1011 FINAL EXAM SET W5-W12
1/206
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
207 Terms

What are proteins
Proteins are complex molecules
Carry out almost all biological functions

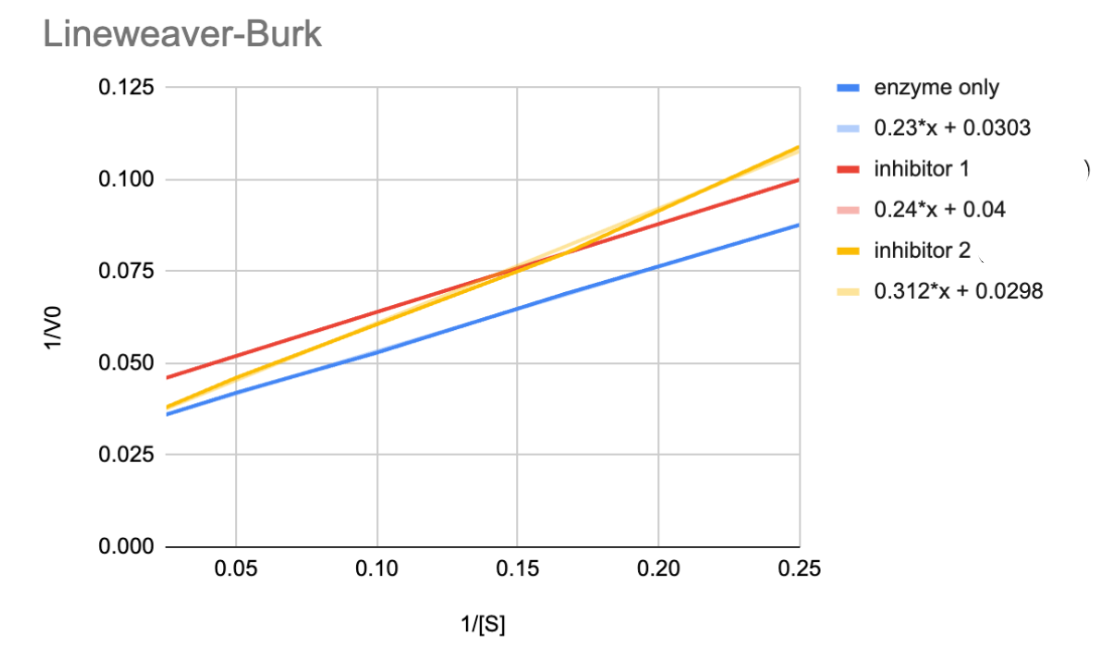
Figure out the nature of Inhibitors 1 and 2
Inhibitor 1 is uncompetitive → we can see this because it is parallel to the enzyme line (no inhibitor)
Inhibitor 2 is competitive → we can see that is will intercept the enzyme line (no inhibitor) line at the same y-axis, this means the Vmax is the same at both conc of substrate, which resemble the graph for competitive inhibition
CALCULATE Km and Vmax for Inhibitor 1
Y-intp → 0.04 therefore the Vmax is 1/Vmax = 1/0.04 = 25.00
X-intp → -0.167 therefore the Km is -1/Km = -(1/-0.167) = 6.00 mM
*When finding the x-intp, make sure you set y=0 and solve for x - BE CAREFUL
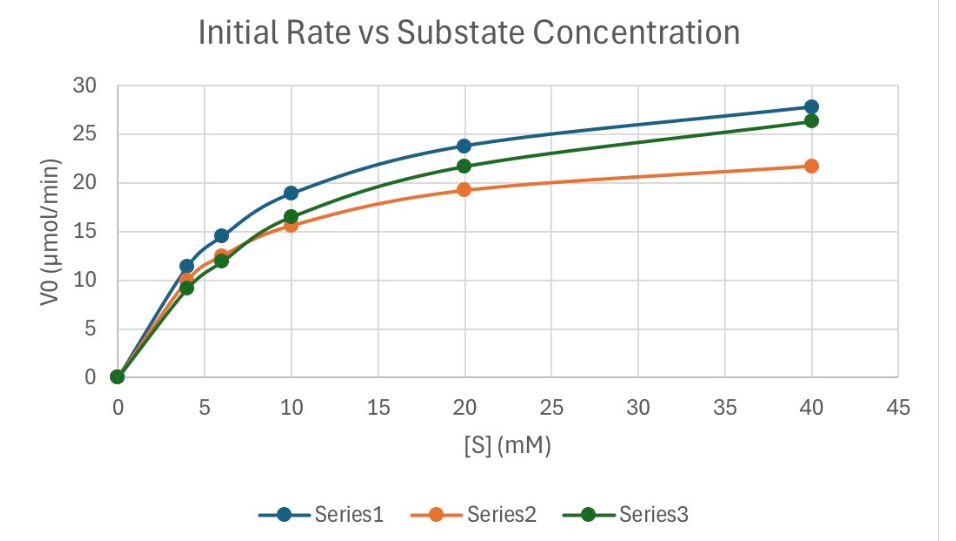
Calculate the Km for Series 1
Infer the Vmax to be about 30μmol/min (this is where the graph should start to plateau out)
Half of Vmax is 15μmol/min
Trace the graph to see the SUBSTRATE CONC at 15μmol/min
This will give about 6mM
How is such diversity of protein functionality achieved?
Protein FUNCTION is dependent upon STRUCTURE, which is dependent upon S-E-Q-U-E-N-C-E
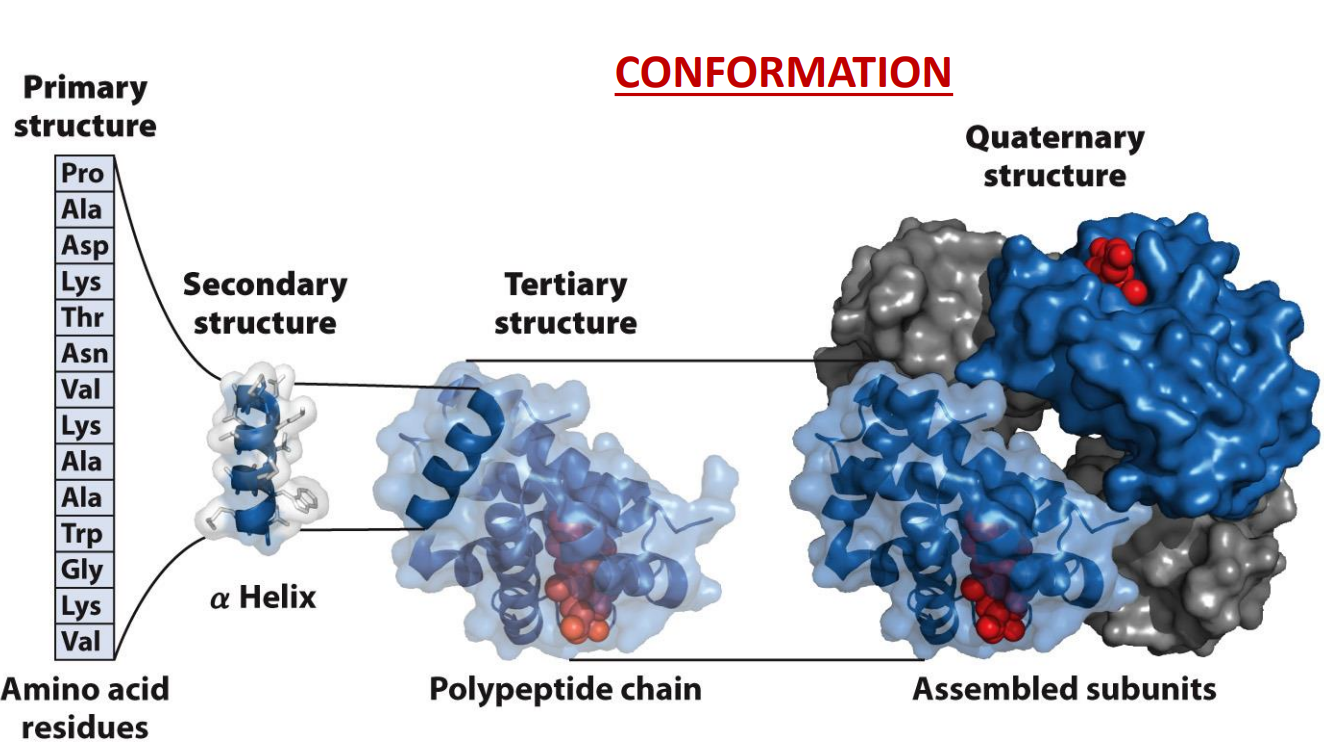
What is the hierarchy of structure in proteins?
Rotation around the peptide bonds in the backbone of a protein means countless possible conformations are possible → functional conformation are termed ‘native’
These ‘conformations’ are when the protein can perform a function
Early observation showed that proteins could be crystalized this suggested that protein had similar 3D structures
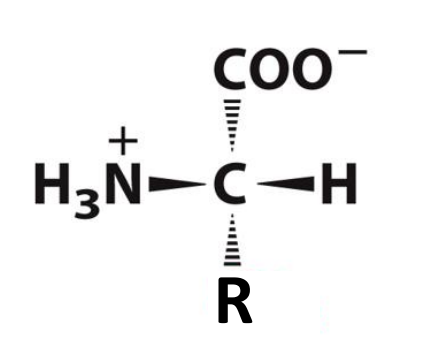
What are amino acids?
Monomers that make up proteins
The backbone of AA is the N, the alpha C and the C in the carbonyl (3 atoms)
There are 20 common AA
Key function of AA:
Capacity of polymerise
Useful acid-base properties
Varied chemical and physical properties (physicochemical)
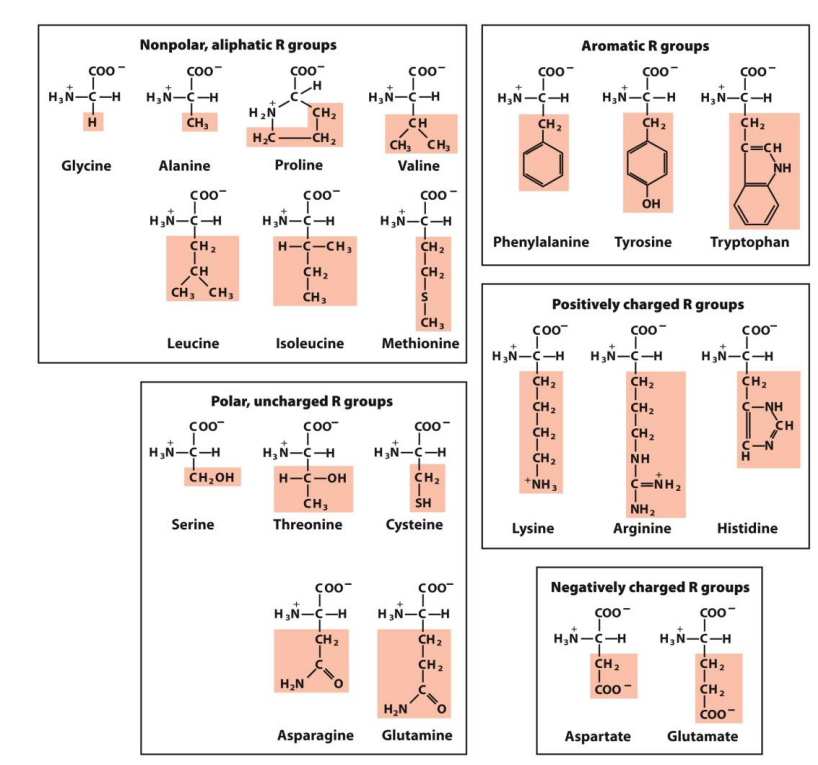
Hydrophobic amino acids are largely buried in the protein interior so that the number of hydrogen bonds and ionic interactions within a protein is maximised
Leucine, Valine and Isoleucine are considered BRANCHED amino acids (BCAAs) - they are essential amino acids, the body cannot synthesise these and they must be obtained from the diet
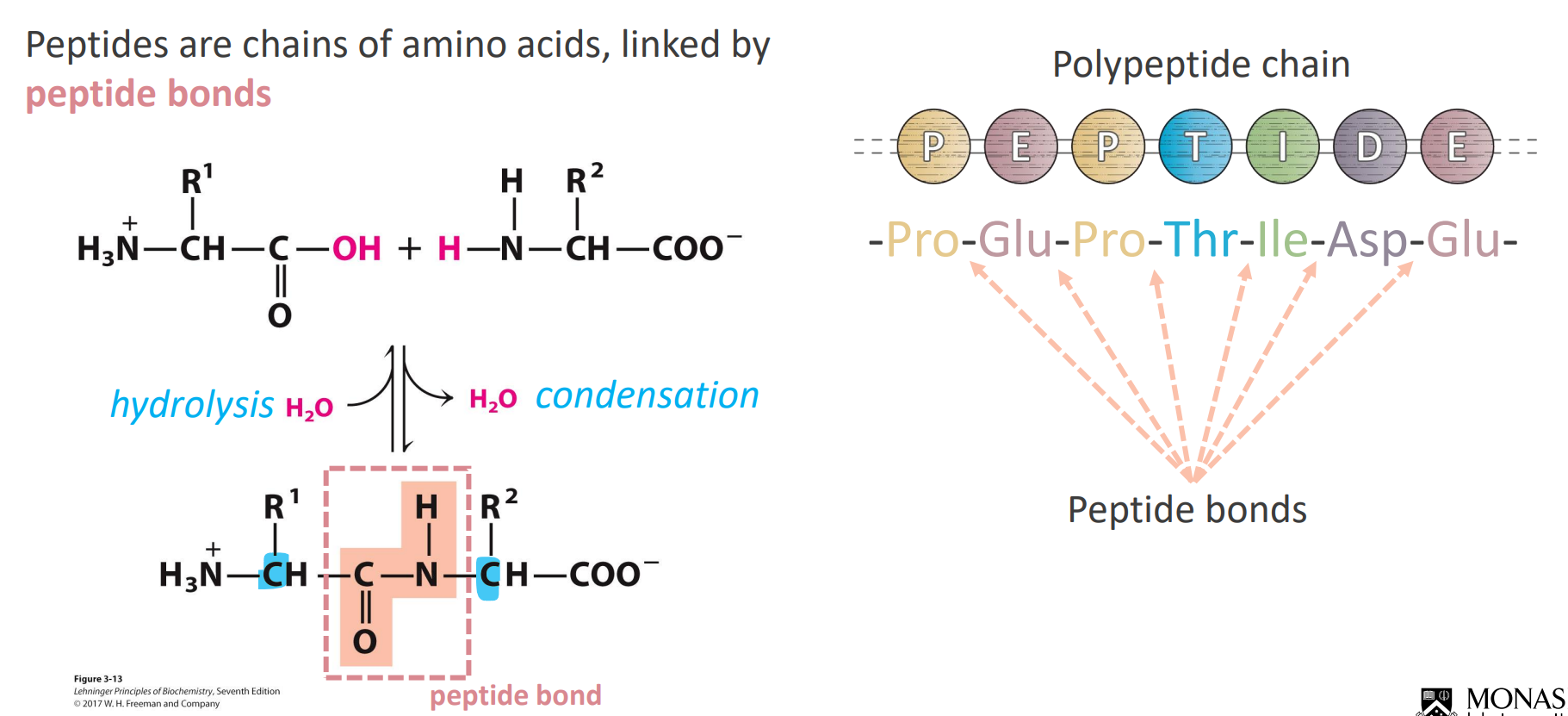
Peptides
Due to electron RESSONANCE the C-N bond has a double bond like characteristic
This makes the bond quite rigid and the CO and NH apart of the peptide become quite planar - no rotation in the peptide bond
We only see rotation on the alpha carbons near the peptide bond (in blue) (hence the R groups orientation)
If asked for the number of amino acids in a polypeptide - COUNT THE R-GROUPS or count the peptide bonds and ADD 1
How does structure arise?
Largely by weak interactions (REVERSIBLE)
Hydrogen bonds
Ionic interactions
Hydrophobic effects
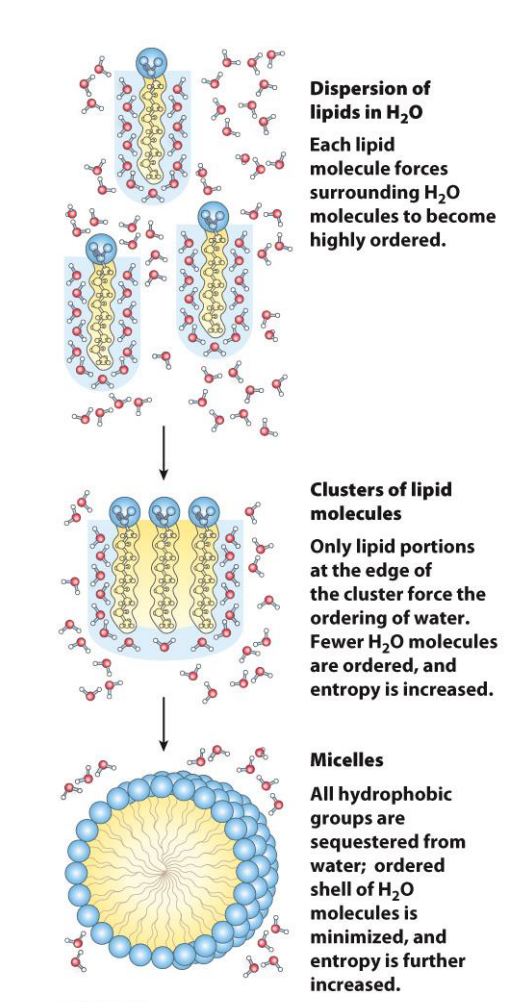
The hydrophobic effect predominates in protein structure and stability
Van der Walls
The release of water molecules from the structured solvation layer around the molecule, as the protein folds, increases the net entropy
Covalent bonds in the peptide backbone also limit structural conformations
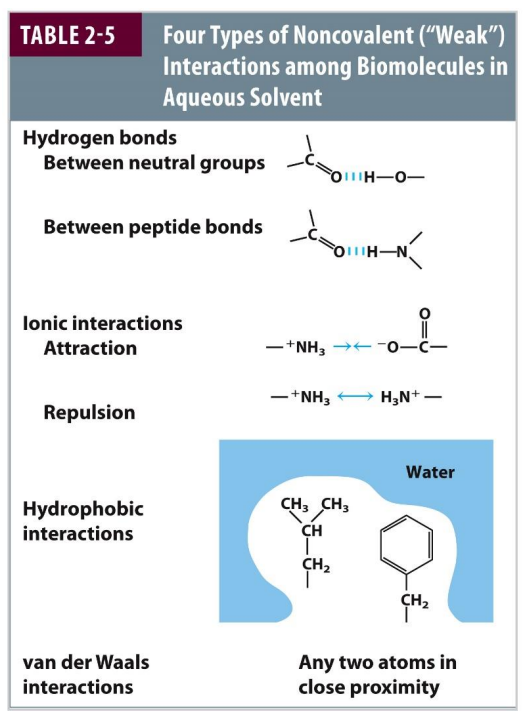
Importance of Hydrogen Bonds
H-bonds are dipole-dipole or charge-dipole interactions
Linear H-bonds are stronger than bent ones
They arise between a covalently bound hydrogen (H-bond doner) and lone pairs of electrons (H-bond acceptor)
They typically involve two electronegative atoms (frequently nitrogen and oxygen).
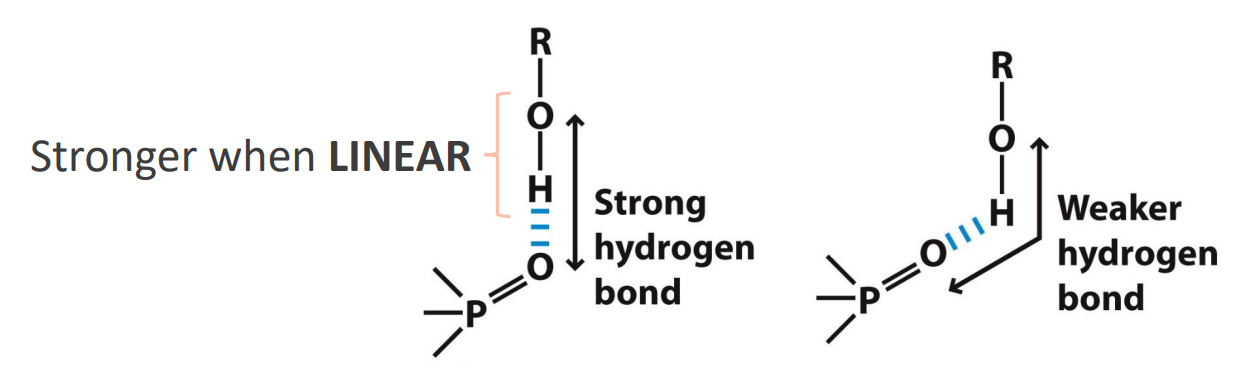
What is the arrangement of amino acids in the Secondary structure?
Several common and stable structure types are observed across proteins (and different amino acids favour different structures)
The Alpha Helix - the R groups determine surface chemistry
The Beta conformation - SHEET
The Beta turn
Secondary structure: Alpha Helix
Backbone atoms wind in right-handed helix
Backbone is twisted (not extended)
One turn every 3.6 residues
Each residue ~100 deg rotation
R groups are all on the outside (B) - helps with stablisation (e.g. ionic bonds that could form (D))
Intrachain H-bonding most important stabilizing feature
CO of res i is H-bonded to NH of res i+4
1 will go with 5
2 will go with 6
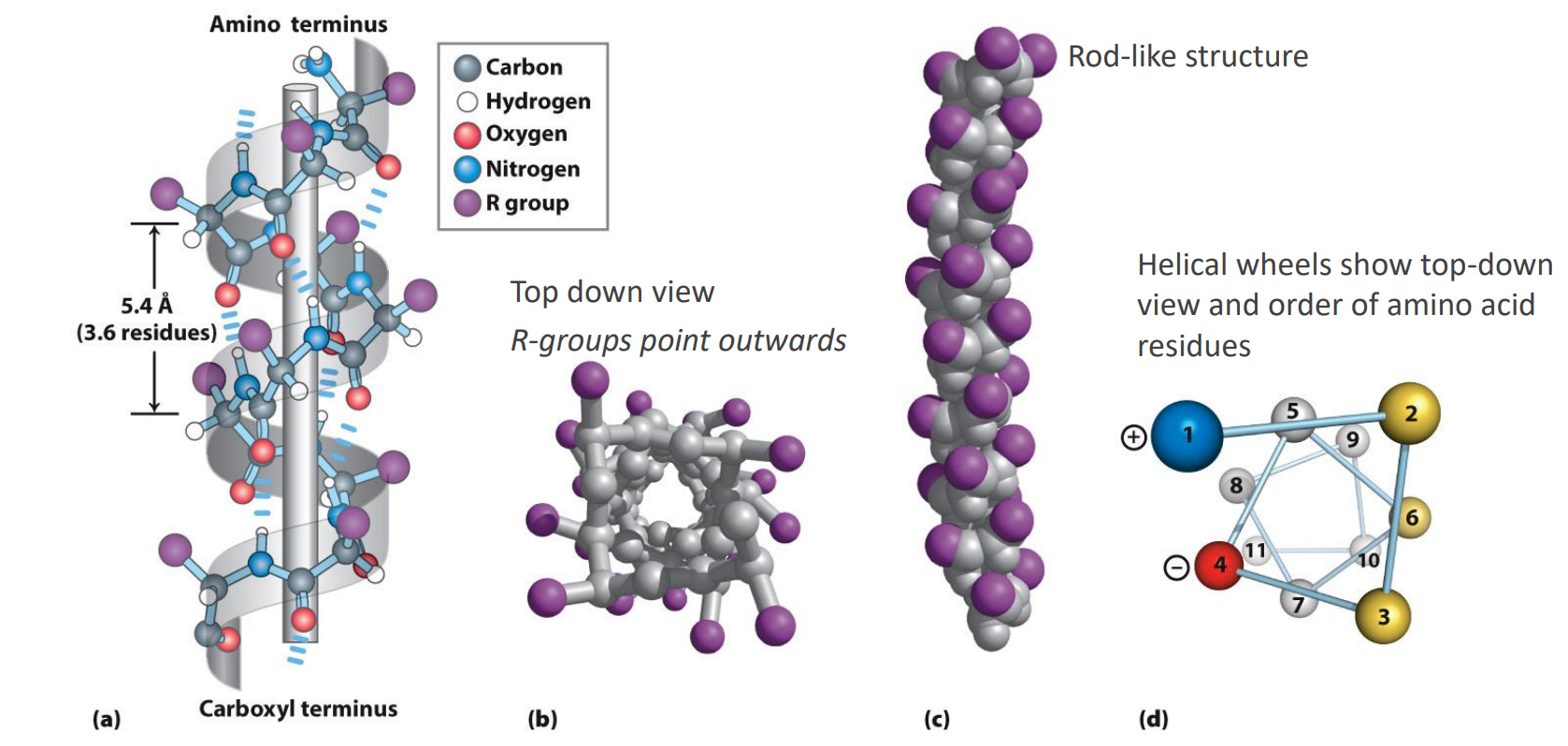
Secondary Structure: Beta Sheets
Extended and more of a zigzag conformation than a helix
Many beta-strands comes together to form a b-sheet
Stabilized by H-bonds
Arrow head is always the C-terminus
Note that the R groups of adjacent amino acids protrude in opposite directions and are above and below the sheet
If the R groups point the same way → parallel, straight H-bonds
Opposite ways → anti-parallel, BENT H-bonds, hence STRONGER
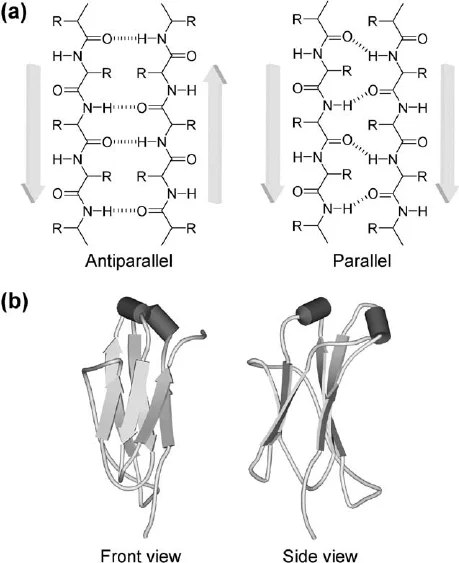
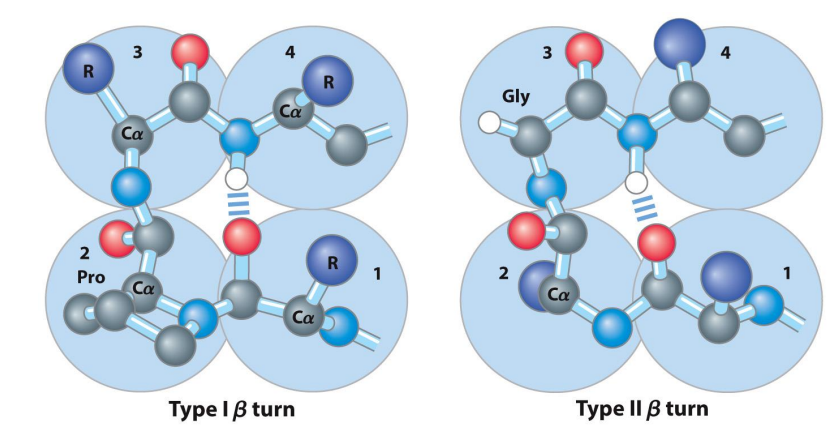
Secondary Structure: Beta Turns
The U-turn of proteins
Allows 180° turn
Connects two ends of anti-parallel beta sheet
Involves four amino acids (specific residues vary)
Glycine is very common at position 3 (type II)
Proline common at position 2 (type I)
Proline has a forced kink that helps with forming Beta-turns
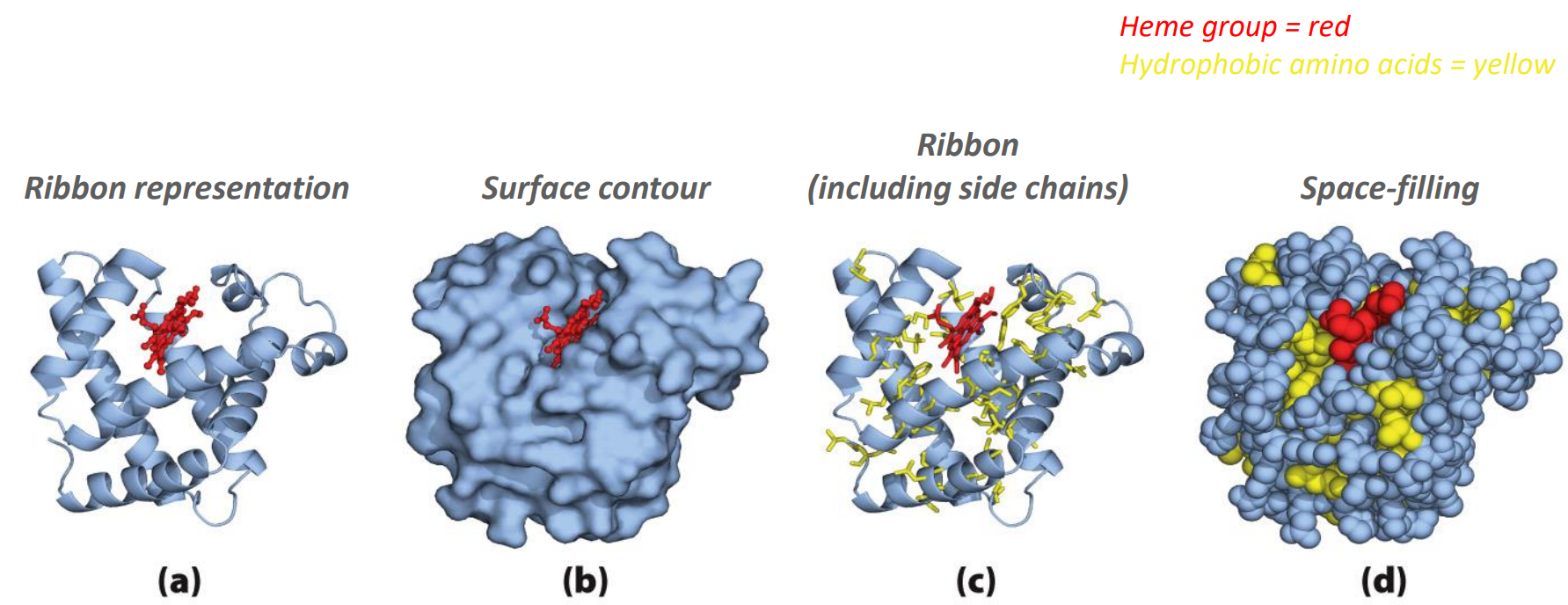
How to display the 3D model of protein structure?
