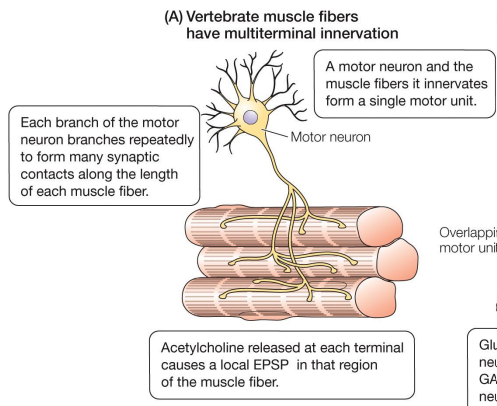
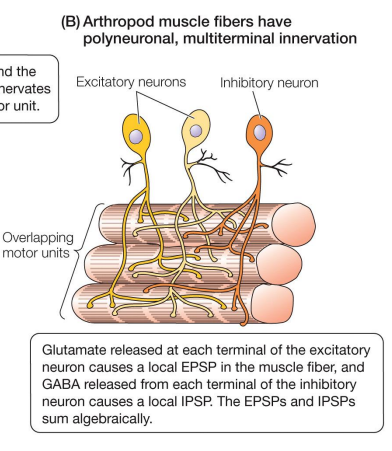
Arthropods neural control
• Each muscle innervated by 1 – 10 motor neuron dendrites
• Each muscle fiber is innervated by many motor neurons
• Motor units overlap
• Each muscle fiber can be innervated by both excitatory and inhibitory motor neurons, allowing for peripheral inhibition
• Excitatory neurotransmitter is glutamate & inhibitory neurotransmitter is GABA (gamma-aminobutyric acid)
Arthropod skeletal muscle
• Arthropods don’t have SO and FG muscle fiber types, they have short sarcomere vs. long sarcomere fibers.
• Short sarcomere muscle fibers contract quickly and long sarcomere muscle fibers contract slowly.
• Most arthropod muscles contain a mixture of long- and short-sarcomere fibers, with some exceptions:
• Flight muscles in Drosophilla are longsarcomere biased
• Muscles in the tails of crawfish are loaded with short sarcomere fibers to allow for sudden & rapid escape.
Vertebrate smooth (unstriated) muscle
• Gastrointestinal, respiratory, reproductive, urinary, and cardiovascular systems, iris of the eye, base of hairs & feathers (vertebrates)
• Single nucleus, no transverse tubules
• Contain actin and myosin, not in sarcomeres
• > thin/thick filaments (actin /myosin)
• Thin actin filaments attach to dense bodies in the cytoplasm (not Z plates)
• Filaments in bundles around the cell’s periphery
• Shorter than most skeletal muscles
• Innervated by the autonomic nervous system
• Smooth muscle hydrolyzes ATP more slowly
• Can contract slowly, for longer periods of time
• Smooth muscle of the esophagus remains contracted except in the presence of food
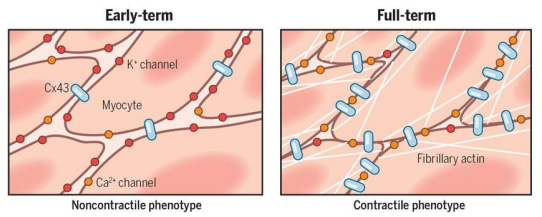
Vertebrate smooth (unstriated) muscle uterine lining
• Mammalian uterus switches between singe- & multi-unit smooth muscle
• Only during late stages of pregnancy when estrogen is highest, the uterine smooth muscle cells form gap junctions that couple adjacent cells
• This allows the uterus to produce coordinated contractions that respond well to local stretch cues.
