The excitable membrane and action potential
1/19
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
20 Terms
What is the resting membrane potential and what are the two factors determining it?
At rest, Vm ≈ −65 mV (inside is ~65 mV more negative than outside).
Determined by:
1. Concentration gradient of ions across the membrane
2. Selective permeability of the membrane to different ions
Why do ions need membrane proteins to cross the lipid bilayer?
The lipid bilayer is highly impermeable to polar species and ions. Thus, for ions to cross the barrier, integral membrane proteins are required to facilitate this transport.
This includes:
Passive transport through channel-mediated or transporter-mediated proteins
Active transport through “pump”-proteins
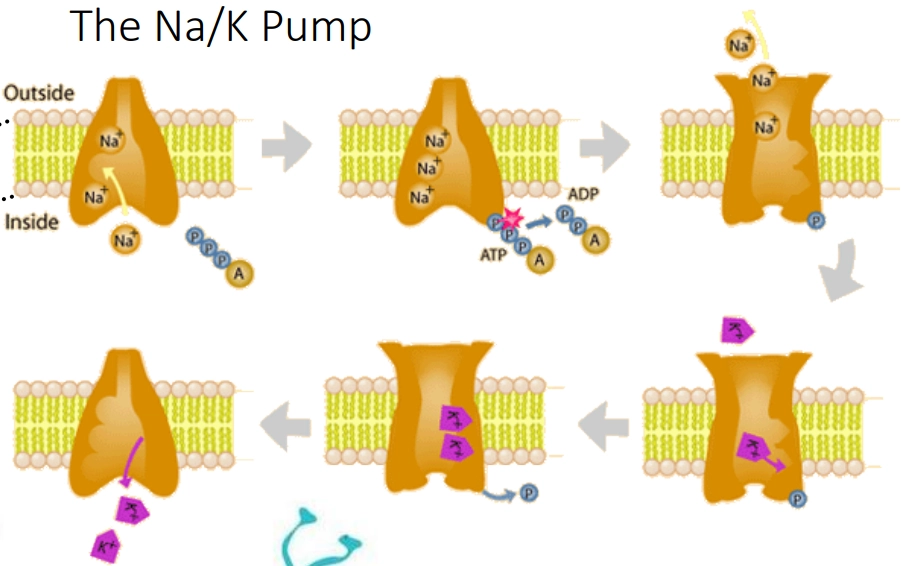
Describe the Na/K pump cycle and its importance.
Ionic gradient under normal conditions:
Na+: high outside, low inside
K+: high inside, low outside
The Na/K pump is an active transporter that is important in setting the membrane potential of neurones.
Inside-facing opening binds 3 Na⁺
1 ATP hydrolysed → conformational change opens outward
3 Na⁺ released outside; pump loses Na⁺ affinity, gains K⁺ affinity
2 K⁺ bind outside (no ATP needed)
Conformational change returns pump to original shape; 2 K⁺ released inside
Net per cycle:
3 Na+ out
2 K+ in
1 ATP hydrolysed
Net outward positive charge
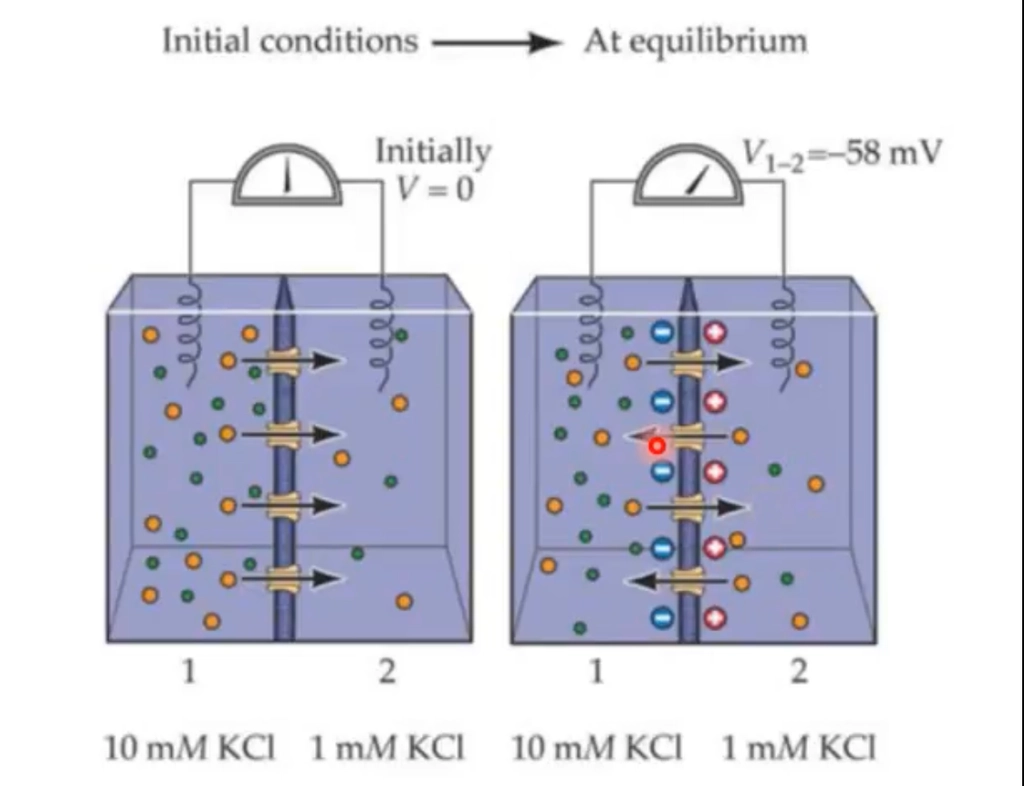
What does the figure tell us about Nernst’s equation?
Shows a KCl gradient (10 mM vs 1 mM) with K⁺-selective channels.
Initially V=0, so K⁺ flows down its concentration gradient (left to right).
As K⁺ moves, charge builds up, creating a voltage that opposes further flow.
At equilibrium, V₁₋₂ = −58 mV —> the Nernst potential for K⁺ at this gradient.
