Chapter 14: Energy Conversion - Mitochondria and Chloroplasts
Energy Conversion: Mitochondria and Chloroplast
- Mitochondria: Occurs in all cells.
- burn food particles to produce ATP by oxidative phosphorylation.
- Chloroplast: it occurs only in plants and green algae.
- harness solar energy to produce ATP by photosynthesis.
Evolutionary origin
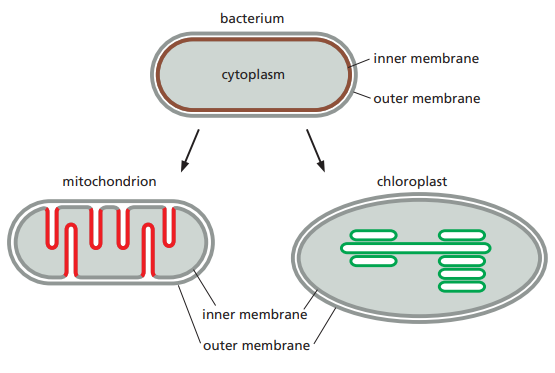
The common evolutionary origin of the energy-converting machinery in mitochondria, chloroplasts, and prokaryotes (archaea and bacteria) is reflected in the fundamental mechanism that they share for harnessing energy. This is known as chemiosmotic coupling.
Signifies a link between the chemical bond-forming reactions that generate ATP and membrane transport processes.
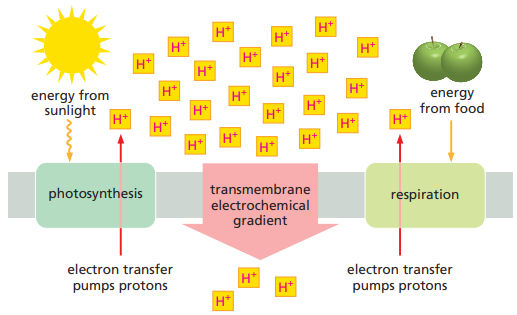
Chemiosmotic Process
Stage 1:
- High-energy electrons (derived from the oxidation of food molecules, from pigments excited by sunlight, or from other sources described later) are transferred along a series of electron-transport protein complexes that form an electron-transport chain embedded in a membrane.
- Each electron transfer releases a small amount of energy that is used to pump protons (H+) and thereby generate a large electrochemical gradient across the membrane.
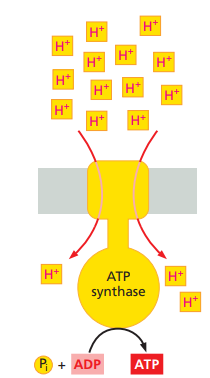
Stage 2:
- The protons flow back down their electrochemical gradient through an elaborate membrane protein machine called ATP synthase, which catalyzes the production of ATP from ADP and inorganic phosphate (Pi).
- This ubiquitous enzyme works like a turbine in the membrane, driven by protons, to synthesize ATP.
In this way, the energy derived from food or sunlight in stage 1 is converted into the chemical energy of a phosphate bond in ATP.
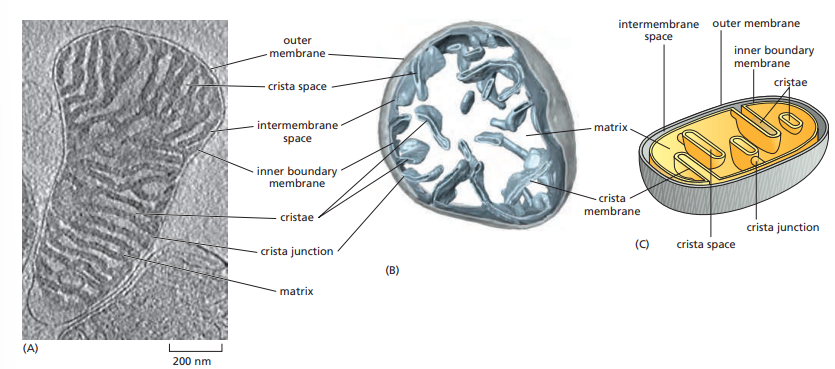
The Mitochondria
Occupy 20% of the cytoplasmic volume of a eukaryotic cell.
Mitochondria has an outer and inner membrane.
The two membranes have distinct functions and properties.
The inner mitochondrial membrane is folded to form invaginations known as cristae.
The narrow gap between the two membranes is called intermembrane space.
The outer mitochondrial membrane is freely permeable to ions and to small molecules as large as 5000 daltons.
