Ribosome & Protein Translation: Eukaryotic
1/21
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
22 Terms
Eukaryotic Ribosome
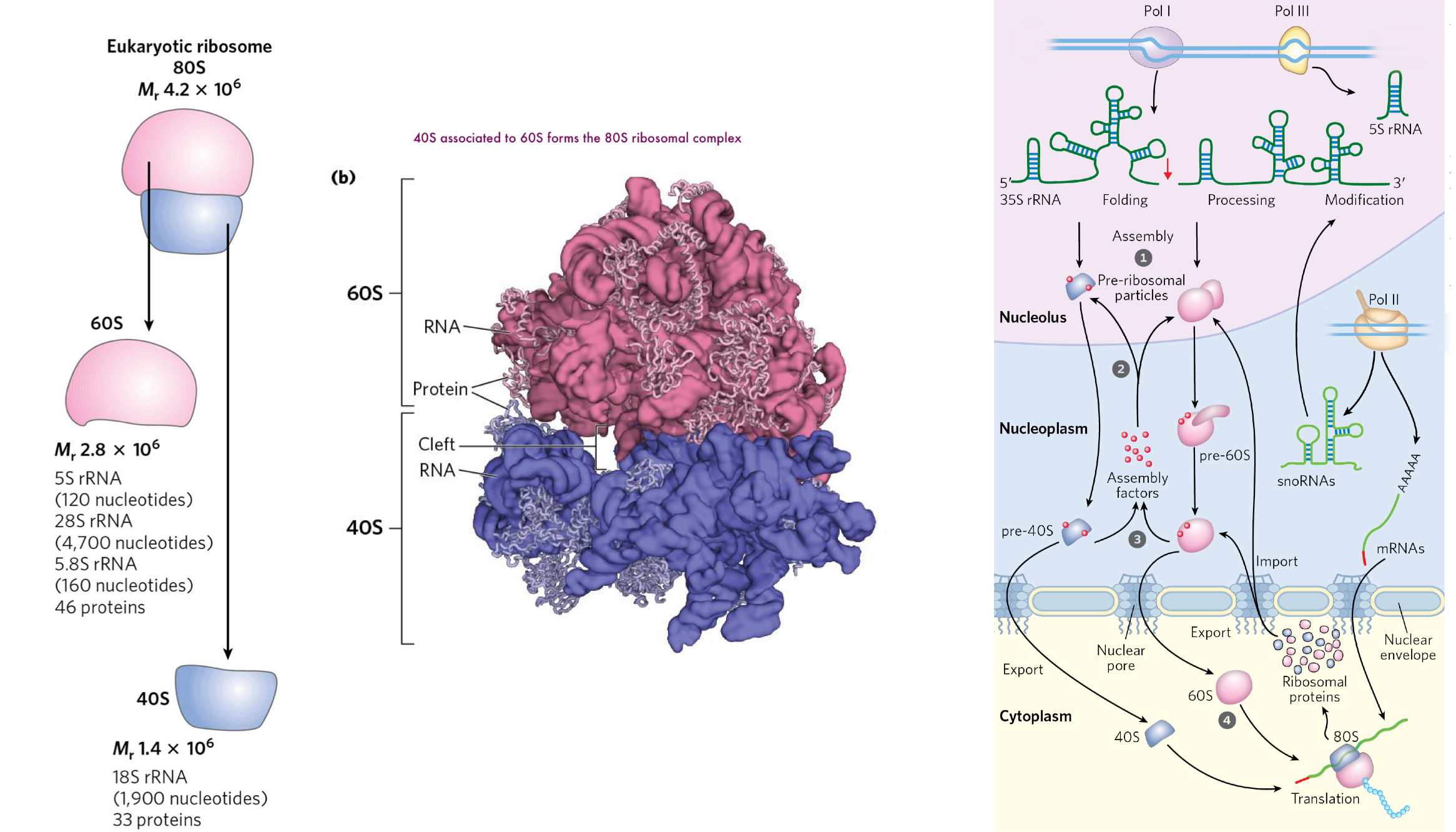
the eukaryotic ribosome is structurally similar to the prokaryotic complex, but larger
assembly of the eukaryotic ribosome requires the coordination of several events across diff cellular compartments
Eukaryotic Translation
key difference: transcription occurs in the nucleus and translation occurs in the cytosol (processes are uncoupled in eukaryotes)
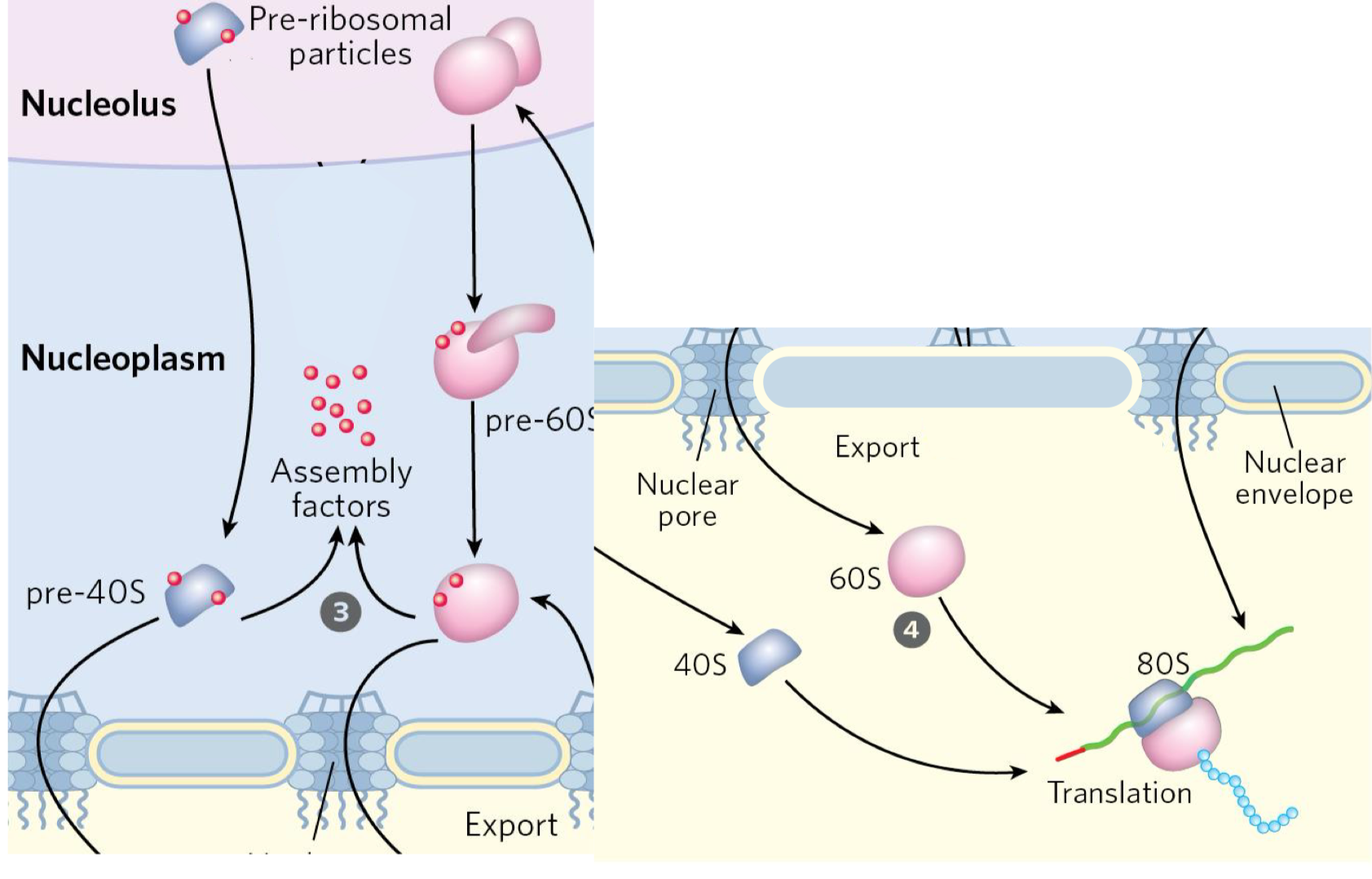
rRNAs are processed in the nucleolus, while small ribosomal proteins are synthesized in the cytosol before being imported and assembled into pre-ribosomal particles (pre-40s and pre-60s)in the nucleolus with the help of assembly factors