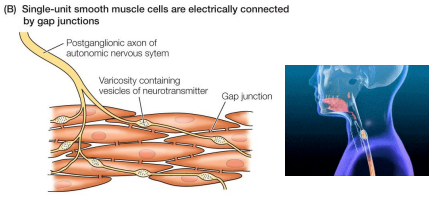
Single-unit smooth muscle
• Muscle cells are connected (electrically coupled) by gap junctions
• Depolarize and contract together, functioning as a single unit
• Less direct neural control (less innervation from ANS)
• Can be activated by stretch
• Ex: gastrointestinal tract, small blood vessels
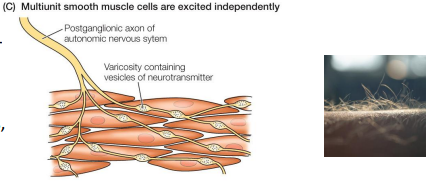
Multi-unit smooth muscle
• Few, if any, gap junctions
• Function more independently from each other
• More direct neural control (more innervation from ANS)
• Not stretch-sensitive
• Ex: hair & feather erectors, eyes, large arteries, respiratory airways
Vertebrate cardiac muscle (its own category)
• Myofibrils are organized into sarcomeres (striated)
• Cells are branched, not straight
• Each cell has a single nucleus
• Intercalated discs (red arrow)
• gap-junction rich
• allows force of contraction to be transferred across cells
• ensures that cells beat nearly synchronously
• controlled by autonomic nervous system
• Local pacemaker cells generate coordinated muscular rhythm
• Action potential of cardiac cells is relatively long (100-500ms)
• Ensure a prolonged contraction suitable for pumping blood, rather than a short muscular twitch
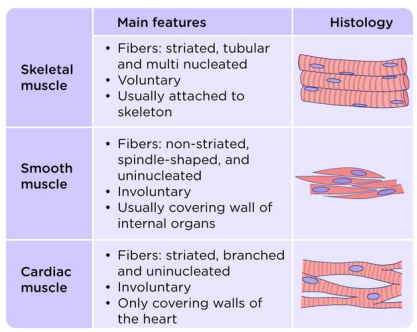
Characteristics of the three major types of muscles in vertebrates
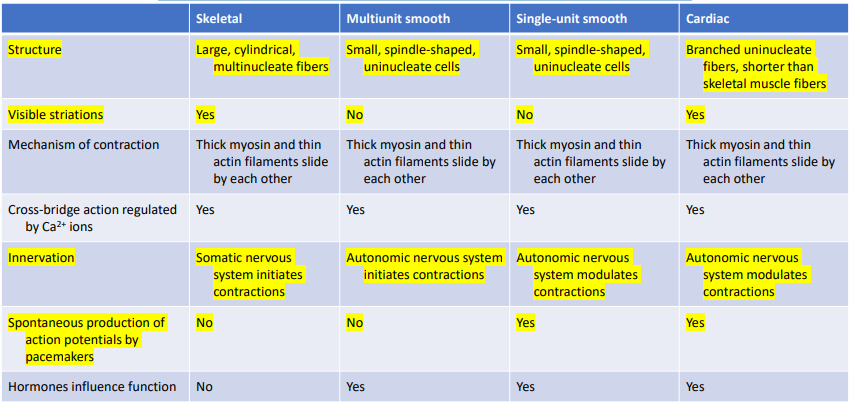
Characteristics of the three major types of muscles in vertebrates
Know why animal bodies need to acquire O2 and void CO2 , & the relative urgencies of each.
They are the primary gasses that are consumed and produced by aerobic catabolism (aka cellular respiration), for instance, see this familiar equation showing glucose metabolism. This is why O2 & CO2 are known as the respiratory gasses.
Why is the need for O2 so much more urgent than for any other material animals exchange with the environment?
The ETC & OXPhos generate the most ATP per molecule of glucose ingested. O2 plays an especially important role as the final electron receptor in the ETC. Not all cells can perform anaerobic respiration (ex. human neural tissue). Those that can must get rid of lactic acid.
The need to void CO2 is important, but not usually as urgent as the need to acquire O2 . • Not voiding enough CO2 can lead to acidification of body fluids & other ill effects, but there are ways the body can deal with this, with a little extra energy expenditure.
O2 & CO2 are the respiratory gasses (involved in cellular respiration) • Transport of O2 & CO2 is usually passive (always passive for O2 ) • Small molecules like these can move by diffusion across membranes • Animals have many adaptations that enable this passive transport across barriers throughout the body
Learn the difference between convective transport (bulk flow) and simple diffusion, and know where each is used in respiratory transport.
Simple diffusion - same mechanisms as solutes diffusing through solutions Has two important rules: 1. Solutes diffuse from areas of high “concentration” to areas of low “concentration” 2. Rate of diffusion is proportional to the difference in concentration
Convective transport (bulk flow) – gases are moved from place to place due to the flow of a fluid into which they are dissolved (ex. in blood as it flows through vessels)
Understand the concept of partial pressures, how they differ from concentration, and be able to calculate them.
Up until now we have used concentration gradients as a proxy for chemical potential, because we’ve been talking about diffusion from one aqueous environment to another • However, this doesn’t work when discussing diffusion from one phase solution to another (ex. O2 in the air diffusing into the blood) • For that reason, we will now be talking about partial pressure gradients instead, a more direct measure of chemical potential. • We notate partial pressure as P with a subscript of the gas name. • For example: 𝑃𝑂2 is the partial pressure of O2
Know what factors affect the rate of respiratory gas diffusion.
Partial pressure of each gas in a mixture is independent of the other gasses in the mixture. • The total pressure (PT ) exerted on the container (in units of Pa or atm) is the sum of the partial pressures of all gases (gas A, gas B, gas C) in the mixture. “Dalton’s Law”: 𝑃𝑇 = 𝑃𝐴 + 𝑃𝐵 + 𝑃�
How do you calculate pressure? • One way is to use the ideal gas law 𝑃𝑉 = 𝑛 𝑅𝑇 • If you know the mass of each gas, the volume of the container, and the temperature: 𝑃 = 𝑛𝑅𝑇 𝑉 P = pressure (Pa) V = volume (m3 ) n = number of molecules of that gas, in units of moles R = universal gas constant (~8.314 m3 ⋅Pa⋅K −1⋅mol−1) T = temperature (K) Gas mixtures One mole contains exactly 6.022×1023 elementary entities, which can be atoms, molecules, ions, or other particles. In these examples, we’re usually going to be talking about molecules.