Notice how the hydrophobic parts are often in the interior of the protein (in order to maximize the strong H-bonds)
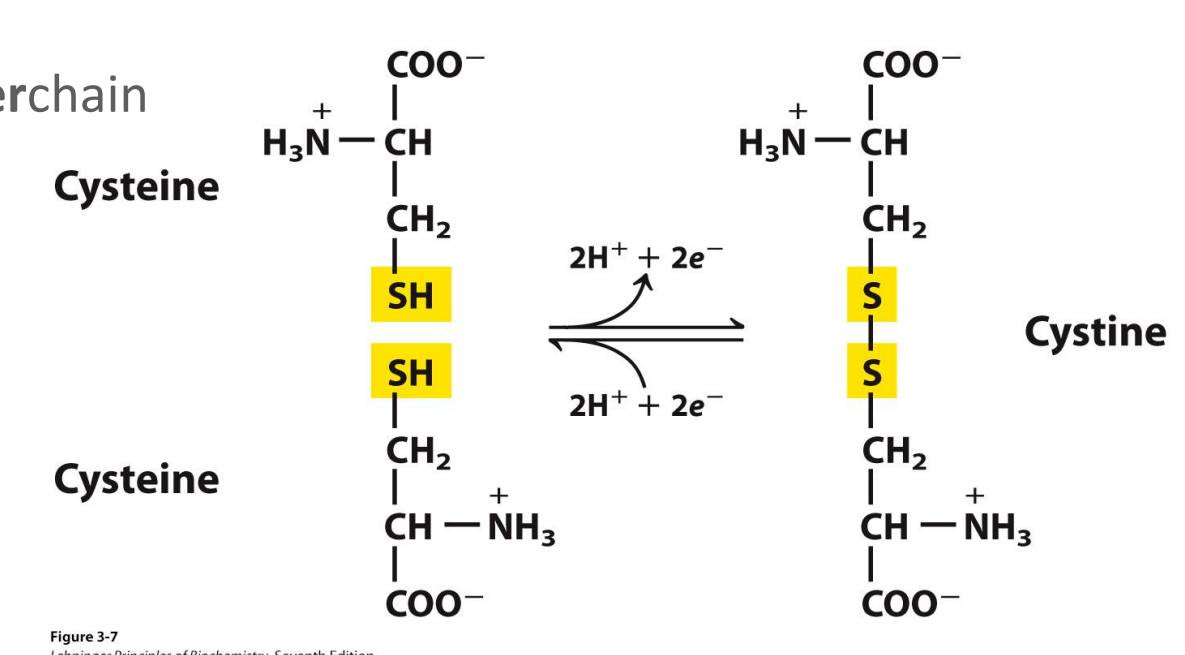
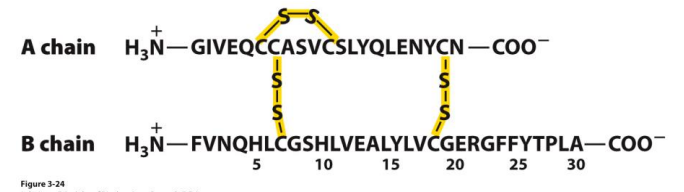
The disulphide bond
Covalent cross-link (bridge) by two amino acid cysteine to form a cystine
Can be intrachain or interchain
Is reversible (depending on the oxidizing or reducing environment)
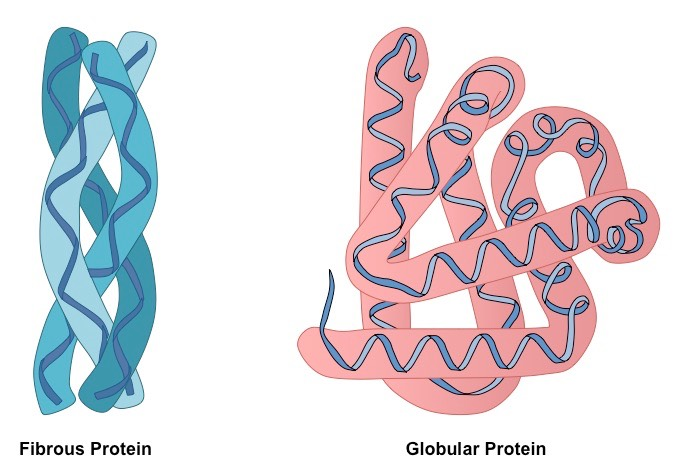
What are the major classes of tertiary structure?
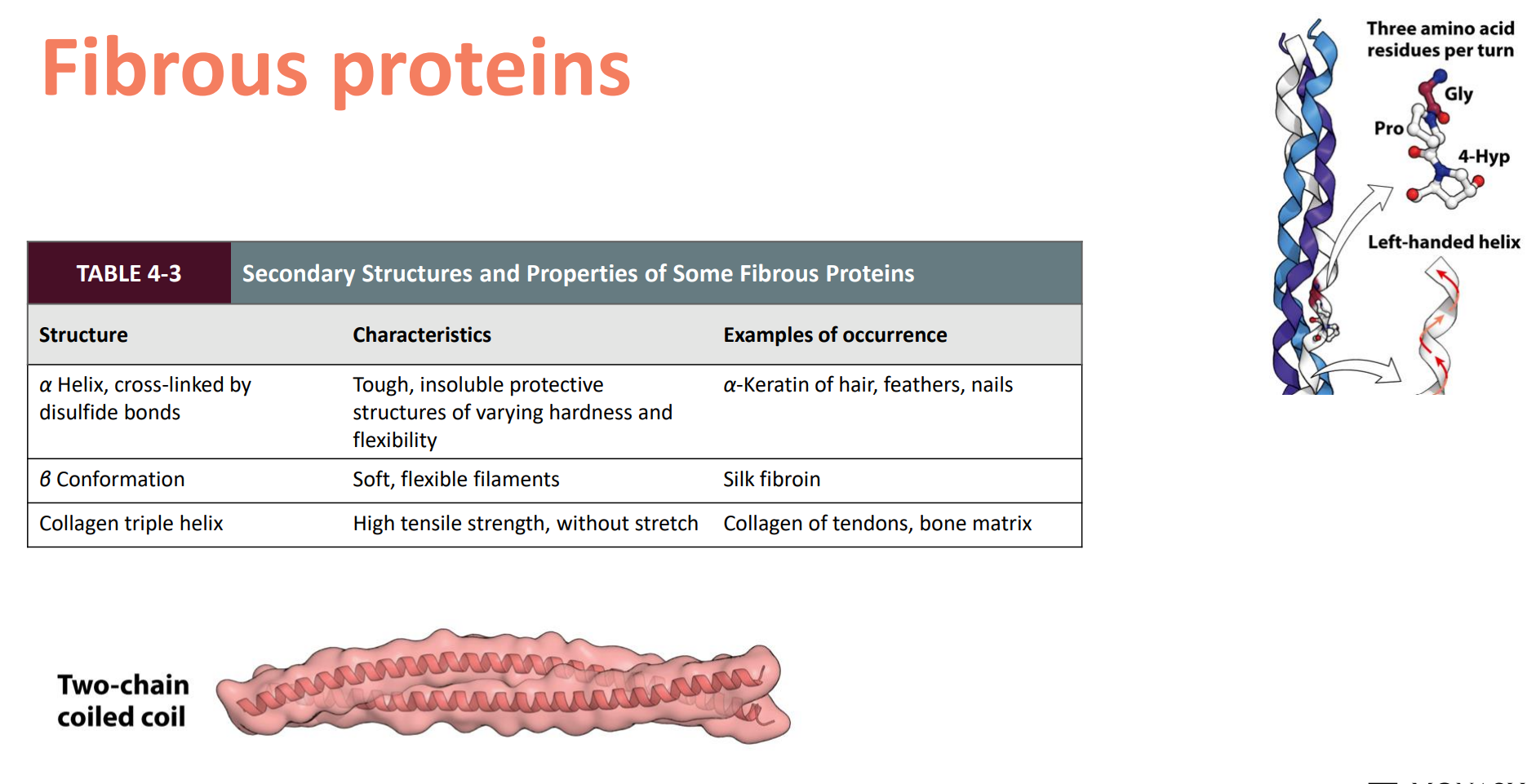
Fibrous — Simple, (usually) repeating secondary structure
Often function in structure for support, shape, protection
Hydrophobic
Keratin, Collagen
Globular — Several types of secondary structure, more folded, more compact
Most other stuff…regulatory proteins, enzymes
Myoglobin, Haemoglobin

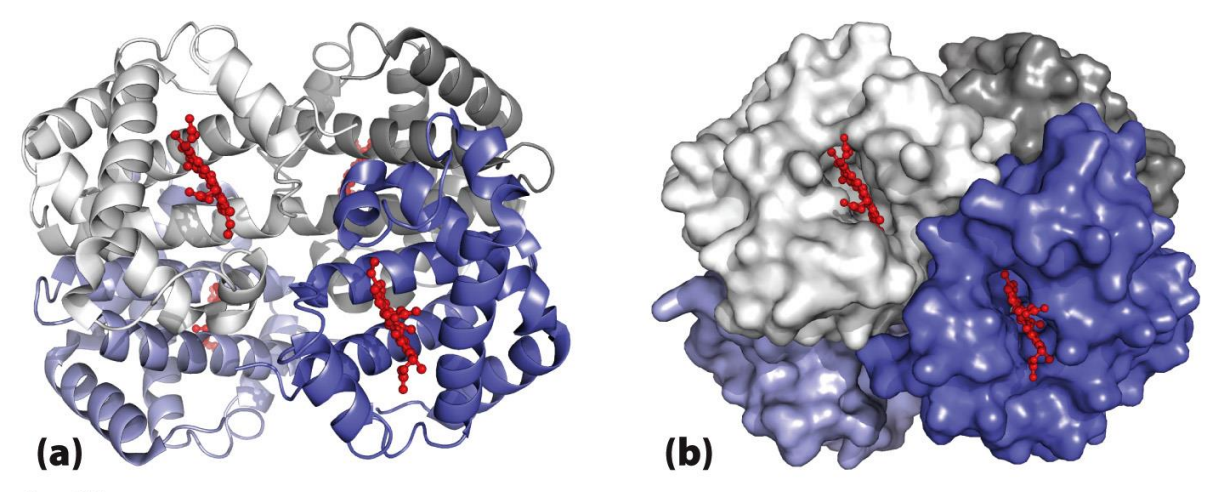
What is Quaternary structure?
A protein with two or more separate polypeptide chains (subunits) - includes 2 (dimer), if 3 it would be a trimer
Subunits can be identical (homodimer) or different (heterodimer)
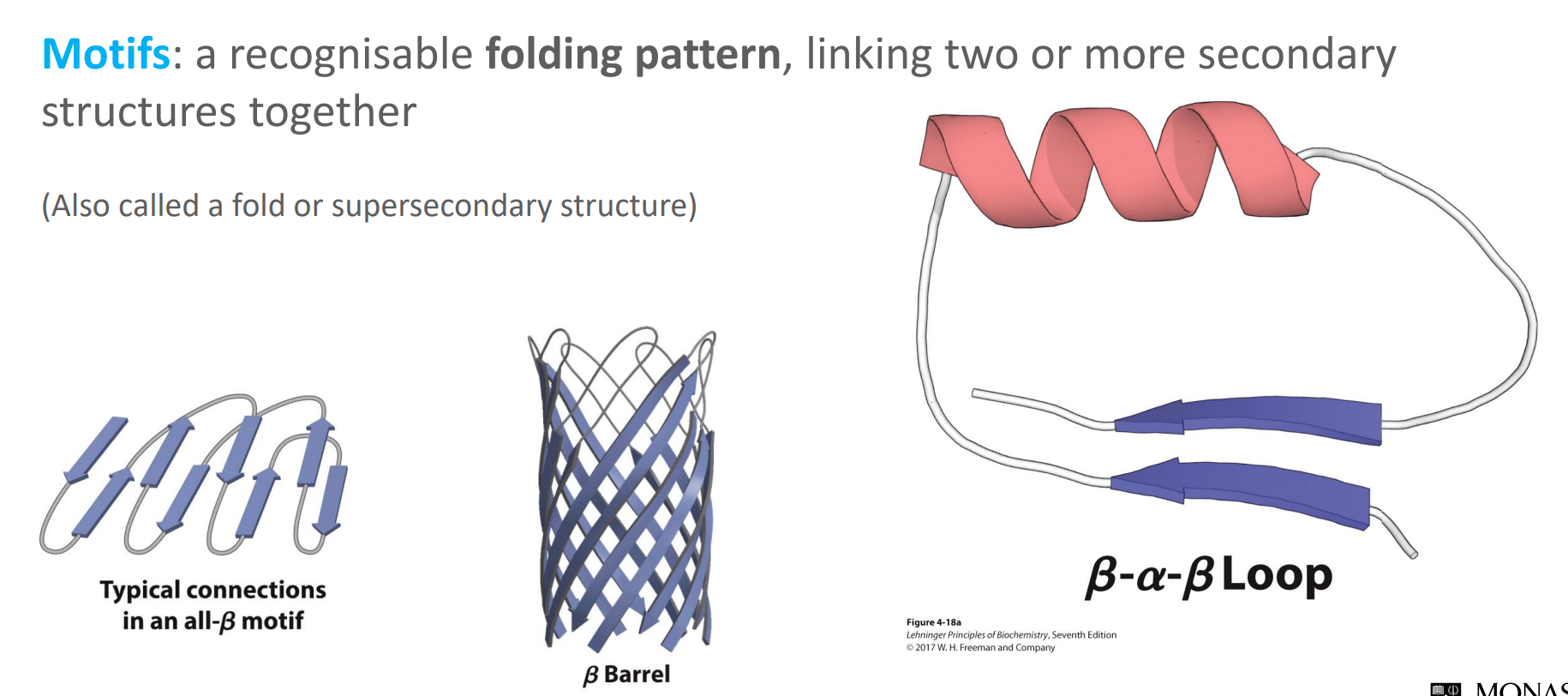
What are protein motifs?
They are a recognisable folding pattern, linking two or more secondary structures together
Also called a fold or supersecondary structure
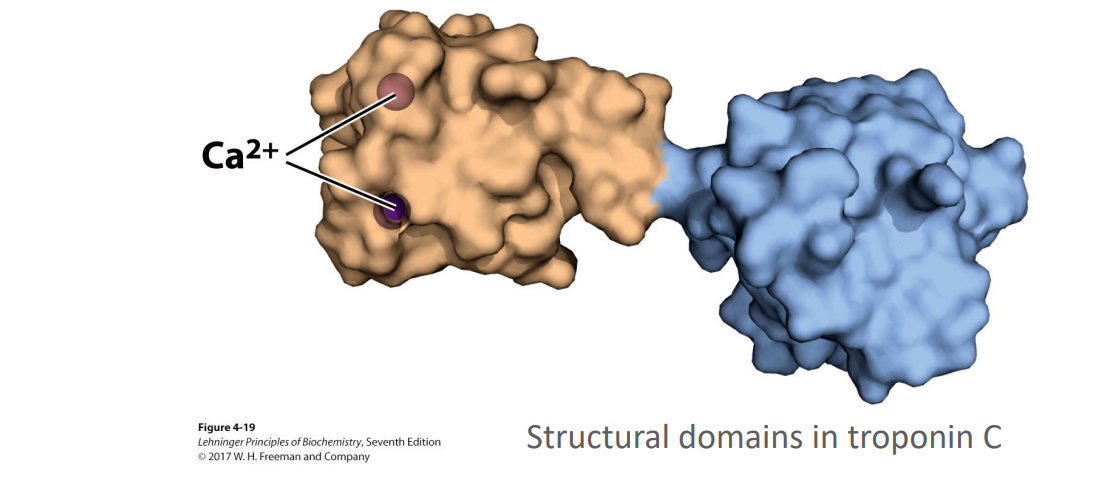
What are protein domains?
These are independently stable parts of a polypeptide chain, or those that can undergo movement as a single entity with respect to the protein
For example the calcium binding domain of this protein
E.g. 1 domain → active site, 2nd domain → the active/inactive binding site (imagine tryptophan regulator protein)
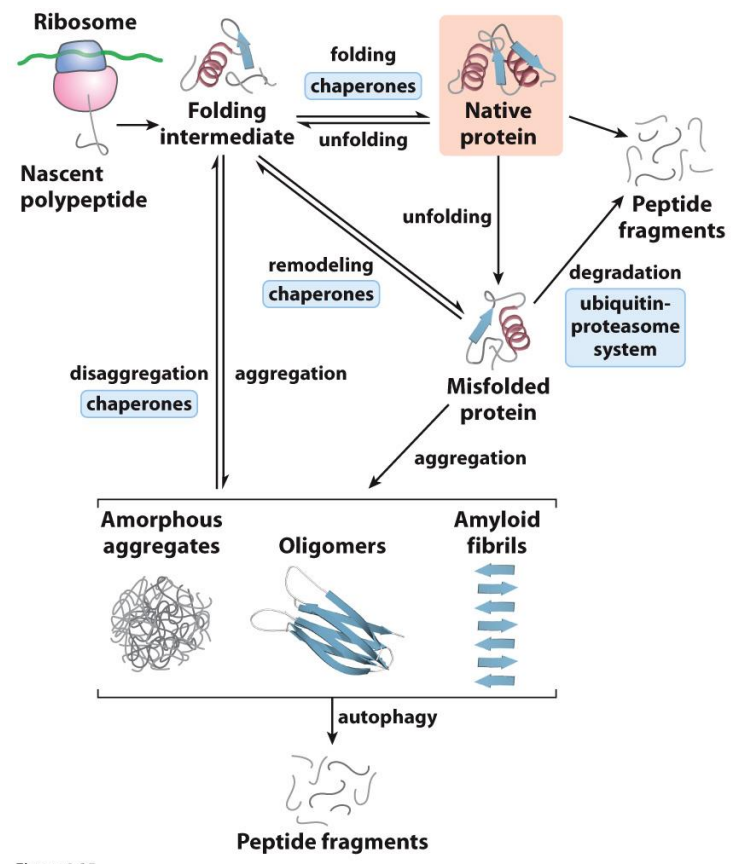
What happens when proteins don’t fold correctly?
Loss of function
Aggregation (Parkinson’s, Alzheimer’s)
These are all in equilibrium systems
Autophagy is when the protein eats itself to try and stop this aggregation
What are the requirements for a protein to fold into its native conformation?
Experiments conducted by Christian B. Anfinsen (1916-1995)
He used RNAase A and questioned:
Is the native conformation of RNAase A required for it to be functional?
What are the requirements for RNAase A to fold into its native conformation?
NOTE the correct disulphide bonds are required

What factors influence the NATIVE formation of the native conformation of proteins?
Disulphide bonds (covalent, so strong and stable)
Weak interactions:
Non-covalent; weak, but overall very stabilising (reversible → breaks and forms over and over again until he most stable form is formed)
Hydrogen bonds, electrostatic interactions, Van der Walls
Hydrophobic effect – burying of hydrophobic groups
Enthalpy
Formation of bonds/interactions decreases enthalpy of a system (favorable; happens spontaneously)
As bonds are formed they release energy into order to stablise the protein → decreases enthalpy
Entropy
Protein structure is a more ordered state (low entropy), so entropy opposes formation of interactions (but … it’s all about overall change in free energy) INCREASE
Protein fold via stepwise pathways?
Secondary structure elements form first
These then fold against each other (various intermediates along the way)
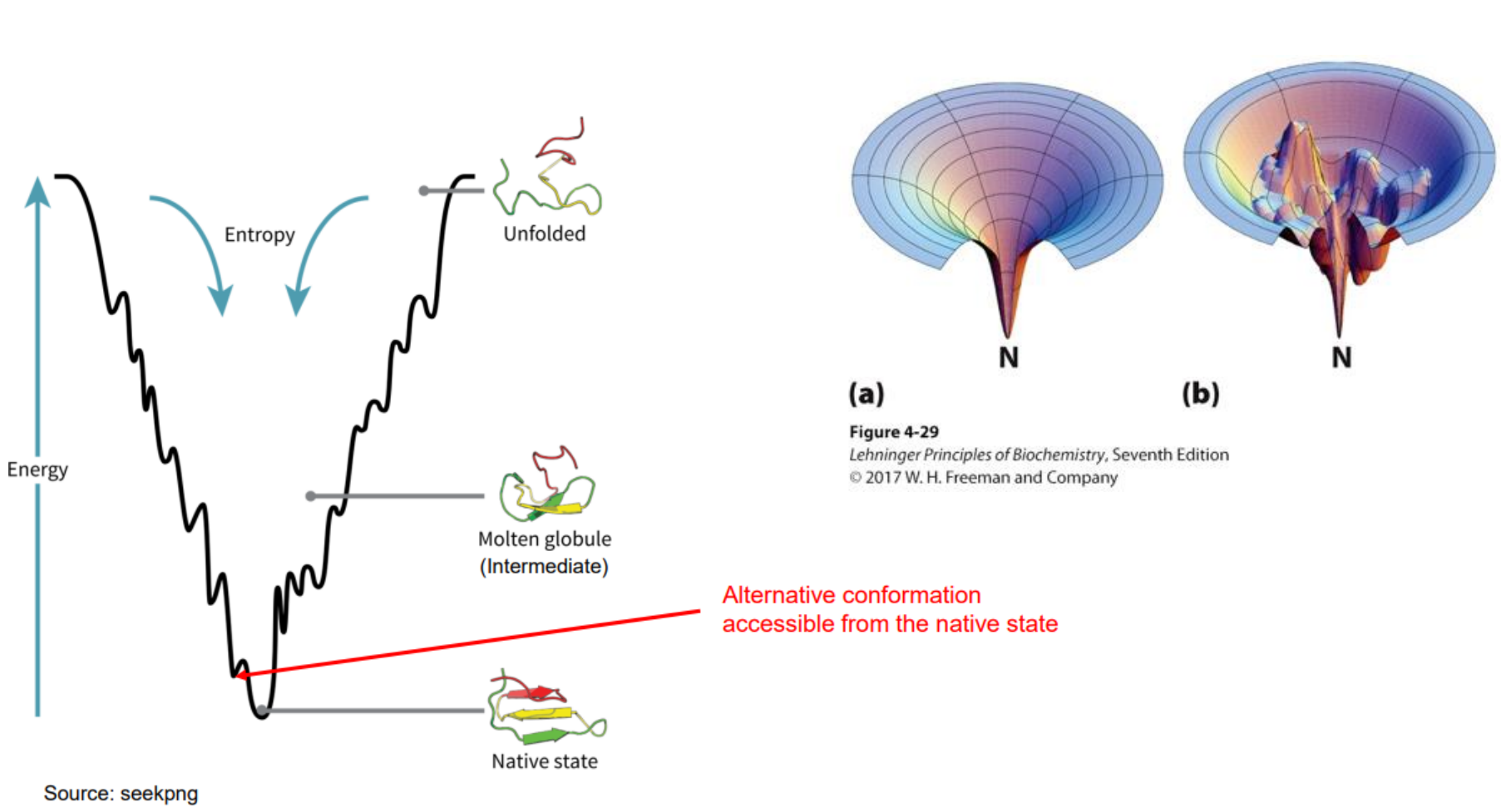
Protein folding and Entropy
A long, floppy unfolded protein has lots of possible shapes → high entropy
When it folds into one specific shape, it becomes more ordered → entropy decreases
Nature hates losing entropy.
So:
Folding decreases entropy = bad for folding
This is why entropy “opposes” folding
When hydrophobic amino acids get buried inside the protein, water molecules are freed from their ordered cages around them.
This increases the entropy of water, which is a huge driving force for folding
So even though the protein becomes more ordered, the water becomes less ordered, and that helps folding happen.