the pre-40s and pre-60s subunits are then exported to the cytosol, and assembly factors are recycled
eukaryotic translation is similar to the prokaryotic process but has major distinctions especially in the initiation process
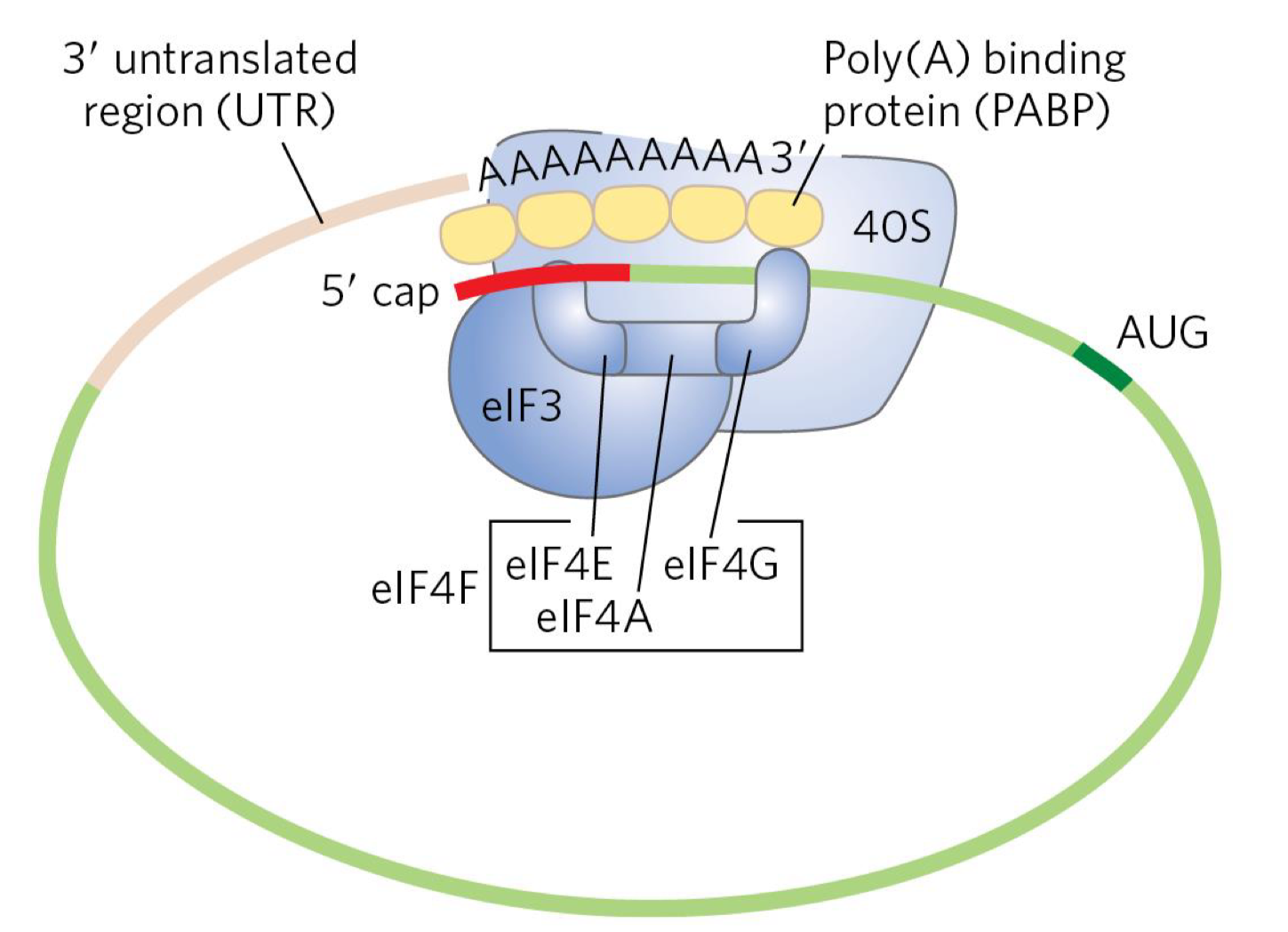
Factors binding to mRNA: PABP
important factors bind to mRNA in the cytosol, including the 3’ poly(A)-binding protein (PABP)
PABP interacts w/ eukaryotic initiation factors (eIFs) bound to the 5’ cap
this interaction promotes circularization of the mRNA, enhancing both translation efficiency and mRNA stability
43S Pre-Initiation Complex
eukaryotic initiation involves multiple eukaryotic initiation factors (eIFs) that assemble with the 40S ribosomal subunit to form the 43S pre-initiation complex
the 43S pre-initiation complex contains the Met-tRNAi (unmodified Met but dedicated initiation tRNA) in the P site delivered by eIF2-GTP
