Bio 131 1st LE
Introduction to Plant Development
Development → process that builds an organism
What has changed?
Analyzes the process by which the change has come about
Approaches to the study of plant development
Analysis of the molecular genetic mechanisms that underlie developmental processes
Characterization of the biochemical reactions that carry out development
Investigations of the structures of cells and how these structures help bring about developmental changes
Investigations of the integrated functions of tissues and organ systems
Genetics of Control of Plant Development
Zygote → single origin of all the cells in plant body
Alll cells in plants contain the same genetic material
Differential gene expression → expression of different genes by cells with the same genome
Differences between cell types are not due to the presence of different genes but due to the expression of different genes
Regulation of Gene Expression
Regulation of gene expression can occur at any of the steps of protein synthesis
Levels of control
Transcriptional level → during formation of primary transcript
Processing level → at the stage of splicing
Transport of mRNA from nucleus to cytoplasm
Translational level
Protein modification/transport
Characteristics of Plant Development
Continuous Development
Reiterative process
Ex: Apical meristems repeat the same developmental patterns to produce an extending root or an extending series of nodes and internodes
Indeterminate → open ended patterns of development
Ex: shoot apical meristem
Determinate → patterns restricted in time and space
Ex: leaf development because leaf meristem activity stops after the leaf is produced
Developmental patterns may switch from indeterminate to determinate, vice versa
Ex: vegetative SAM becomes determinate if it becomes a floral meristem
Ex: determinate plant organs may give rise to indeterminate adventitious root or shoot buds
Plastic Development
Development can be adjusted according to the prevailing environmental conditions
Important for sedentary organisms a.k.a plants
Cabomba caroliniana
Their underwater leaves are feathery → protects them from damage by lessening their resistance to moving water
Their surface leaves are pads that aid in flotation
Both leaves are genetically identical, but different environments result in the turning on or off of different geens during development
Regeneration and Totipotency
Totipotency → ability to become any organ or cell and can give rise to a complete plant
Callus → amorphous mass of cells which can then reorganize and differentiate like the cells of meristems
Regeneration may or may not involve callus formation
In the case callus formation is not needed, missing tissues are directly replaced by highly organized cell proliferation and differentiation
A comparison of plant and animal development
Post-Embryonic vs. Embryonic developemnt
Animal development is almost synonymous with embryogenesis
Ex: most adult organs are formed during embryogenesis
Animal body plans are predetermined by embryonic development
Angiosperm embryogenesis is concerned with establishment of the meristems so major organs and tissue systems are not yet found in mature embryo