Examples of Fibrous proteins
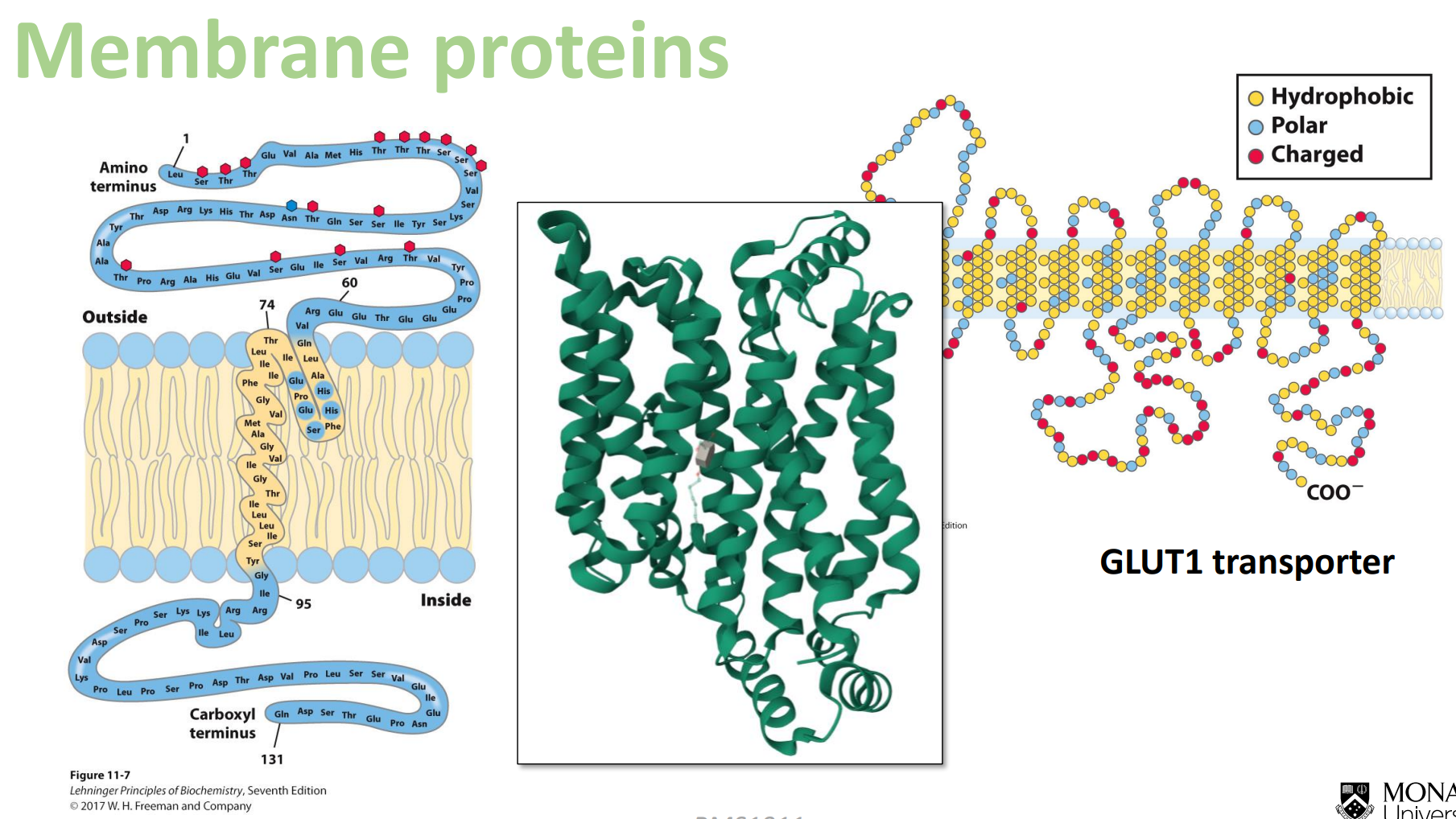
Membrane proteins
Look at how when you approach the hydrophobic fatty acid tails, the amino acids are hydrophobic in the middle, and then on the top and the bottom we have the polar amino acids again
Globular protein - Myoglobin
Found mainly in muscle tissue, where its role is to bind to oxygen (delivered to tissue via hemoglobin in the blood) and supply it to mitochondria
It's quite a small molecule. It only has a single polypeptide chain of 153 amino acids. TERTIARY STRUCTURE
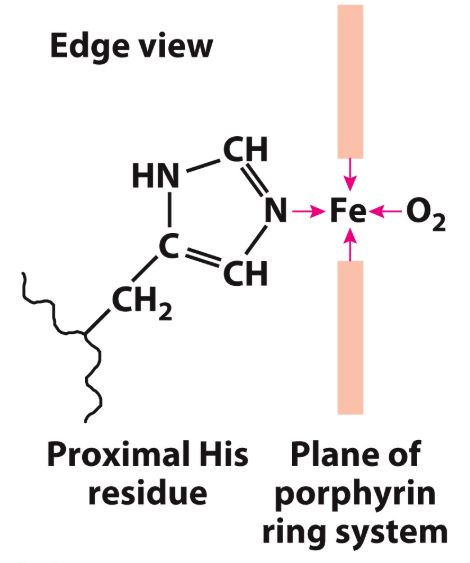
The structure has 8 α-helices, along with a pocket that binds porphyrin ring (shown in red) that has an iron bound at its centre
The iron is what binds oxygen
This iron-bound porphyrin ring is known as "heme" and is technically a "prosthetic group"(A prosthetic group is a compound permanently associated with a protein that helps with the protein's function.)
It's actually the side chain nitrogen of a histidine (His) amino acid that helps to interact with the heme:
Myoglobin binds oxygen with high affinity, and this property helps it to acquire the oxygen from incoming molecules of haemoglobin
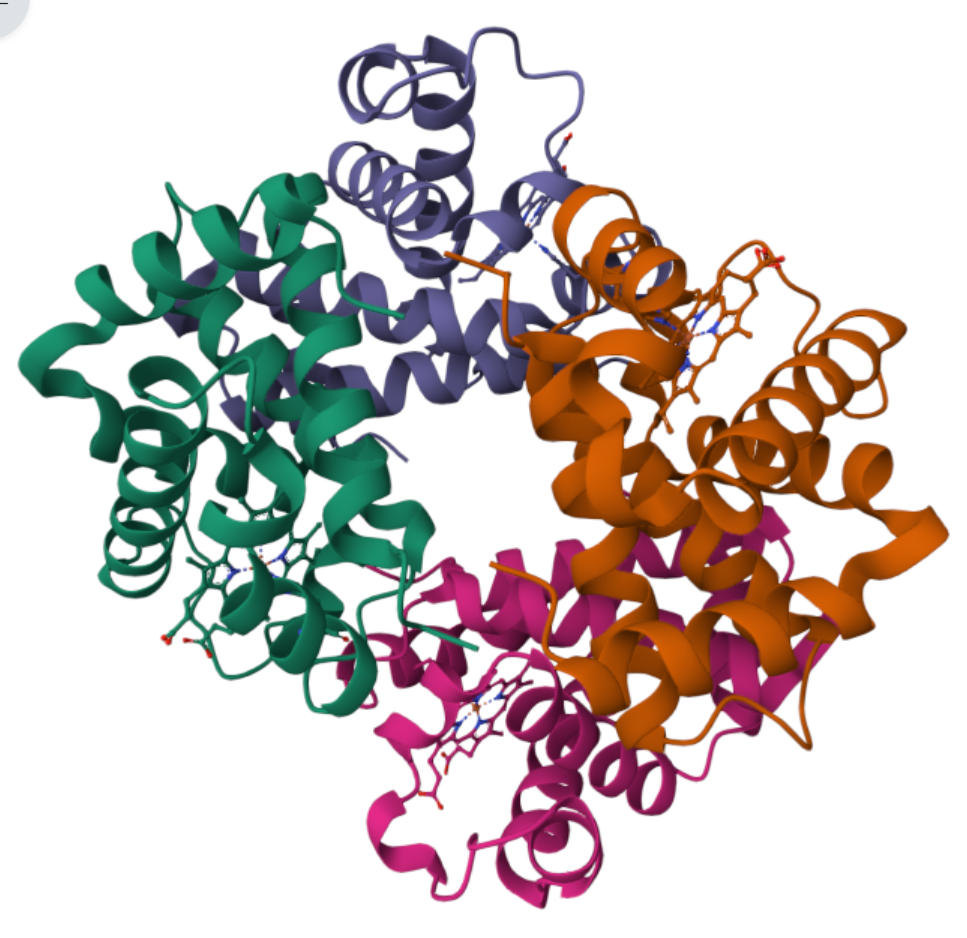
Globular protein - Haemoglobin
QUANTERNARY STRUCTURE
four polypeptide chains (two alpha chains, two beta chains). Each is very similar in sequence and structure to myoglobin.
picks up oxygen from the lungs and transports it to tissues, where it hands it off to myoglobin
Haemoglobin's structure is critical to its function. Binding of oxygen to one subunit increases its affinity for oxygen in the other subunits. This occurs due to structural changes in the molecule.
Without oxygen, haemoglobin is said to be in the "T state" (think "tense"; it's really tense because it's desperate to have oxygen)
When oxygen binds, there is a subtle shift in the conformation of the surrounding amino acids, and it shifts to the "R state" (think "relaxed"; it can breathe a sigh of relief now it has oxygen!)
This is a really subtle change, but it has a knock-on effect to the adjacent subunits, making them more amenable to oxygen binding
This is known as cooperativity, or cooperative binding. Binding of a ligand (in this case oxygen) to one subunit impacts the ability of another subunit to bind. In this case, the impact is positive
What's the point of all this? It means that haemoglobin is more likely to bind oxygen when it is in plentiful supply (in the lungs) but will release that oxygen when it is in an environment of lower oxygen concentration (the tissue). It allows a trade-off in affinity for oxygen, aiding its function.
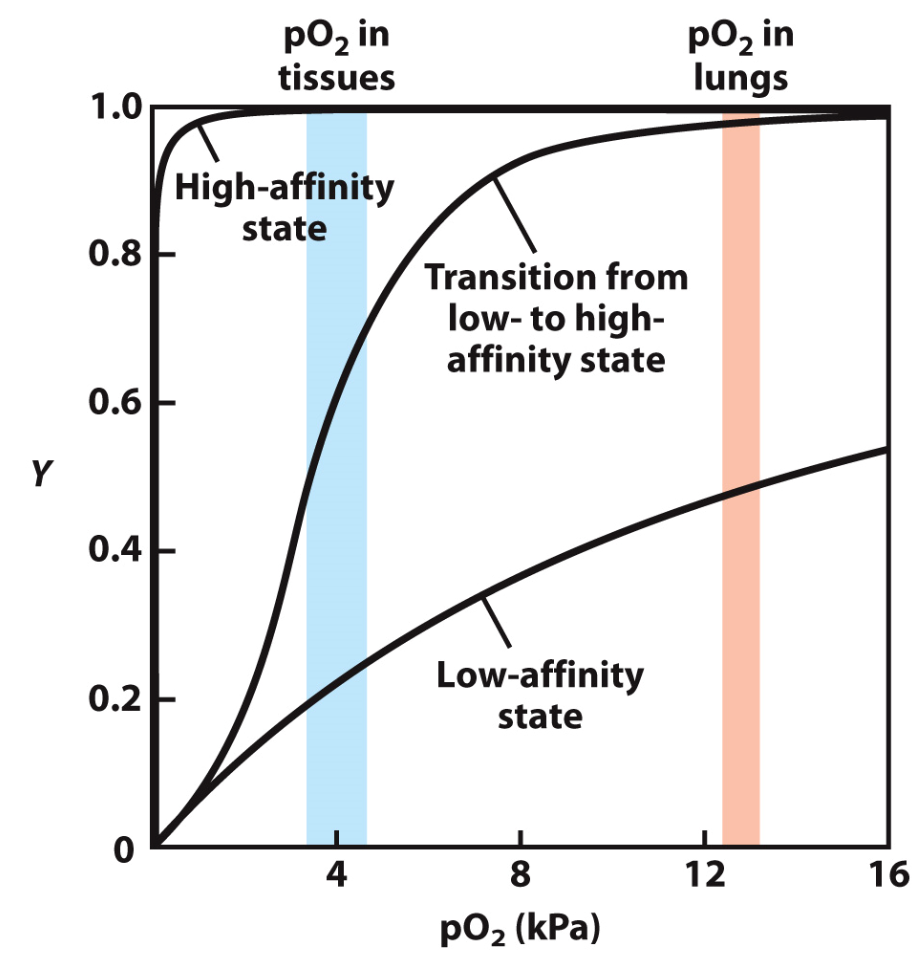
The Graph
The "High-affinity state" line depicts the situation if haemoglobin permanently adopted a high affinity (i.e. a high capacity to bind) oxygen. Even in the low-oxygen environment of tissues it would still be holding on tight to oxygen, preventing it from effectively reaching the cells of the tissue.
The "Low-affinity state" line depicts a situation if haemoglobin always had a low affinity (i.e. a low capacity to bind) for oxygen. If this were the case, it would readily let go of oxygen in tissue (desirable), but wouldn't be able to acquire much oxygen in the lungs (undesirable)
The reality is that due to cooperativity of binding, haemoglobin can shift between high-affinity (in the lungs) to low-affinity (in the tissue), all due to the amount of oxygen in the environment. This is shown in the middle line labelled "Transition from low- to high-affinity state".
What are enzymes and why do we need them?
Enzymes increase the rate of (catalyse) biochemical reactions - they are biological catalysts
They are essential to life - reactions would occur too slowly without enzymes
Many enzymes are part of coordinated pathways, catalysing stepwise reactions
They are highly specific for their substrates
Enzymes are not used up in the process
The majority of enzymes are proteins (globular proteins) (some exceptions such as ribosomes which are RNA not a protein)
The native structure of the protein is critical for the activity of the enzyme
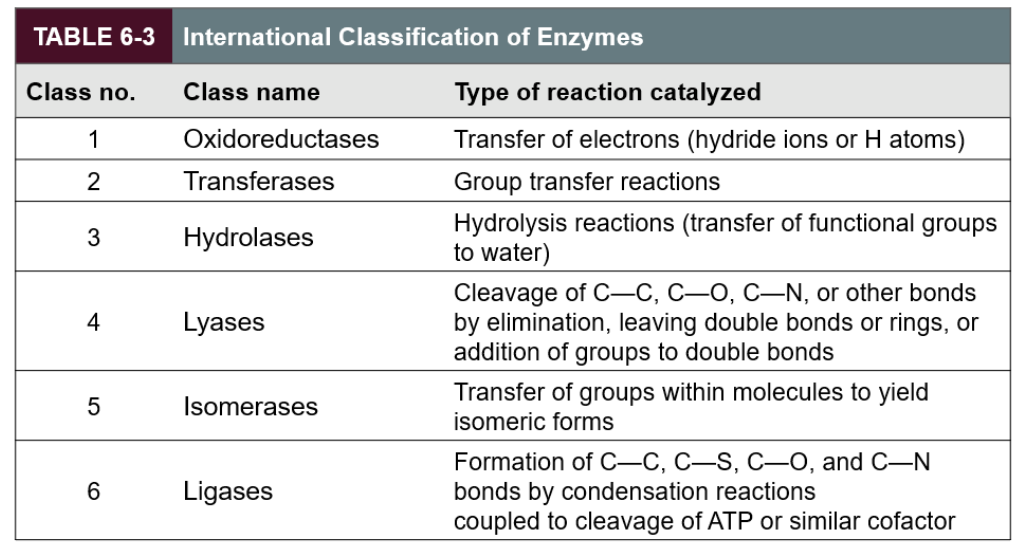
What is the classification of enzymes?
What happens in an enzyme-catalsed reaction?
You have enzyme and substrate
You form enzyme-substrate (ES) complex
Induced fit at the active site
Weak substrate binding, but sufficient enough to be specific
E changes conformation to optimally bind to the transition state
Tighter than substrate binding
Lowers activation energy
You form enzyme-product (EP) complex
Product is bound weakly
You have enzyme and product
E releases P
E is not used up- It can catalyse another round and so on…
*Where old chemical bonds must break and new chemical bonds must form
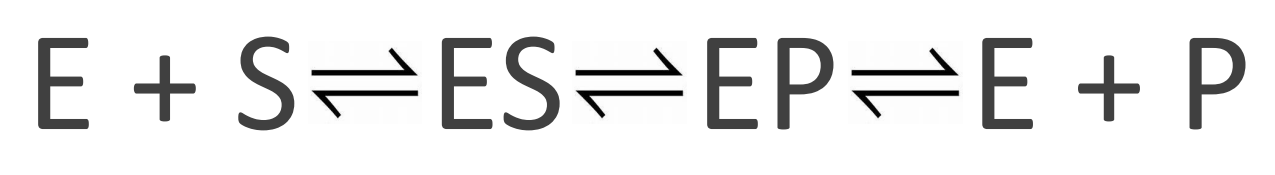
How can a reaction be sped up?
Raise the temperature (gives molecules more energy, which helps to overcome the activation energy barrier)
Lower the activation energy (via enzyme)
Change the pathway (mechanism) of the reaction (via enzyme)
*NOTE THE EQUILIBRIUM DOES NOT NOT NOT CHANGE, will not lead to more or less S/P (depends on environment this happens in)
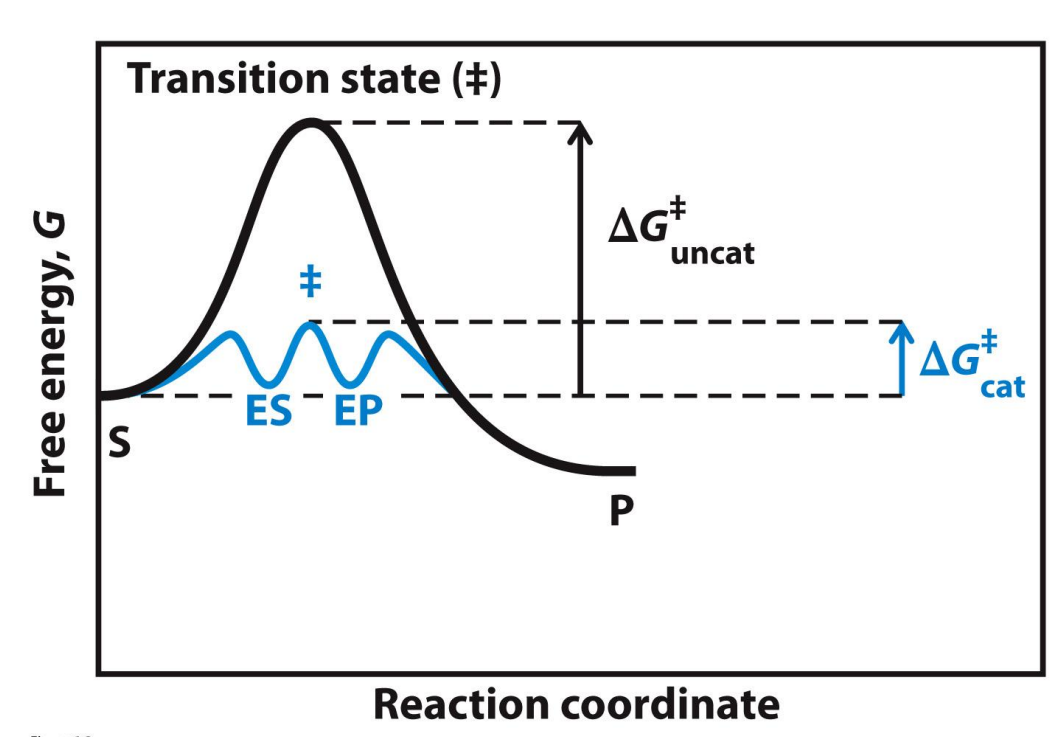
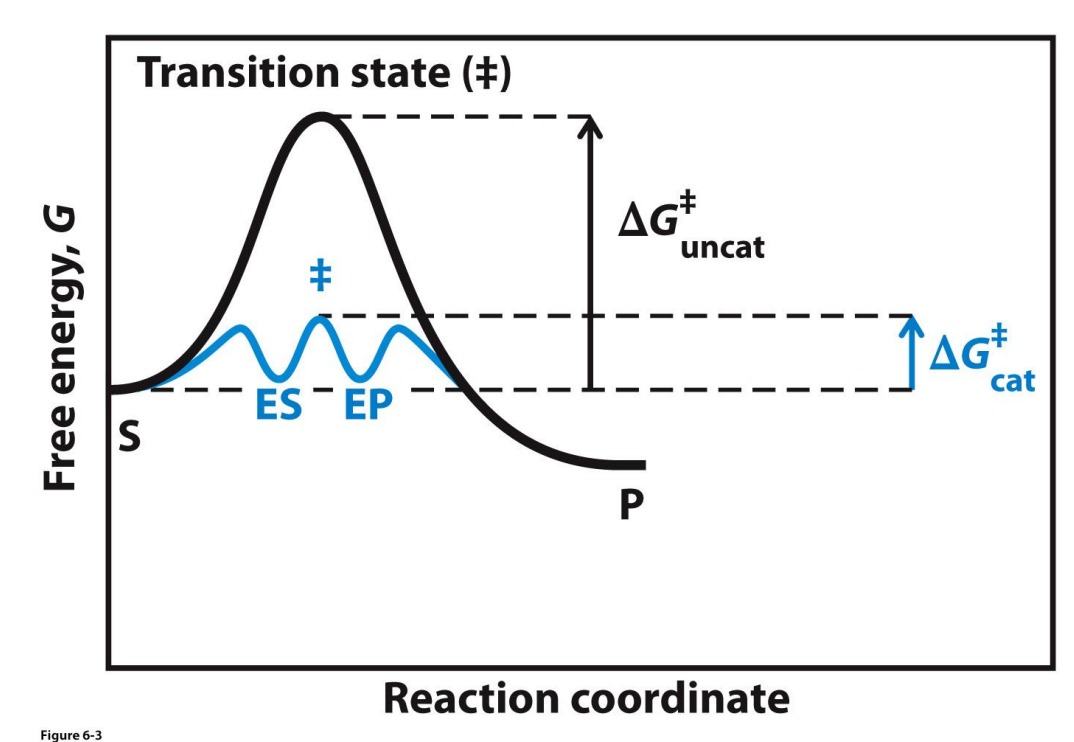
Catalysed vs Uncatalysed reactions
Activation energy has decreased from the uncat to the cat → the catalysed reaction can occur faster
The change in free energy is unchanged (ΔG is the same - not shown here)
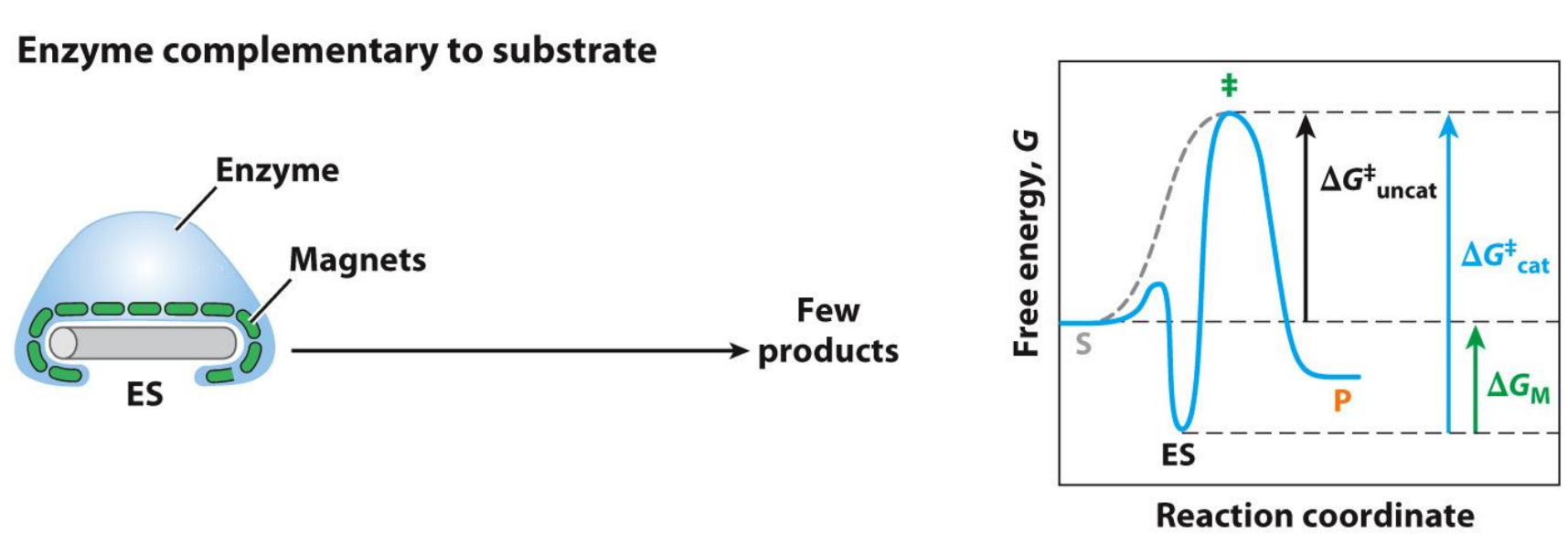
Modelling Catalysed and Uncat reactions on graphs - Stickase
Enzyme needs to be complementary to the transition state not the substrate
Try to think of this as an induced fit model
The problem with the enzyme complementary to the substrate is that you now lock yourself in the ES complex, where the connection between the enzyme and substrate is way too strong and thus requires way more activation energy (free energy change) to enter the transition state - hence the reaction rate is very slow and you can see this on the graphs
Note how the graphs ends and starts at the same points (the ground state of S and P is the same)
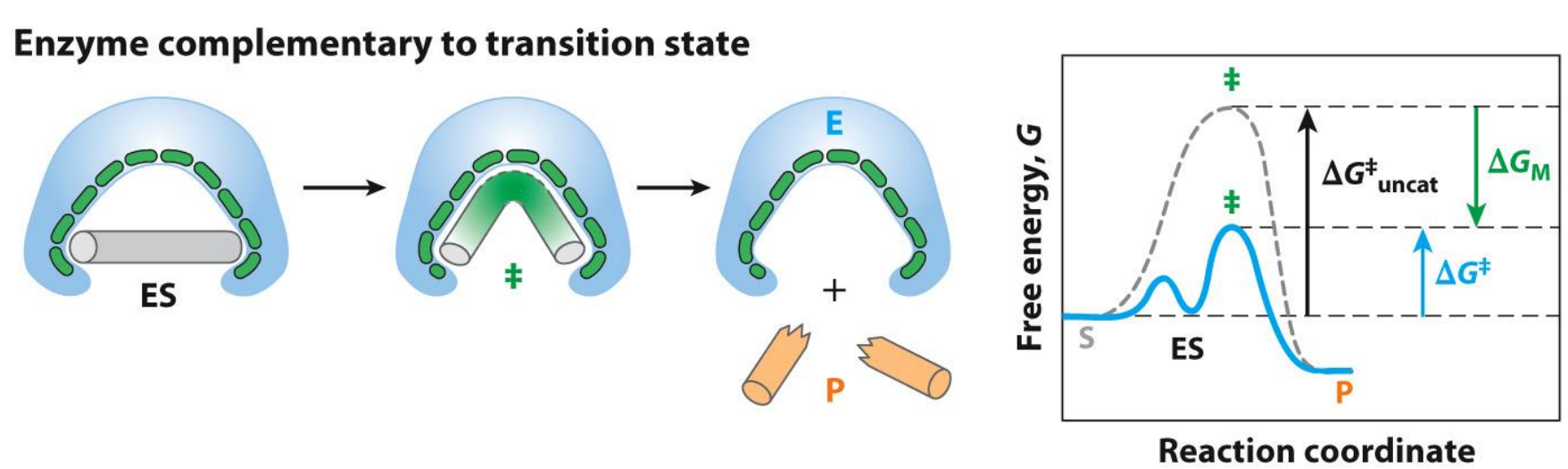
What are some Enzyme Strategies?
Transition state stabilisation — lowers activation energy
Specific binding — allows discrimination of correct substrate
Induced fit — stabilised formation of transition state
Entropy reduction — brings molecules in close proximity
Desolvation — can make functional groups more reactive
Acid-base catalysis — functional groups that can protonate or deprotonate substrate
Covalent Catalysis — temporary formation of covalent bonds
How does Binding interactions drive enzyme reaction?
Weak binding interactions between enzyme and substrate provides driving force for catalysis, especially those formed in the transition state
Such interactions also drive specificity of enzyme reactions
What are some amino acids that often play a role in enzyme active sites?
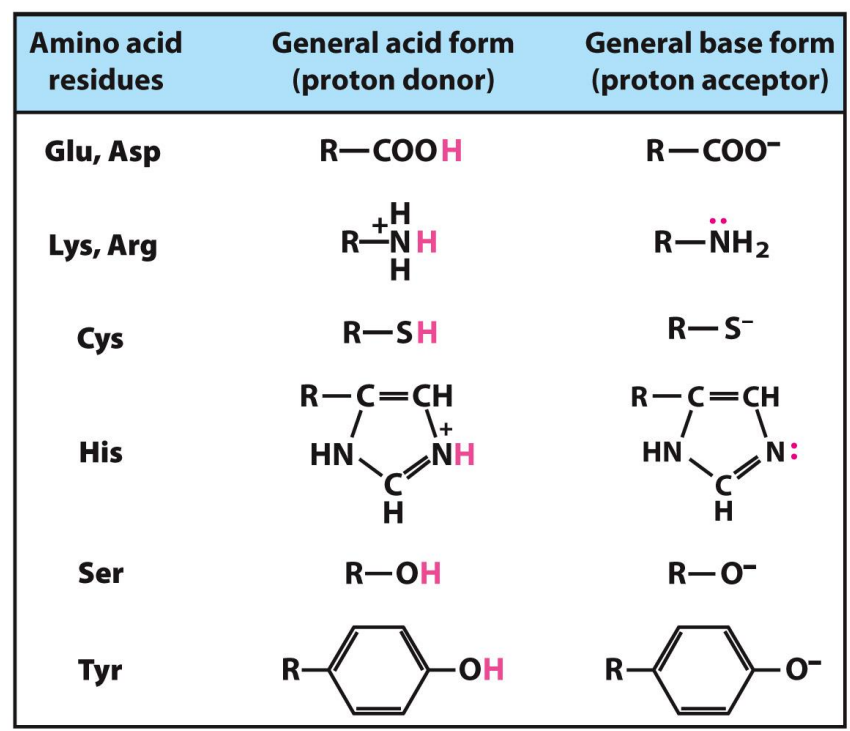
What is a Cofactor and Coenzymes?