AKA Equilibrium potential (Eion)
Net ion movement = 0.
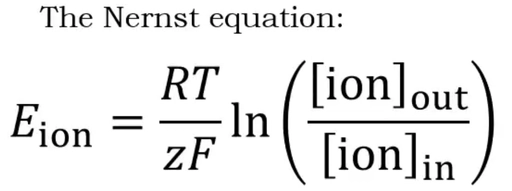
Nernst equation?
Describes the voltage that arises because of the difference in concentration of a specific ion type.
Eion = equilibrium potential for that ion
R = gas constant, T = temperature (Kelvin)
z = valence of ion (e.g. +1 for K⁺, +2 for Ca²⁺, −1 for Cl⁻)
F = Faraday's constant
[ion]out / [ion]in = concentration ratio outside/inside
How does Ohm's law apply to ion channels?
The movement of ions is described as the current (I)
At equilibrium, the current = 0
Ohm’s law predicts the direction and amplitude of the current flow
Conductance (G) is the specific permeability; represent the portion of channels that are open
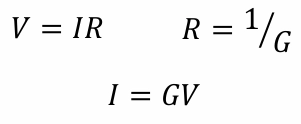
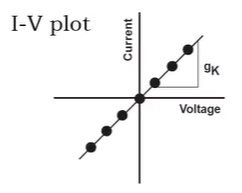
I-V plot?
X-axis: voltage (V)
Y-axis: current (I)
Slope: conductance (G)
More open channels → higher G → steeper slope on I-V plot
0 channels open → no G → flat line along x-axis
Concentration gradient → G (slope) shifts so that x-intercept is the Eion
Positive and negative currents.
Cations (+):
Positive current = outward flow
Negative current = inward flow
Anions (-):
Positive current = inward current
Negative current = outward current
What is the reversal potential (Vrev)?
The voltage at which the net current of a system is 0.
Vrev can be used interchangeably with Eion for one type of ion
Why is the membrane more permeable to K⁺ than Na⁺ at rest?
Leak channels are open at rest and are mostly selective for K⁺. These keep the membrane potential close to the K⁺ equilibrium potential (EK ≈ −90 mV), pulling Vrest negative.
What is the concentration gradient of Na⁺, K⁺, Ca²⁺, and Cl⁻ relative to the cell?