The inner mitochondrial membrane is a diffusion barrier to ions and small molecules.
- It also contains the machinery for electron transport and ATP synthesis.
The membranes of the cristae:t
- They are continuous with the boundary membrane.
- It contains the ATP synthase enzyme that produces most of the cell’s ATP.
- They also contain the large protein complexes of the respiratory chain—the name given to the mitochondrion’s electron-transport chain.
The mitochondrial matrix contains a large variety of enzymes, including those that convert pyruvate and fatty acids to acetyl CoA and those that oxidize this acetyl CoA to CO2 through the citric acid cycle.
TCA (tricarboxylic acid cycle) or Kreb’s cycle or citric acid cycle
- This cycle was discovered by H.A. Krebs.
- TCA cycle occurs in the mitochondrial matrix.
- All the enzymes of the TCA cycle, except succinate dehydrogenase (in the inner mitochondrial membrane) present in the matrix.
- During Krebs cycle acetyl Co-A is completely oxidized into CO2.
- Kreb’s cycle is also called the citric acid cycle because 1st compound is citric acid. In this acid, 3 carboxylic groups are found so the process is also called TCA (Tricarboxylic Acid) cycle.
- In Kreb’s cycle oxaloacetic acid (OAA) is the first member and it also act as first acceptor of Acetyl Co-A.
- The TCA cycle starts with the condensation of the acetyl group with oxaloacetic acid and water to yield citric acid. The reaction is catalyzed by the enzyme citrate synthase and a molecule of CoA is released.
- Citrate is then isomerized to isocitrate. It is followed by two successive steps of decarboxylation.
- In the remaining steps of citric acid cycle, succinyl co-A is oxidised to OAA allowing the cycle to continue. During the conversion of succinyl co-A to succinic acid and a molecule of GTP is synthesized.
- Also there are three points in the cycle where NAD+ is reduced to NADH + (H+) and one point where FAD+ is reduced to FADH2.
- The continued oxidation of acetyl CoA via the TCA cycle requires the continued replenishment of oxaloacetic acid, the first member of the cycle. In addition it also requires regeneration of NAD+ and FAD+ from NADH and FADH2 respectively.
Oxidation or dehydrogenation occurs at 4 places in one Krebs cycle, results in the formation of 3NADH, 1 FADH2. Along with 1GTP (ATP) produced by substrate level phosphorylation in each turn of TCA cycle equals to 12 ATP.
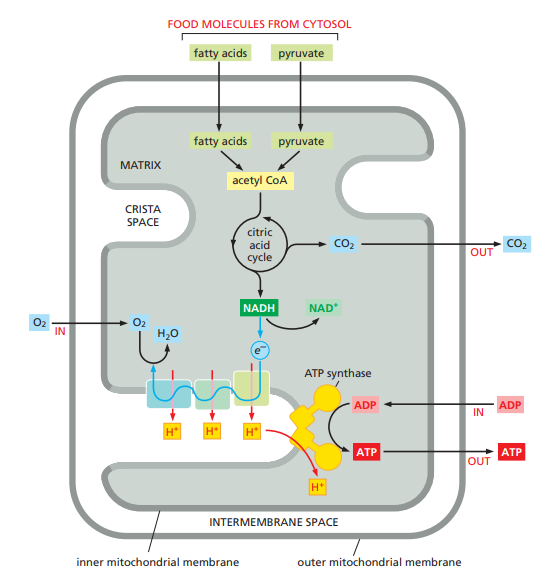
Electron Transport System (ETS)/ Oxidative Phosphorylation OR Respiratory Chain & (Terminal Oxidation of NADH + (H+) & FADH2)
All the reduced hydrogen acceptors like NADH + H+ and FADH2 move to the ETS where they release their hydrogen and get reoxidized to NAD+ and FAD+ so that they can again enter into the respiration process.
ETS is the chain of some hydrogen and electron carriers present in the inner mitochondrial membrane.
The significance of ETS is to remove hydrogens from reduced hydrogen acceptors NADH + H+ & FADH2. During this process, hydrogen acceptors get reoxidized and ATP are produced.