Major organs and systems are only formed after seed germination → post-embryonic
Plants can adapt their body plan to environment changes → great plasticity
Nature of Cell commitment for differentiation
Plants can retain totipotency
Ex: mesophyll cells can be made to re-differentate into tracheary elements
Animals are irreversibly committed to a developmental pathway
Ex: neurons cannot be induced to become muscle cells
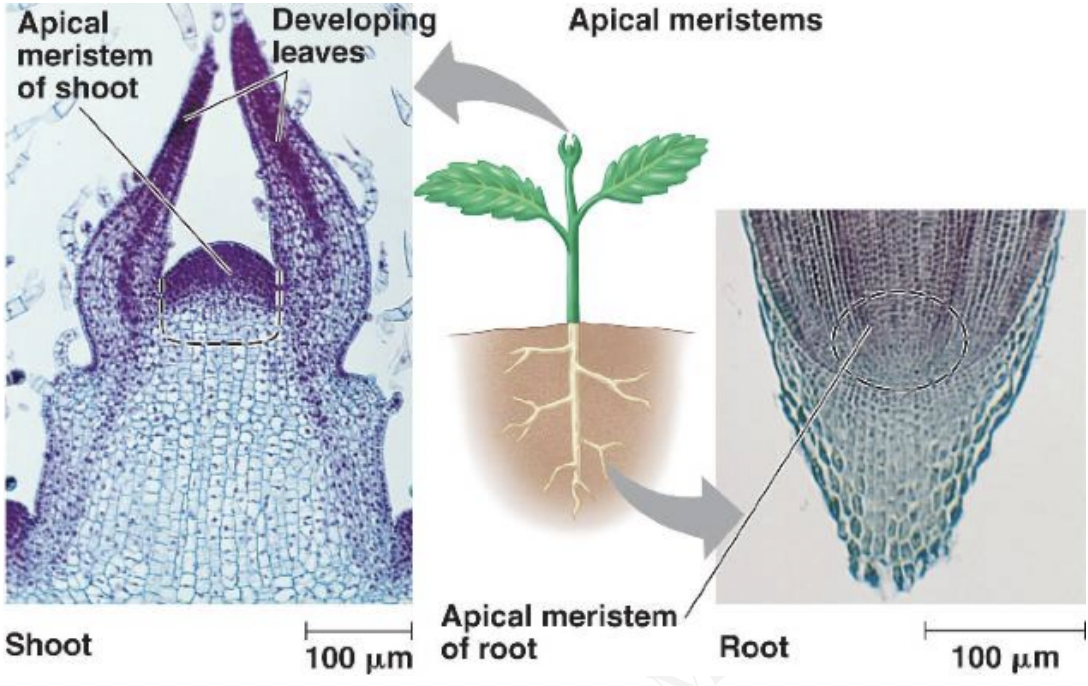
Cell movement and planes of cell division
Animal cells are motile and cell migration is important → gastrulation
Developmental fate of plant cells determined by its position in the plant body
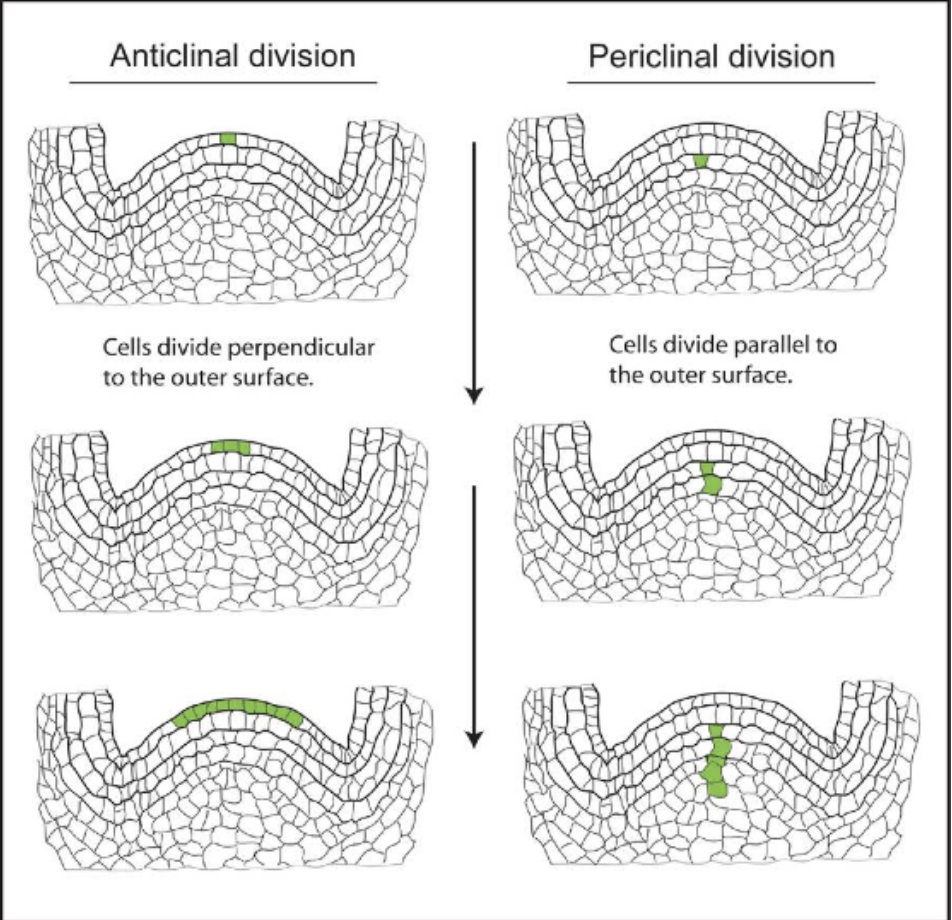
Anticlinal division → cell plates perpendicular to surface → expansion of surface
Periclinal division → cell plates parallel to surface → protrusion from surface
Ex: epidermis → anticlinal division → expands as a single layer covering entire surface
Ex: pericycle → periclinal division → outgrowth of lateral roots in primary root
Variety of plant organs and cell types
Higher animals have a greater variety of organs and cell types than higher plants
Embryos of eudicots have 4 organs
Plumules → embryonic axis above the cotyledons containing epicotyle
Cotyledons
Hypocotyls → embryonic axis below the cotyledons but above radicle
Radicle → Embryonic root
Mature plants have 3 vegetative organs
Stems
Roots
Leaves
Flowers have 4 organs
Sepals
Petals
Stamens
Pistils
40 cell types in plants compared to hundreds of cell types in higher animals
Intercellular signaling network between plant cells is less complicated

Control of Cell Fate
In multicellular organisms, cell fates are influenced by activities of neighboring cells
Regulation of cell fate dependent on ability of cells to transduce intercellular and extracellular information into changes in gene activity
Both plants and animals adopted the transcriptional cascade as principal mechanism for cell fate determination
How plant and animals police cell fates
Animal cells require survivial factors from neighboring cells
Programmed cell death → happens in the absence of survival factors + safeguards that is disabled in cancer cells
Plants cells have the capacity for apoptosis but it is not invoked by isolation
A plant cell displaced out of its normal position will just sqitch to a fate appropriate to its new position
Consequences of Autotrophy vs Heterotrophy
In plants, assimilation of energy and of nutrients and water are achieved at separate sites (leaves and root hairs respectively)
Ex: more leaves → more sunlight and CO2 absorbed
Ex: more roots → more water and mineral uptake
In animals, energy and nutrients come from food
Ex: development of new organs will not improve feeding ability
Ex: changes in environmental conditions cause a change in animal behavior
Thus, different morphology between plants and animals
Model organisms in Plant development
Model organism → short generation time, characterized genome, similar to a member of a particular group
Ex: Arabidopsis thaliana → mustard family
What makes Arabidopsis thaliana a good model organism?