similar to prokaryotic initiation, eIFs block the A site and prevent premature association with the 60S large subunit
also like in prokaryotic initiation, there are 2 tRNAs that recognize Met (AUG) codons, one is used during elongation and one during initiation
these tRNAs are activated w/ unmodified Met
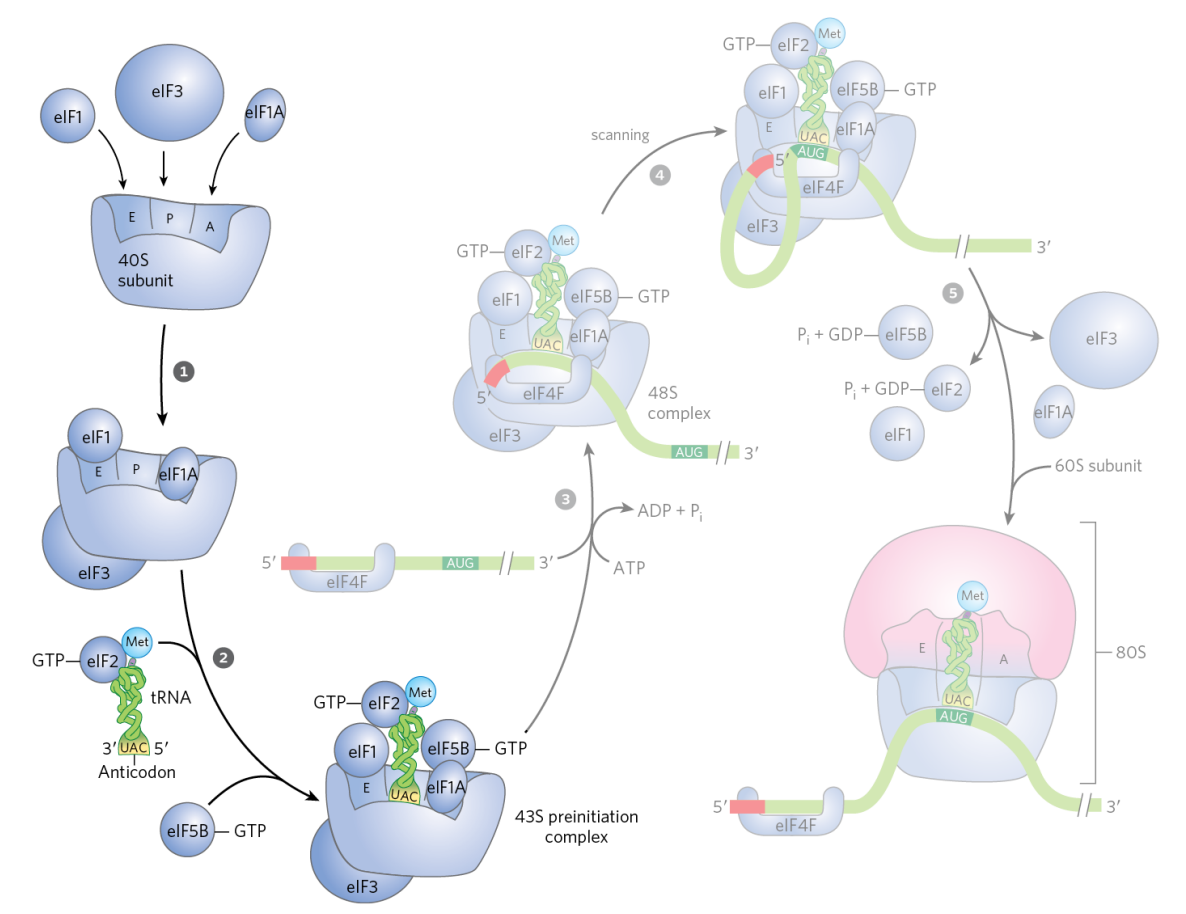
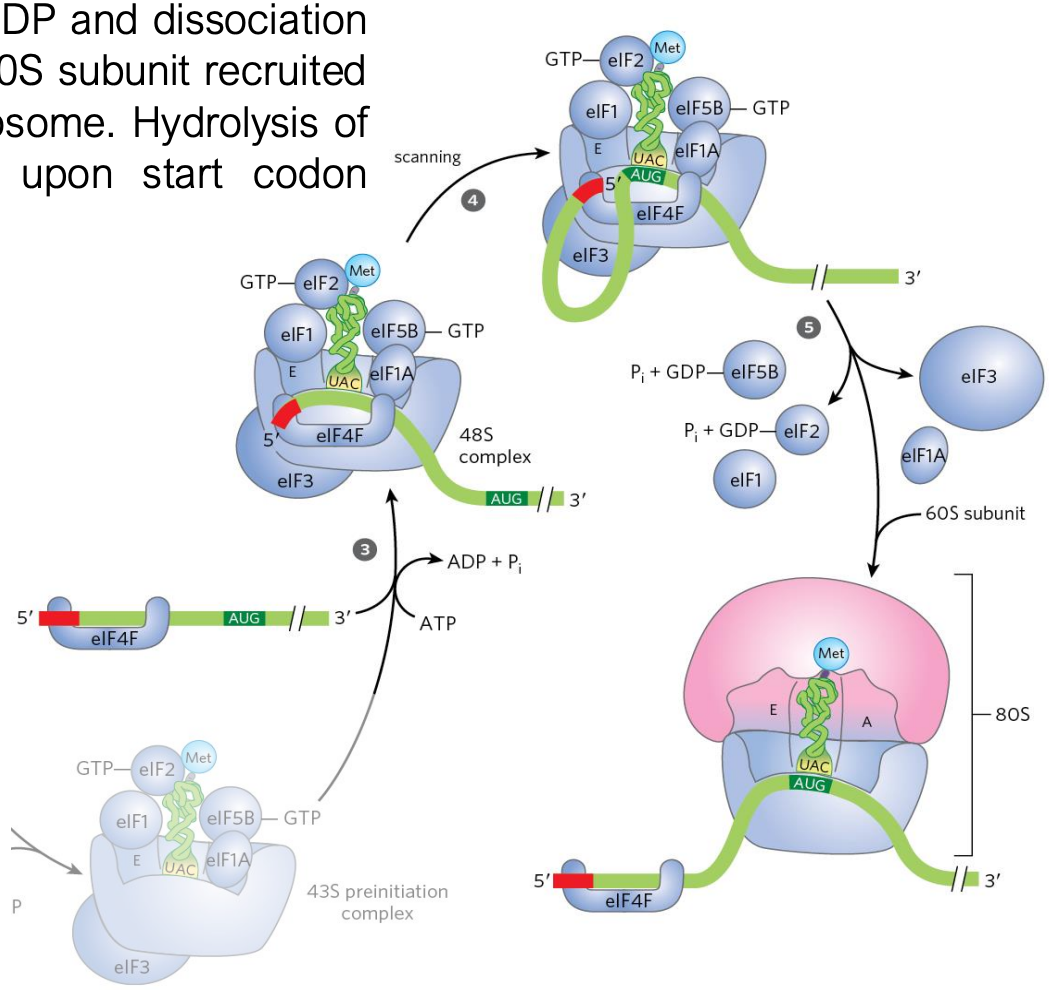
mRNA Scanning & Start Codon Recognition
with the assistance of additional eIFs, the mRNA is recruited
the 43S pre-initiation complex after mRNA has been recruited (48S initiation complex) scans along the mRNA to identify the AUG start codon (the start codon can be put in the A site)
the scanning process requires helicase activity and ATP hydrolysis to unwind 2º structures in the mRNA