A cofactor is one or more inorganic ions (non-protein) that are involved in the active site reaction. For example, Fe2+, Mg2+, Zn3+.
A coenzyme is a larger organic/metalloorganic molecule that acts as a transient carrier of a functional group necessary for the enzymatic reaction.
For example, flavin adenine dinucleotide (FAD) that transfers electrons
A lot of coenzymes derive from vitamins
They temporarily bind to the enzyme and helps transfer atoms or electrons.
*All coenzymes are cofactors, but not all cofactors are coenzymes
What is a Prosthetic group?
A prosthetic group is a coenzyme or metal ion that is covalently bound (sometimes they can be very tightly associated if not covalently bound) to the enzyme itself.
Biotin, derived from vitamin B7, is often found as a prosthetic group; for example, acetyl-coA carboxylase contains covalently-bound biotin (this is a key enzyme in fatty acid synthesis).
How to quantify enzyme kinetics?
Plateaus out as substrate is all used up
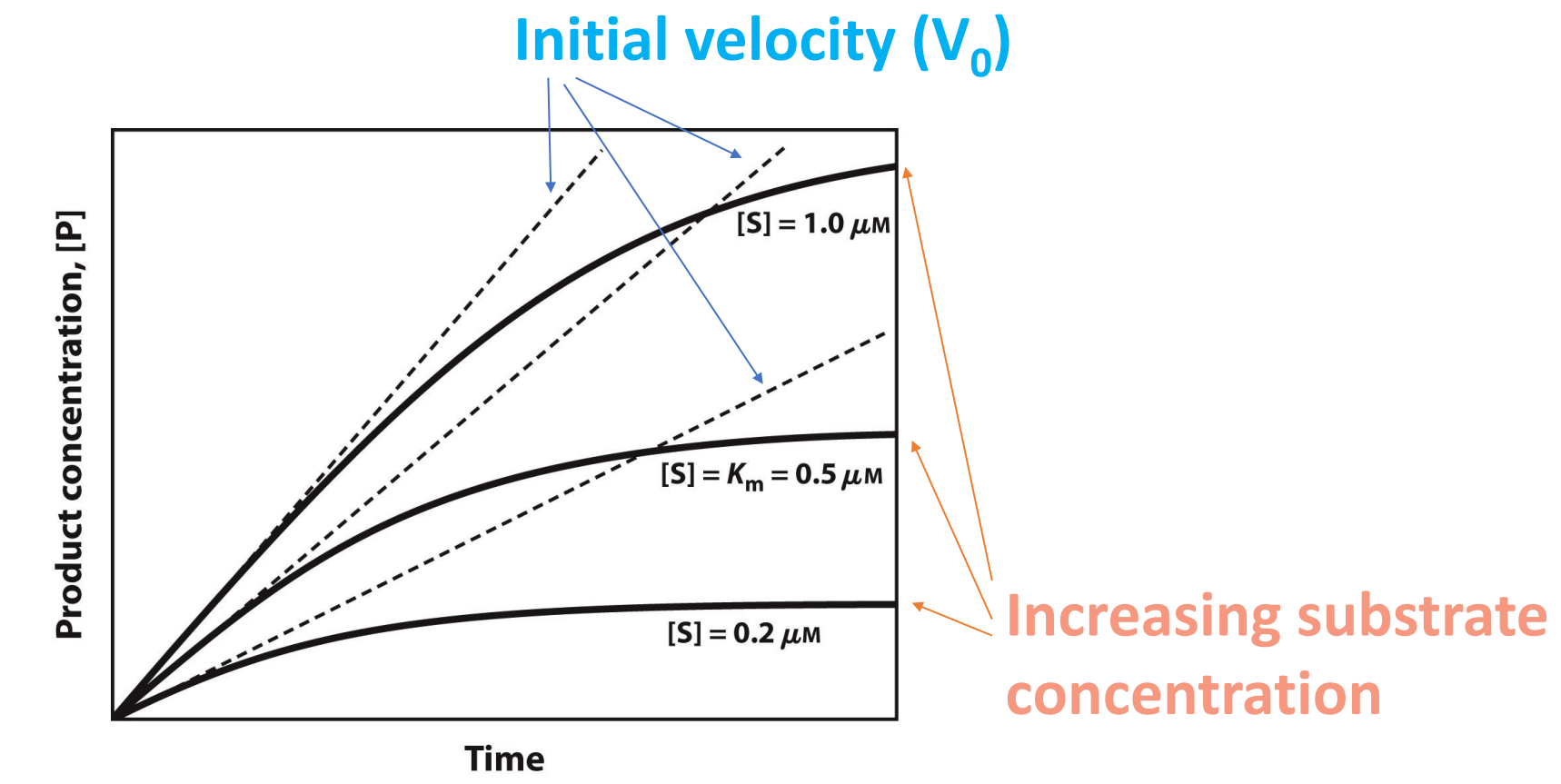
Why is it important to understand Enzyme kinetics?
It gives us an understanding of how enzymes work
Helps us compare between different enzyme
It allows a prediction of rates of reactions
It provides a means to better engineer enzymes with increased/modulated activity
Graphing V0 against [S]
Beware of units
Velocity eventually reaches a max, where all enzyme active sites as used up… cannot theoretically go faster
Vmax is an asymptote, the line will never touch it
Vmax is always proportional to enzyme conc, is we use half enzyme conc, Vmax will also be halved
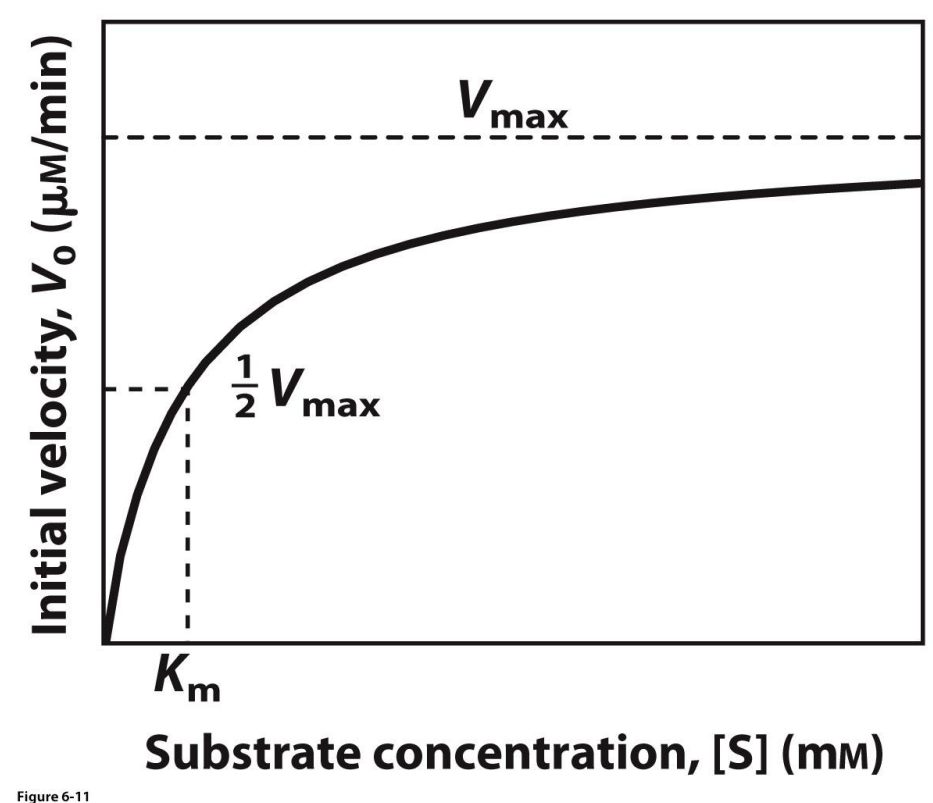
What are some important terms relating to enzyme kinetics?
V0 -- the initial velocity of a reaction at a given substrate concentration. It has units of concentration of product per reaction time.
Vmax -- the maximal velocity for a reaction at a given concentration of enzyme. It has the same units as V0.
Km -- the Michaelis constant. This is the substrate concentration that results in a reaction rate equal to one-half Vmax. Km is a constant. It has units of substrate concentration.
Kcat -- a new term I'm introducing now. This is the maximal rate for a given molecule of enzyme at saturation with substrate; it represents the number of substrate molecules converted into product in a given unit of time. Think of a single molecule of enzyme swimming in a vast sea of substrate. Kcat is constant. It's sometimes referred to as an enzyme's turnover rate. It has units of "per time".
How do the terms relating to enzyme kinetics change
V0 -- changes with [S], but eventually begins to plateau as rate approaches Vmax.
Vmax -- this is proportional to the concentration of enzyme (i.e. [E]). So if you keep throwing in substrate but maintain [E], you'll approach Vmax. But if you then double the amount of enzyme then you'll double the theoretical Vmax. So, Vmax scales with [E].
Km -- it's constant! It doesn't change. It's not dependent on [E].
Kcat -- can be calculated if you know Vmax and enzyme concentration. So, it's independent of [E] because Kcat is a measure of the rate of a single molecule of enzyme.
What is the Km Constant?
Km → Michaelis Constant, it is defined by an equation, conceptually is the amount of substrate that gives half Vmax (the substrate concentration at which V0 is one-half Vmax)
V0 = Vmax[S]/Km+[S]
Km is constant! It doesn't change with enzyme concentration
A lower Km essentially means the enzyme is more efficient. (Sometimes you might see Km referred to as a measurement of enzyme affinity -- i.e. the strength of binding -- for its substrate, but this isn't strictly true.)
So, a lower Km means the enzyme requires less substrate to achieve a given reaction rate compared to enzymes with a higher Km
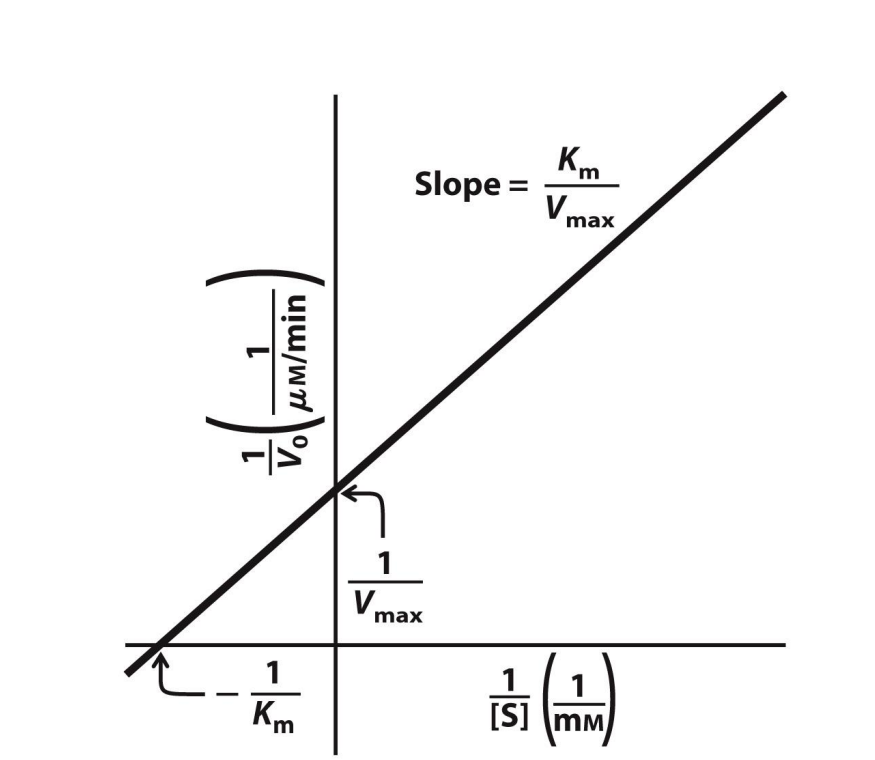
Plotting 1/V0 against 1/[S]
Double reciprocal plot, more technically known as a Lineweaver-Burk plot
Super useful
Y-axis intercept → 1/Vmax
If the y-axis intercept value was 0.01, then Vmax = 1/0.01 = 100 µM per min
X-intercept → -(1/Km)
If the x-axis intercept value was -0.5, then Km = -(1/-0.5) = 2 mM
Slope → Km/Vmax
*The units are given as one OVER whatever the original units were
What is enzyme inhibition?
The action of enzymes can be inhibited
Two broad classes of inhibition: irreversible and reversible
Irreversible – results in permanently non-reactive form of enzymes (often via covalent binding of inhibitor) (stops reaction) (e.g. Aspirin)
Reversible – inhibitor binds reversibly; action of enzyme can be restored after removal of inhibitor (slows down reaction)
Many pharmaceutical agents are enzyme inhibitors
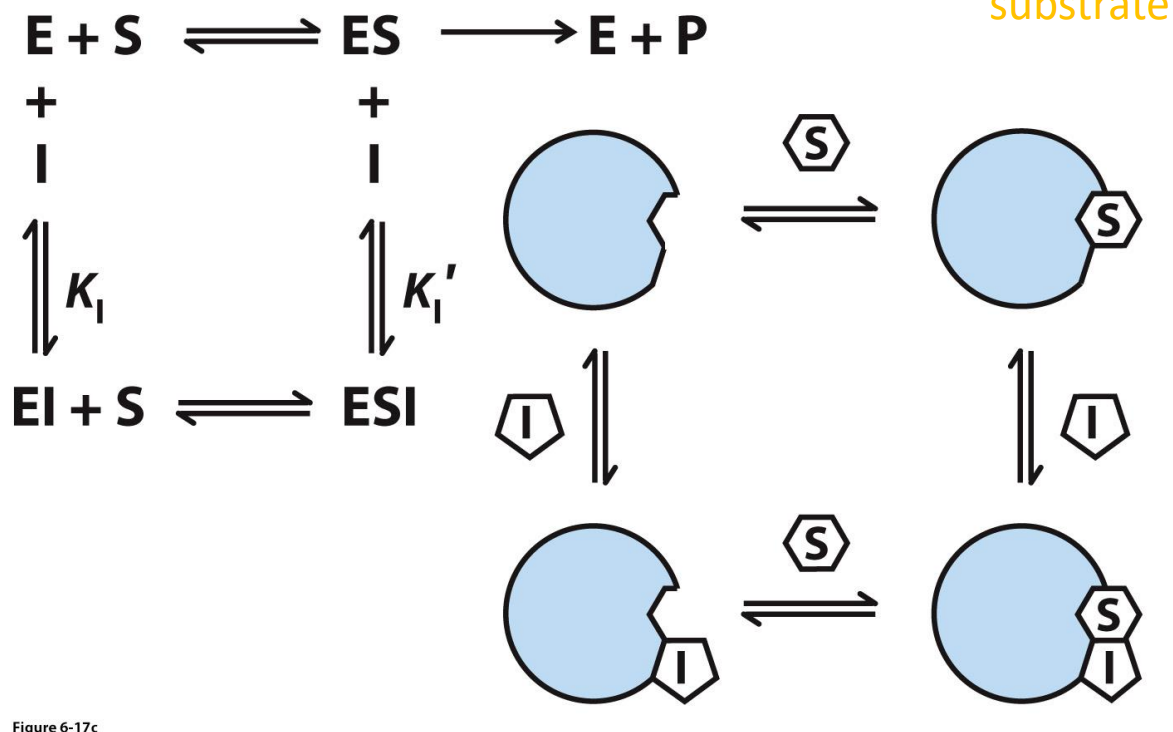
Competitive inhibitor
Inhibitor competes with substrate and binds to the ACTIVE SITE
Statins that reduce cholesterol are considered a Reversible Competitive inhibitor
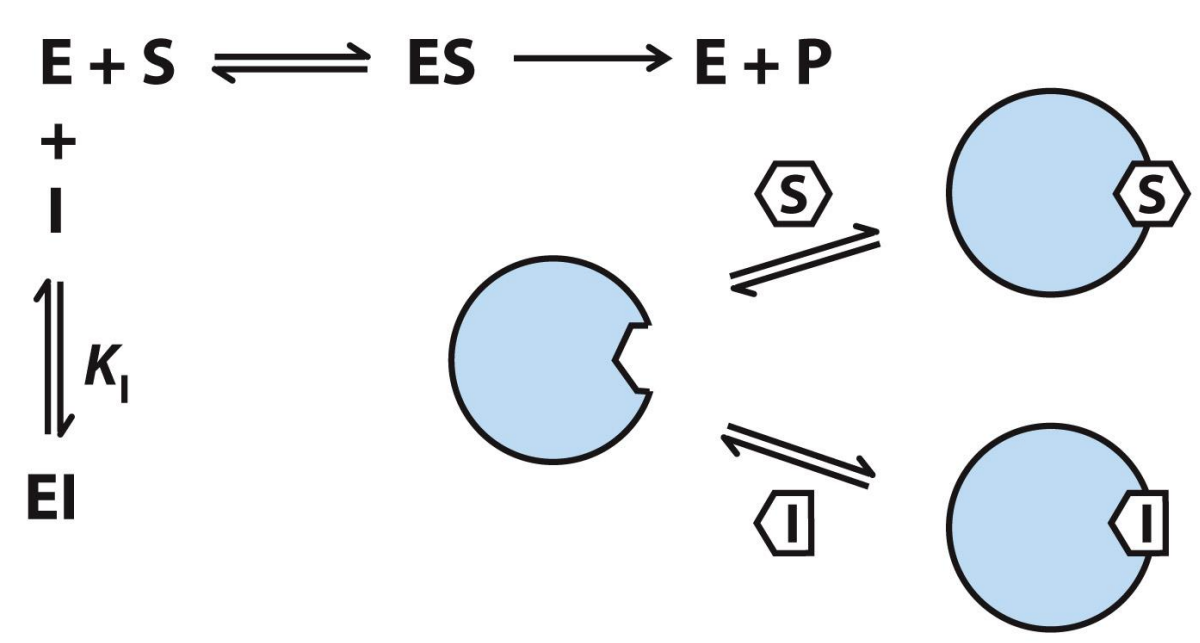
Uncompetitive inhibition
Inhibitor binds to separate site on ONLY the enzyme-substrate complex
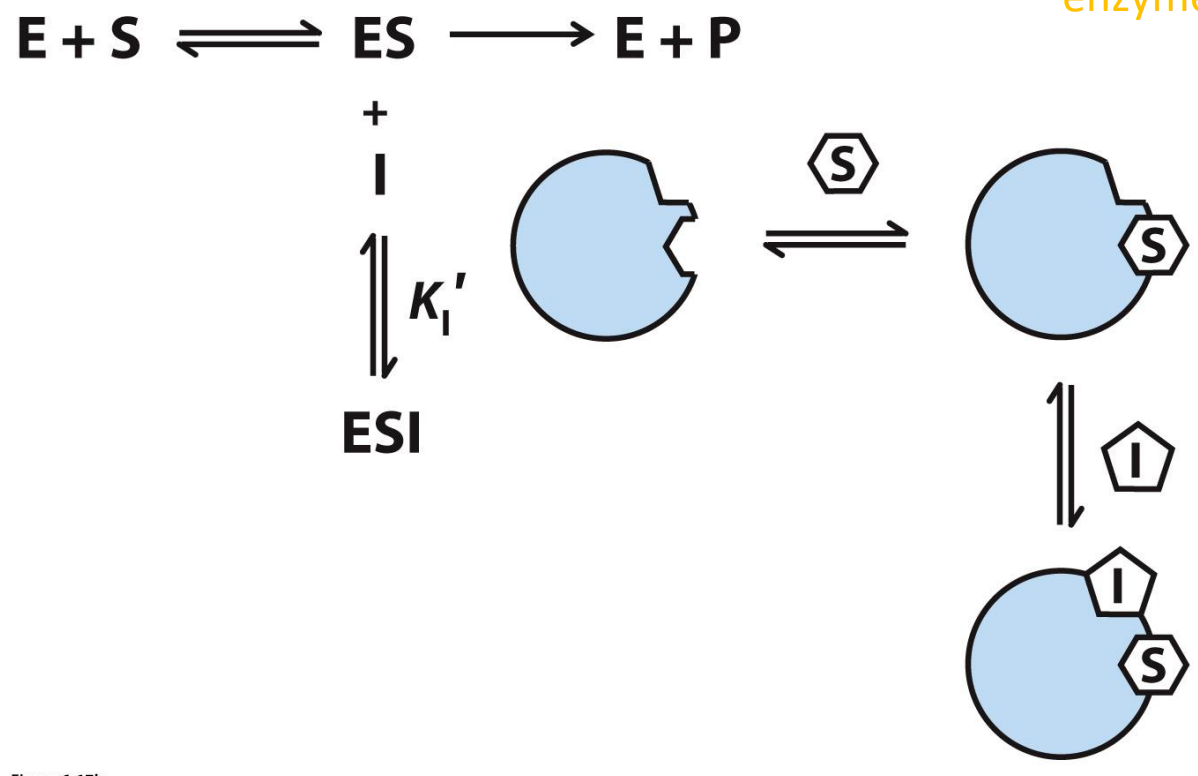
Mixed inhibition
Inhibitor binds to separate site on either the enzyme or enzyme-substrate complex
Competitive Inhibitor graphs
Remember, in competitive inhibition, the inhibitor (I) is competing with the substrate. Shovel enough substrate in and you'll eventually out-compete the inhibitor
Thus, Vmax doesn't change. We can easily see that in the graph because the lines all intersect the y-axis at the same point
However, because you need more substrate to achieve Vmax, the apparent Km (denoted αKm) changes
Note that as the concentration of inhibitor increases, the x-axis intercept shifts to the right; because this represents -1/Km then this means that shifting to the right results in a larger Km
Plug in some entirely theoretical numbers to check things:
Without inhibitor, if the x-axis intercept was -2 (assume the units are 1/µM as per the figure), then that means the Km would be = -(1/-2) = 0.5 µM
With inhibitor, if the x-axis intercept was -0.25 (i.e. shifted to the right along the axis), then that means the apparent Km would be = -(1/0.25) = 4 µM
Therefore, the apparent Km increases as [I] increases
![<ul><li><p>Remember, in competitive inhibition, the inhibitor (I) is <em>competing</em> with the substrate. Shovel enough substrate in and you'll eventually out-compete the inhibitor</p></li><li><p>Thus, <span style="color: rgb(238, 189, 149);"><strong><em>V</em><sub>max</sub> doesn't change</strong></span>. We can easily see that in the graph because the lines all intersect the y-axis at the same point</p></li><li><p>However, because you need <em>more</em> substrate to achieve <em>V</em><sub>max</sub>, the <strong><em>apparent</em> <em>K</em><sub>m</sub> (denoted </strong><span><strong>α<em>K</em><sub>m</sub>) </strong></span><strong>changes</strong></p></li><li><p>Note that as the concentration of inhibitor increases, the x-axis intercept shifts to the right; because this represents -1/Km then this means that shifting to the right results in a <strong><em>larger</em> <em>K</em><sub>m</sub></strong></p></li><li><p>Plug in some entirely theoretical numbers to check things:</p><ul><li><p>Without inhibitor, if the x-axis intercept was -2 (assume the units are 1/<span>µM as per the figure)</span>, then that means the Km would be = -(1/-2) = <strong>0.5 </strong><span><strong>µM</strong></span></p></li><li><p><span>With inhibitor, if the x-axis intercept was -0.25 (i.e. shifted to the right along the axis), then that means the apparent Km would be = -(1/0.25) = <strong>4 µM</strong></span></p></li><li><p><span><strong><em>Therefore, the apparent </em></strong></span><span style="color: rgb(151, 239, 241);"><strong><em>K<sub>m</sub> increases as [I] increases</em></strong></span></p></li></ul></li></ul><p></p>](https://assets.knowt.com/user-attachments/60cba7fe-8103-45f4-8ebc-681533384ad3.png)
Uncompetitive inhibition graphs
The bottom line represents the situation without inhibitor, then the two lines above it are with increasing inhibitor concentration. Note that both the y-intercept and x-intercept change with increasing [I]
This means that Vmax is decreasing with increasing [I]. (The y-axis is 1/Vmax; therefore a larger Vmax results in a small value on the y-axis.)
At the same time, the x-axis intercept is shifting left, towards a more negative value. This means that the apparent Km is decreasing.
Think about why we see this pattern for an uncompetitive inhibitor: the inhibitor binds the ES complex and so you can think of it as removing free enzyme from the pool
With less enzyme around, the apparent Vmax decreases (recall how Vmax is dependent upon enzyme concentration)
As a consequence, the [S] required to reach half Vmax (i.e. Km) declines
Also think about the km decreasing because as you remove the ES from the equation, the reactions shifts to the right (to maintain equilibrium) and therefore the affinity between E and S increases and thus why Km actually decreases
![<ul><li><p>The bottom line represents the situation without inhibitor, then the two lines above it are with increasing inhibitor concentration. Note that both the y-intercept and x-intercept change with increasing [I]</p></li><li><p>This means that <span style="color: rgb(210, 232, 161);"><strong><em>V</em><sub>max</sub> is <em>decreasing</em> </strong>with increasing [I]</span>. (The y-axis is 1/<em>V</em><sub>max</sub>; therefore a larger <em>V</em><sub>max</sub> results in a small value on the y-axis.)</p></li><li><p>At the same time, the <em><u>x-axis intercept is shifting left</u></em>, towards a more negative value. This means that the <span style="color: rgb(194, 127, 229);"><strong><em>apparent</em> <em>K</em><sub>m</sub> is decreasing</strong></span>.</p></li><li><p>Think about why we see this pattern for an uncompetitive inhibitor: the inhibitor binds the ES complex and so you can think of it as <em>removing</em> free enzyme from the pool</p></li><li><p>With less enzyme around, the <em>apparent</em> <em>V</em><sub>max</sub> decreases (recall how <em>V</em><sub>max</sub> is dependent upon enzyme concentration)</p></li><li><p>As a consequence, the [S] required to reach half <em>V</em><sub>max</sub> (i.e. <em>K</em><sub>m</sub>) declines</p><ul><li><p>Also think about the km decreasing because as you remove the ES from the equation, the reactions shifts to the right (to maintain equilibrium) and therefore the affinity between E and S increases and thus why Km actually decreases </p></li></ul></li></ul><p></p>](https://assets.knowt.com/user-attachments/1712d424-6836-4ca1-b4bb-020c03bddaab.png)
Mixed Inhibition graphs
Apparent Vmax is decreasing (the y-axis intercept is increasing)
In the image shown, apparent Km is increasing (the x-axis intercept is increasing), but for these types of inhibitors it can actually go one way or the other, depending on the precise action occurring
Why is this the case? Recall that a mixed inhibitor is capable of binding both the free enzyme as well as the ES complex. For Vmax the situation is as for uncompetitive inhibition, where some fraction of [E] is removed entirely due to the inhibitor binding the ES complex
For Km the situation is complicated by whether the inhibitor has a higher affinity for the E or ES complex. In the former, it would be like the competitive example, where more substrate is required to compensate and thus the apparent Km increases
If the inhibitor has a higher affinity for the ES complex then it will decrease Km as per the uncompetitive example shown before.
![<ul><li><p><span style="color: rgb(241, 139, 252);"><strong><em>Apparent V</em><sub>max</sub> is decreasing</strong></span> (the y-axis intercept is increasing)</p></li><li><p><span>In the image shown, </span><em>apparent K</em><sub>m</sub> is <strong>increasing</strong> (the x-axis intercept is increasing), but for these types of inhibitors it can actually <span style="color: rgb(243, 185, 185);"><em>go one way or the other, depending on the precise action occurring</em></span></p></li><li><p>Why is this the case? Recall that a mixed inhibitor is capable of binding both the free enzyme as well as the ES complex. For <em>V</em><sub>max</sub> the situation is as for uncompetitive inhibition, where some fraction of [E] is removed entirely due to the inhibitor binding the ES complex</p></li><li><p>For <em>K</em><sub>m</sub> the situation is complicated by whether the inhibitor has a higher affinity for the E or ES complex. In the former, it would be like the competitive example, where more substrate is required to compensate and thus the <em>apparent</em> <em>K</em><sub>m</sub> increases</p></li><li><p>If the inhibitor has a higher affinity for the ES complex then it will decrease <em>K</em><sub>m</sub> as per the uncompetitive example shown before.</p></li></ul><p></p>](https://assets.knowt.com/user-attachments/8c1f28bc-8ddb-461b-b69b-05c015fbf0d0.png)
What are Regulatory Enzymes?