High outside: Na⁺, Ca²⁺, Cl⁻
High inside: K⁺
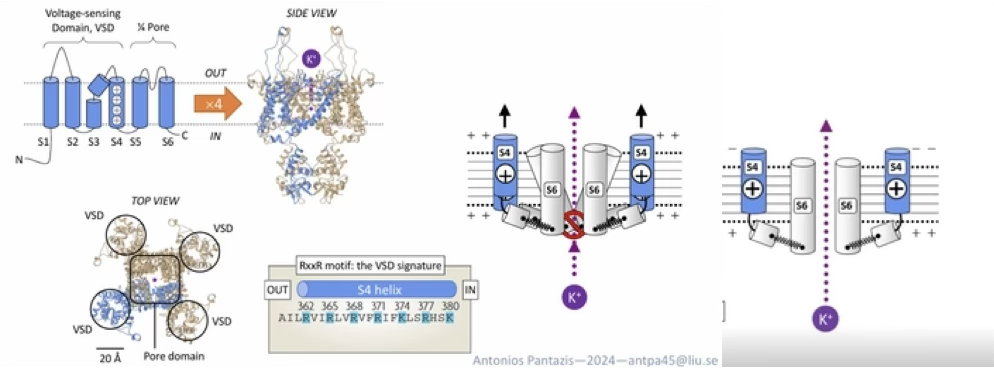
Describe the structure and gating mechanism of a voltage-gated K⁺ channel.
A tetramer of 4 symmetrical subunits, each with segments S1–S6:
S1–S4: voltage-sensing domain (VSD). S4 has positively charged residues every 3rd amino acid
At rest (negative inside): S4 is pulled inward, keeping pore closed
Upon depolarisation: S4 is repelled upward (outward), changing pore conformation → K⁺ can flow
Conductance follows a sigmoidal relationship with voltage
What are the 3 main voltage clamp techniques?
Cell-attached recording: pipette on membrane surface with suction; records single channel currents
Whole-cell recording: stronger suction breaks membrane; records currents from all channels in the cell
Two-electrode voltage clamp (TEVC): used with Xenopus laevis oocytes; channel gene injected, two electrodes control Vm and record current
How did the voltage-gated Na⁺ channel evolve and how does it differ structurally from KV?
NaV evolved from KV via 2 gene duplication events, making it a tandem concatemer
4 domains in one polypeptide (pseudo-tetrameric)
Each domain has accumulated different mutations → different functional properties per domain
Eukaryotic CaV channels share the same architecture
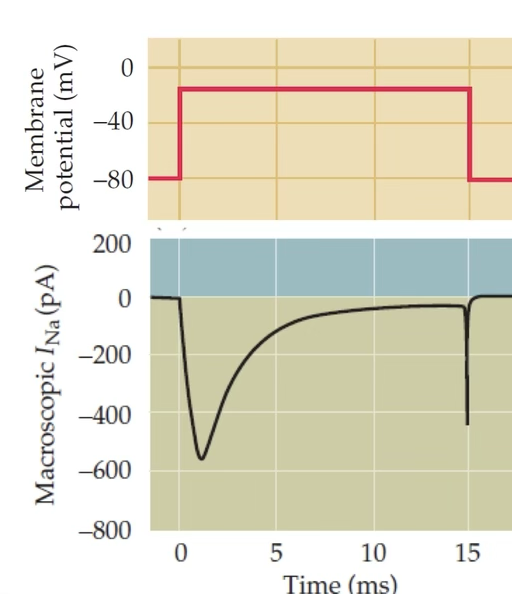
How does a NaV channel open and inactivate? What is the IFM motif?