Small Size
Short lifecycle → 6-8 weeks from germination to production of new crop seeds
Self-fertilizing → bisexual flowers and can self-pollinate so recessive mutations quickly become homozygous and mutatnt phenotype observed faster
High seed Production
Small genome
Ideal for genetic characterization of mutants
Use of mutants in the study of plant development
Most mutations result in a loss of function of a gene
Ex: MALE STERILITY or MS1 gene → no stamen development → thus this gene is required for stamen development
Nomenclature for genes identified by mutation
Full descriptive names of the wild-type gene should be capitalized and italicized (i.e. ALPHABETICA)
Full descriptive names of the mutant should be in lower case and italized (i.e. alphabetica)
Mutant gene symbols should be written in lower case letters and italicized (i.e. abc) while wild type gene symbols should be capitalized and italicized (i.e. ABC)
Protein products of genes should be written in capital letters without italics (ABC)
Ex:
Wildtype gene: SHORT INTEGUMENt 1 (SIN1)
Mutant gene: short integument 1 (sin1)
Protein product: SIN1
Plant Developmental Processes
Development vs. Growth
Development is the progression from earlier to later stages in maturation
Process whereby tissues, organs, and whole plants are produced
Involves growth, morphogenesis, and differentiation
Growth is the irreversible change in size of cells and plant organs due to both cell division and cell enlargement
Enlargement requires a change in elasticity of the cell walls together with increase in size and water content of vacuole
Determinate growth when an organ or part or whole organism reaches a certain size and then stops growing
Indeterminate growth when cells continue to divide indefinitely (i.e. plants)
Cell division is the number of cells increasing due to mitosis
Symmetry, rate, and orientation greatly affects cell fate
Cell enlargement is the size of a cell increasing due to an increase in the volume of its protoplasm
Differentiation, dedifferentiation, and redifferentiation
Differentiation is the process in which generalized cells specialize into the morphologically and physiologically different cells
It is a function of which particular genes are either expressed or repressed thus guided by gene expression
In plants, it involves changes in size, biochemistry, structure, and function of cells
Ex: meristematic cells of procambium and vascular cambium differentiate into xylem elements