only after GTP hydrolysis and dissociation of initiation factors is the 60S subunit recruited to form the functional 80S ribosome
hydrolysis of eIF2-associated GTP occurs upon start codon recognition
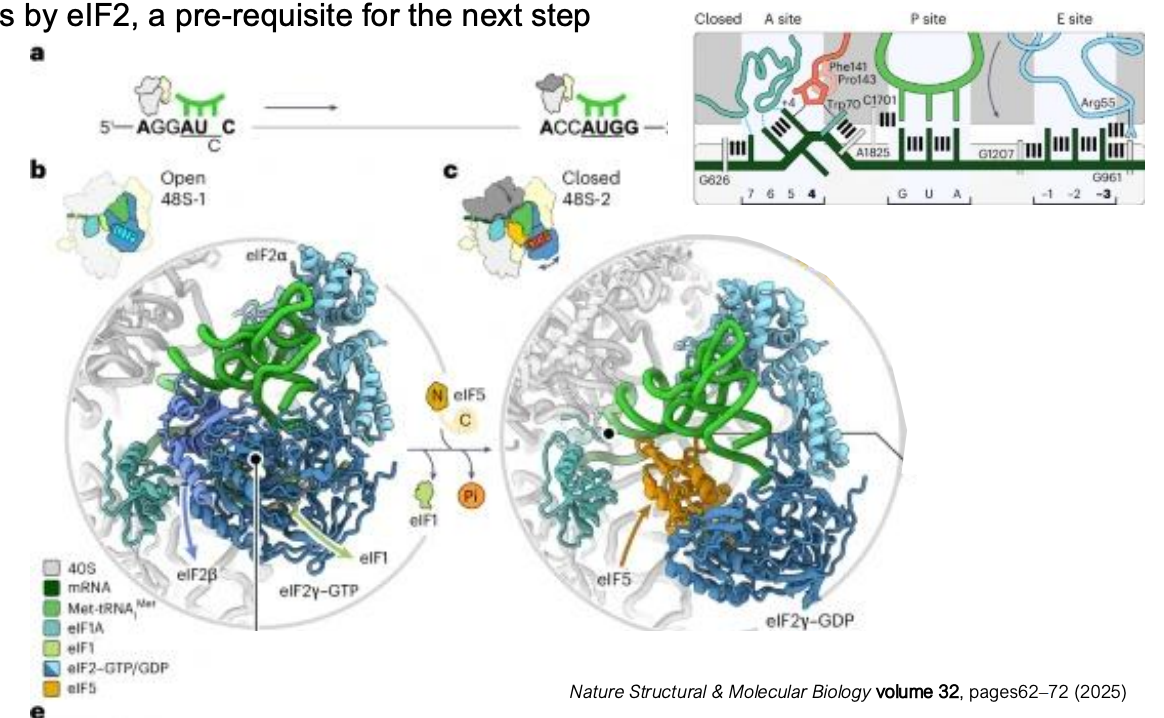
Kozak Sequence
Kozak sequence is a conserved motif and a consensus sequence that surrounds the AUG start codon in eukaryotic mRNAs
it helps the scanning ribosome recognize the correct initiation site
in contrast, the Shine-Dalgarno sequence in prokaryotes is located upstream of the start codon and base-pairs with 16S rRNA to place AUG on the P site
the Kozak sequence typically contains a purine (A/G) in position -3 and a G in +4

Start Codon Recognition: Open → Closed Complex Transition
during scanning, in the open conformation, eIF1 prevents the association of eIF5
recognition of Kozak sequence elements (position -3 and +4) stabilizes multiple interactions with initiation factors and rRNA, promoting formation of the closed complex