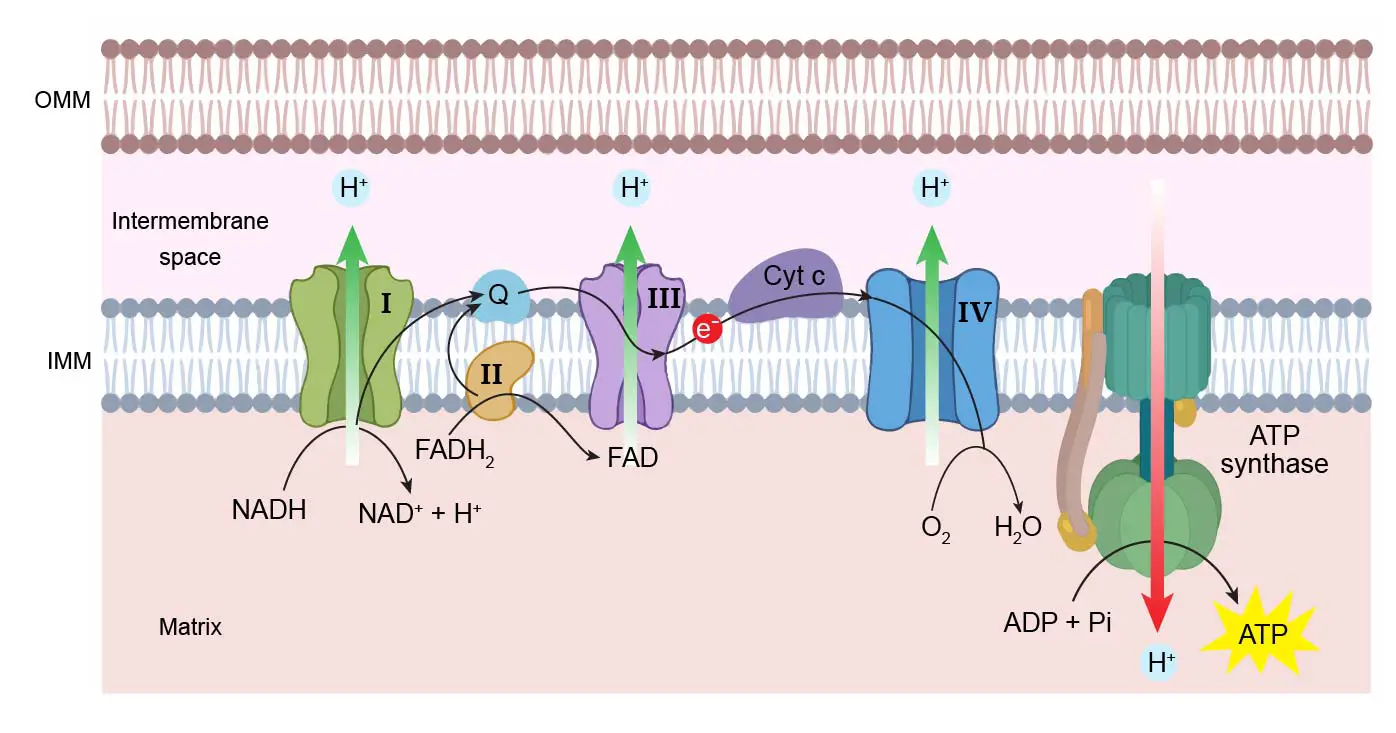
Now components of ETS are categorised as:
- Complex I: (NADH dehydrogenase complex): FMN, Fe-S
- Complex II: (Succinic dehydrogenase complex): FAD, Fe-S
- Complex III: (Cytochrome bc1 complex): Cytochrome b-Cyt c1, Fe-S
- Complex IV: (Cytochrome c oxidase complex): Cyt a and Cyt a3, 2Cu centers.
- Complex V: (ATP synthase/ ATPase/Oxysome): Fo (integral) - F1 (peripheral)
Electrons from NADH + H+ produced in the mitochondrial matrix during citric acid cycle are oxidised by an NADH dehydrogenase (complex I), and electrons are then transferred to ubiquinone located within the inner membrane.
Ubiquinone also receives reducing equivalents via FADH2 (complex II) that is generated during the oxidation of succinate in the citric acid cycle.
The reduced ubiquinone (ubiquinol) is then oxidised with the transfer of electrons to cytochrome c via cytochrome bc1 complex (complex III).
Cytochrome c is a small protein attached to the outer surface of the inner membrane and acts as a mobile carrier for transfer of elctrons between complex III and IV.
Complex IV refers to cytochrome c oxidase complex containing cytochromes a and a3, and two copper centres.
When the electrons pass from one carrier to another via complexes I and IV in the electron transport chain, they are coupled to ATP synthase (complex V) for the production of ATP from ADP and inorganic phosphate.
The number of ATP molecules synthesised depends on the nature of the electron donor. Oxidation of one molecule of NADH + H+ gives rise to 3 molecules of ATP, while that of one molecule of FADH2 produces 2 molecules of ATP.