Cells undergoing differentiation may undergo major structural changes
Ex: differentiation of tracheary elements involves losing the protoplasm and developing a secondary cell wall
Dedifferentiation is the reversal of cell development in plants so that the differentiation that had occurred previously is lost and the cell becomes more generalized in structure
Living differentiated cells that have lost the capacity to divide can regain capacity of division
Ex: in the formation of meristems, both interfascicular cambium and cork cambium develop from fully differentiated cells
Redifferentiation is the differentiation after dedifferentiation takes place either forming the same mature cell type or an entirely different one
Ex: callus tissue produced from a mature leaf explant can be induced to differentiate into roots or shoots
Pattern Formation
Process whereby organisms create spatially-ordered and reproducible structures
Organisms must noy only generate different cell types, they must also ensure that the different cells are correctly arranged in time and space
Organized cell growth and cell architecture
Preferential elongation of cells along certain axes is a major determinant of form
Orientation of cell growth is in the plane perpendicular to the orientation of the cellulose microfibrils in cell wall
Elongation favored when CMFs are oriented transversely to the direction of growth
Elongation limited when CMFs are oriented in oblique or longitudinal direction
Control of cell proliferation
Placement and activity of meristems are major determinants of shape and form
Lateral meristems account for increases in girth
Apical meristems account for elongation of plant axis
External cues
Initial arrangement of cells may use external signals in order to create a spatial pattern
Phototropism/gravitropism
Assymetric cell divisions
Early plant embryo results in unequally sized daughter cells
Creates polarity and permit unequal distribution of cellular material
Ex: first division of the angiosperm zygote results in a smaller apical cell and a larger basal cell
Lateral inhibition
Interactions between neighboring cells that prevent both from adopting the same fate
Ex: during stomata formation, precursor cells inihibit the development of other precursor cells close to them, ensuring spaces in between
Programmed cell death
Tissue can be sculpted by the selective death of some cells
Ex: formation of holes in Monstera leaves
Plant cell division - finding the sweet spot for cell plate insertion