closed complex: Met-tRNAi is properly positioned in the P site
eIF1 is evicted
eIF5 (GTPase activating protein) can now associate and promote GTP hydrolysis by eIF2
eIF5 is brought into proximity with eIF2-GTP
GTP hydrolysis signals to ribosomes that the correct AUG is placed in the P site and to proceed with the last stage of initiation
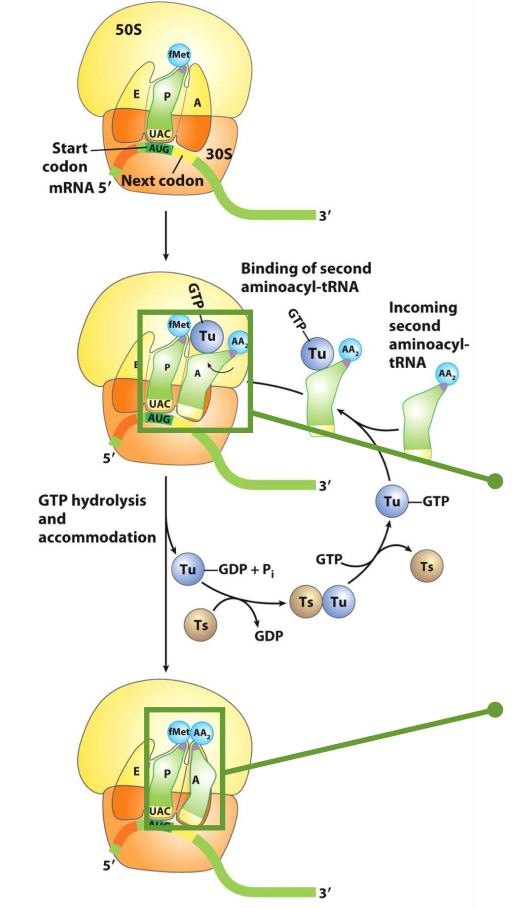
Accuracy of Translation Elongation
translation has no true proofreading, so why isn’t it more error-prone?
because timing is everything!
GTP hydrolysis be EF-Tu (prokaryotes)/eEF1A (eukaryotes) acts as a kinetic checkpoint linking translation speed to accuracy
role: delivers aminoacyl-tRNAs to the A site
GTP hydrolysis only occurs if codon-anticodon pairing is correct