Passage of 4H+ through F0 particle or proton channel leads to synthesis of 1 ATP.
Although the aerobic process of respiration takes place only in the presence of oxygen, the role of oxygen is limited to the terminal stage of the process.
- Yet, the presence of oxygen is vital, since it drives the whole process by removing hydrogen from the system. Oxygen acts as the final hydrogen acceptor.
Unlike photophosphorylation, where it is the light energy that is utilised for the production of proton gradient required for phosphorylation, in respiration it is the energy of oxidation-reduction utilised for the same process. It is for this reason that the process is called oxidative phosphorylation.
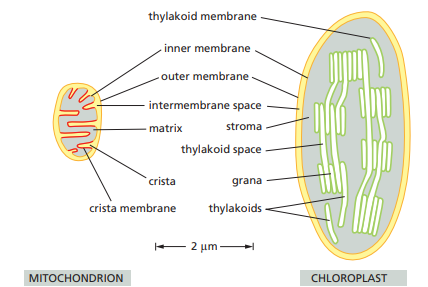
Chloroplast
Photosynthesis occurs in a specialized intracellular organelle.
Chloroplasts use chemiosmotic mechanisms to carry out their energy interconversions in much the same way that mitochondria do.
They have:
- a highly permeable outer membrane
- a much less permeable inner membrane, in which membrane transport proteins are embedded
- a narrow intermembrane space in between.
The inner chloroplast membrane surrounds a large space called the stroma, which is analogous to the mitochondrial matrix.
The chloroplast has its own genome and genetic system.
The stroma therefore also contains a special set of ribosomes, RNAs, and chloroplast DNA.
The inner membrane of the chloroplast is not folded into cristae and does not contain electron-transport chains.
The electron-transport chains, photosynthetic light-capturing systems, and ATP synthase are all contained in the thylakoid membrane.
- It is a separate distinct membrane that forms a set of flattened, disc-like sacs called thylakoids.
Light Reaction
Light reactions or the photochemical phase include light absorption, water splitting, oxygen release, and formation of high energy chemical intermediates like ATP and NADPH.
Photosystems are required for this process.
Emerson and Arnold worked on chlorella and gave concept of two photosystem or two pigment system.