Cytokinesis in cells of flowering plants is achieved through the construction of a new cell wall form the inside out
Cell plate is initiated between daughter nuclei after mitosis and expands centrifugally to form a new cell wall sandwiched between new plasma membranes
Plant cytokinesis proceeds through a microtubule-dependent mechanisms
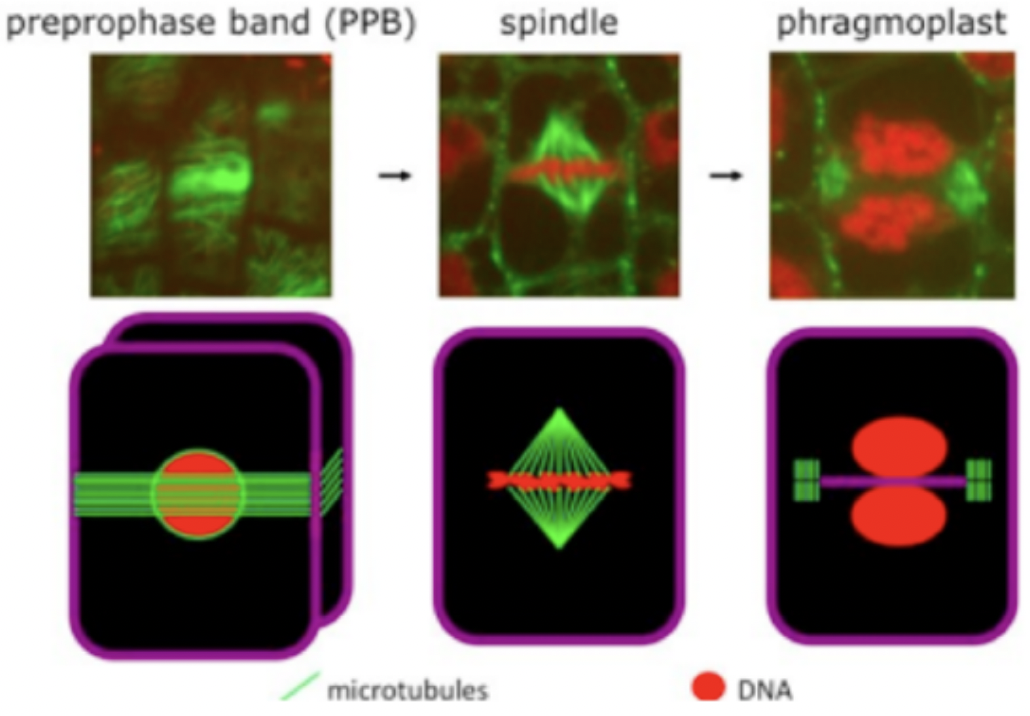
Involves mitotic spindle, pre-prophase band and phragmoplast
Pre-prophase band (PBB) is a ring like microtubule and actin structure formed around the pre-mitotic nucleus
During mitosis, actin dissociates from the PPB and the accumulation of a yet unknown factor marks division site
Found in the G1 phase of cell cycle and disappears when M phase starts
Mitotic spindle gives rise to Phragmoplast that expands toward the cell cortex
Composed of two opposing disks of parallel microtubules and actin filaments
+ ends near the equatorial plane and - ends near the poles
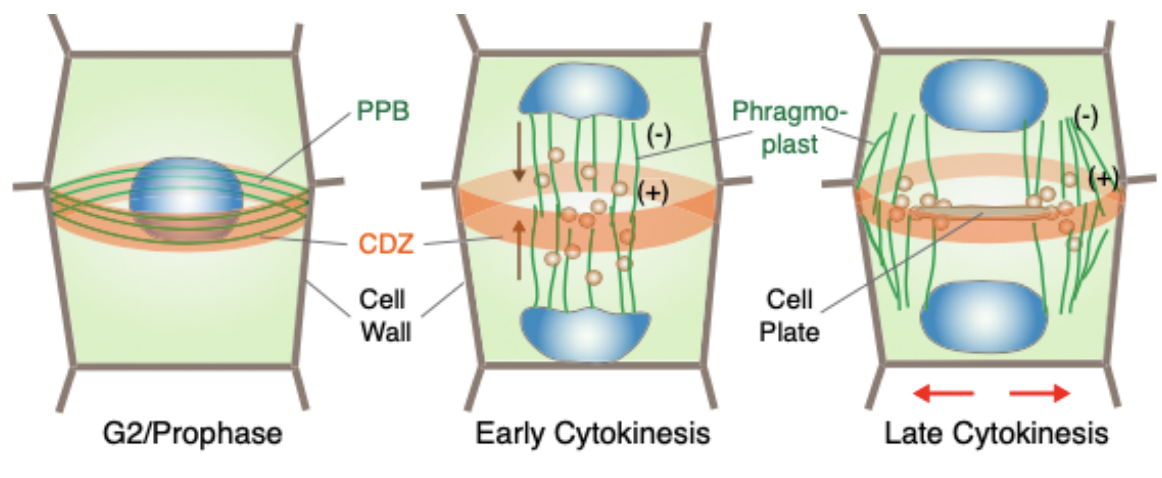
Cortical division zone (CDZ) is the division plane in plant cells that is outlined by the PBB in response to selection cues
Phragmoplast guides the vesicles containing cell wall material toward the division plane for fusion (brown arrows)
Cell plate and phragmoplast expand toward the CDZ in a centrifugal direction (outward pointing red arrows)
Subsequent membrane trafficking and fusion at its edge leads to cell plate formation initiated through the fusion of golgi-derived vesicles
Phragmoplast guides the vesicles to the cell plate containing polysaccharides, proteins, and membranes
Plant Cell Wall
Meristematic cells are usually isodiametric and then differentiate by developing distinct forms to acquire specific functions
Dynamic structures that act as an exoskeleton by participating in the establishment and maintenance of cell shape and by protecting the cell content
Primary cell walls are made of glucan-based cellulose microfibrils embedded in a highly hydrated matrix composed of
pectins,
hemicelluloses,
structural proteins, and
proteoglycans
Also extensible to allow cell expansion which is driven by strong intracellular turgor pressure
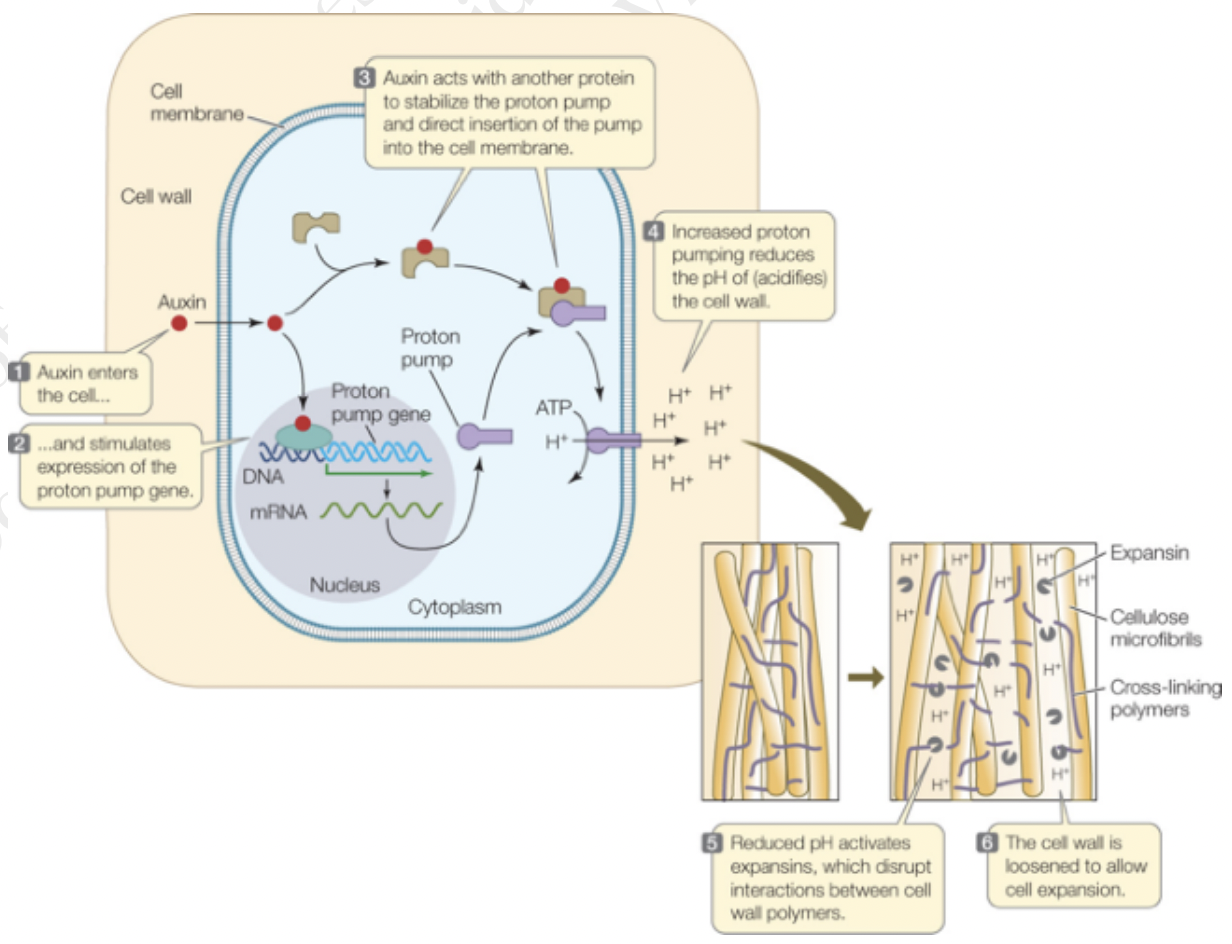
Auxin is a stimulator of cell elongation, as it increases cell wall extensibility and regulates cell wall properties by initiating wall loosening
Secondary cell walls are usually present in specialized, non-growing cells
Cellulose microfibrils
Embedded in components such as non-cellulosic polysaccharides and structural proteins
