NRS 305 Final
1/72
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
73 Terms
population regulation
population size and growth are limited by density dependent and density independent factors
density dependent
increasing population size causes per capita growth rate to decrease. example: limited food leads to competition in population
density independent
affects per capita growth rate regardless of population density. example: natural disasters
whooping cranes
low chick survival is a major limiting factor that keeps small population size from growing. Management strategy: increase chick survival
bighorn sheep
predation matters but varies too much in survival and reproductive rates across locations. Management strategy: must be site specific, some populations unaffected while others are struggling
vancouver marmots
very small population sizes to the point little predation is impactful. Management strategy: reduce all mortality sources, even if they seem minor
white sharks
increasing shark populations are linked to increasing seal populations. Management strategy: population regulation involved intricate food webs, must consider entire ecosystems and not just one species
new england cottontails
high predation rates are a major limiting factor during reintroductions. Management strategy: focus on predator control or habitat managements during reintroduction of population
primary focus on top down
humans are predators and see other predators as competition, predators often pose conservation and management issues (white tail deer overpopulation because no predators), anti-predator adaptations (camouflage, porcupine spikes), predators mainly blamed for declining prey populations
White tail deer in Wisconsin
severe winters, multiple sources of mortality (bears, cars, humans), bottom up controls, wolf and deer populations are coincidental, deer populations increases during more mild winters even when wolves present
dP/dt=caRP-dP
lotka volterra predator model
c
LV, capture efficiency of predation
a
LV, efficiency of converting food into predator growth
R
LV, prey population size
P
LV, predator population size
d
LV, predator death rate
LV predator assumptions
predators only eat one species at a time, predator efficiency does not change with prey abundance
dR/dt=rR-cRP
lotka volterra prey model
r
LV, prey growth rate
LV prey assumptions
prey limited only by predators, predator efficiency does not change with prey abundance
snowshoe hare, canadian lynz
rare cyclic relationship between predator prey populations predicted simple models, predicts boom-bust cycles
Lemming cycles
pattern does not fit simple LV cyclic model, population peaks don’t occur in regular intervals, smooth increases and sudden crashes, populations influenced by multiple factors not just predation
support top down control
if lemmings are mainly controlled by predation, then population shouldn’t crash when predators are excluded from the experiment
support bottom up
if lemmings are mainly controlled by bottom up, lemming survival and reproduction rates are higher when more food is added
predator efficiency
the proportion of the prey population that a predator can successfully capture and consumer per unit time, small amount of prey means high efficiency, large amount of prey means less efficiency
type I functional response
rate of consumption per predator is proportional to prey density (constant slope, no satiation)

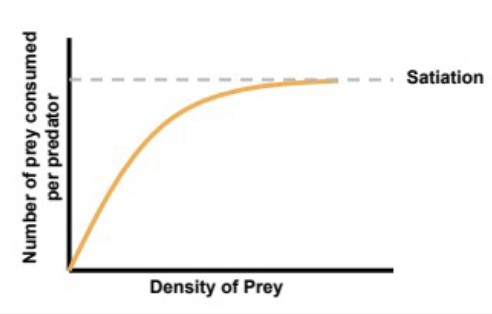
type II functional response
rate of increase in prey consumption per predator starts high, diminished and ultimately plateaus with increasing prey density
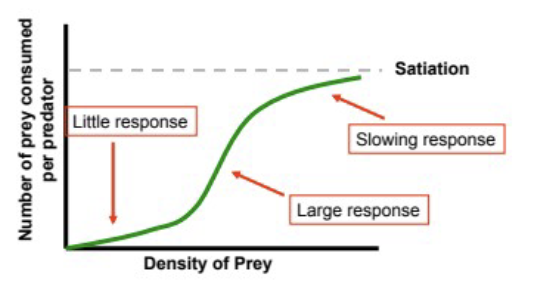
type III functional response
predator response to prey is depressed at low prey densities, then accelerates, then plateaus once reaching satiation
Hen harriers and red grouse
when red grouse populations are low → hen harriers dont focus on them, red grouse becomes more abundant → prey switching, efficiency increases
hen harriers concentrate on red grouse → leads to satiation and efficiency levels off
search image
when a predator begins to recognize and focus on a specific type of prey, increase predator efficiency in functional relationship between predator and prey populations
prey switching
predator switches from consuming one prey to another depending on which is most abundant, functional models assume no prey switching, but as prey population declines predator may switch to another prey species weakening their interactions
immigration and emigration
not included in functional response, assumes populations are closed, may help prey populations avoid crashing, can lead to prey switching
nelson and mech (2006)
wolves heavily preyed on deer in that area → depleted deer populations → wolves switched to new prey, moose → shows prey switching allows predators to persist
territorial behavior
puts a cap on how many predators can occupy an area, predator density cannot freely increase as prey becomes abundant
1910, swam over
when and how did moose colonize isle royale
1948, crossed ice bridge
when and how did wolves colonize isle royale
factors influence moose populations
overbrowsing of balsam fir, malnourishment, ticks
factors influence wolf populations
inbreeding due to low population size
lynx-hare cycle
classic, predator driven, predictable cycles, wolf-moose cycle has lagged responses and irregular cycles in comparison