The groups of photosynthetic pigments in the thylakoid membrane are known as photosystem.
The pigments are organised as two discrete photochemical light harvesting complexes (LHC) within the photosystem I and photosystem II.
In every photosystem there is a reaction centre surrounded by accessory pigments.
The reaction centre is different in both the photosystems.
- In PS I the reaction centre chlorophyll-a has an absorption peak at 700 nm, hence it is called P700.
- In PSII it has absorption peak at 680 nm, and it is called P680.
These are named in the sequence of their discovery, and not in the sequence in which they function during the light reaction.
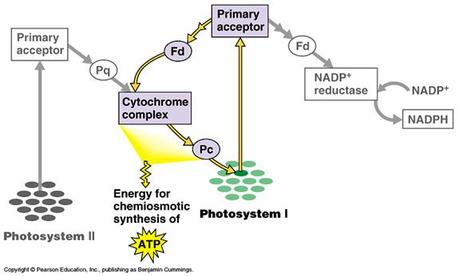
The electron transport/ phosphorylation
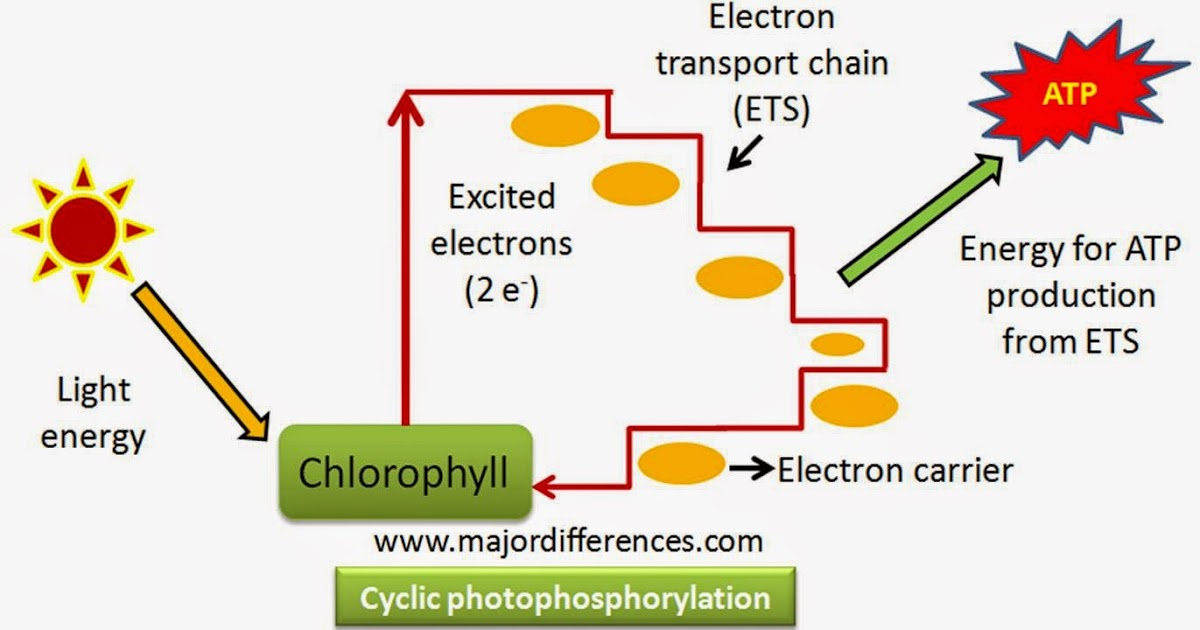
- Cyclic photophosphorylation:
Light Absorption: Chlorophyll and other pigments in the thylakoid membrane absorb light energy from the sunlight.
Electron Excitation: The absorbed light energy excites electrons in the chlorophyll molecules, raising them to higher energy levels.
Electron Transport: Excited electrons are passed through a series of electron carriers embedded in the thylakoid membrane. As electrons move along this transport chain, they release energy.
ATP Synthesis: The released energy is utilized to pump protons (H+) across the thylakoid membrane, establishing an electrochemical gradient. The accumulated protons create a potential energy difference, which is harnessed by ATP synthase, an enzyme complex embedded in the membrane. ATP synthase uses this energy to convert ADP (adenosine diphosphate) and inorganic phosphate (Pi) into ATP.
Return of Electrons: After releasing energy, the electrons that were excited in step 2 return to the chlorophyll molecule from which they originated. This cyclic flow of electrons is characteristic of cyclic photophosphorylation.
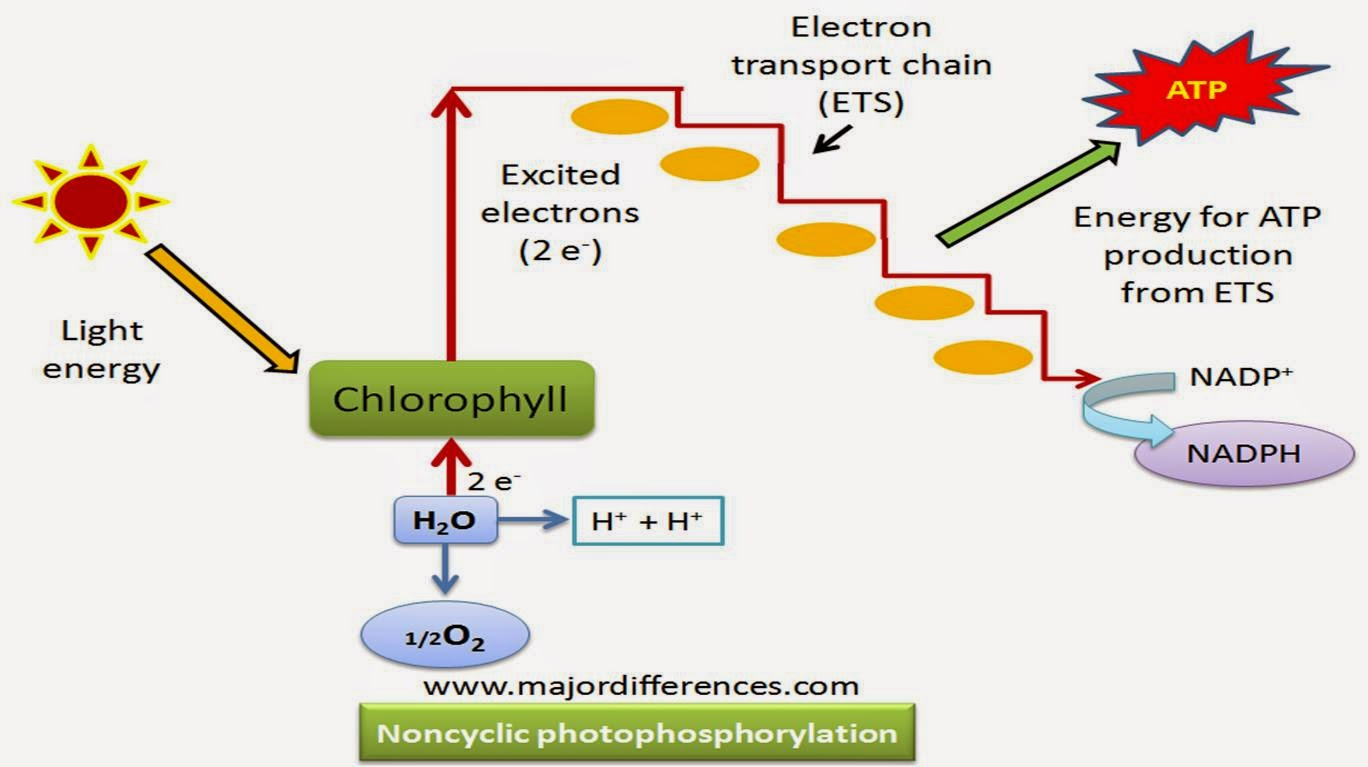
- Non-Cyclic photophosphorylation:
Non-cyclic photophosphorylation is a process that occurs during the light-dependent reactions of photosynthesis.