Determine the direction of cell expansion
Cellulose synthesis takes place beneath the cell wall at the plasma membrane via cellulose synthases
Cellulose microfibril patterning is mediated via cortical microtubules and cellulose synthases
Microtubules as raild road tracks thus controlling microfibril organizationg by guiding cellulose synthase complexes
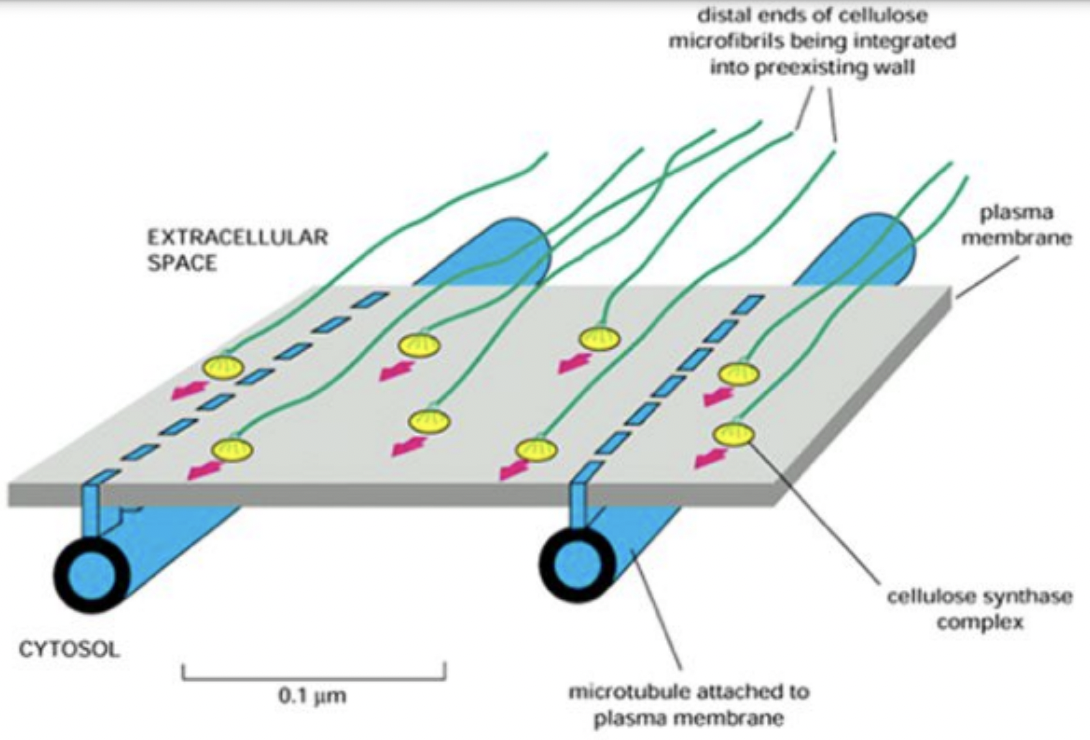
Interaction between Autonomous and microtubule guidance systems controls cellulose synthase trajectories
Cellulose synthase trajectories remained the same when microtubules were disrupted
Cellulose synthase complexes can interact with trails left by other complexes → autonomous mechanism
This mechanism can be overridden by the microtubule guidance system
Dual guidance model
Autonomous system with interactions between cellulose synthases and microfibrils maintain aligned cellulose synthase trajectories
Microtubule guidance system allowing alignments to be steered by environmental and developmental cues
Hemicelluloses and Pectins
Hemicellulose xyloglucans are found mainly in primary cell walls and participate in cell wall extension during cell elongation
influence wall extensability and stiffness
Pectins are important in the regulation of wall properties by controlling wall porosity and hydration causing wall swelling and thus influences wall thickness
Adjust wall extensibility by influencing the alignment of cellulose microfibrils
Form the middle lamella → adhesive compartment between two adjacent cell walls
Composed of highly heterogenous polysaccharides
Homogalacturonan
Rhamnogalacturonan
Rhamnogalacturonan II
Xylogalacturonan
Structural proteins
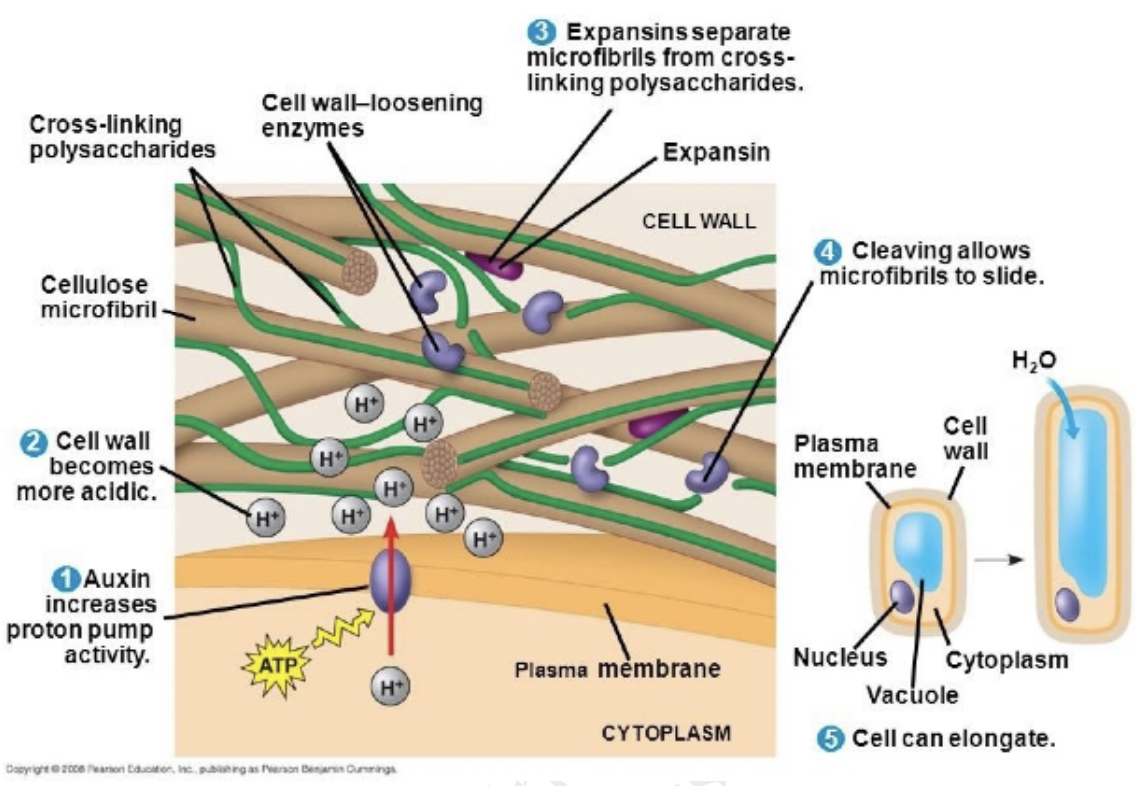
Expansins are cell wall loosening proteins and enhances wall expansion in acidic pH
Extensins are required for cell wall assembly
Arabinogalactan proteins AGPs play a role in plant protection known to specifically control pollen tube growth but also regulated overall plant development
Role of Auxin in Wall extension
Controls plant growth and development by promoting cell division (proliferation), growth (expansion, elongation), and differentiation
Auxin or indole-3-acetic acid (IAA)
Responsible for cell wall loosening and cell expansion via modifications of cell wall composition
Causes pectin polymerization
Increases pectin viscosity and xyloglucan depolymerization
Activates the expression of cell wall related genes
Stimulates the synthesis of proton pumps leading to apoplast acidification