A lot of enzymes are part of sequential pathways:
Reactions of one enzyme produce products that become substrates for the next enzyme
This is especially prevalent in metabolism
Lots of metabolic pathways have key steps that are regulated by one or more regulatory enzymes
Allosteric enzymes are one type of regulatory enzyme
What are Allosteric Enzymes?
Multi-domain or quaternary structure
Involve reversible, non-covalent binding of regulatory compounds called allosteric modulators (“allosteric effectors”)
These can even be the substrate/product themselves
Binding occurs at a separate site (allosteric site)
Binding of modulator induces conformational change that affects substrate binding (either positively or negatively)
Allosteric can BOTH increase or decrease the rate of reaction (whereas inhibitors only slow or stop reactions)
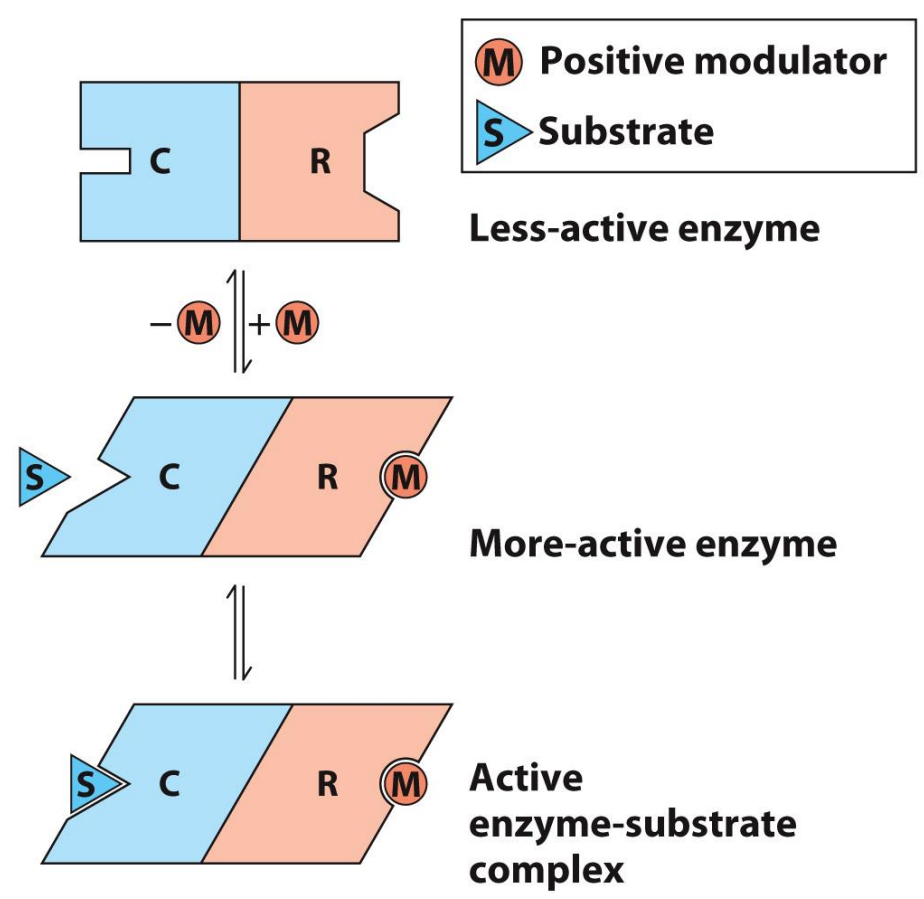
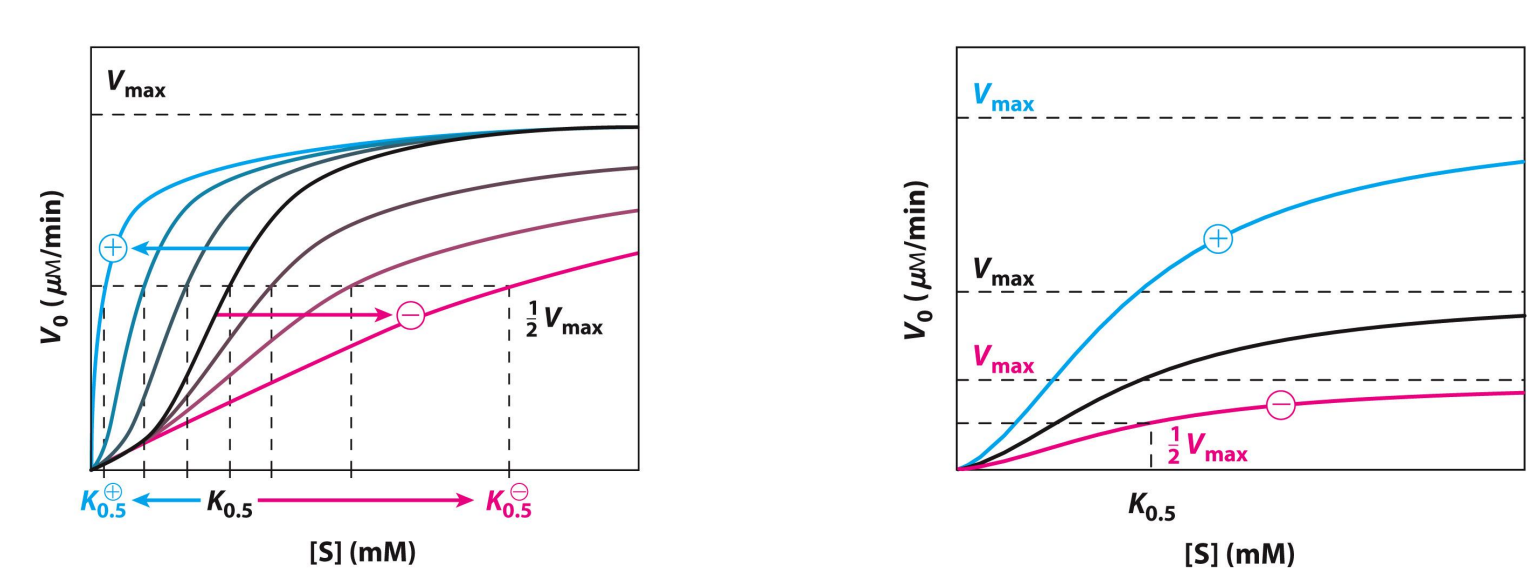
Allosteric Enzyme Kinetics (negative or positive modulator)
Behavior doesn’t obey Michaelis-Menten kinetics
Besides is an example for when the substrate itself is a positive modulator of activity
High-activity R state → Relaxed
Low-activity T state → Tense, looking to bind
K0.5 changes
Km is not given
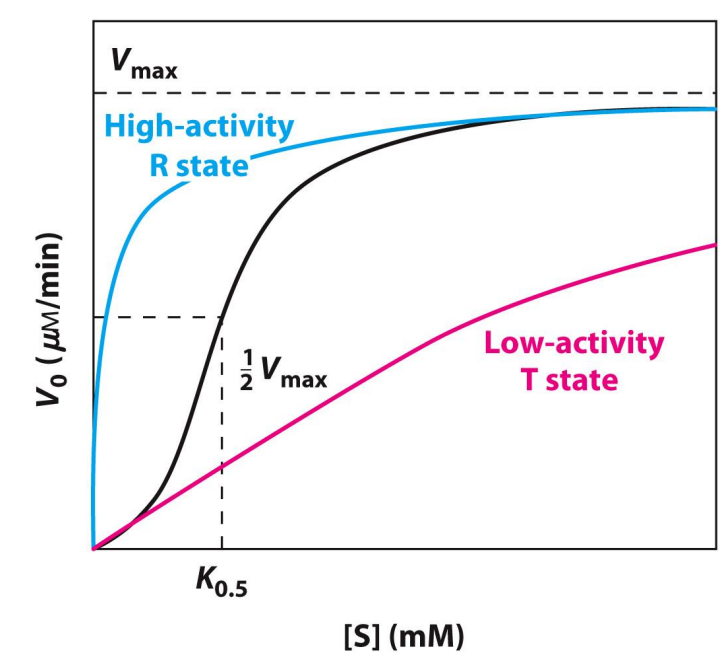
What are Isozymes?
Isozymes are enzymes that catalyse the same reaction but have different primary structures (they’re also encoded by different genes)
A good example of this is hexokinase, which catalyses the phosphorylation of glucose to glucose-6-phosphate
Isozymes - Hexokinase Example
What is Metabolism?
Refers to all the reactions that occur in our body that provide the body with energy to carry out all its bodily processes
Metabolism is defined as the sum of all the chemical reactions taking place in a cell or organism that allows life to be sustained. This highly coordinated activity and includes many reaction pathways, includes:
Obtaining chemical energy from macromolecules
Converting nutrient molecules to cellspecific molecules
Synthesising macromolecules from precursor molecules
Synthesising and degrading cell-specific molecules
A chemical reaction occurs when atoms have enough energy to combine or change bonding partners
Metabolism and Energy
Metabolic reactions involve energy changes
Energy is the defined as the capacity to do work or the capacity for change
In Biochemical reactions, energy; is the capacity for change
Energy changes are usually associated with changes in the chemical composition and properties of molecules

What are the types of Energy?
All forms of energy are either:
Potential energy—energy stored as chemical bonds, concentration gradient, charge imbalance
Kinetic energy—the energy of movement
Energy can be converted from one form to another

What is Bioenergetics?
Is the quantitative study of energy transductions (the changes of one form of energy to another)
The law of thermodynamics (overview)
The laws of thermodynamics apply to all matter and all energy-transforming reactions
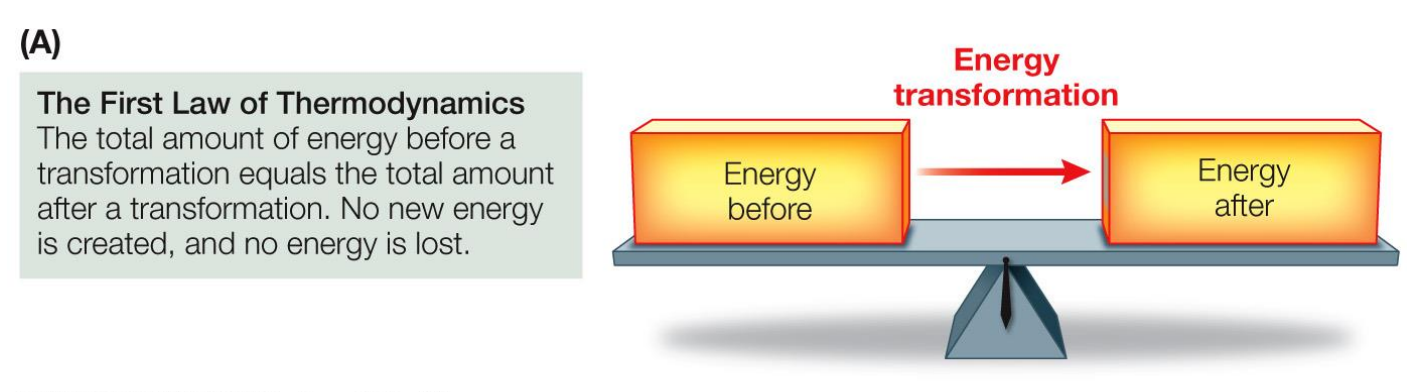
1st Law of Thermodynamics
Energy is neither created nor destroyed
When energy is converted from one form to another, the total amount of energy before and after the conversion is the same
2nd Law of Thermodynamics
When energy is converted from one form to another, some of that energy becomes unavailable to do work
No energy transformation is 100% efficient, some energy is unavailable (due to disorder or entropy)
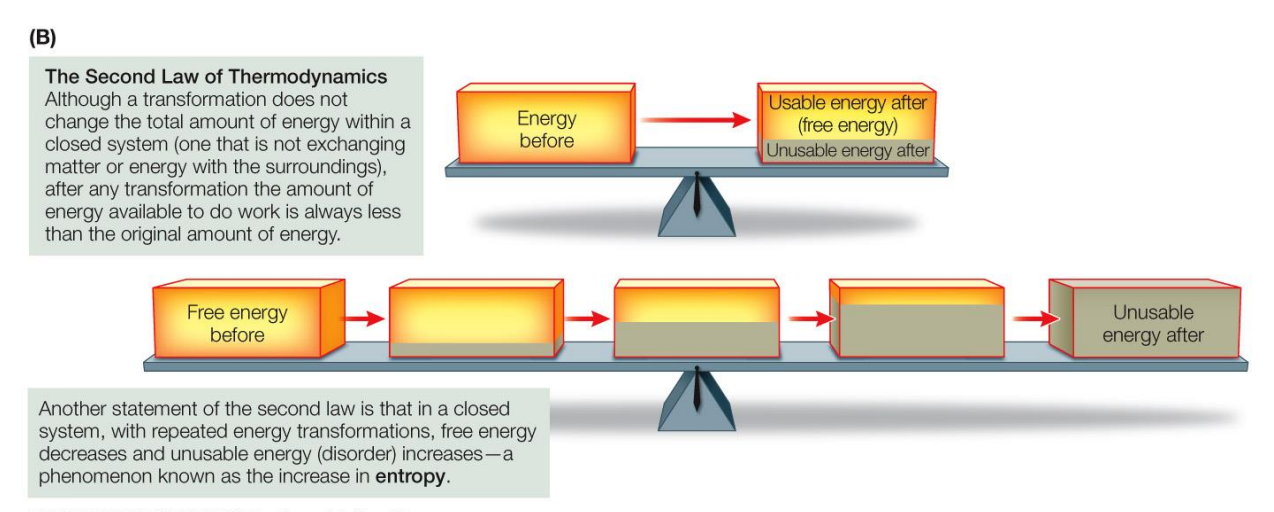
What is Entropy?
Entropy is a measure of the disorder in a system
It takes energy to impose order on a system. Unless energy is applied to a system, it will be randomly arranged or disordered
During physical exercise, muscles convert chemical energy (from food) to mechanical energy (contraction)
Not all of this energy is available to do work, some of it is released as heat energy due to entropy
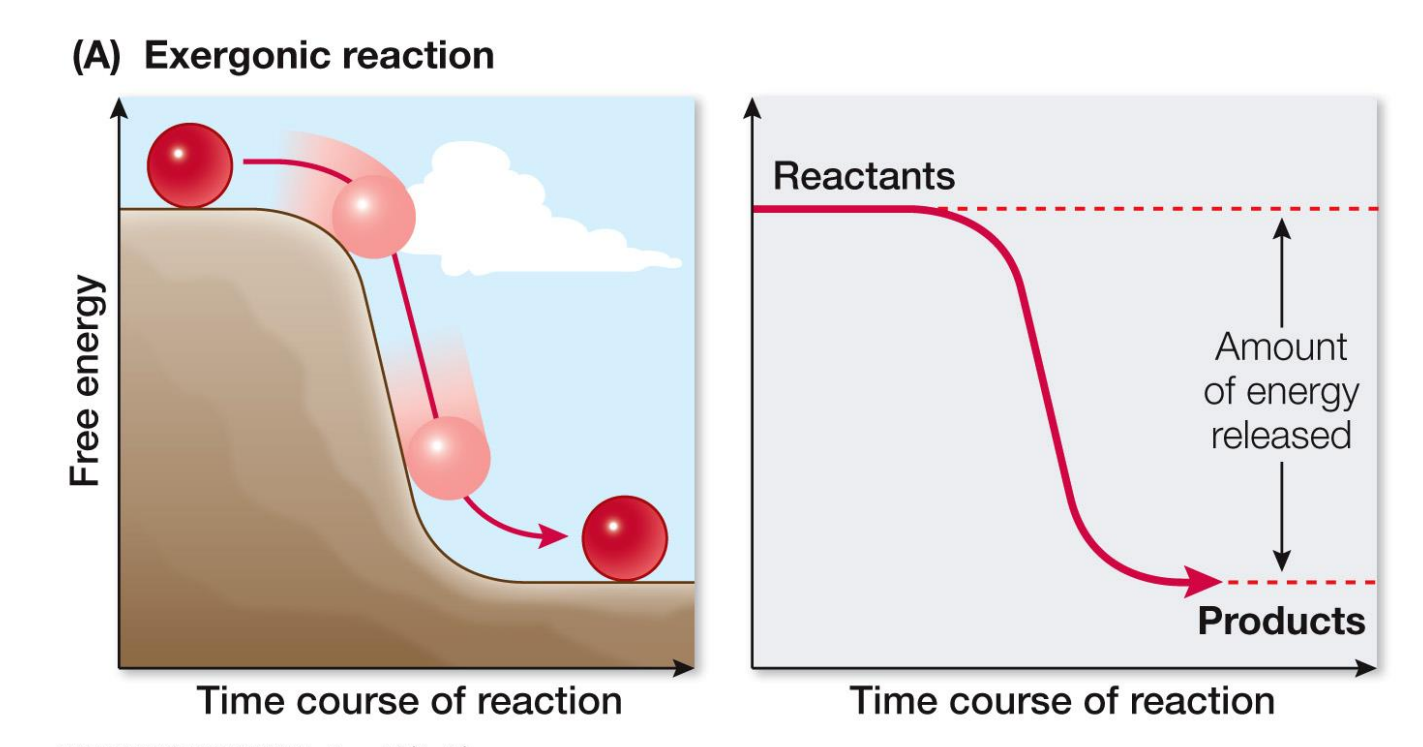
What is Free Energy?
Free energy is a measure of the capacity of a system to do work
We can get a negative, positive or 0 answer this can tell us the type of reaction

What are the types of energy conversions?
If ΔG is –ve, free energy is released (EXERGONIC)
If ΔG is +ve, free energy is required (ENDERGONIC)
If free energy is not available, the reaction does not occur
If ΔG = 0, the reaction is at chemical equilibrium – A <=>B (Concentrations of A and B will determine the direction of the reaction)
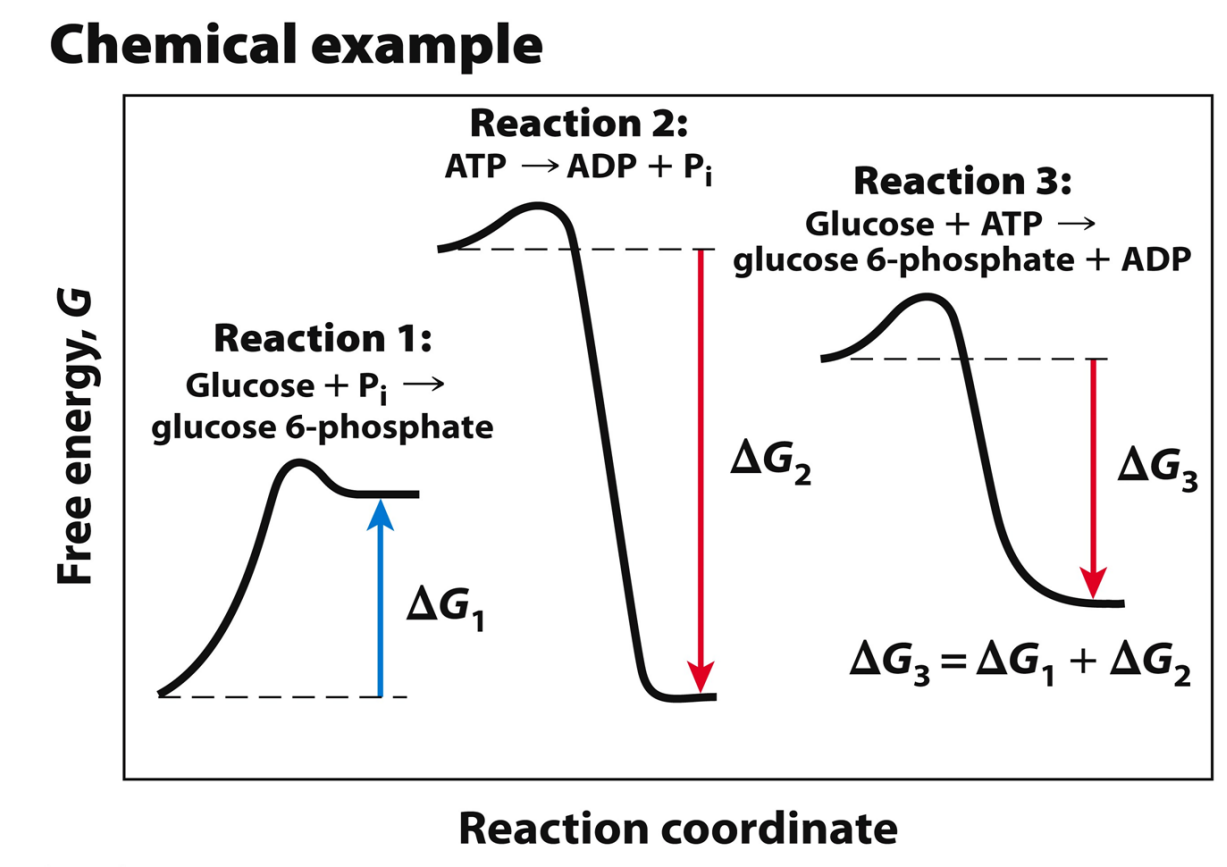
What are coupled reactions?
Many biological reactions are thermodynamically unfavourable (require energy or are endergonic)
To overcome this, endergonic reactions are coupled with exergonic (or energyreleasing) reactions—to do biological work
The reactions of metabolism harness energy-releasing reactions to drive energy-requiring reactions
What is Anabolism and Cataboilism
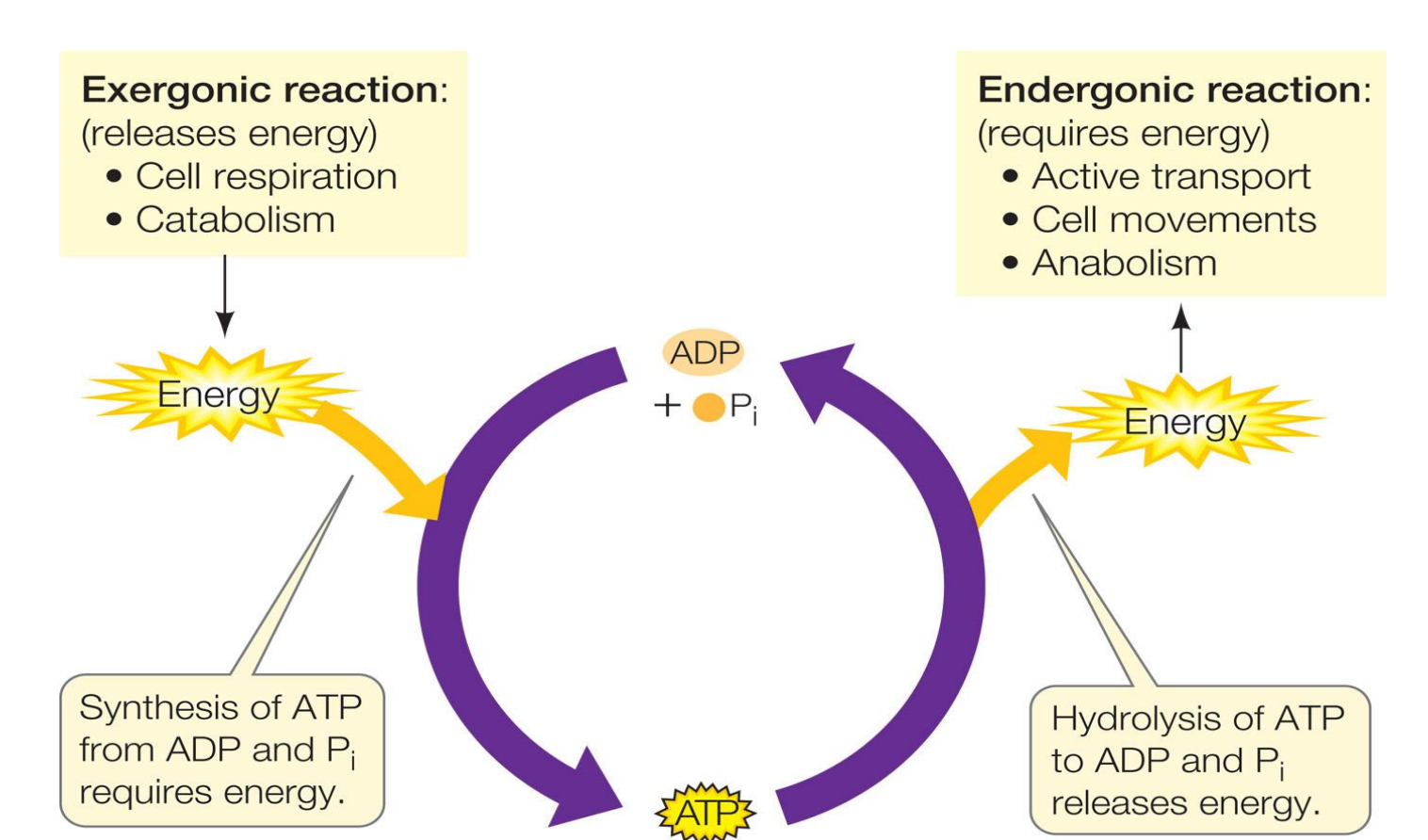
ANABOLISM
Uses precursor molecules to synthesise complex macromolecules
Requires an input of energy
Energy is captured in chemical bonds and stored as potential energy
CATABOLISM
Breakdown of complex molecules
Release of energy stored in chemical bonds
Occurs via oxidative pathways
Released energy is recaptured in new chemical bonds
Example of Chemical Coupling
Chemical coupling of exergonic and endergonic reactions allows otherwise unfavourable reactions to take place.
The “high-energy ” molecule (ATP) reacts directly with the metabolite that needs “activation.”