It takes place in the thylakoid membrane of chloroplasts in plants, algae, and some bacteria.
The primary goal of non-cyclic photophosphorylation is to produce both ATP (adenosine triphosphate) and NADPH (nicotinamide adenine dinucleotide phosphate).
The process begins with the absorption of light energy by chlorophyll and other pigments in the thylakoid membrane.
The absorbed light energy excites electrons in chlorophyll, raising them to higher energy levels.
Excited electrons are transferred to electron carriers in a series of redox reactions, creating an electron transport chain.
As electrons move along the electron transport chain, they release energy that is used to pump protons (H+) across the thylakoid membrane, establishing an electrochemical gradient.
The accumulated protons create a potential energy difference, which is harnessed by ATP synthase to produce ATP through the process of chemiosmosis.
Simultaneously, during non-cyclic photophosphorylation, NADP+ (nicotinamide adenine dinucleotide phosphate) molecules accept the electrons at the end of the electron transport chain, becoming reduced to NADPH.
NADPH, along with ATP, serves as a source of energy and reducing power for the subsequent light-independent reactions (Calvin cycle) of photosynthesis.
The ultimate purpose of non-cyclic photophosphorylation is to capture and convert light energy into chemical energy in the form of ATP and NADPH, which are used to fuel the synthesis of carbohydrates and other organic compounds in the Calvin cycle.
Where are the ATP & NADPH used?
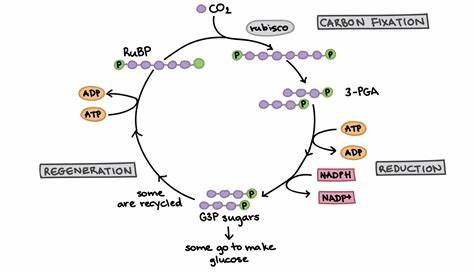
The Calvin cycle or C3 Pathway
The Calvin cycle, also known as the light-independent reactions or dark reactions, is a series of biochemical reactions that occur in the stroma of chloroplasts in plants, algae, and some bacteria.
Its primary function is to convert carbon dioxide (CO2) into carbohydrates, particularly glucose, through a series of enzymatic reactions.
The Calvin cycle is named after Melvin Calvin, who elucidated its major steps during photosynthesis.
The cycle consists of three main phases: carbon fixation, reduction, and regeneration of the CO2 acceptor molecule.
Carbon Fixation:
- CO2 molecules from the atmosphere are captured and attached to a five-carbon sugar called ribulose-1,5-bisphosphate (RuBP).
- This reaction is catalyzed by the enzyme RuBisCO (ribulose-1,5-bisphosphate carboxylase/oxygenase).
- The resulting unstable six-carbon molecule splits into two three-carbon molecules called 3-phosphoglycerate (3-PGA).
Reduction:
- ATP and NADPH, generated during the light-dependent reactions, provide the energy and reducing power for the conversion of 3-PGA into glyceraldehyde-3-phosphate (G3P).
- Some of the G3P molecules produced are used to regenerate RuBP, while others are utilized to synthesize glucose and other carbohydrates.
Regeneration of the CO2 Acceptor:
- The remaining G3P molecules in the cycle undergo additional reactions to regenerate the initial CO2 acceptor molecule, RuBP.