Acts on the cytoskeleton through Rho GTPases and promotes trafficking of vesicles containing new cell wall material
Recall
[1] auxin activates H+ -ATPases
[2] apoplast acidification
[3] wall loosening proteins like expansins become active
[4] causes wall enlargement
[5] changes trigger the cell to activate calcium channels to pump calcium into the wall
[6] pH increases
[7] growth cessation
Auxin Signaling and Gene Expression
Auxin acts through the transport inhibitor resistance 1 or Auxin signalling F-box (TIR1) nuclear auxin receptor family
F-box proteins target other proteins for degradation via ubiquitin degradation pathway
TIR1 proteins bind to auxin → auxin acts as a glue → proteins bind to their targets
Auxin/Indole-3-Acetic Acid (AUX/IAA) is a target of TIRs which represses auxin-mediated transcription through the interactions with Auxin Response Factors (ARFs) in the absence of auxin
Binding of TIR promotes degradation of AUX/IAAs
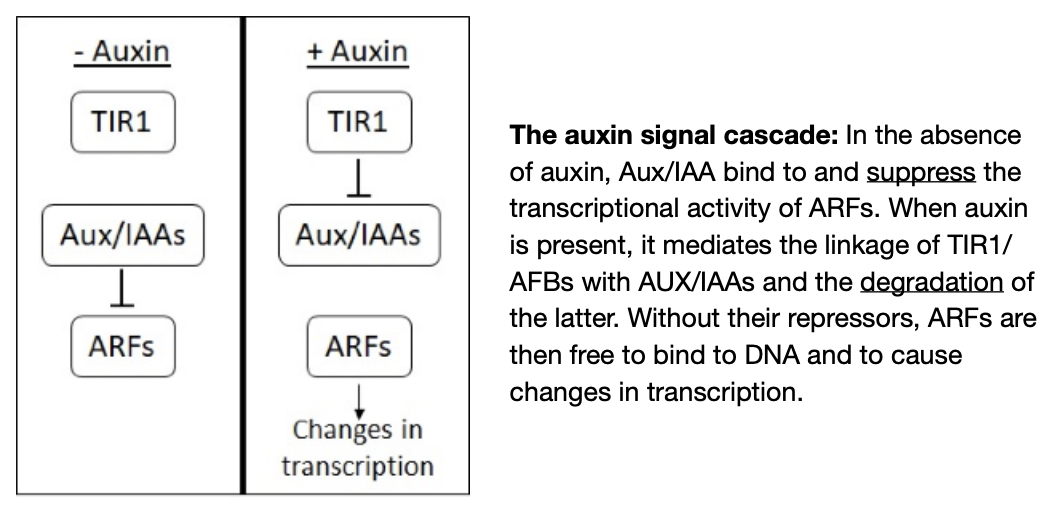
Auxin is absent
ARFs bind to Aux/IAAs (repressor)
Aux/IAA suppresses the ability of ARFs to enhance gene transcription
Binding of Aux/IAA to ARFs brings Aux/IAA into contact with promoters of auxin regulated genes where Aux/IAA repress the expression of these genes by recruiting other factors to make modifications to the DNA structure
Auxin is preset
Binding of auxin to TIR1/AFBs allows them to bind to Aux/IAAs → Aux/IAAs are marked for degradation through proteosomal activity → frees ARF proteins → activate or repress genes at whose promoters they are bound
TIR1 → protein receptor, ARFs → transcription regulator, Aux/IAA → inhibitor of ARFs
Auxin and Cell wall related genes
Auxin treatment results in the upregulation of key genes related to cell wall components
Reported genes are not necessarily related to cell elongation (wall expansion) and could be linked to different auxin-driven processes such as cell division, growth or differentiation
Movement of materials through apoplast
Interconnected cell walls of plant form the apoplast
Apoplast acts as a barrier and a channel for intercellular signals
They allow movement of small negative charged molecules
Apoplastic route is continuous with the lumen of xylem vessels (except for endodermis cell walls which are suberized)
Molecules secreted into the cell wall may be transported via the xylem providing a unidirectional route (root to shoot) for long range developmental signals
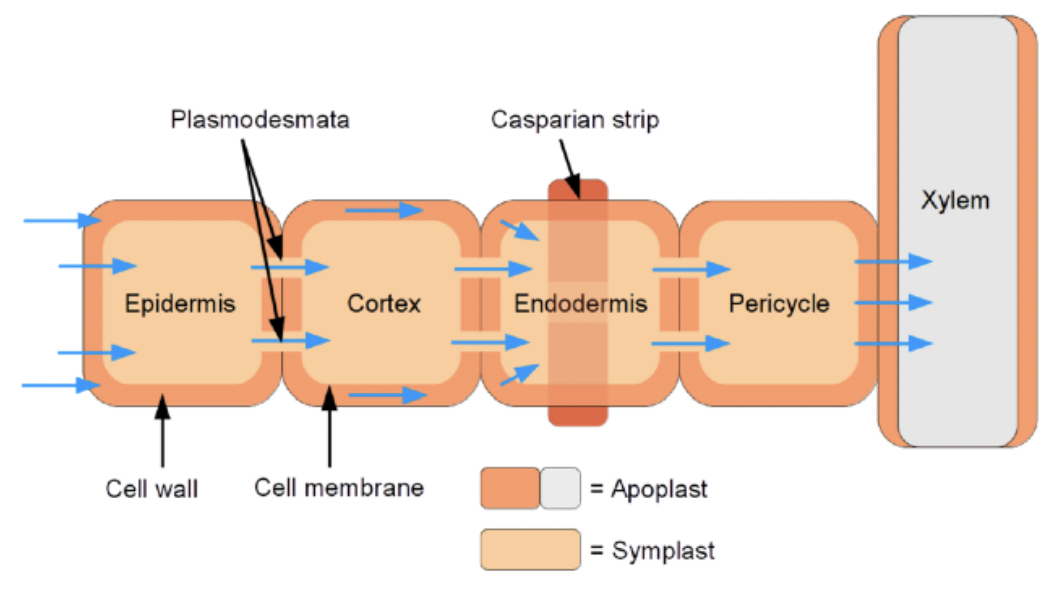
Movement of materials through plasmodesmata