The final reaction will be: Glucose + ATP → GLU 6-P + ADP (deltaG = 13.8 + (-30.5) = -ve
The energy requiring reaction harnesses the energy from the energy releasing reaction
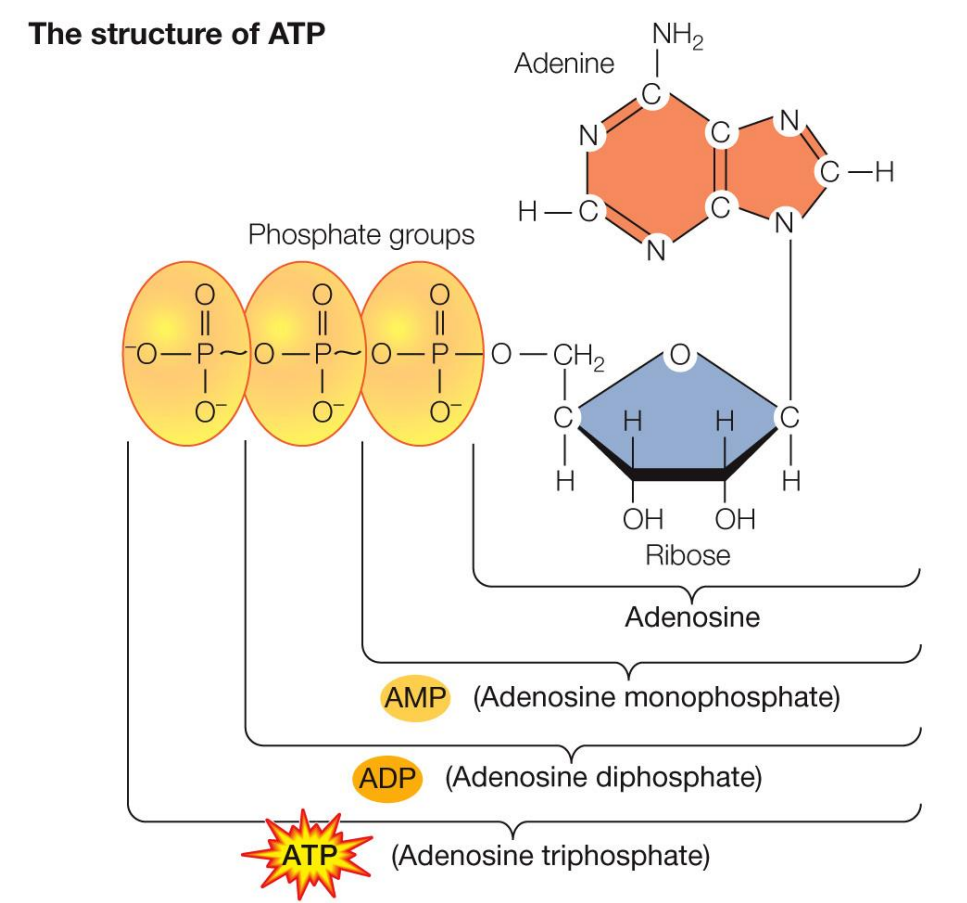
What is the structure of ATP (adenosine Triphosphate)
ATP is considered the energy currency, because the hydrolysis of the phosphate groups release energy
How do electrons work again
Capturing ELECTRONS is the key to harnessing energy
A substance that loses electrons is OXIDISED
A substance that accepts electrons is REDUCED
AN OIL RIG CAT
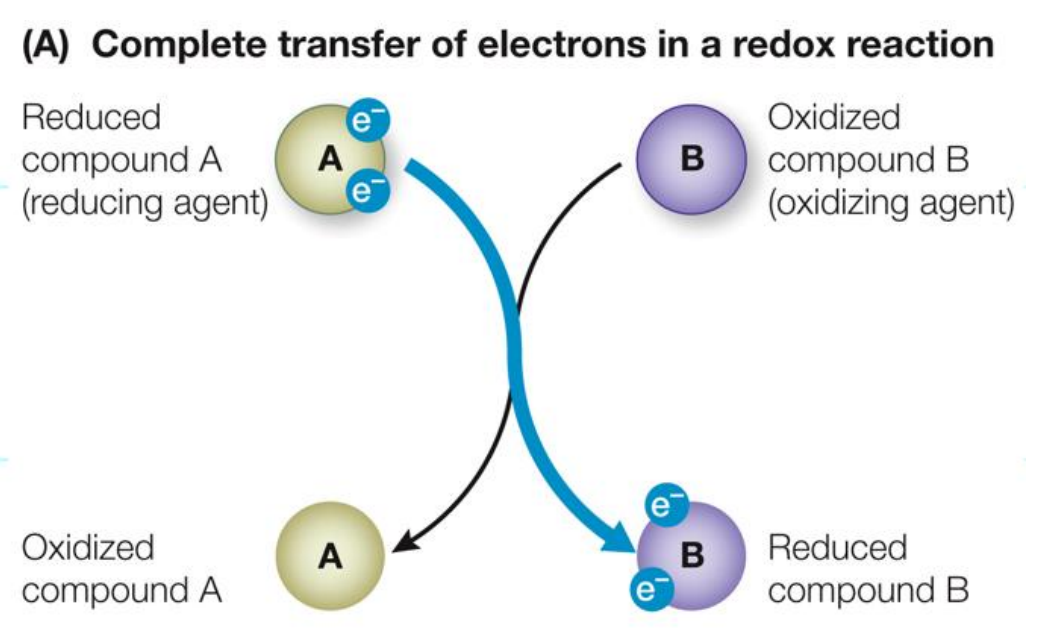
Biological Oxidation-Reduction Reactions
Electrons are transferred from one molecule (electron donor) to another (electron acceptor) in one of 4 ways in biological systems
Directly as electrons
Fe2+ + Cu2+ → Fe3+ + Cu+
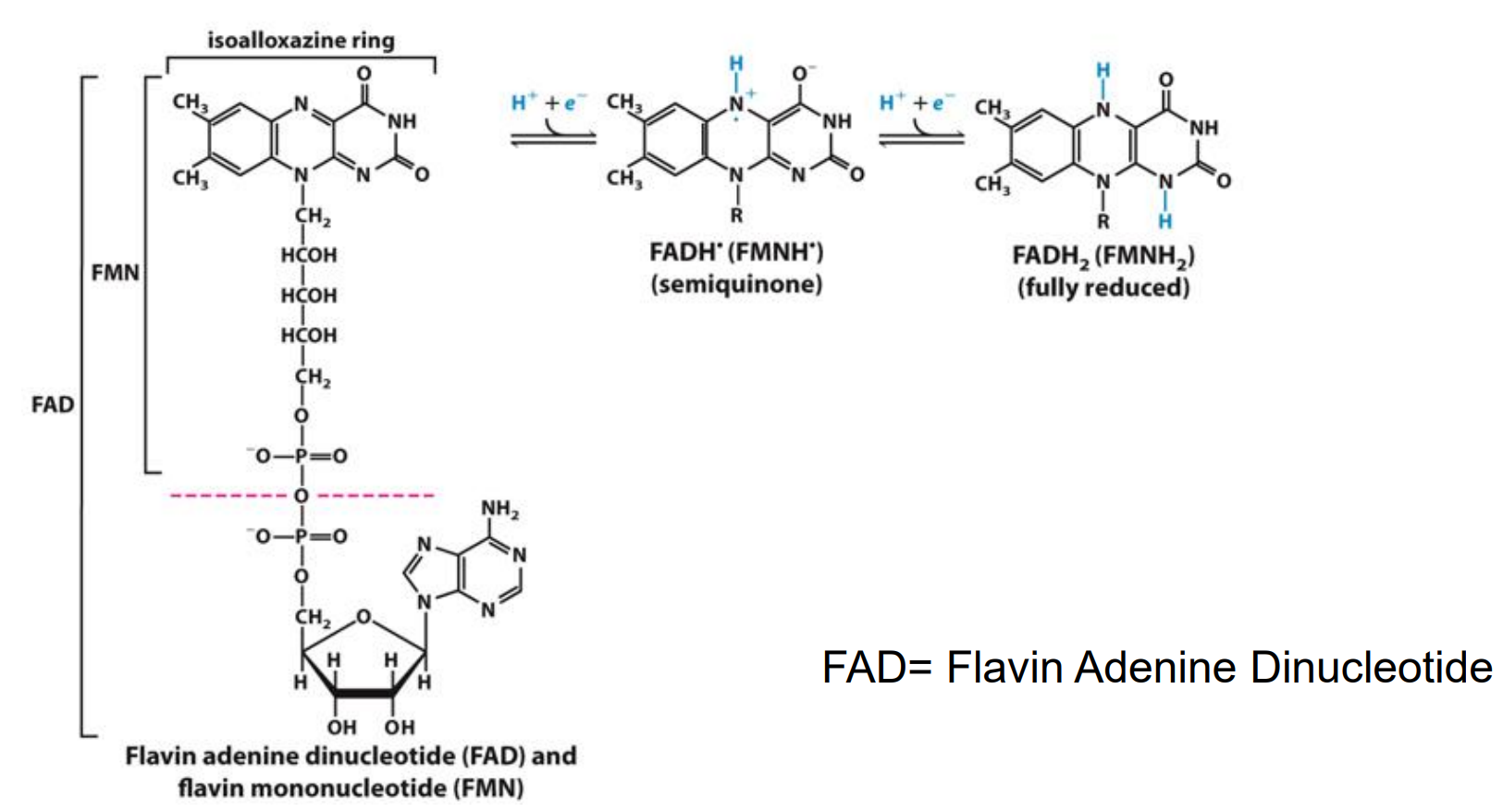
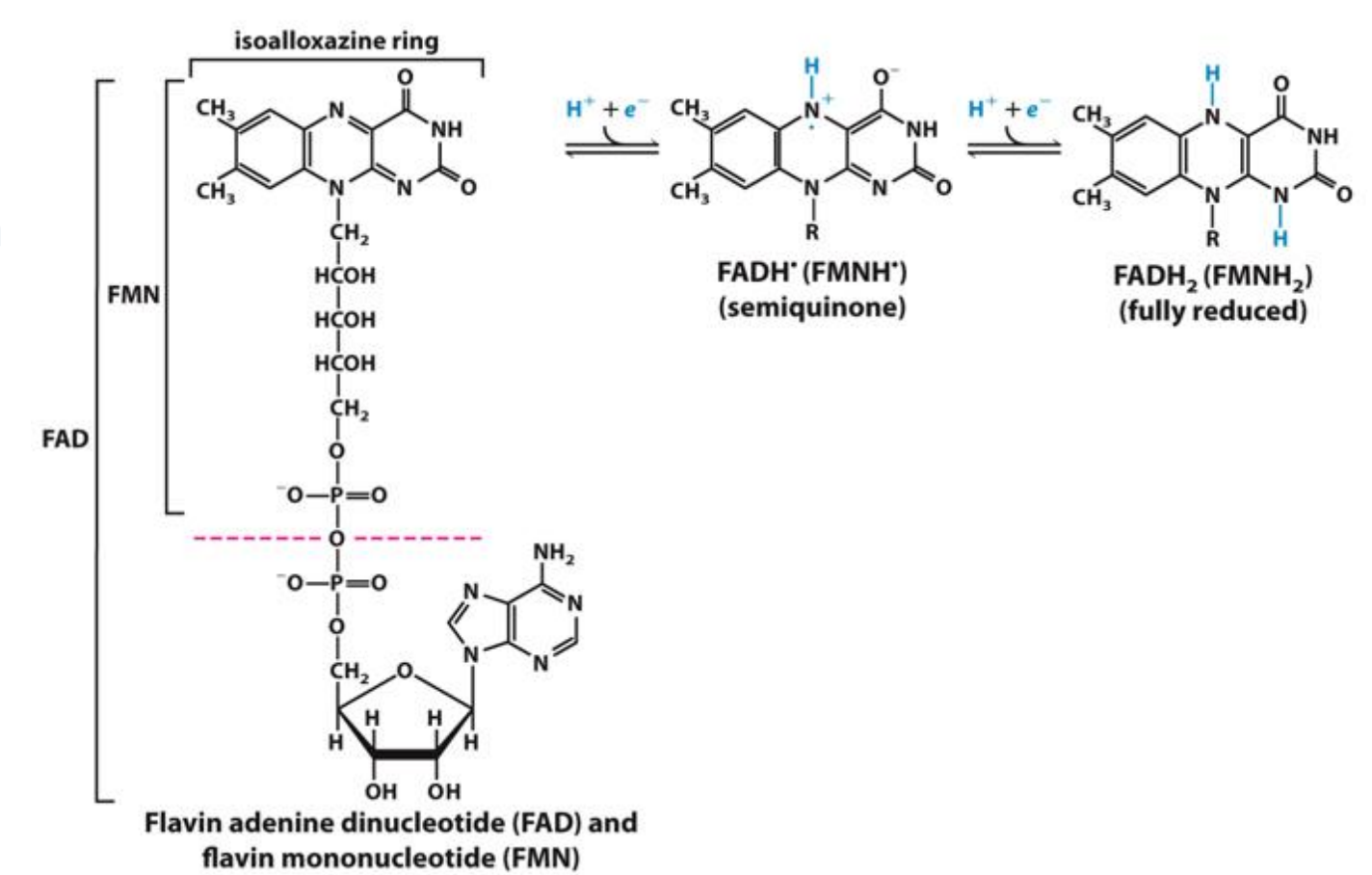
As hydrogen atoms (H+ + e-) (FAD)
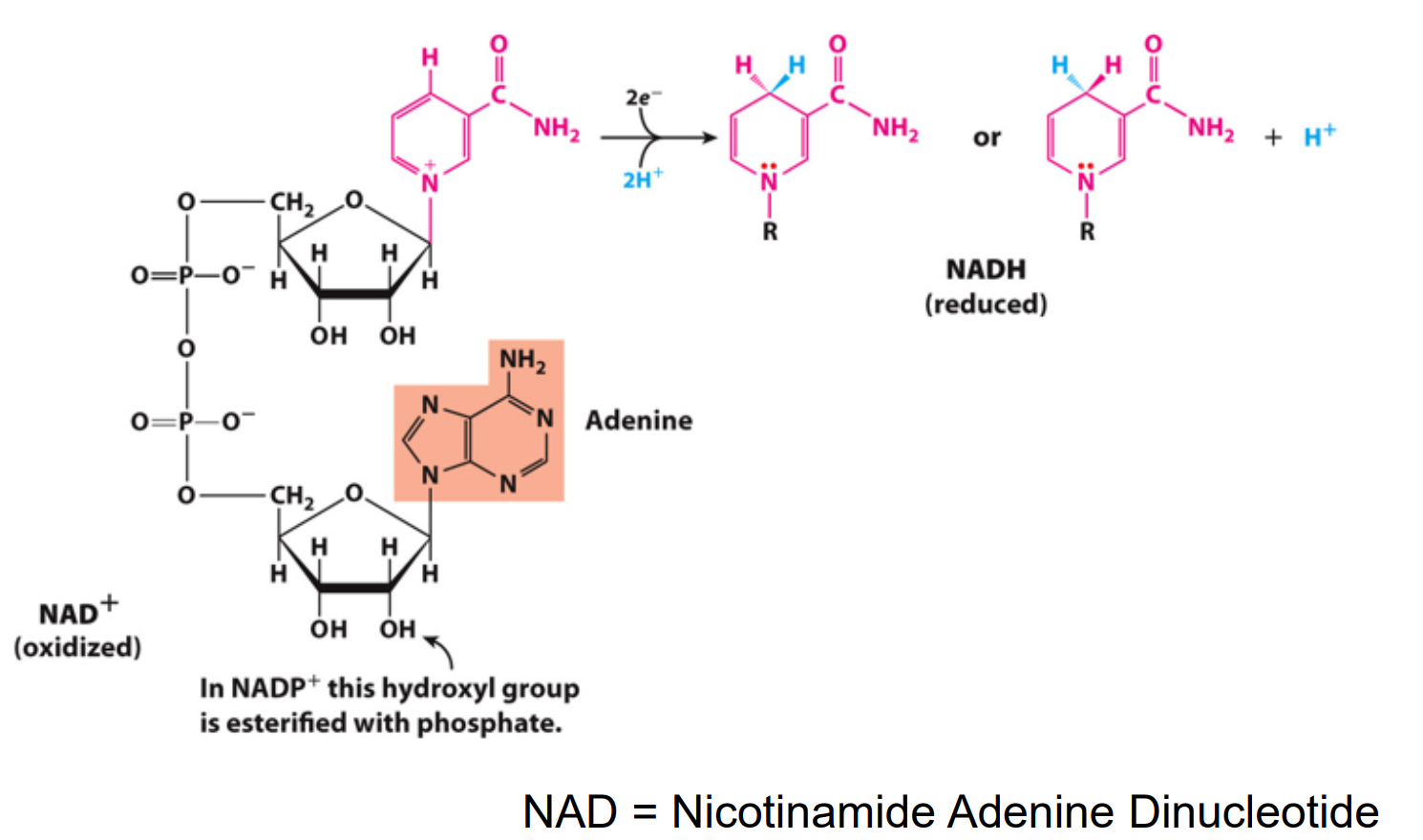
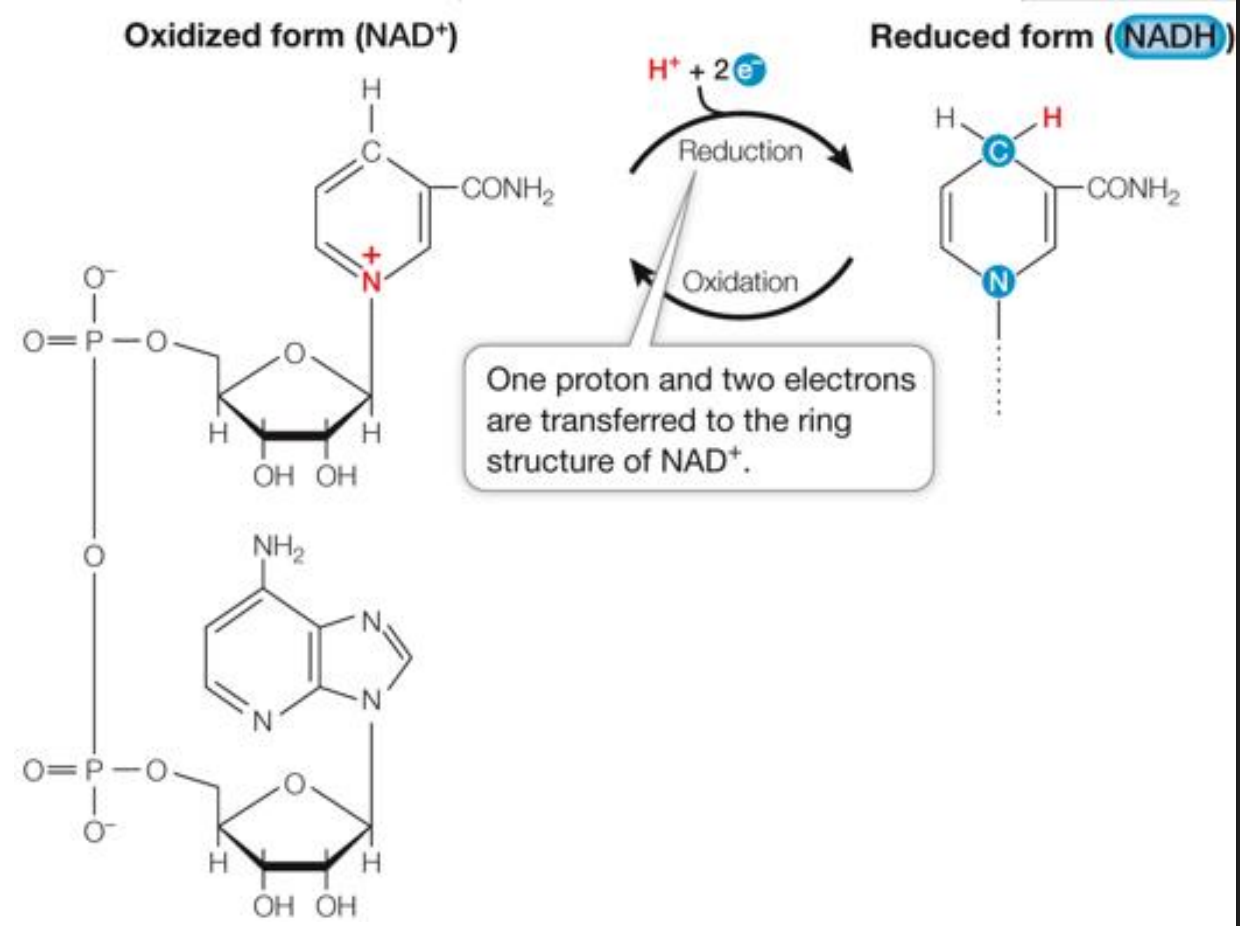
Transfer as a hydride ion (which is made of a 2e- and 1H+) (NAD)
Direct combination with oxygen
Oxygen combines with an organic reductant and is covalently incorporated to the product
R-CH3 + 1/2O2 → R-CH2-OH (oxidation of a hydrocarbon to an alcohol)
What are the most common electron carriers/acceptors?
NAD accepts e- and will be reduced to NADH
FAD will accept 2e- and will be reduced to FADH2
The molecules in orange are the electron carriers
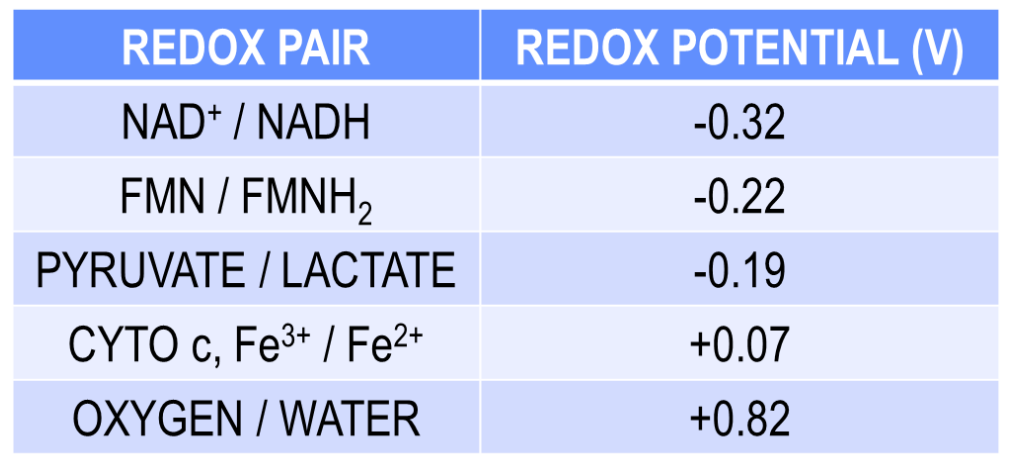
NAD+/NADH as Electron Carriers in Redox Reactions
O2 accepts electrons from NADH:
NADH + H+ + ½ O2 → NAD+ + H2O
The reaction is exergonic: ΔG = –52.4 kcal/mol
O2 is the oxidizing agent, reduced to water
Spontaneously occurring energy producing reaction
FAD/FADH2 as Electron Carriers in Redox Reactions
O2 accepts electrons from FADH2 and is reduced to water
The reaction is exergonic: ΔG = –40 kcal/mol
Spontaneously occurring energy producing reaction
What does the transfer of electrons to oxygen depend on?
The transfer of electrons to oxygen depends on:
SPECIFIC CARRIERS in the RESPIRATORY CHAIN or THE ELECTRON TRANSPORT CHAIN
REDOX POTENTIAL
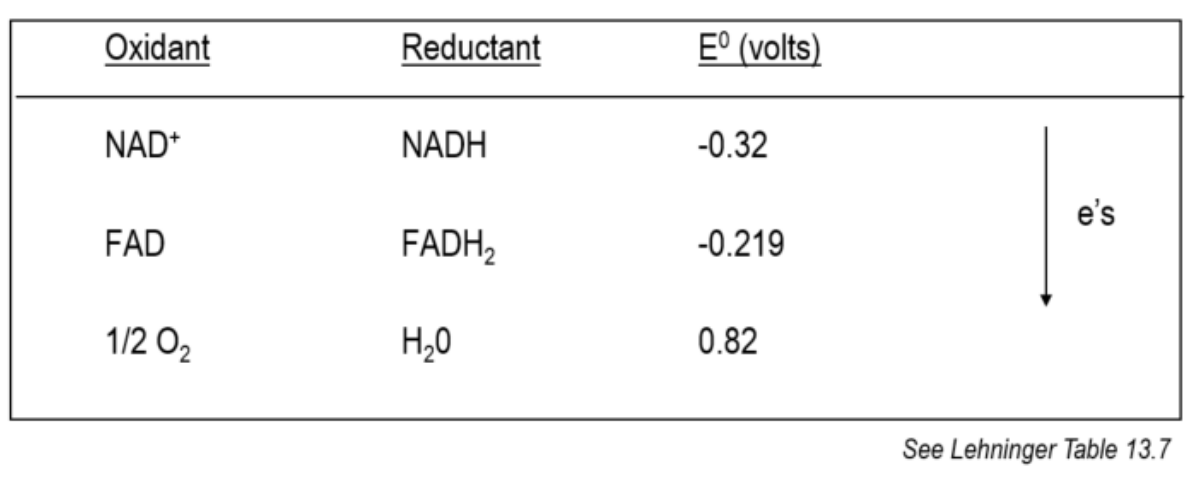
What is Redox Potential?
When two conjugate redox pairs are in solution;
The transfer of electrons from the donor to the acceptor depends on the relative affinity of the electron acceptor for the electrons (look in the image at the way the e- move)
Electrons always “fall” toward the species with the highest (most positive) reduction potential, because that species is the strongest oxidant — the best electron acceptor.
A reductant is the opposite: it is an electron donor. Electrons are not attracted to donors; donors push electrons away.
Free energy exchange in oxido-reduction reactions is proportionate to the ability of reactants to donate or accept electrons
ΔG can be expressed as an oxidation-reduction or redox potential E0
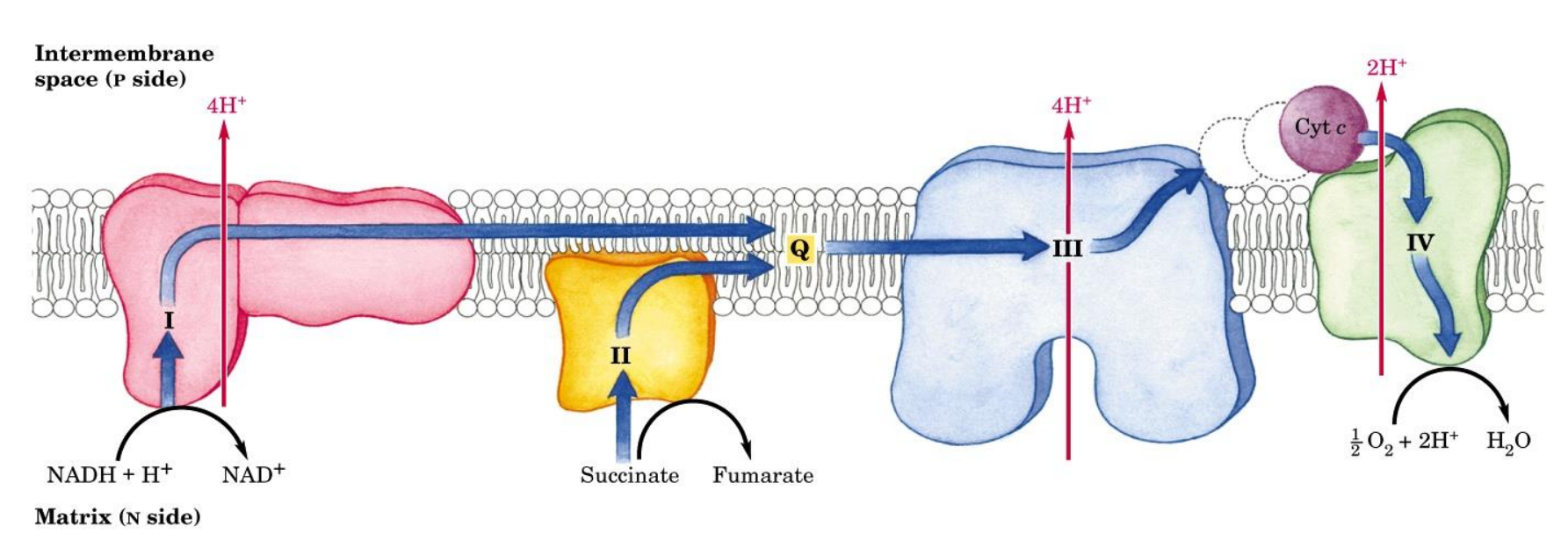
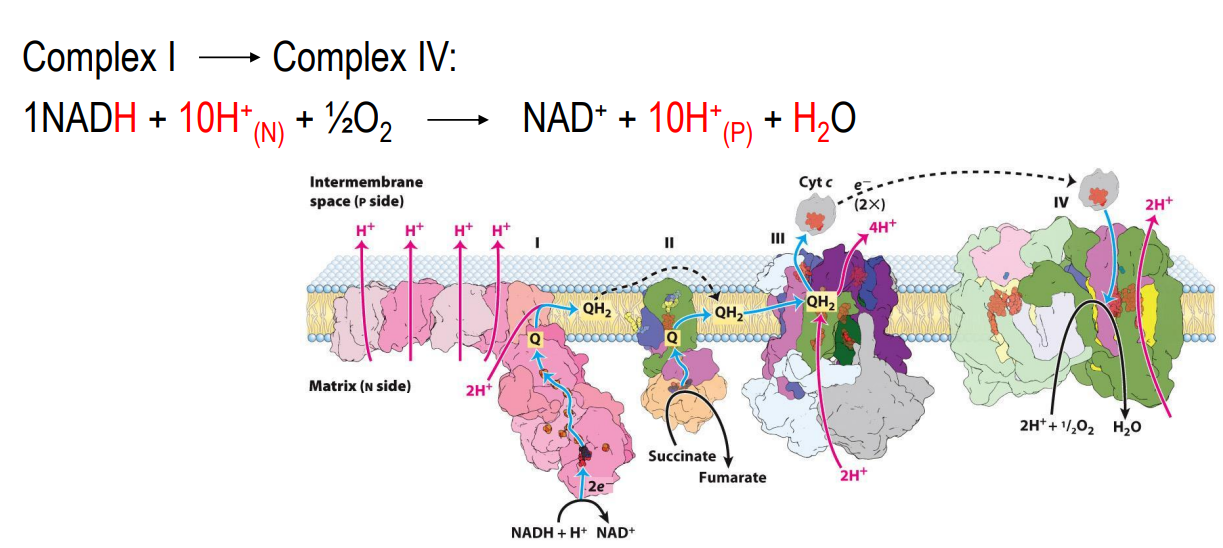
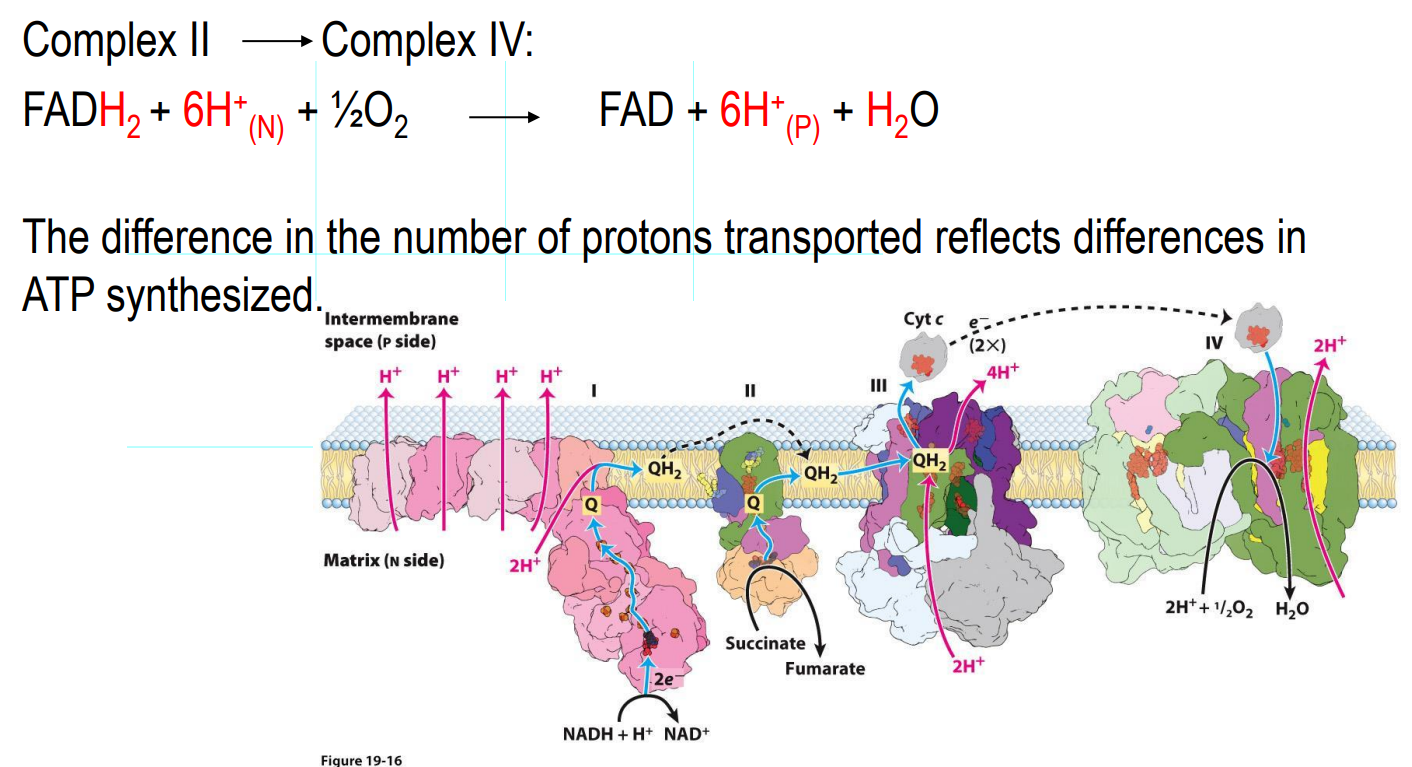
What are the main structures involved in electron transport
Complex I: NADH dehydrogenase
Complex II: Succinate dehydrogenase
Complex III: Ubiquinone:cytochrome c oxidoreductase
Complex IV: Cytochrome oxidase
Q: Coenzyme Q
Cyt c: Cytochrome C

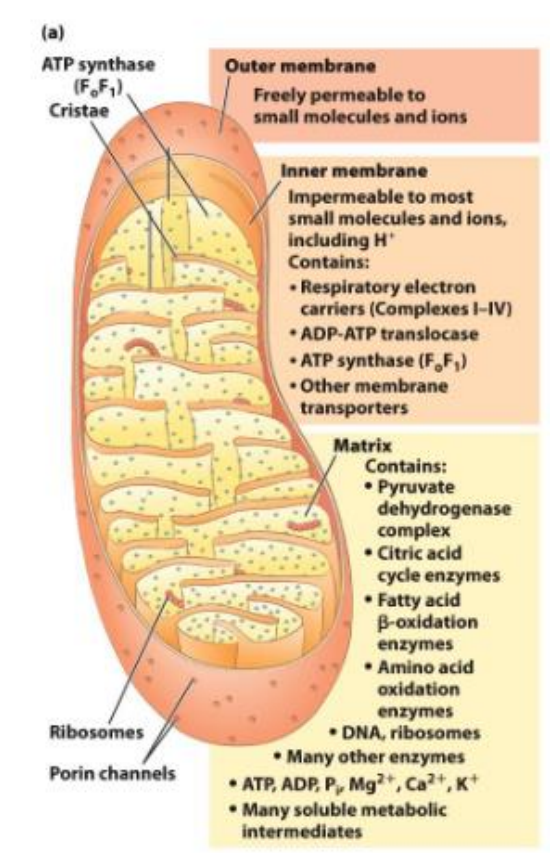
Where is the electron transport chain located?