- ATP is consumed in these reactions to rebuild the RuBP molecules, preparing them for another round of carbon fixation.
The Calvin cycle is a cyclic process, meaning that some of the intermediate compounds are recycled to sustain the continuous production of carbohydrates.
The cycle must take place multiple times to generate one molecule of glucose, as six cycles are required to fix six molecules of CO2.
The Calvin cycle is crucial for the synthesis of organic molecules, including glucose, which serves as a primary energy source and building block for plants and other organisms in the food chain.
It operates in close coordination with the light-dependent reactions, as the ATP and NADPH produced during the light reactions power the Calvin cycle's energy-demanding reactions.
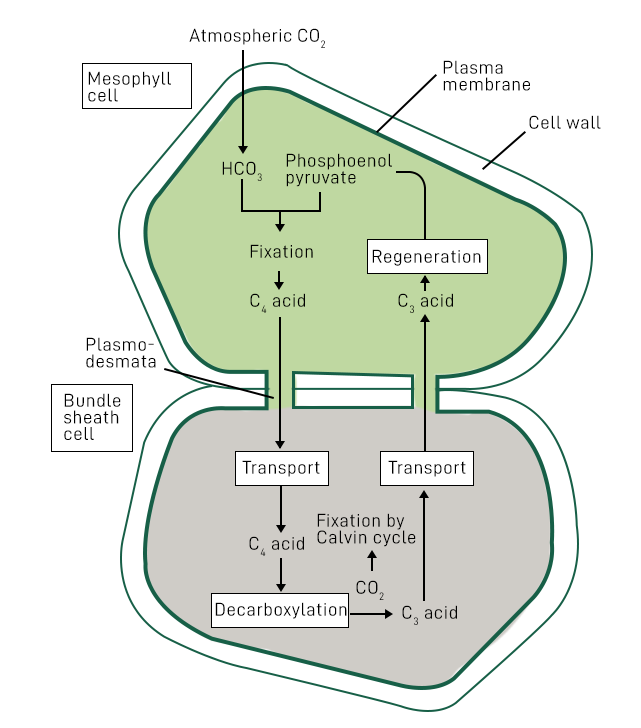
C4 Pathway/ Hatch & Slack Pathway
The C4 pathway is an alternative photosynthetic pathway that some plants have evolved to enhance carbon dioxide (CO2) fixation, particularly in hot and dry environments.
It is named after the four-carbon compound (C4) that is initially formed during the pathway.
The C4 pathway is found in plants such as maize (corn), sugarcane, sorghum, and certain grasses.
Unlike the typical C3 pathway, where CO2 fixation and the Calvin cycle occur in the same cells (mesophyll cells), the C4 pathway separates these processes into different cell types.
The initial CO2 fixation step occurs in specialized cells called mesophyll cells, which are located near the surface of the leaf.
In mesophyll cells, CO2 combines with a three-carbon molecule, phosphoenolpyruvate (PEP), to form a four-carbon compound called oxaloacetate (OAA) in the presence of the enzyme PEP carboxylase.
The four-carbon compound, OAA, is converted to malate or aspartate and transported to bundle sheath cells, which are located deeper within the leaf.
In the bundle sheath cells, CO2 is released from the transported four-carbon compounds and enters the Calvin cycle, where it is utilized for carbohydrate synthesis.
The separation of CO2 fixation and the Calvin cycle into different cells reduces the effects of photorespiration and increases the efficiency of carbon fixation, especially under high light and high temperature conditions.
The C4 pathway requires additional energy in the form of ATP to power the biochemical reactions involved in the initial CO2 fixation and the transport of four-carbon compounds.
Overall, the C4 pathway allows plants to minimize water loss through reduced stomatal opening, increase photosynthetic efficiency, and adapt to environments with high light, high temperature, and low CO2 conditions.
It is an example of biochemical and anatomical adaptations that enable certain plants to thrive in arid or semi-arid regions.