A glass bottle contains 1 mole of Nitrogen (N2 ), 0.5 moles of Oxygen (O2 ), and 0.25 moles of Argon (Ar). Together, this mixture exerts a total pressure of 8 Pa on the bottle. What are the partial pressures of Nitrogen, Oxygen, and Argon?
Here are the equations you’ll need: 𝑃𝑇 = 𝑃𝐴 + 𝑃𝐵 + 𝑃𝐶 1 = 𝑋𝐴 + 𝑋𝐵 + 𝑋𝐶 𝑋𝐴 = 𝑛𝐴 𝑛𝑇 𝑃𝐴 = 𝑋𝐴𝑃𝑇 PA = partial pressure (Pa) of gas “A” PT = Total pressure nA = moles of gas A nT = total moles of all gasses in the container XA = molar ratio of gas A Answer: PN2 = ~ 4.568 Pa PO2 = ~ 2.288 Pa PAr = ~1.143 Pa
A glass bottle contains 0.625 moles of Nitrogen (N2 ), 0.25 moles of Oxygen (O2 ), and 0.125 moles of Argon (Ar). Together, this mixture exerts a total pressure of 8 Pa on the bottle. What are the partial pressures of Nitrogen, Oxygen, and Argon?
Here are the equations you’ll need: 𝑃𝑇 = 𝑃𝐴 + 𝑃𝐵 + 𝑃𝐶 1 = 𝑋𝐴 + 𝑋𝐵 + 𝑋𝐶 𝑋𝐴 = 𝑛𝐴 𝑛𝑇 𝑃𝐴 = 𝑋𝐴𝑃𝑇 PA = partial pressure (Pa) of gas “A” PT = Total pressure nA = moles of gas A nT = total moles of all gasses in the container XA = molar ratio of gas A
Gas in aqueous environments
Gases such as O2 can exist in aqueous solutions, not as bubbles, but as molecules interspersed among molecules of water, also known as “dissolved” gases • This is not the same as air bubbles
The partial pressure of a gas dissolved in an aqueous solution is equal to the partial pressure of the same gas in a gas phase at equilibrium.
Partial pressure and concentration in aqueous environments are proportional, but the absorption coefficient varies greatly and depends on temperature and salinity. • Cold water can “hold” (absorb) more dissolved O2 than warm water • Fresh water “holds” (absorbs) more dissolved CO2 than salt water
Diffusion of gases
One of the first places you see inward diffusion of O2 happening is at the interface between the organism’s respiratory surface and the fluid environment • Two rules: 1. Diffuse down partial pressure gradients 2. Difference in partial pressure drives diffusion rate
The rate of gas diffusion is directly proportional to the size of the partial pressure gradient. • The equation for the rate of gas diffusion: 𝐽 = 𝐾 𝑃1 −𝑃2 𝑋 J = rate of gas movement P1 = relatively higher partial pressure P2 = relatively lower partial pressure X = distance between them K = Krogh diffusion coefficient (depends on several factors) • For O2 , K is about 200,000 times greater in air than in water. • For CO2 , K is about 9,000 times greater in air than in water.
Convective transport of gases: Bulk flow
• Besides diffusion, this is the other major mechanism of respiratory gas transport • Far faster than diffusion • More effective for long distance transport • Costs metabolic energy • Two major types in animal bodies: • Breathing (moving gas mixtures around) • Pumping of body fluids (ex. blood flow)
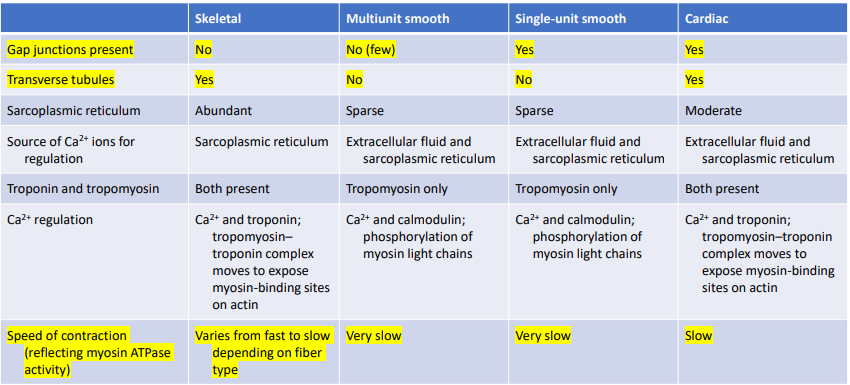
Learn the difference between unidirectional flow and tidal flow.
Unidirectional flow – occurs through a tube • Ex. flow of blood through a blood vessel 2. Tidal flow – back and forth flow; occurs in cavities with a single opening for entrance & exit • Ex. flow of air in mammalian lungs
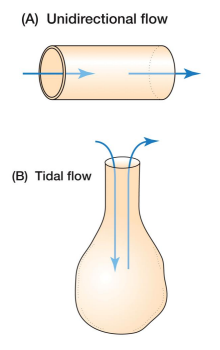
Be able to explain the concept of an oxygen cascade.