Opening (m-gate): VSDs I–III sense depolarisation > at −40 mV, pore opens → Na⁺ flows in
Inactivation (h-gate): VSD IV controls inactivation. The IFM motif (Isoleucine-Phenylalanine-Methionine) on the III–IV intracellular loop physically blocks the open pore from the cytosolic side
Open probability: pO = m³ × h
h = 1 → not inactivated; h = 0 → fully inactivated (no current despite open pore)
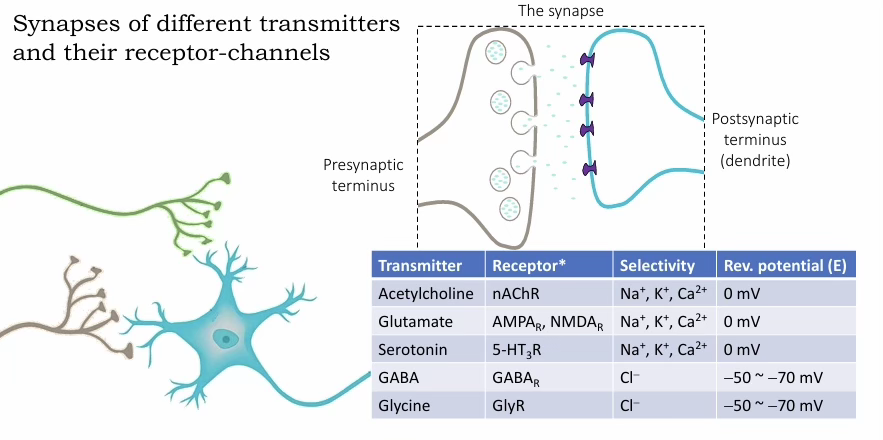
Synaptic signalling
Glutamate / ACh / Serotonin (cation channels): Vrev ≈ 0 mV — permeable to Na⁺, K⁺ (sometimes Ca²⁺); depolarising → excitatory
GABA / Glycine (Cl⁻ channels): Vrev ≈ −50 to −70 mV — permeable to Cl⁻; near or below resting potential → inhibitory/stabilising
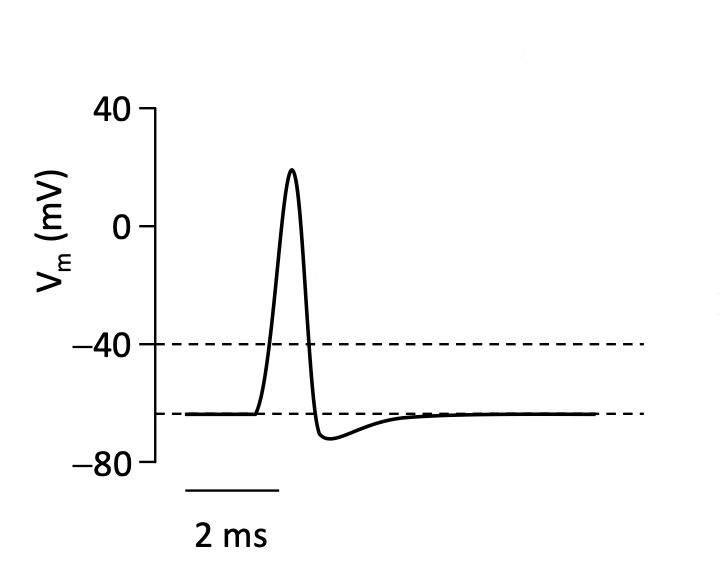
Describe the phases of the action potential.
Initiation: Integration at soma; Axon initial segment decides whether AP is carried out or not → if Vm > −40 mV (threshold), NaV channels open → Na⁺ influx → depolarisation → more NaV open (Hodgkin cycle / positive feedback)
Peak (30–40 mV): Vm approaches ENa; peak is reached where net current = 0 momentarily; NaV inactivation begins; KV channels start opening
Termination: KV open + NaV inactivated → repolarisation toward EK → undershoot (hyperpolarisation) because KV slow to close → leak channels restore Vrest
What are the different refractory periods?
Absolute refractory: no AP possible regardless of stimulus (NaV inactivated)
Relative refractory: need stronger-than-normal stimulus (some NaV inactivated, some KV still open)
Why do refractory periods exist and why do we need them?
Refractory periods exist because:
NaV channels are inactivated (need time to recover)
KV channels are still open and opposing depolarisation
Function: prevents backward propagation of the AP — the region behind the AP is refractory, so the signal can only travel forward along the axon.
What solution do vertebrates use to propagate signals faster?
Myelination: myelin sheath (from oligodendrocytes in CNS, Schwann cells in PNS) wraps the axon and provides electrical insulation, thereby decreasing membrane capacitance and increasing resistance, leading to faser signal propagation.
Voltage-gated ion channels are only at nodes of Ranvier (gaps between myelin). The AP 'jumps' from node to node → saltatory conduction, which is much faster and more energy efficient.