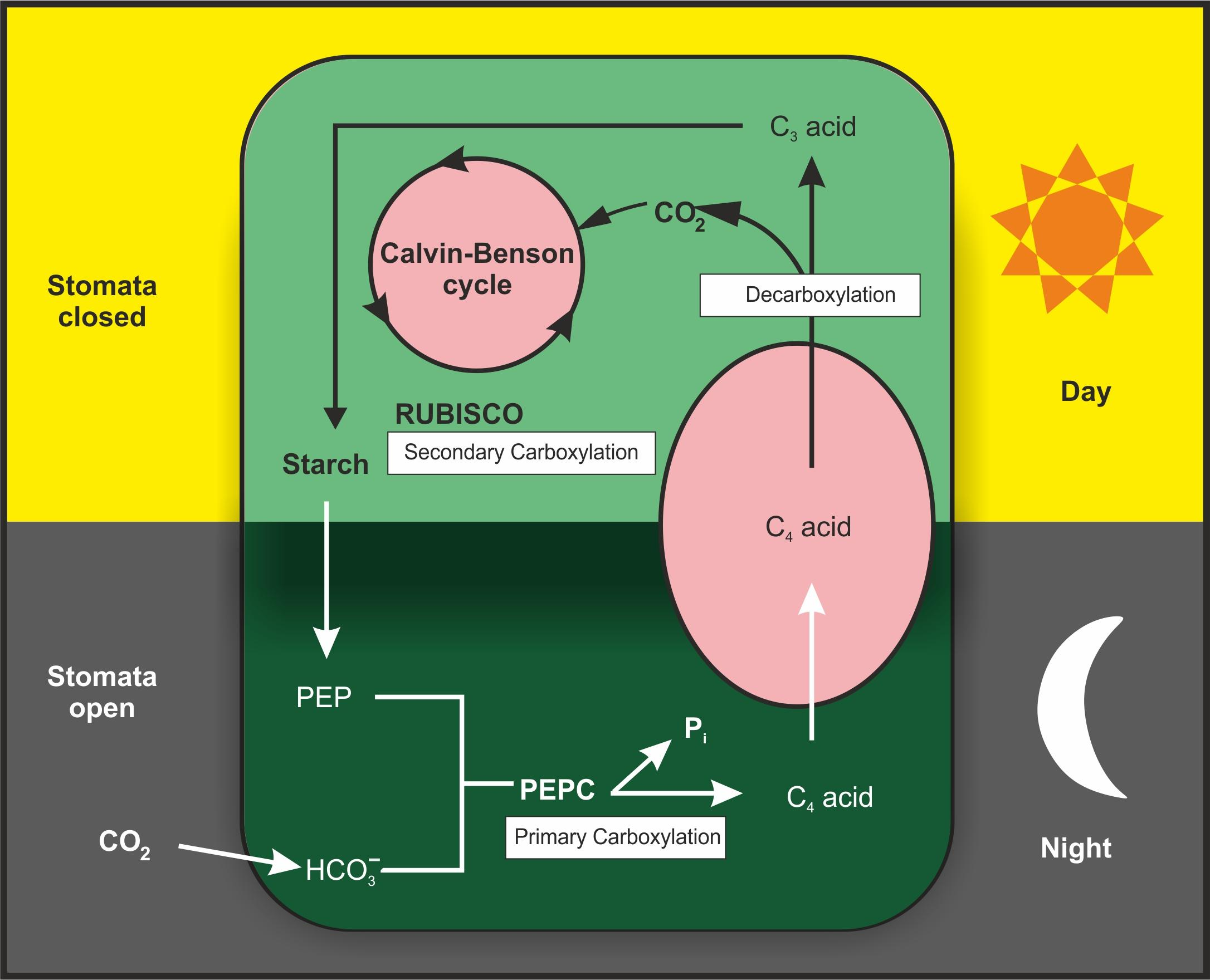
CAM Pathway
The CAM pathway is a specialized form of photosynthesis observed in certain plants, particularly succulents, that grow in arid or semi-arid environments.
CAM plants include species such as cacti, pineapple, and some orchids.
The CAM pathway allows plants to conserve water by opening their stomata and conducting gas exchange during the night, when temperatures are cooler and humidity is higher.
During the night, CAM plants open their stomata and take in carbon dioxide (CO2), which is converted into a four-carbon organic acid called malate or another similar compound.
The malate is stored in large vacuoles within the plant's cells until the following day.
During the day, when the stomata are closed to prevent water loss, the stored malate is broken down, releasing CO2 that enters the Calvin cycle for photosynthesis.
The CO2 released from malate enables carbohydrate synthesis and the production of energy-rich molecules like glucose.
The CAM pathway allows plants to efficiently use CO2 by storing it at night and releasing it during the day, avoiding excessive water loss through transpiration.
This pathway maximizes water-use efficiency in arid environments, where water availability is limited.
CAM plants have adapted their leaf anatomy and physiology to carry out this pathway, including specialized cells and biochemical mechanisms that regulate the storage and release of CO2.
The CAM pathway is an example of a plant's ability to adapt to water-stressed conditions and efficiently utilize available resources.