The transfer of oxygen from the environment to the mitochondria can be thought of as an oxygen cascade, where potential energy is incrementally lost. • This is like a cascade losing potential energy in the form of height over its length. • In the case of O2 transport, this takes the form of incremental decreases in the partial pressure of O2 . • At each step, O2 is diffused at a rate dependent on the difference in partial pressures. • Like a waterwheel, there is a minimum chemical potential needed to transport enough O2 for mitochondrial function. • In humans, mitochondrial 𝑃𝑂2must be > 0.001 atm to support aerobic respiration. • Hence, systemic blood must stay > 0.09 atm
Be able to give 2 reasons why it is more difficult for water-breathing animals to extract oxygen from their environments than air-breathing animals.
Water is denser and more viscous than air, so water-breathers must expend more energy to move water through their bodies via bulk flow than airbreathers spend to move air via bulk flow.
O2 is also less soluble in water compared to air, so air can “hold” more molecules of at any given temperature. • Due to its salinity, sea water is even less soluble to O2 than freshwater. • For all of them, solubility of O2 decreases as temperature increases.
Respiration
exchange of respiratory gasses (O2 & CO2 ) • Can be internal (moving them around in the body) or external.
External respiration
aka “breathing”; the process by which an animal gains O2 from its environment, & voids CO2 into its environment (exchange between outer fluid & respiratory exchange membrane)
Dual breather or bimodal breather
can breathe both air & water
Respiratory-exchange (or gas-exchange) membrane
1 to 2 layers of epithelial cells that are where external respiration occurs
Ventilation
forced flow (convection) of air or water into & out of structures/surfaces used for external respiration • Not all animals ventilate!
Fundamental concepts of external respiration
O2 always via diffusion • CO2 usually via diffusion, but transported actively (as HCO2 ) in some freshwater animals • In some animals the whole body surface is a gas-exchange membrane. • Other animals have specialized breathing surfaces/structures where this occurs
Types of specialized breathing structures:
Lungs – invaginated into the body & contain environmental medium • Gills – evaginated into the environment & surrounded by environmental medium • external gills – not in a superficial body cavity • internal gills – in a superficial body cavity • Requires ventilation (costs ATP) • Enables better canalization
Ventilation
the forced flow (convection) of air or water into & out of structures/surfaces used for external respiration
Passive ventilation
current is environmental • does not cost metabolic energy (ATP) • current is less reliable
Active ventilation
animal creates the current • costs metabolic energy (ATP) • current is more reliable
Types of active ventilation
Nondirectional – fluid flows across the gas-exchange membrane in many directions • Bidirectional – fluid flows to & from the gas-exchange membrane via the same passages • This is also known as tidal gas exchange • Unidirectional – fluid is pumped over the gas-exchange membrane in a one-way path • Co-current gas exchange (also known as concurrent gas exchange) • Countercurrent gas exchange – most fish have this • Cross-current gas exchange – birds have this
Be able to calculate the O2 uptake of a breathing organ & oxygen extraction efficiency.
The rate of O2 uptake by a breathing organ is affected by: 1. The flow of the medium (volume/time) 2. The amount of O2 removed from each unit of volume 𝑅𝑎𝑡𝑒 𝑜𝑓 𝑂2 𝑢𝑝𝑡𝑎𝑘𝑒 𝑚𝐿 𝑚𝑖𝑛 = 𝑉𝑚𝑒𝑑𝑖𝑢𝑚(𝐶𝐼 − 𝐶𝐸) 𝑉𝑚𝑒𝑑𝑖𝑢𝑚 = “rate of ventilation ” the rate air or water flows through the breathing organ (L/min) 𝐶𝐼= O2 concentration of the inhaled medium (mL / L) 𝐶𝐸= O2 concentration of the exhaled medium (mL / L)

Water entering the mouth of a fish contains 6 mL O2 /L. Water exiting the gills contains 4 mL O2 /L. The rate of ventilation is 0.5 L/min. What is the rate of O2 uptake?
𝑅𝑎𝑡𝑒 𝑜𝑓 𝑂2 𝑢𝑝𝑡𝑎𝑘𝑒 = 𝑉𝑚𝑒𝑑𝑖𝑢𝑚(𝐶𝐼 − 𝐶𝐸)
𝑅𝑎𝑡𝑒 𝑜𝑓 𝑂2 𝑢𝑝𝑡𝑎𝑘𝑒 = 0.5 (6 − 4)
𝑅𝑎𝑡𝑒 𝑜𝑓 𝑂2 𝑢𝑝𝑡𝑎𝑘𝑒 = 1 𝑚𝐿 𝑂2/𝑚𝑖𝑛
𝑉𝑚𝑒𝑑𝑖𝑢𝑚 = rate air or water flows through the breathing organ (L/min)
𝐶𝐼= O2 concentration of the inhaled medium (mL / L)
𝐶𝐸= O2 concentration of the exhaled medium (mL / L)
oxygen utilization coefficient (aka oxygen extraction coefficient)
an indication of how efficiently an animal uses the O2 in the fluid pumped through a respiratory organ. It is expressed as a percentage of the originally available O2 that was present in the medium upon inhalation.