The electron transport chain is located in the inner mitochondrial membrane
In the table above, we can see the transfer of electrons from the electronegative to the electropositive, indicated how the molecules at the top can readily donate e- and how oxygen at the bottom is a strong acceptor of electrons
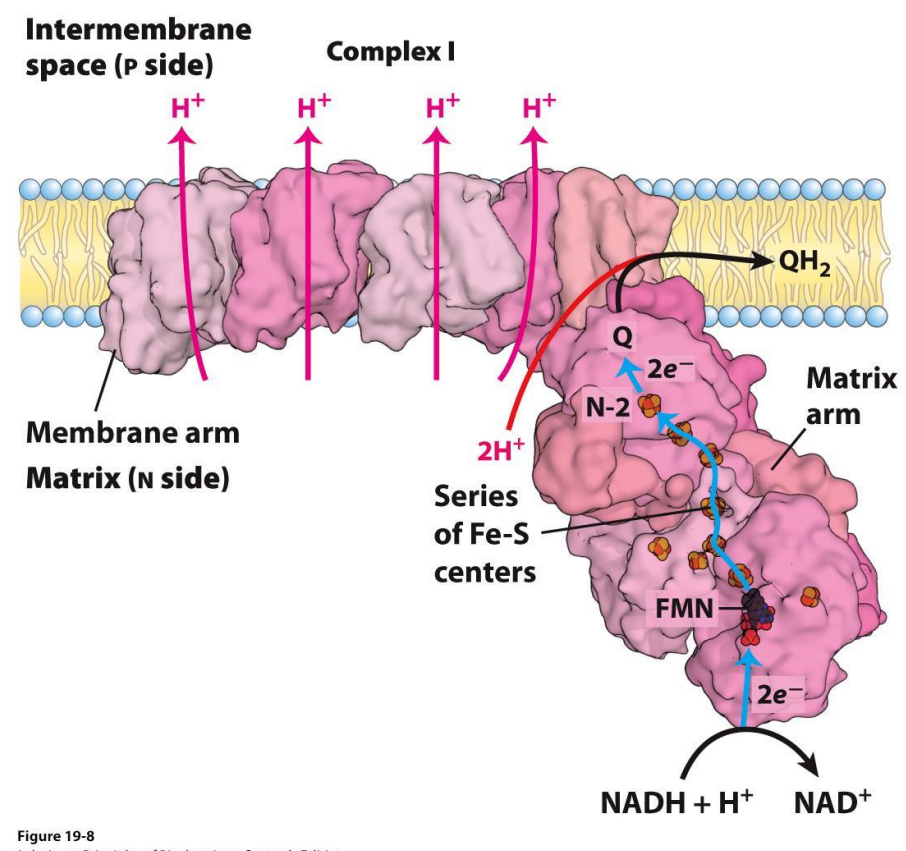
Electron Transport Chain — Complex I
NADH dehydrogenase complex
Multi subunit complex (around 45)
Accepts electrons from NADH
Electrons are transferred to coenzyme Q (quinone), which accepts these e- and will be reduced to QH2 (Ubiquinone)
QH2 is a mobile carrier (moves freely in the inner membrane to Complex III)
As electrons move from one carrier to the next, there is a release of free energy, and therefore that free energy means there is sufficient energy in the cell to expel 4 protons from the matrix to the intermembrane space
Every 2e- from NADH → EXPELS 4 PROTONS
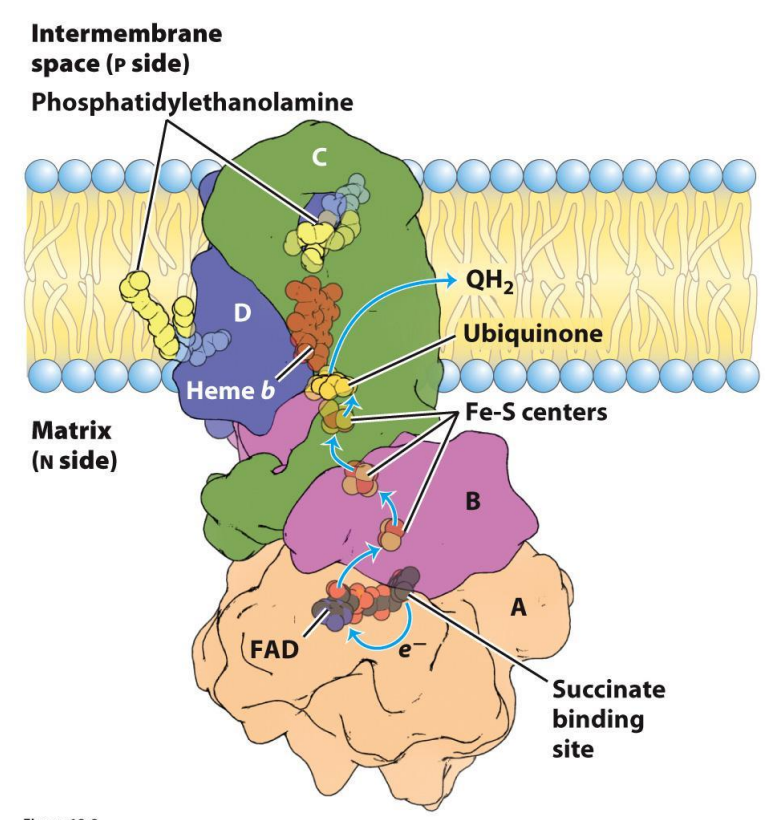
Electron Transport Chain — Complex II
Succinate dehydrogenase
Accepts electrons from FADH₂ (from the Krebs cycle)
Passes electrons to Coenzyme Q (quinone), which accepts these e- and will be reduced to QH2 (Ubiquinone) which will carry electrons to Complex III
Single enzyme with dual roles:
Citric acid cycle (later)
Capture and donate electrons in the ETC
DOES NOT EXPEL PROTONS
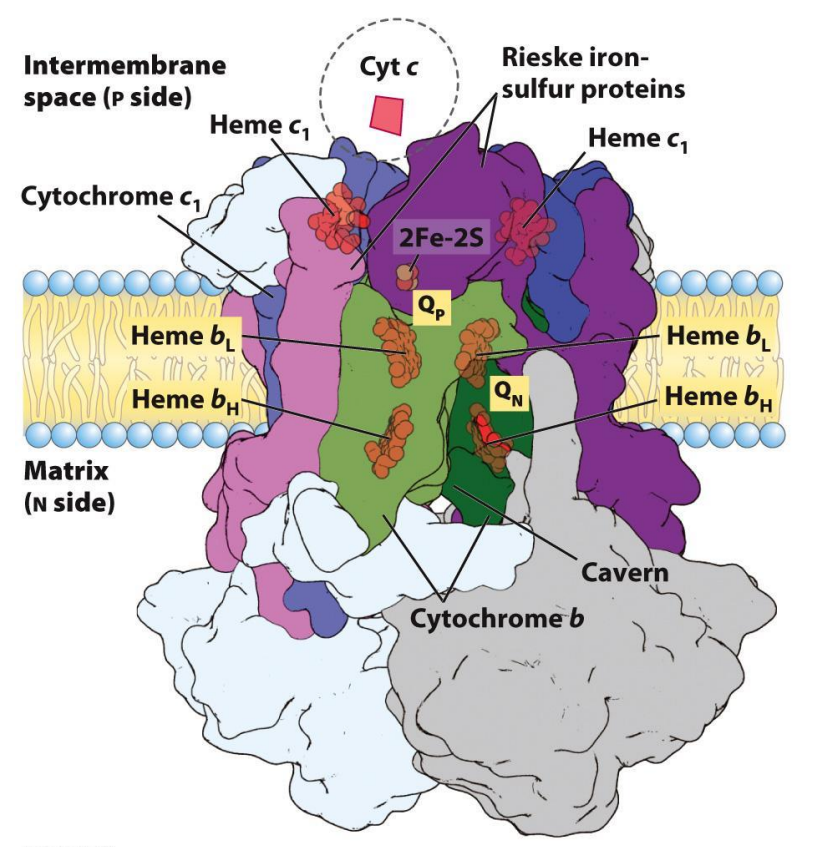
Electron Transport Chain — Complex III
Cytochrome C oxidoreductase
Receives electrons from QH2
Electrons are transferred to Cyt c (mobile carrier in the intermembrane space)
There is a transfer of electron, which means there is free energy released which is sufficient enough to EXPELS 4 PROTONS
Electron Transport Chain — Complex IV
Cytochrome oxidase
Accepts electrons from cytochrome c
Electrons are transferred to oxygen
O2 + 4H+ → 2H2O (oxygen is reduced thus is the oxidizing agent at the end)
The final electron acceptor station
Release of free energy → EXPELS 2 PROTONS
ETC Summery
The complexes are found embedded in the inner mitochondrial membrane
Take a look at BOTH equations
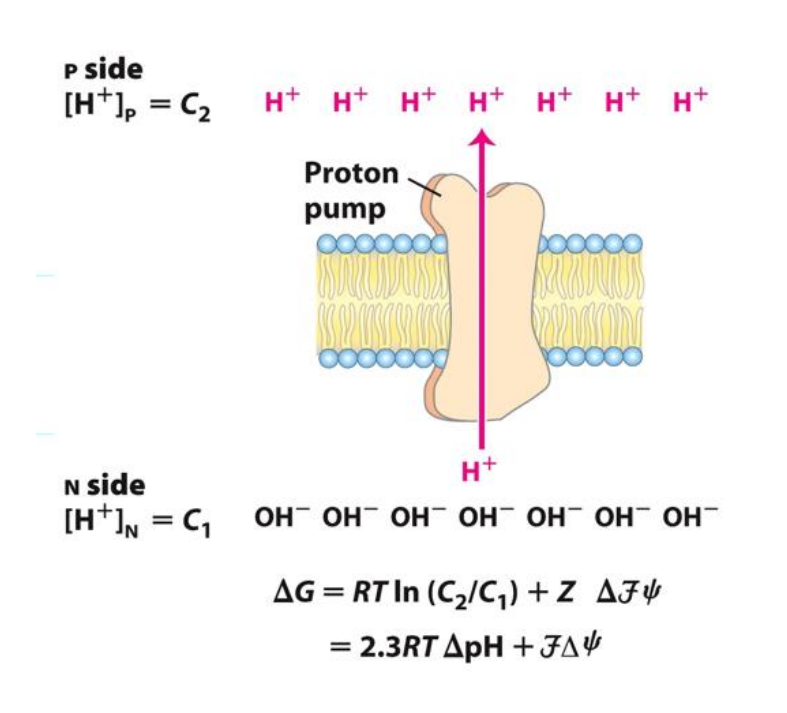
What does it mean for a proton gradient to be established?
The resulting proton gradient causes… (potential energy)
Chemical potential energy produced by the pH gradient
Matrix pH 8
Outside pH 7
Also generates an Electrical potential energy (Voltage gradient)
The matrix is electronegative compared with the outside space
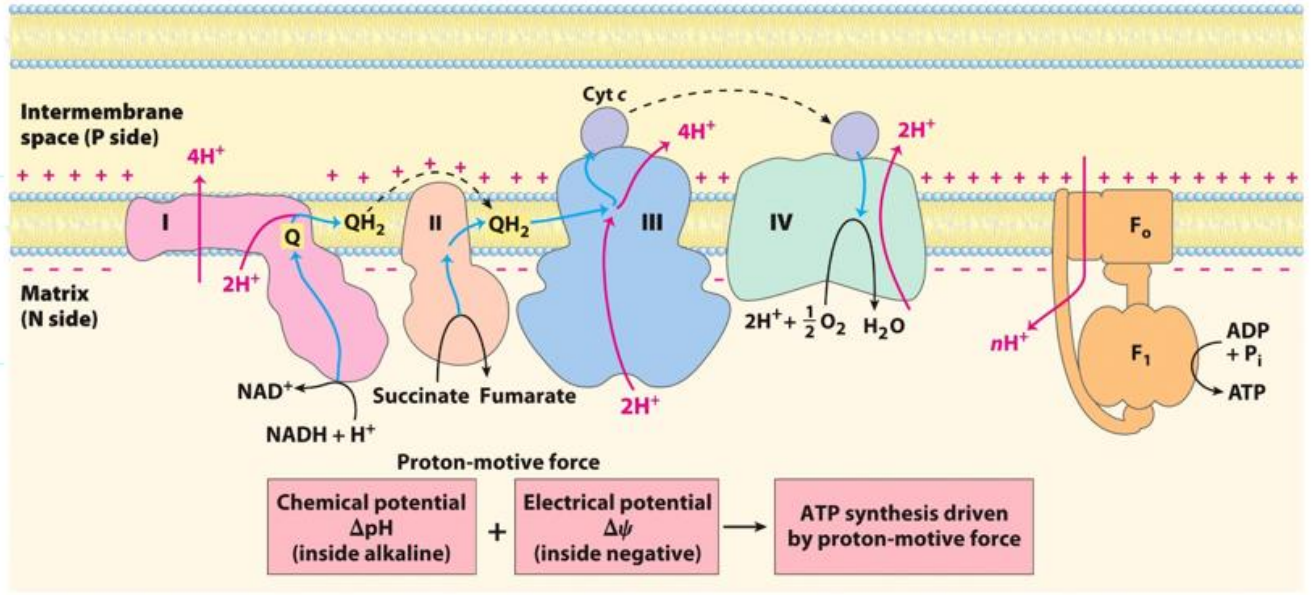
What is the chemiosmotic model for ATP synthesis
The energy released from the transfer of electrons (from NADH and FADH2 formed as a result of biological oxidation reactions) is stored as a transmembrane difference in proton concentration. This drives ATP synthesis.
Chemical potential + electrical potential → ATP synthesis driven by proton motive force
Complex V - ATP synthase complex
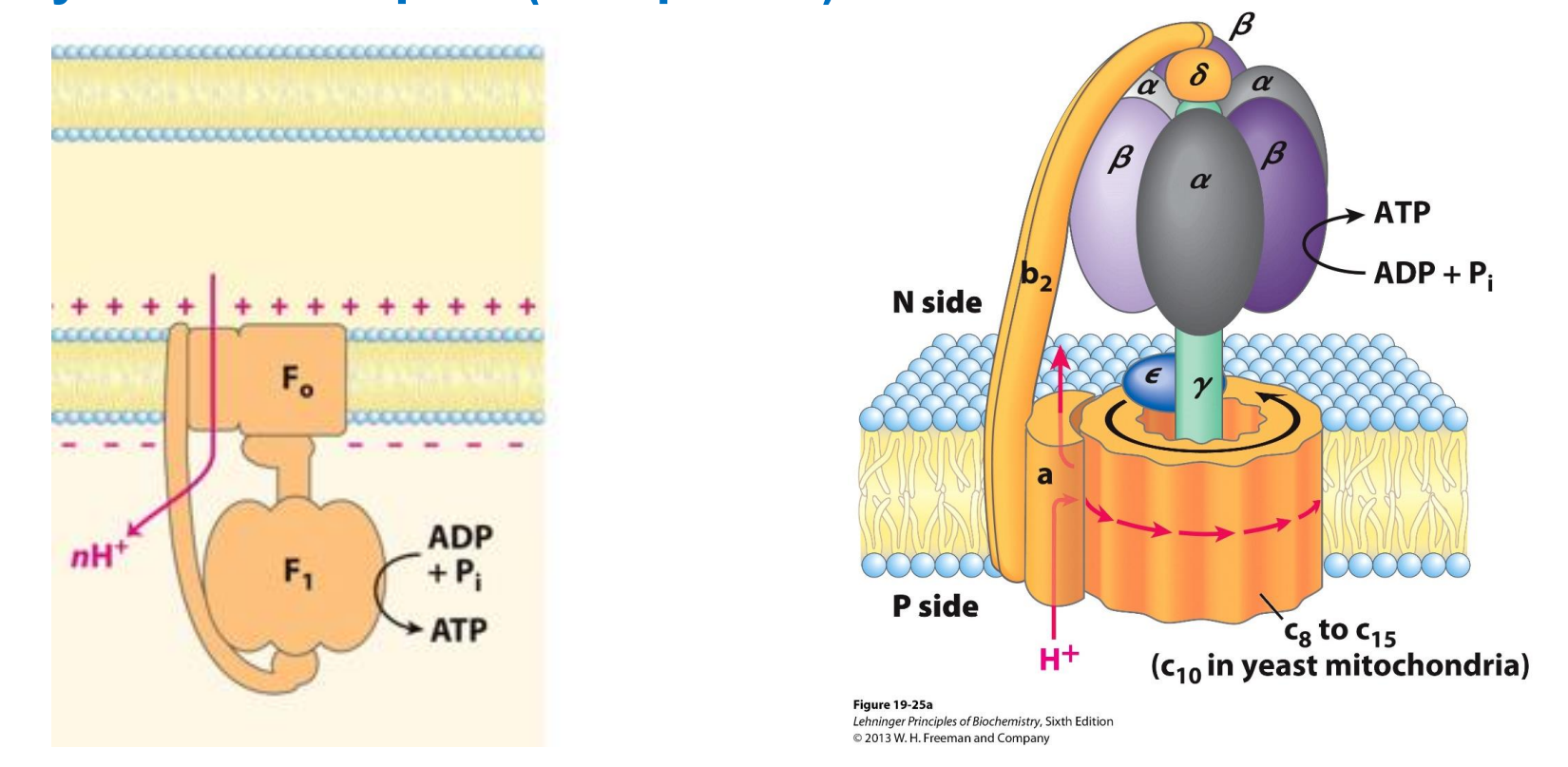
Complex V - ATP synthase complex
Experimental values show;
4 protons = synthesise ONE ATP
Made up of 2 subunits Fo (oligomycin - an antibiotic) and F1 (has catalytic activity, responsible for the phosphorylation of ADP to ATP)
Acts as a channel to allow the reentry of protons back into the matrix
The rotation of Fo subunit as H+ load in causes a confirmational change to F1 → this conformation change activates the enzyme ATP synthase → once activated will phosphorylate ADP to form ATP
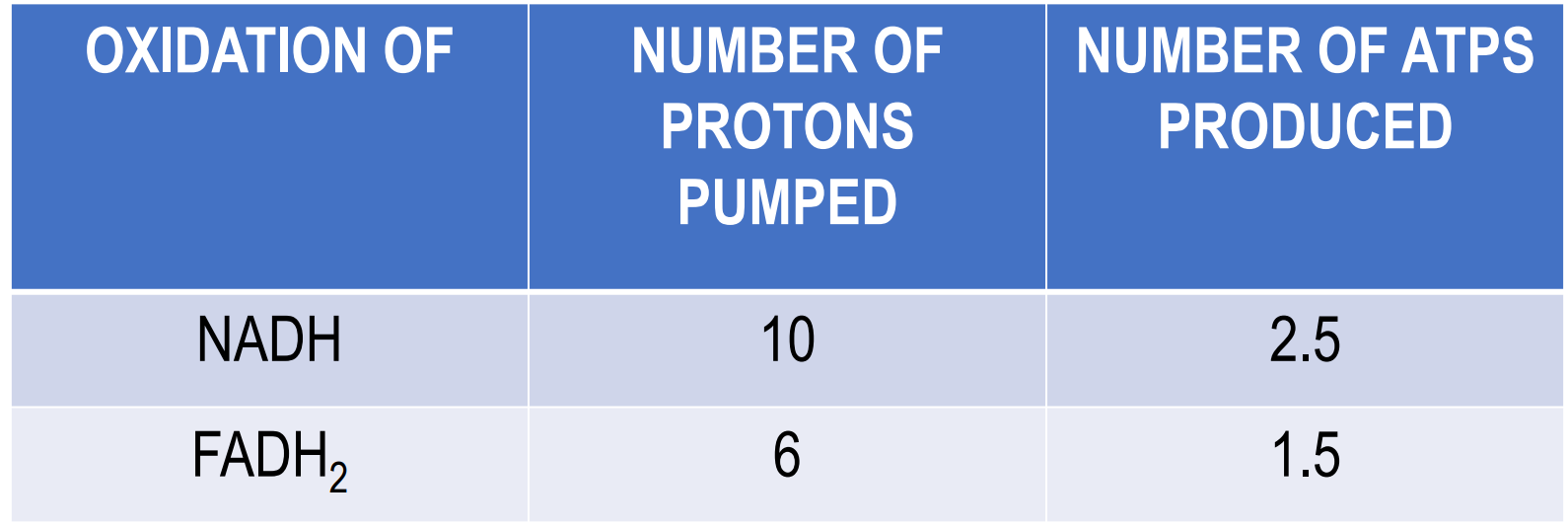
How many ATPs are synthesised when NADH and FADH2 are oxidised?
4 protons = synthesise ONE ATP
We know that the transfer of electrons from NADH to oxygen ∆G = -190 kJ/mol, which is a HUGE energy release
ATP synthesis is endergonic (requires energy): ADP + Pi → ATP ∆G = +52 kJ/mol
But luckily it offsets the transfer of electrons from NADH to oxygen
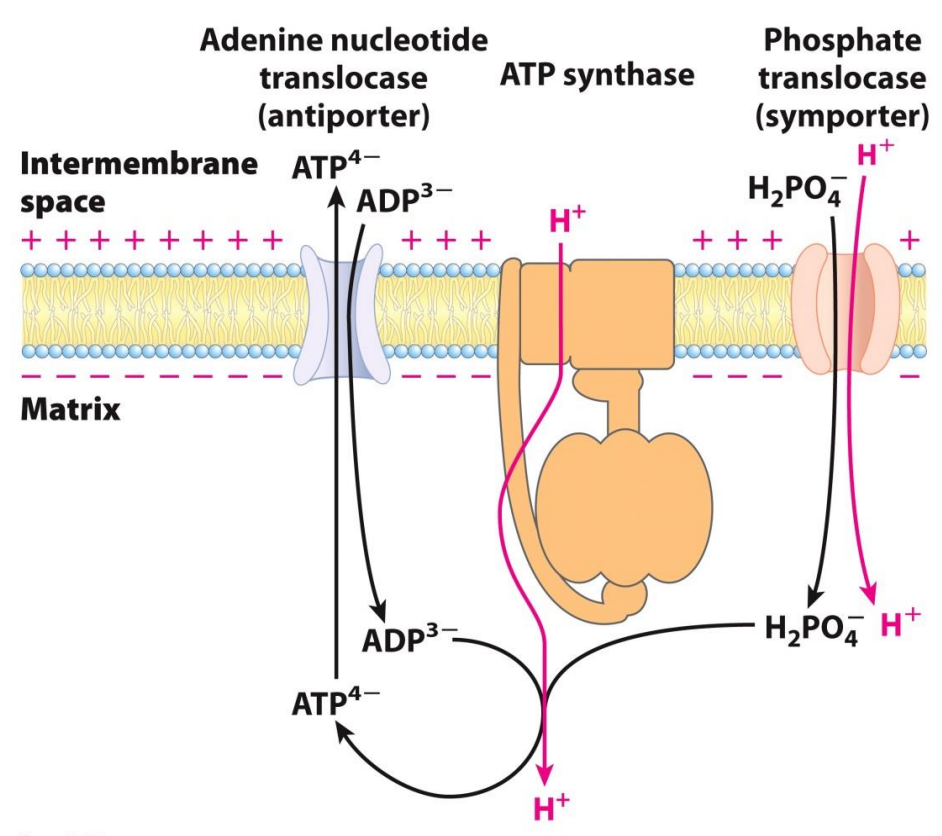
What happens to the ATP that is synthesized?