Prokaryotic Accommodation
accommodation is tightly linked to translation speed & accuracy
it consists of a conformational change that repositions the tRNA within the A site, allowing peptide bond formation to occur
accommodation only takes place after GTP hydrolysis and dissociation of elongation factor (Ef-Tu or eEF1A) in its GDP-bound form
Prokaryotic Accommodation: Mech
upon initial binding, the incoming AA-tRNA is not positioned in a configuration that permits peptide bond formation (the 2 AA’s are far away from each other)
incorrect tRNAs typically dissociate from the A site before GTP hydrolysis occurs, ensuring that only correctly paired AA tRNAs remain associated prior to accommodation
following GTP hydrolysis, Ef-TU (GDP) is released, allowing accommodation to proceed
once the incoming AA-tRNA is fully accommodated and correctly positioned, peptide bond formation can occur
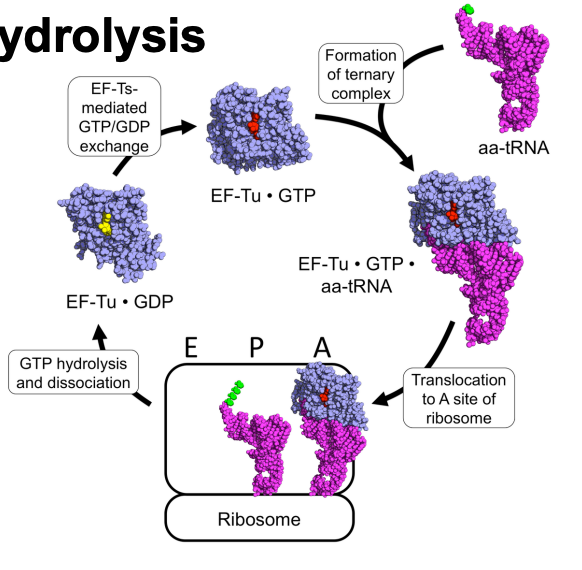
GTP Hydrolysis
GTP binds non covalently to translation factors
upon GTP hydrolysis (GDP + Pi), the translation factor undergoes a conformational change
each GTP-binding TF has a slow intrinsic GTPase activity
GTP hydrolysis is stimulated by a GAP (GTPase-activating protein)
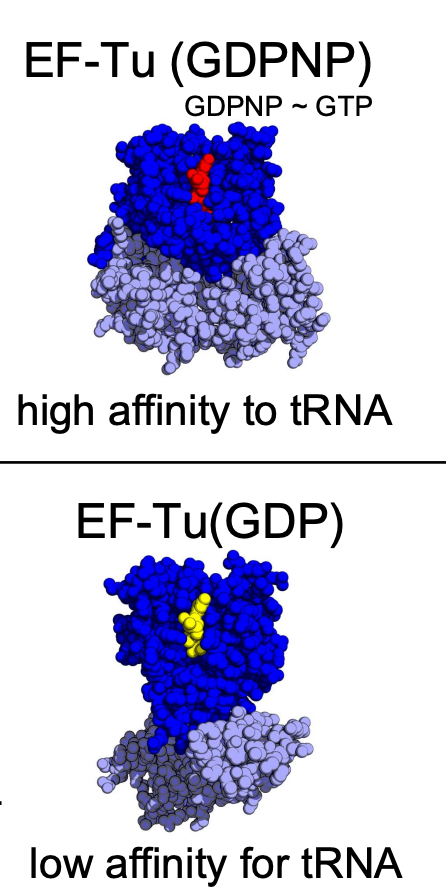
TF Conformational Change
the configuration of elongation factors is different when it is associated w/ GDP vs. an analog of GDP
when EF-Tu elongation factor is bound to GDP, there’s high affinity to charged tRNA
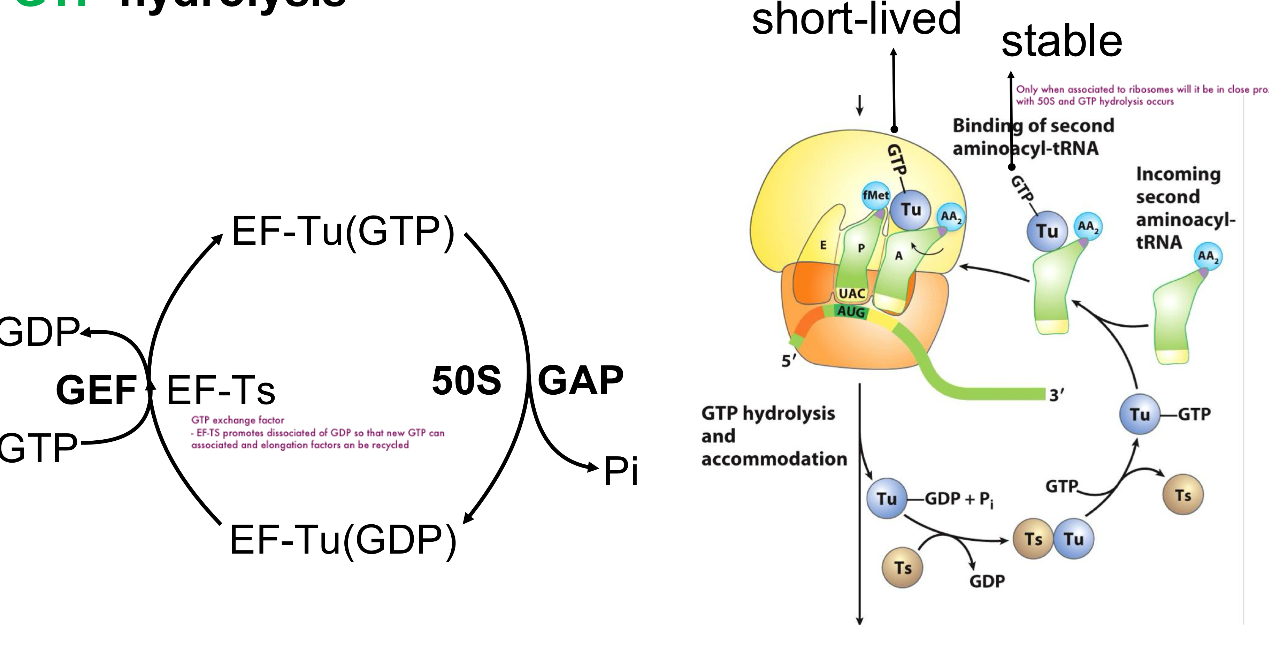
GTP Hydrolysis Continued
GTP hydrolysis by Ef-Tu is stimulated by the 50S ribosomal subunit acting as a GAP (GTPase activating protein)
this timing ensures hydrolysis occurs only for correctly paired tRNAs