𝑂𝑥𝑦𝑔𝑒𝑛 𝑢𝑡𝑖𝑙𝑖𝑧𝑎𝑡𝑖𝑜𝑛 𝑐𝑜𝑒𝑓𝑓𝑖𝑐𝑖𝑒𝑛𝑡 = 100(𝐶𝐼 − 𝐶𝐸)/𝐶𝐼
𝐶𝐼= O2 concentration of the inhaled medium (mL / L)
𝐶𝐸= O2 concentration of the exhaled medium (mL / L)
Water entering the mouth of a fish contains 6 mL O2 /L. Water exiting the gills contains 4 mL O2 /L. The rate of ventilation is 0.5 L/min. What is the oxygen utilization coefficient?
𝑂𝑥𝑦𝑔𝑒𝑛 𝑢𝑡𝑖𝑙𝑖𝑧𝑎𝑡𝑖𝑜𝑛 𝑐𝑜𝑒𝑓𝑓𝑖𝑐𝑖𝑒𝑛𝑡 = 100(𝐶𝐼 − 𝐶𝐸)/𝐶𝐼
𝑂𝑥𝑦𝑔𝑒𝑛 𝑢𝑡𝑖𝑙𝑖𝑧𝑎𝑡𝑖𝑜𝑛 𝑐𝑜𝑒𝑓𝑓𝑖𝑐𝑖𝑒𝑛𝑡 = 100(6 − 4)/6
𝑂𝑥𝑦𝑔𝑒𝑛 𝑢𝑡𝑖𝑙𝑖𝑧𝑎𝑡𝑖𝑜𝑛 𝑐𝑜𝑒𝑓𝑓𝑖𝑐𝑖𝑒𝑛𝑡 = 33%
A cat ventilates 1.2 L of air every minute. Air entering the cat’s nostrils contains 6 mL O2 /L. Air exiting the nostrils contains 3 mL O2 /L. What is the oxygen utilization coefficient and the rate of O2 uptake?
𝑂𝑥𝑦𝑔𝑒𝑛 𝑢𝑡𝑖𝑙𝑖𝑧𝑎𝑡𝑖𝑜𝑛 𝑐𝑜𝑒𝑓𝑓𝑖𝑐𝑖𝑒𝑛𝑡 = 100(𝐶𝐼 − 𝐶𝐸)/𝐶𝐼
𝑅𝑎𝑡𝑒 𝑜𝑓 𝑂2 𝑢𝑝𝑡𝑎𝑘𝑒 = 𝑉𝑚𝑒𝑑𝑖𝑢𝑚(𝐶𝐼 − 𝐶𝐸)
𝑉𝑚𝑒𝑑𝑖𝑢𝑚 = rate air or water flows through the breathing organ (L/min)
𝐶𝐼= O2 concentration of the inhaled medium (mL / L)
𝐶𝐸= O2 concentration of the exhaled medium (mL / L)
Tidal gas exchange (bidirectional)
Breathing organ is never completely emptied between breaths • Fresh & stale media mix. • The O2 partial pressure of medium in contact with the gas exchange membrane is therefore often much lower than the partial pressure of the inhaled medium
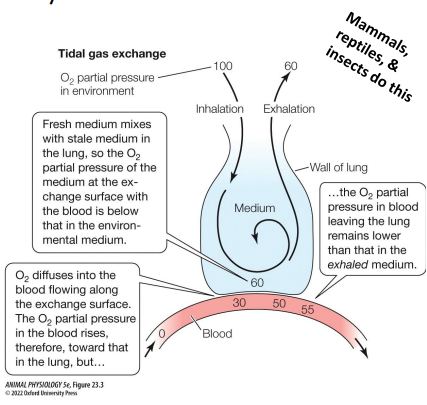
Unidirectional ventilation
Cocurrent (or concurrent) gas exchange – medium & blood flow in the same direction • Countercurrent gas exchange – medium & blood flow in opposite directions
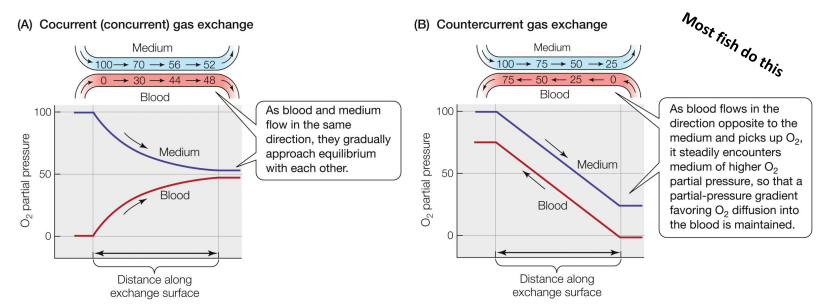
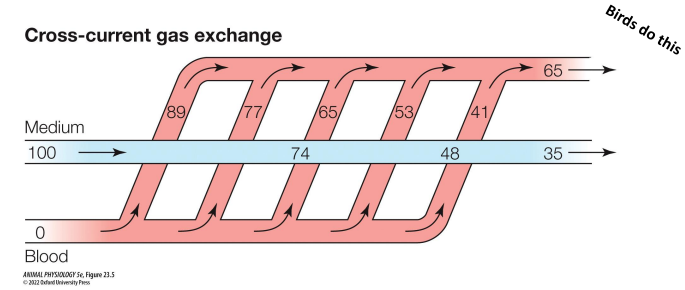
Cross-current gas exchange
medium splits into multiple vessels that cross the path of the medium
Relative efficacy of each active ventilation strategy:
Countercurrent gas exchange (1st place) • Cross-current gas exchange (2nd place) • Tidal gas exchange (3rd place tie) • Cocurrent gas exchange (3rd place tie)
Rhythmogenesis
the generation of motor neuron impulses that stimulate rhythmic muscular contractions driving active ventilation
Breathing by fish
Many young fish respire exclusively via diffusion across the skin • Large-bodied adults use other strategies • Countercurrent gas exchange is the norm in most modern fishes, such as in this teleost • Cocurrent gas exchange is thought to be the ancestral state
Breathing by teleost (bony) fish
Tuna are especially efficient, with oxygen extraction coefficients exceeding 60%, double that of many other bony fish • Countercurrent gas exchange • Internal gills • Obligate ram ventilators • Ram ventilation – mouth is opened and water is forced over the gills by forward movement • By contrast, many other fish only use ram ventilation when they are swimming fast enough. Tuna have no choice.