rmax
isle royale, maximum per capita growth rate of resource recruitment (balsam fir), how fast plant can grow when not limited, represents growing season and climate conditions
k
isle royale, carrying capacity in absence of consumers, max amount of plants environment can support, represents habitat productivity
a
isle royale, area searched per unit time by consumer, how quickly moose can find and consume plants, higher means stronger pressure and less plant biomass
h
isle royale, handling time for each item, limits how much moose can eat, satiation
c
isle royale, coefficient for converting resource consumption into offspring, higher means more offspring per unit food
d
isle royale, consumer per capita mortality rate
a
tri-trophic model, maximum rate moose can eat plants, higher means stronger grazing and reduced plant biomass, represents foraging ability and feeding time
b
tri-trophic model, plant biomass at which consumption is half of maximum, low means moose feeding rates are high even when plants are scarce, high means moose need lots of plants to feed effectively
d
tri-trophic model, amount of food needed just to survive (no growth or reproduction), if intake is less than → moose will starve, if intake is more than → moose can grow and reproduce
e
tri-trophic model, conversion efficiency of turning food into offspring, higher means faster population moose growth
A
tri-trophic model, consumption rate of wolves on moose, how effectively wolves kill moose, represents hunting success
B
tri-trophic model, half saturation content for wolves, low → wolves feed efficiently even when moose density is low, high → wolves struggle at low moose density
D
tri-trophic model, amount of food needed for wolves to survive, if intake is less than → wolves decline, if intake is greater than → wolves grow and reproduce
E
tri-trophic model, efficiency of converting moose into new wolves, higher → more wolf reproduction per moose eaten
isle royale models
complex, chaotic patterns, should expect inconsistency of moose and wolf relationship over the years
wolf territoriality
limits wolf population growth as wolf density increases, as wolf density increases there is more competition and reduced reproduction or survival, density dependent self regulates, LV model assumes wolves increase indefinitely
stochastic events on isle royale
severe winters (moose starvation), disease outbreaks (canine parvovirus in wolves), inbreeding of wolves, variation in plant growth, human intervention (2018-2019)
landscape of fear
prey respond to predators by changing their behaviors based on perceived risk, prey adjust where they feed, move, and rest, moose will reduce feeding efficiency due to risk of wolves
predator efficiency decreases, decrease prey feeding
vigilance of prey increases →
increasing group size
lower predation risk, individuals vigilance reduces, higher predator detection
selfish herd benefits
individuals in a large herd would use other members from the herd as protection against predators, lowers likelihood and predator attacks
trophic cascade
when one trophic level cascades down through multiple layers of the food web, indirect effects on multiple levels
wolves changed rivers
wolves are reintroduced → wolves reduce elk numbers, make elk avoid river areas → fewer elk leads to more aspen, willow, and cottonwood → more vegetation causes less erosion and beavers dams alters rivers
mesopredator release
mid sized predator populations explode after removal of a top predator from food chain
aspen
before wolves → heavily browsed by elk, shrubby growth, little recruitment. After wolves → ripple el al. found increased height and new cohorts of young trees, reduced elk browsing led to more growth
willow
before wolves → heavily grazed and suppressed biomass. After wolves → increased height and crown volume, expansion in riparian areas where elk avoided, behavioral changed in elk led to less browsing pressure
cottonwood
before wolves → poor or no recruitment, missing younger age classes. After wolves → recovery was patchy and site dependent, reduced elk browsing some regeneration
ripple et al. (2011)
measured height of young aspen stems and browsing status, found that stems grew taller after wolves reintroduced
Kauffman et al. (2010)
used broader sampling compared to ripple et al. and found no significant recruitment of aspen, only protected areas (enclosures) showed recovery
Mech (2012)
found many aspen still showed no recovery, willow recovery is inconsistent and based on water availability, beaver populations increased due to willow recovery and not solely due to wolves, and elk decline was also due to humans, grizzlies, and climate variation
feedback loop
high water table → better willow growth → more food for beavers → dams raise water table, can’t tell if driving factor for willow growth is due to wolves or beavers
brice et al. (2021)
criticized aspen stem studies were biased → five tallest stems were chosen, used random sampling and found evidence for recovery was weaker
ripple-beschta-painter
measured five tallest stems, useful for detecting maximum potential growth, not representative of overall population, goal was to detect potential recruitment
MacNulty et al. (2025)
flawed methods used for measure willow crown volume (distorted shapes), sampling was uneven across time, photos only highlighted certain areas that were recovering