Correct codon–anticodon pairing stabilizes the tRNA long enough for GTP hydrolysis
Incorrect tRNAs dissociate before GTP hydrolysis
GDP to GTP exchange is then mediated by EF-Ts acting as a GEF (GTP exchange factor)
EF-Ts promote dissociation of GFP so that new GTP can associate and elongation factors can be recycled
Ef-Tu-GTP is regenerated
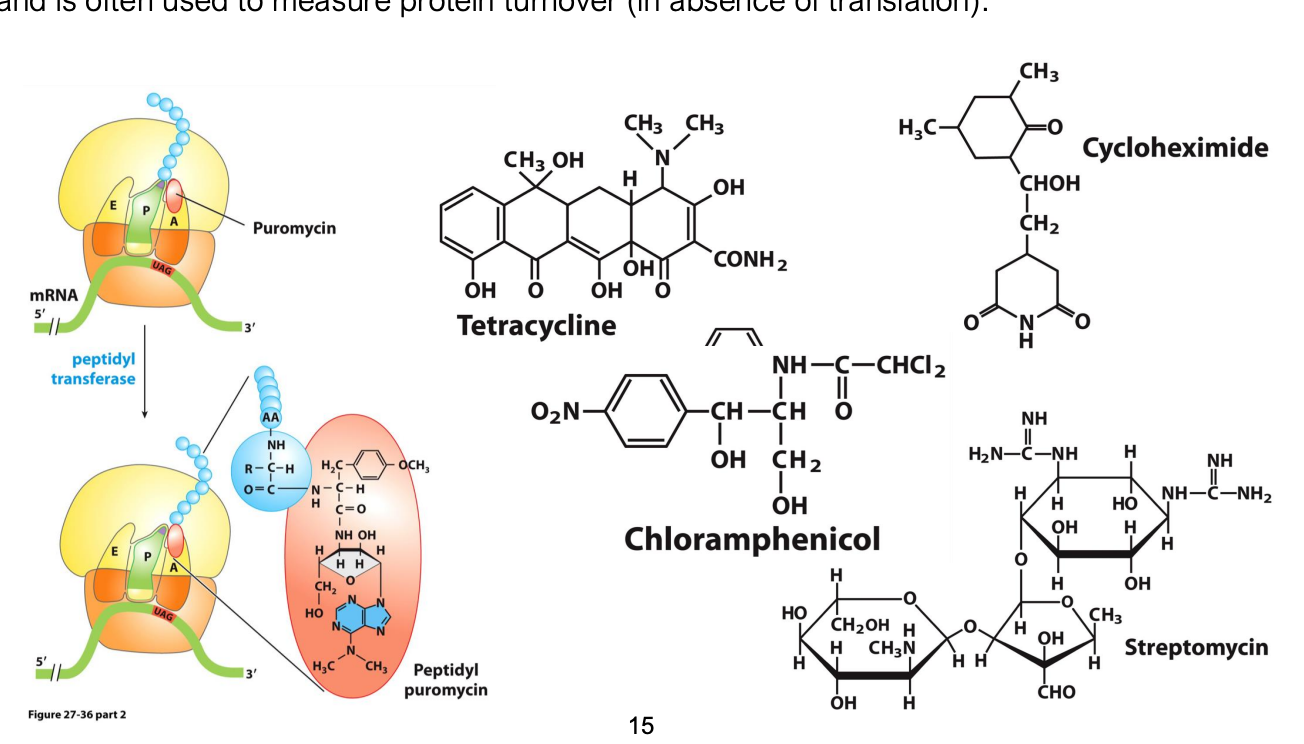
Antibiotics & Translation Inhibition
several small molecules inhibit translation
because many of these molecules inhibit prokaryotic but not eukaryotic ribosomes, they can be used as antibiotics
e.g. puromycin prematurely terminates translation by forming a peptide bond to release the elongated polypeptide, releasing it from the tRNA
Cycloheximide inhibits eukaryotic ribosomes and is often used to measure protein turnover (in absence of translation)