Another key feature enabling tuna to use ram ventilation to efficiently remove O2 from water: • Reinforced gills stabilize lamellae in high velocity waterflow
Breathing in teleost fish (most bony fish)
Instead of relying exclusively on ram ventilation, most other bony fish pump water over their gills • Gill ventilation is usually driven by buccal-opercular pumping. • Buccal pressure pump – positive pressure generated in the buccal cavity pushes water over gill surfaces • Opercular suction pump – negative pressure generated in the opercular cavity sucks water over gill surfaces • Water flows from areas of high pressure to those of low pressure. • Breathing cycle alternates between them to generate nearly continuous unidirectional flow
Breathing: lungfish
~400 spp. of fish have an air-breathing organ derived from various organs • Lungfish (6 spp.) are among our closest living fish relatives • Intensely in-folded, high-surface-area lungs (specialized swim bladders) • Lungfish have both gills & lungs • Evolutionarily, gill areas reduce in size as lineages of fish evolve to be more heavily reliant on air-breathing. • High O2 in the blood can be lost over gill surfaces to relatively O2 -poor water
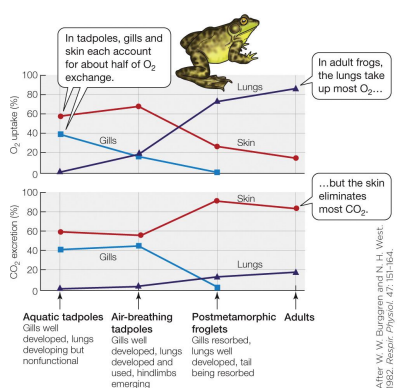
Breathing: amphibians
Can change between water & air breathing over time / during development • Many remain dual breathers as adults (breathing air & water) • The lung of a frog is a single sac with lots of pockets. • Gas exchange can occur over gills, skin, lungs, & the buccal cavity
Pulmonary breathing cycle of a bullfrog
Gasps air into buccal cavity 2. The glottis opens and exhaled air leaves the lungs and exits the body 3. Air in the buccal cavity is pushed into the lungs.
Which site does most of the gas exchange changes developmentally. Though lungs become increasingly important for O2 exchange, most voiding of CO2 is done by the skin.
Breathing by birds
Use cross-current gas exchange. • More efficient than mammalian • In addition to lungs, birds have air sacs that pump air unidirectionally across the lungs • Takes 2 breaths for air to enter & leave: • First inhalation: air sacs expand. Air drawn into posterior sacs • First exhalation: air sacs compress. Air pushed into lungs • Second inhalation: air sacs expand. Air in lungs drawn into anterior sacs. • Second exhalation: air sacs compress. Air is exhaled
Use cross-current gas exchange. • More efficient than mammalian • In addition to lungs, birds have air sacs that pump air unidirectionally across the lungs • Takes 2 breaths for air to enter & leave: • First inhalation: air sacs expand. Air drawn into posterior sacs • First exhalation: air sacs compress. Air pushed into lungs • Second inhalation: air sacs expand. Air in lungs drawn into anterior sacs. • Second exhalation: air sacs compress. Air is exhaled
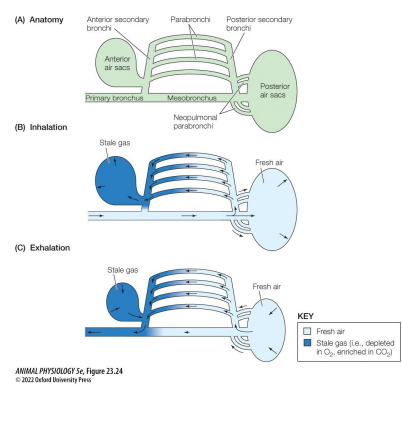
Breathing by birds pt 2
Main site of gas exchange is across the thin, vascularized walls of the parabronchi within the lungs • Bird lungs are hard and rigid, they don’t expand & contract! • Flow is posterior to anterior within the lungs
Breathing by mammals
Mammals & birds are endotherms with high metabolic rates, which demands a high O2 supply, but we also breathe air rather than water, which is easier • tidal flow a.k.a. bidirectional flow • Alveolus “hollow cavity“ – sac-like site of gas exchange • ~500 million alveoli (each 0.25 mm) in the human lung. • In total 130 m2 of area! • Pulmonary surfactants – mostly phospholipid mixture that alter surface tension in the alveoli, preventing their collapse; synthesized by epithelial cells • Trachea, bronchi, & bronchioles are conducting airways (not involved in gas exchange). • The respiratory airways of lungs are the places where gas exchange occurs (respiratory bronchioles, the alveolar ducts, & alveolar sacs, in mammals).