Proton Motive Force drives ATP synthesis as well as transports substrates into the matrix and products out of the matrix
Inner membrane space is very impermeable, then how does it get all the substrates to synthesis ATP and what happens to the ATP
ADP comes from outside and is transported by the Adenine nucleotide translocase (antiporter) which is a transport protein
The phosphate (in the form of dihydrogen phosphate) comes from outside and is transported by a phosphate translocase (symporter) which is a transport protein
Now substrates are in the matrix for the phosphorylation to happen
The ATP synthesized is then transported out via the Adenine nucleotide translocase (antiporter)
The Adenine nucleotide translocase brings a charge of -3 inside (ADP) and then releases a -4 charge (ATP) outside, since the outside is more positive the transport of ATP (the more negative one) is favored
What is Oxidative Phosphorylation?
The process by which the oxidation of NADH or FADH2 (transfer of electrons to oxygen) results in the synthesis of ATP
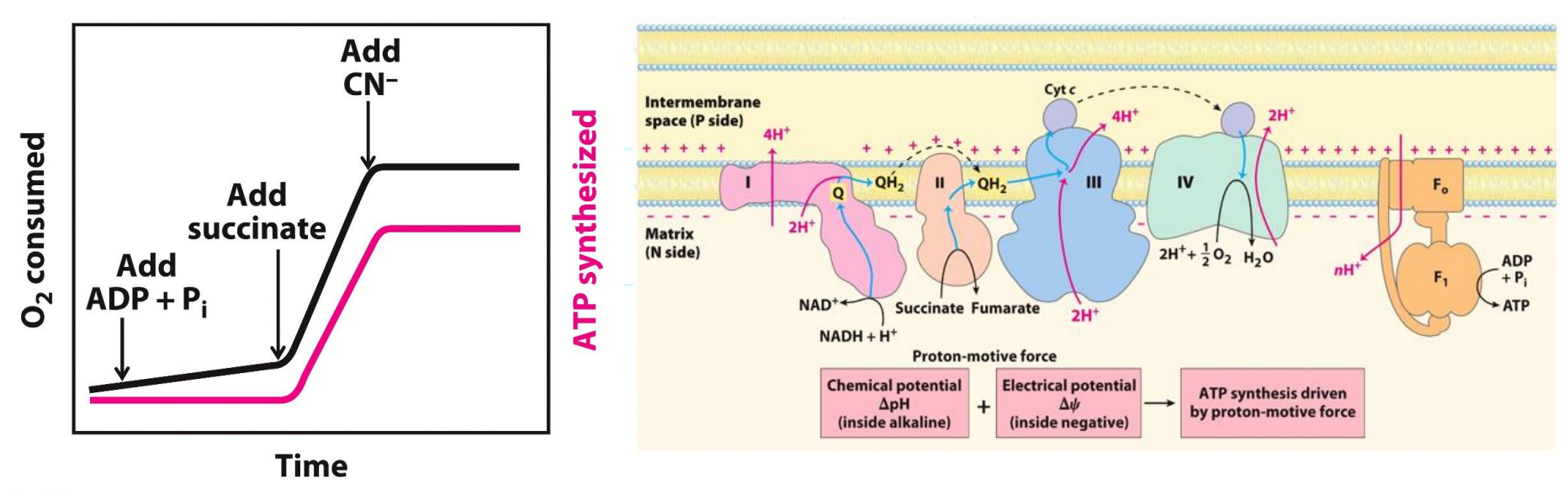
The electron transport chain and ATP synthesis are COUPLED
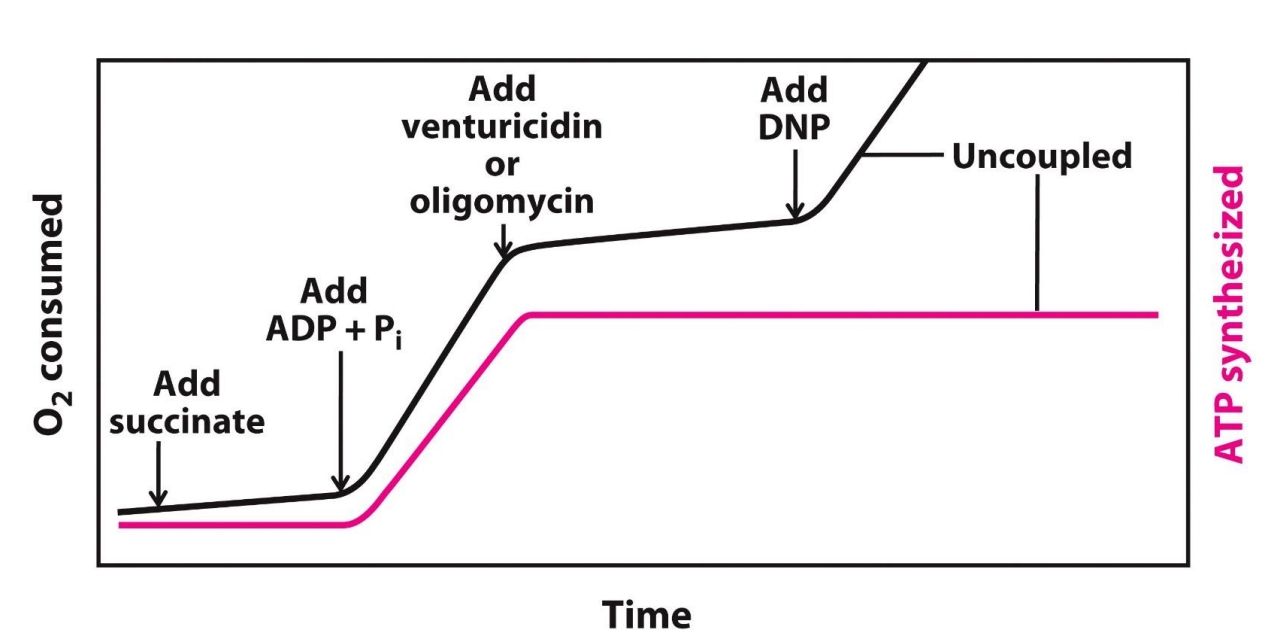
O2 consumed → reduction of O2 to H2O (GRAPH UP)
At the start both graphs are flat since not much O2 is being consumed and not much ATP is being synthesized
Then they added succinate → this oxidizes to fumarate, during this reaction FAD is reduced to FADH2, FADH2 then passes its electrons to the reaction
Now the O2 consumed (reduced) and ATP synthesis is steadily increasing - means electrons are being transported now
Add CN- (cyanide)
This inhibits complex IV, this means the electrons are not transported to oxygen, ATP synthesis is also come to a halt (plateau)
THIS SHOWS US that if the ETC is inhibited, ATP synthase is also inhibited and thus these processes are COUPLED
Succinate added, adds electrons, but the graphs shows that O2 is not reduced as much and ATP is not synthesized as much (naturally as there is no substrate)
Thus now add the substrates ADP and Pi, now with BOTH the substrates and electrons, more O2 is consumed and as a result more ATP is synthesized
Now add Venturicidin or oligomycin to inhibit ATP synthesis → now we see ATP synthesis comes to a halt as a result the ETC also comes to a halt
THIS SHOWS US that if the ATP synthase is inhibited, the ETC is also inhibited and thus these processes are COUPLED, INTACT mitochondria
Now uncouple the process by adding DNP (an uncoupler), then we see ATP synthesis continue to be inhibited, but ETC can continue to take place (not having to occur at the same time)
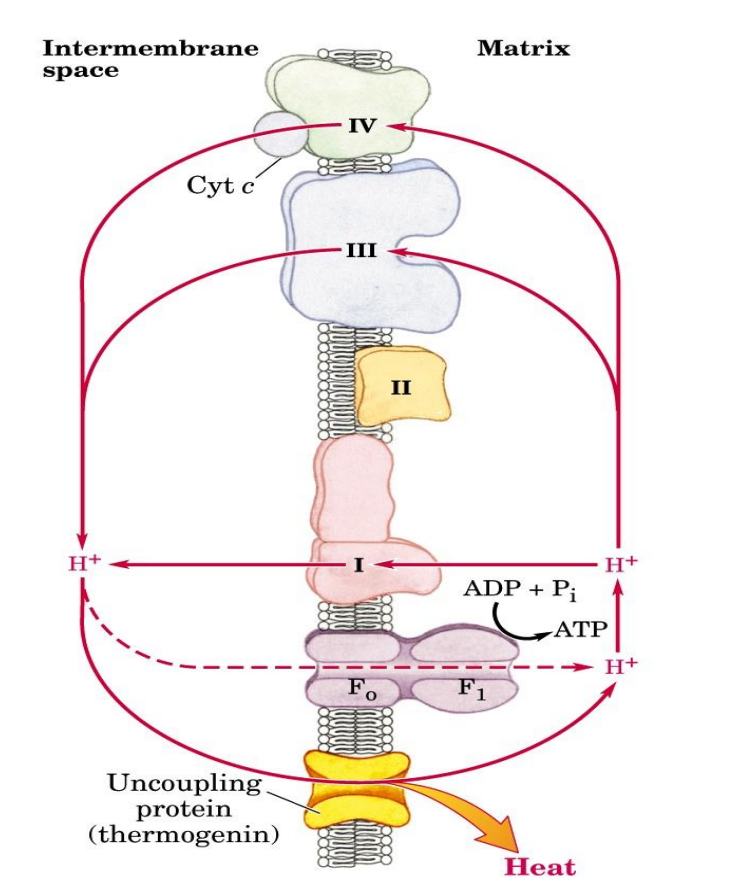
The electron transport chain and ATP synthesis can be UNCOUPLED, HOW?
Due to…
Mechanical damage
Leakages → hard to establish a PMF
Chemical reagents (e.g. 2,4-dinitrophenol (DNP))
DNP is a hydrophilic proton carrier (uncoupler), therefore it can diffuse into the matrix, carrying those protons which will then dissipate the proton gradient and PMF that had been established, now ATP synthesis does not occur, only the ETC can take place thus why it is an uncoupler
Natural uncouplers
Commonly found in newborns
Uncoupling protein 1
Thermogenin found in brown adipose tissue - which has a brown color due to an excess of mitochondria which therefore has a high conc of cytochrome C
Again provides a channel for protons to enter the matrix, thus dissipates the protons gradient and PMF, therefore ATP is not produced BUT the energy produced is released as heat energy which is important to help new borns/animals to adjust to the temp of the environment
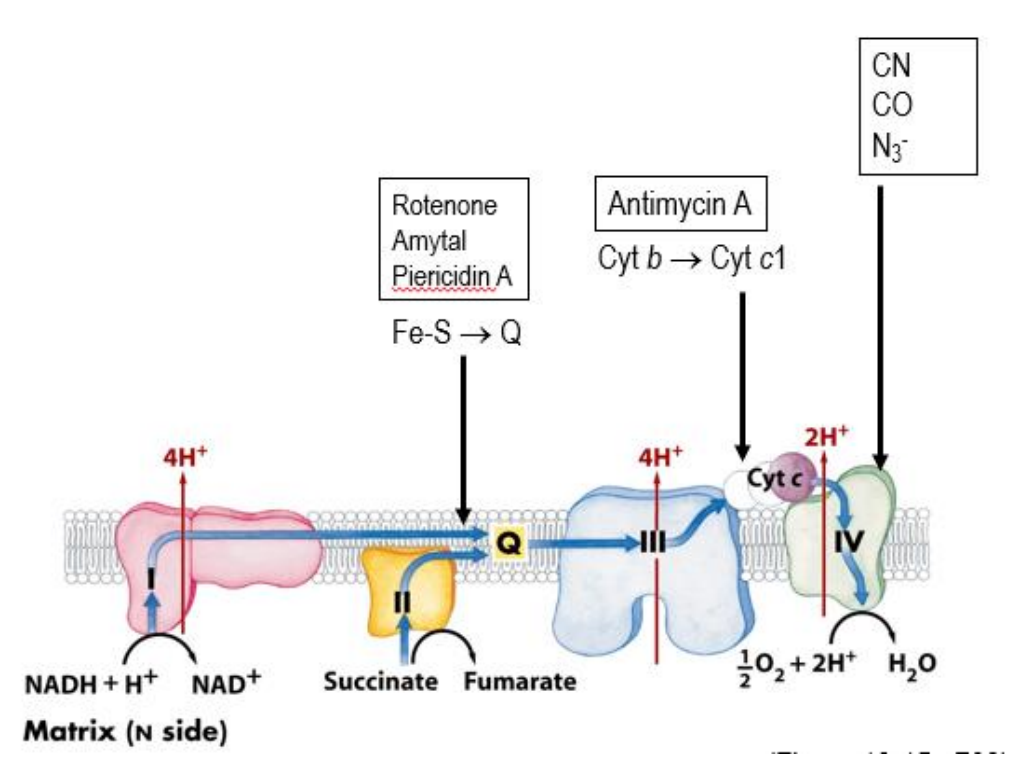
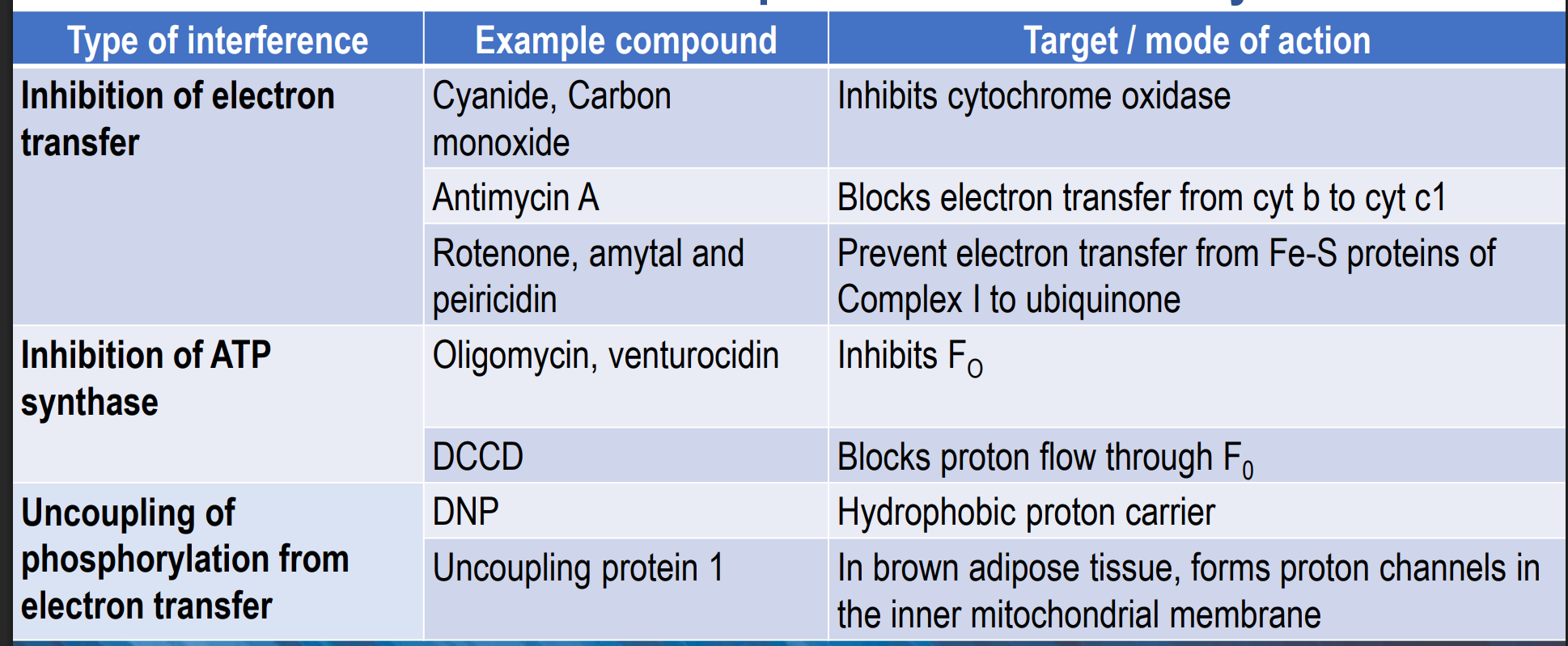
Whoa are the inhibitors of the electron transport chain and ATP synthesis?
Venturicidin and Oligomycin (inhibit the Fo complex of the ATP synthase)
Look at all the ones given in the image above (these inhibit the ETC)
THESE CEASE PRODUCTION - doesn’t decrease it
How is oxidative phosphorylation controlled?
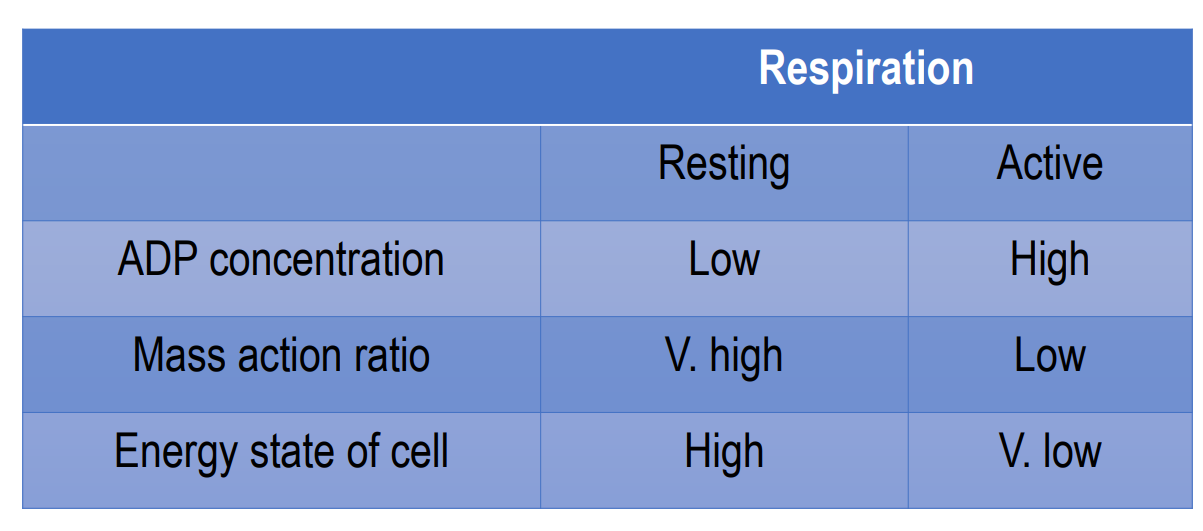
ATP synthesis is regulated by the energy needs of the cell
Depends on the mass action ratio [ATP] / [ADP][Pi] (product/substrate)
High energy energy, lots of ATP, low ADP → respiration is resting
ATP decrease, ADP increase → increase in respiration, coupled with oxidative phosphorylation, mass action ratio is low
ATP and ADP back to normal → respiration resting
![<ul><li><p>ATP synthesis is regulated by the energy <em>needs </em>of the cell</p></li><li><p>Depends on the mass action ratio [ATP] / [ADP][Pi] (product/substrate)</p></li><li><p>High energy energy, lots of ATP, low ADP → respiration is resting </p></li><li><p>ATP decrease, ADP increase → increase in respiration, coupled with oxidative phosphorylation, mass action ratio is low</p></li><li><p>ATP and ADP back to normal → respiration resting </p></li></ul><p></p>](https://assets.knowt.com/user-attachments/81a1d2c7-bcad-4e80-866a-2f991e62ccd5.png)
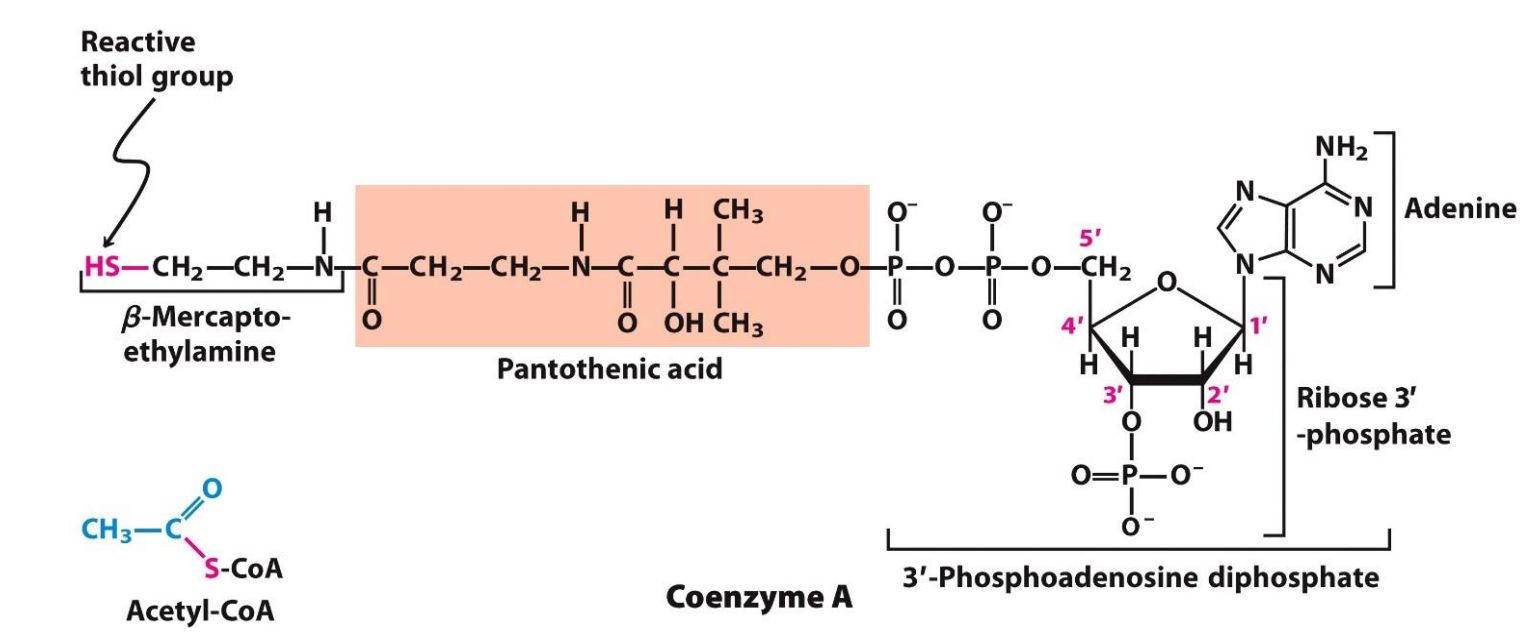
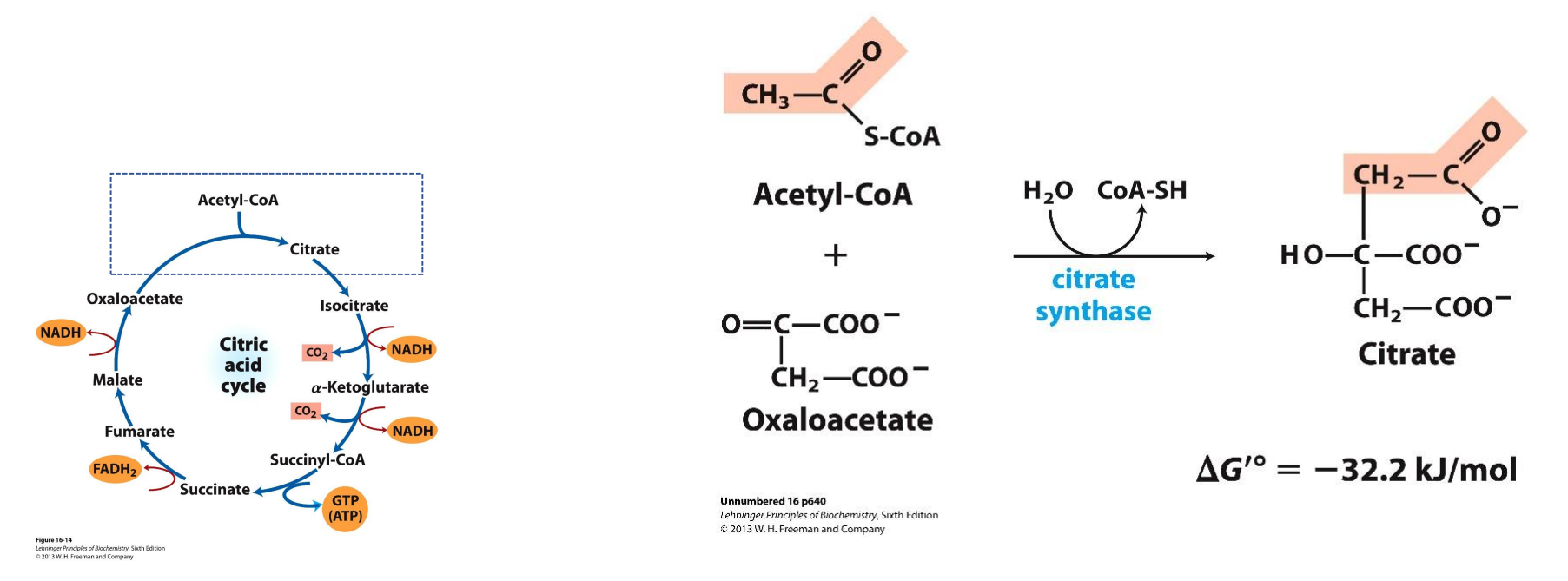
What is Acetyl CoA?
Make up of Coenzyme A + Acetate - has 2 Carbon atoms
Coenzyme A is derived from pantothenic acid (part of the vitamin B group) and has a reactive thiol group (-SH) which condensed with the acetate to form Acetyl CoA (by forming a thiol ester linkage, which is very important because it is the hydrolysis of this bond in Acetyl CoA that releases a HUGE amount of energy (-31.4 KJ/mol) which is sufficient to dive the ENTIRE CAC)
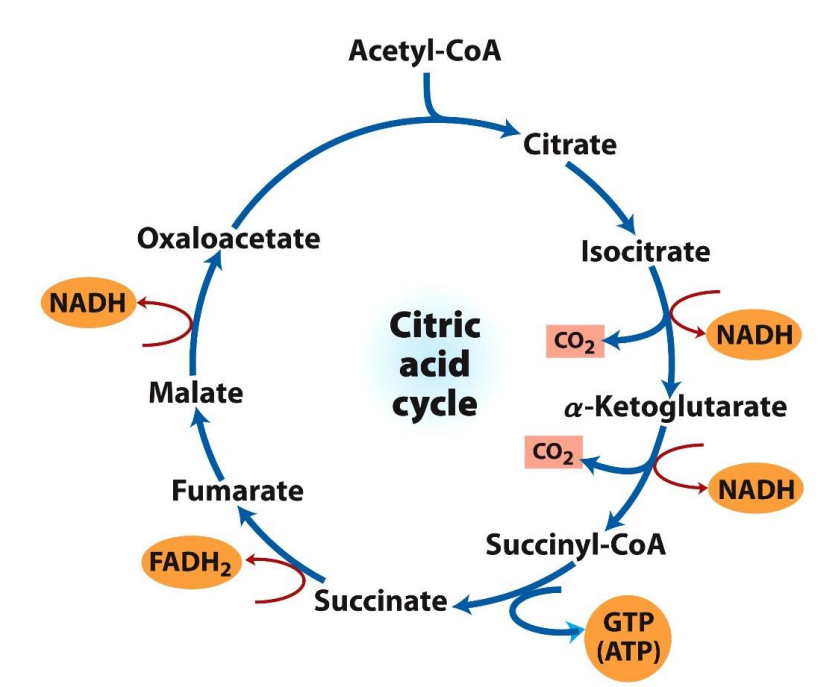
Where does the Citric Acid Cycle (CAC) take place?
In the mitochondrial Matrix
The 2 carbon atoms in Acetyl CoA are released as CO2 through multiple reduction/oxidation reactions, these electrons released are captured to form the NADH, FADH2 and GTP
The citric acid cycle is aerobic and therefore cannot occur in the absence of oxygen
Even though oxygen is not used directly in the cycle, it is required indirectly because NADH and FADH2 must be recycled, This regeneration happens only in the ETC which can only run if oxygen is present as the final electron acceptor
CAC: Step 1 Condensation
2C Acetyl CoA + 4C Oxaloacetate → 6C Citrate
The energy to carry out the reaction is provided by the hydrolysis of the thiol ester linkage in Acetyl CoA and it provides a huge surplus of energy as seen with the DG value given