When mammals breathe (active ventilation), they do not come close to completely filling or completely voiding their lungs of air • The tidal volume is the volume of air inhaled & exhaled per breath. • In humans at rest, lung vol at the end of inhalation is 2900 mL, volume at the end of exhalation is 2400 mL. So resting tidal volume is 500 mL.

Expiratory reserve volume
vol of air that can be expelled beyond resting exhalation
Inspiratory reserve volume
vol of air that can be inhaled beyond resting inhalation
Vital capacity
sum of resting tidal volume, expiratory reserve volume, and inspiratory reserve volume
Breathing by mammals pt 2
Contraction of the diaphragm creates negative pressure • This pulls air into the lungs • Ventilatory rhythm is set by the PreBötzinger complex, a cluster of ventilatory neurons in ventrolateral medulla of the brainstem. • Low blood O2 is detected by the aortic bodies & carotid bodies. These relay info to the brain stem. • Controls based on sensing excess CO2 are most potent. When CO2 rises H+ does too. • Both arterial CO2 levels & acidification of cerebral spinal fluid are monitored. Increases in either increase ventilation
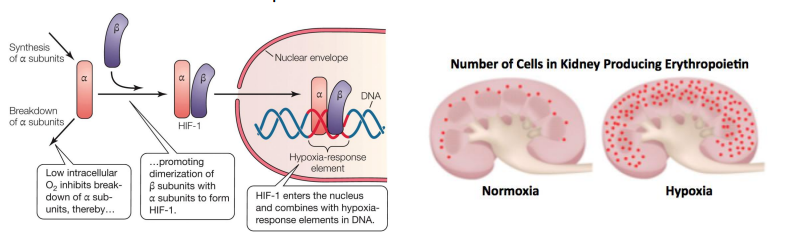
How does an animal cell detect Low O2 ?
Hypoxia inducible factors 1 and 2 (HIF 1 & HIF 2)
Alpha & beta subunits bind in low-O2 scenarios, forming a transcription factor • In the vertebrates this stimulates the production of erythropoietin. • Erythropoietin (EPO) is a hormone produced by the peritubular cells of the kidney, that stimulates red blood cell production.
Breathing by aquatic inverts
• Many aquatic animals exchange respiratory gases across body surfaces (especially small or thin animals)
• Sponges, jellyfish, comb jellies, corals
• No respiratory system
• No circulatory system
• Most metabolically active cells in a jellyfish are near the internal or external surface
• Larger aquatic invertebrates: Convergent evolution of gills
• Either passive ventilation or non-directional active ventilation
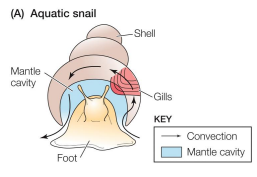
Breathing by aquatic inverts: Aquatic Snail
• gill leaflets
• counter-current gas exchange
• ventilation via cilia
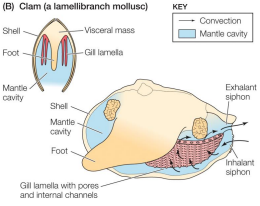
Breathing by aquatic inverts: Clam
• sheetlike gills
• counter-current gas exchange
• Ventilation via cilia
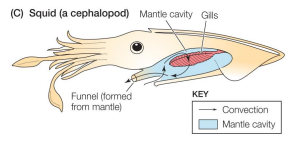
Breathing by aquatic inverts: Squid
• feathery gills
• counter-current gas exchange
• Ventilated by muscular contraction
• High ventilation, remove less O2 from water
Breathing by terrestrial gastropods
• Slugs & land snails
• The walls of the mantle cavity are the site of gas exchange
• Some don’t ventilate
• Some ventilate by raising & lowering the floor of the cavity
• Tidal / bidirectional
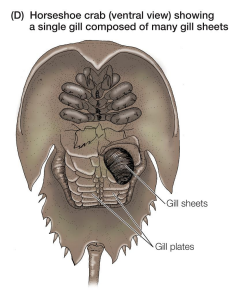
Breathing by aquatic inverts: horseshoe crab
• Book gills – gill structures consisting of many sheets of tissue arrayed like pages of a book
• Respiratory organ of horseshoe crabs
• Rhythmic flapping motion of gill plates ventilates the book gills
• Non-directional ventilation
Breathing air with book lungs (Arachnids)
• Book lungs – lungs within which sheets of gas-exchange tissue alternate with sheetlike air spaces
• like pages of a book: separated by thin layers of air
• Akin to the book gills of horseshoe crabs
• Some are diffusion lungs (no active pumping)
• Some ventilated by pumping motions
• Bidirectional / tidal
Breathing by aquatic inverts: Crustaceans
• Gills arise from base of thoracic legs
• Central axis & many lamellar plates
• Gills covered in an especially thin cuticle (they must molt!!!)
Breathing by tracheate arthropods
• Tracheae – gas-filled, tube-shaped invaginations used by some terrestrial arthropods (including insects)
• Branch into smaller tracheoles
• Bring air directly to the cells in need
• Invaginations of epidermis (they molt!!!)
• Spiracle – site where tracheal system opens to the atmosphere (on the abdomen &/or thorax)
• Hemolymph sometimes lacks O2 transport pigments
• Respiratory system does most of the work
All tissues in insect bodies have tracheae, but the most metabolically active tissues (high ATP use) have especially high “tracheation”
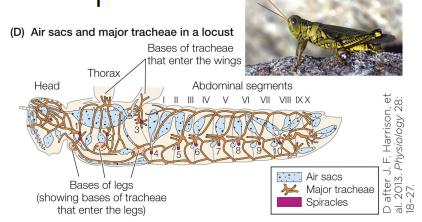
Breathing by tracheate arthropods Pt 2
• Many highly mobile insects can compress air sacs to actively pump air through the tracheal system
• Grasshoppers, bees, etc.
• Usually compressed via muscular contraction
• Often this is visible to the naked eye (abdomen movements)
• More recently, physiologists discovered microscopic ventilation
• rhythmic tracheal compressions are an important mechanism for ventilation in a far wider array of insects.
• Often not visible without x-rays… or a freshlymolted translucent insect such as can be seen in the video to the right.
Breathing by tracheate arthropods Pt 3
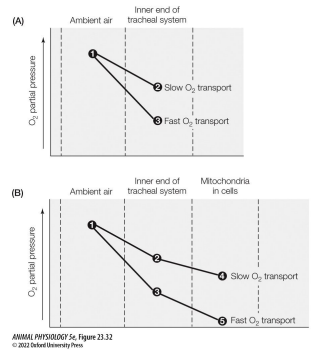
• Though the respiratory systems of terrestrial arthropods are different from those of vertebrates, the same basic principles apply (e.g. O2 cascades)
• O2 homeostasis is aided by higher diffusion occurring when high mitochondrial use of O2 drives intracellular O2 partial pressures down.
• Regulation of the ventilation rate is also important
Respiratory pigments
metalloproteins able to undergo reversible combination with molecular oxygen
• Not all animals have / require respiratory pigments
• All respiratory pigments
• contain metal atoms
• have a distinct color (hence, “pigments”)
• Function of respiratory pigments
• Increase the O2 carrying capacity of blood (or other body fluids)
• Only bind with O2 at specific binding sites
• Combination of O2 & respiratory pigments is “oxygenation”, not “oxidation” • Partial (not complete) transfer of electrons
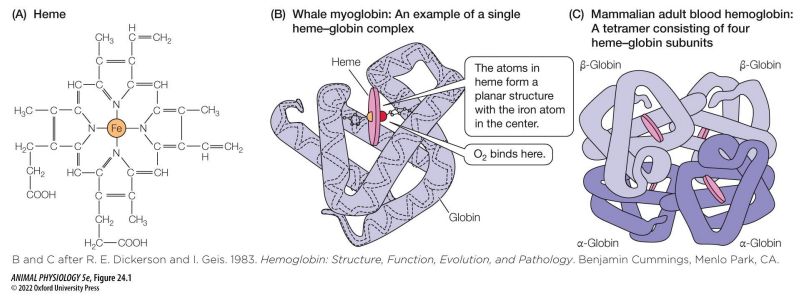
Hemoglobin
• 2 defining characteristics:
1. Contain a heme (a particular metalloprotein with ferrous iron in it)
2. The heme is noncovalently bonded to a globin protein.
• Most widespread respiratory pigment in kingdom Animalia
Hemocyanin
• Copper-based
• Many arthropods & molluscs
Chlorocruorins
• Resemble hemoglobin (iron-based)
• annelids
Hemerythrin
• Iron-based, but lacks heme
• Several invertebrate phyla
Why are respiratory pigments not enzymes?
• They do not undergo conformational change when they bind to O2
• They do not catalyze reactions, they are only carriers
However, due to the rules of kinematics they follow “affinity curves” or “O2 binding curves” resembling the enzyme kinematic curves you learned about earlier in this course. For this reason, they are sometimes referred to as “honorary enzymes”.
O2 binding characteristics of respiratory pigments