Plasmodesmata connect adjacent cells
Each plasmodesmata is lined by a membrane continuous with the plasma membrane of the connected cells
Symplast formed by the connected protoplasm of cells throughout the plant
Symplast includes the cytoplasm of the phloem elements and provides another route besides the xylem for long range bidirectional transport
Symplastic communication includes
Passive diffusion for smaller molecules
Active transport for large nucleic acids and proteins
Some parts act so well connected they act as a syncytium
Review: Angiosperm Life Cycle
Angiosperm Life Cycle
Dominated by the spore generating sporophyte stage rather than sexual gametophyte stage
Produce two types of spores: male and female gametes
Megaspore produced in a carpel (ovary + ovules)
Each ovule contains a megasporangium where megaspores are produced
Megasporocyte within each megasporangium is a megaspore mother cell
Megasporophyte produces 4 megaspores where 1 survives and develops into an embryo sac consisting of an egg and other cells
Microspores originate from anthers at the tips of stamens
Within an anther are microsporangium containing microsporocyte
Microsporocyes produce microspores via meiosis
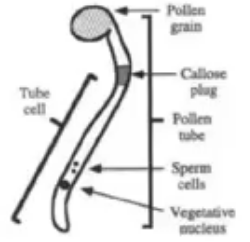
Miscrospore develops into a pollen grain which contains a tube cell and a generative cell
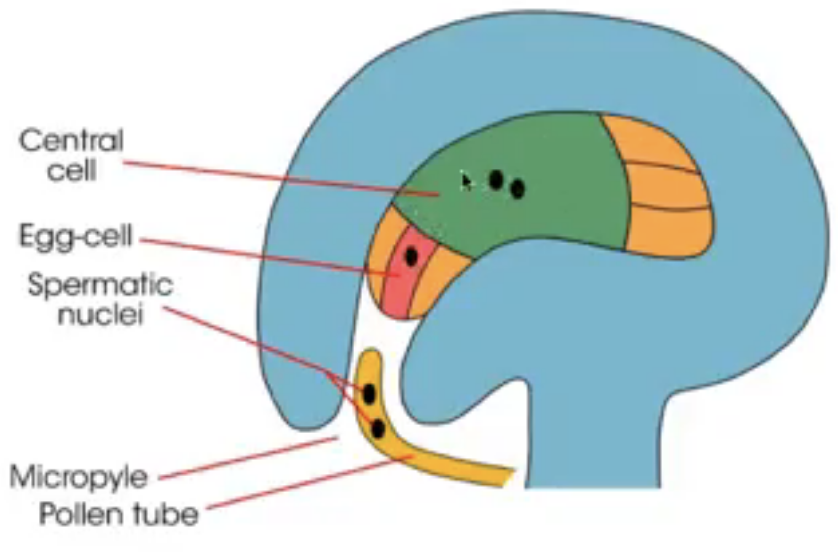
Once the pollen reaches the stigma, the upper part of the carpel, tube cell becomes a pollen tube that extends to the ovule containing the embryo sac
Generative cell divides to form 2 sperm which are released together in an act of double fertilization
One sperm fertilizes the egg and one fertilizes the central cell
Fertilized egg develops into a zygote then into an embryo
Fertilized central cell becomes an endosperm
Endosperm and embryo are packed into a seed
Sporophyte and Gametophyte generation
A haploid generation alternates with a diploid generation where diploid is the sporophyte and haploid is the gametophyte
Both sporophyte and gametophyte are always multicellular
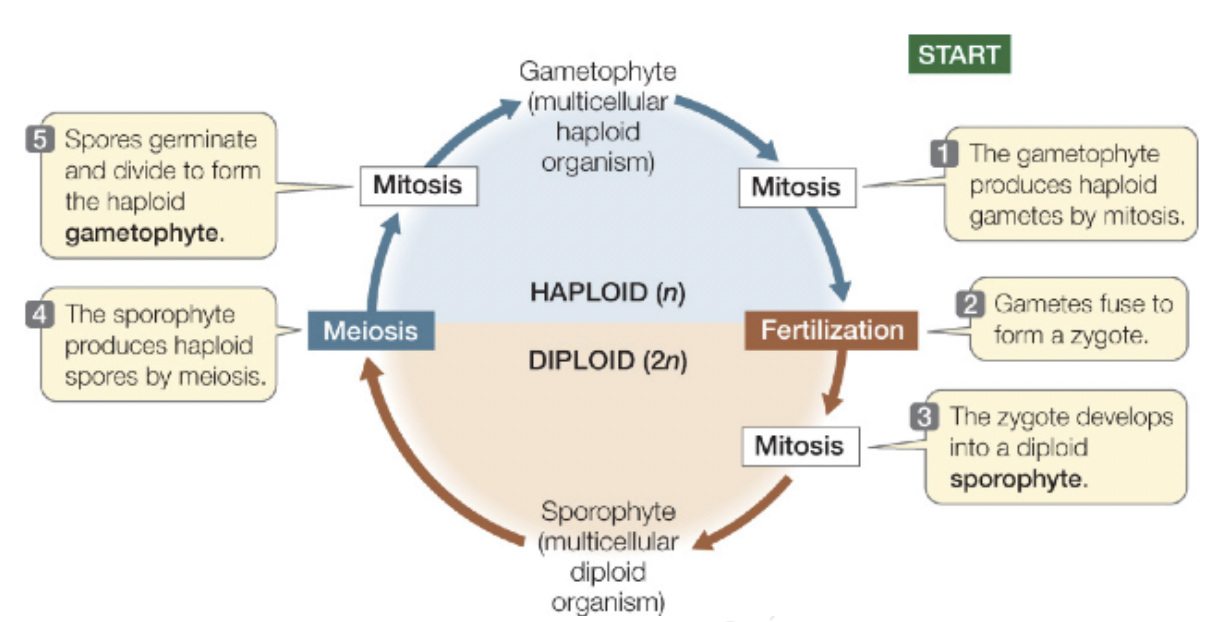
Sporophyte (2n) generation produces unicellular spores (n) via meiosis
Spores (n) germinate and divide via mitosis to produce multicellular gametophytes (n)
Gametophyte produces gametes egg (n) and sperm (n) via mitosis
Two haploid gametes join to form a diploid zygote
Zygote (2n) divides to form multicellular sporophyte (2n)
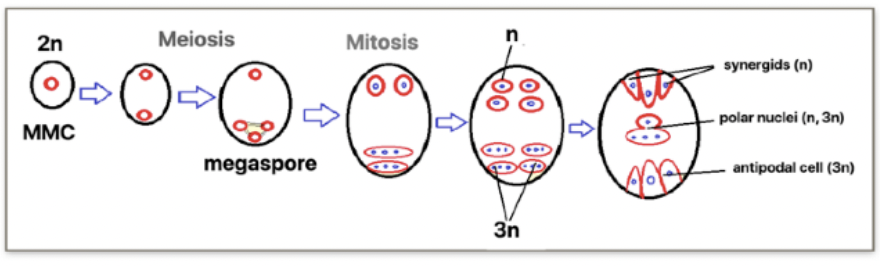
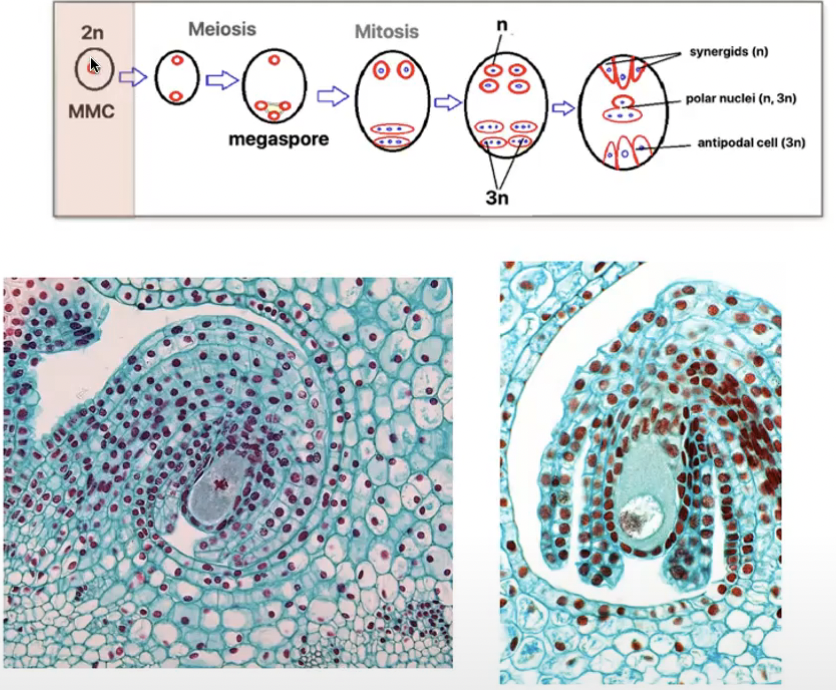
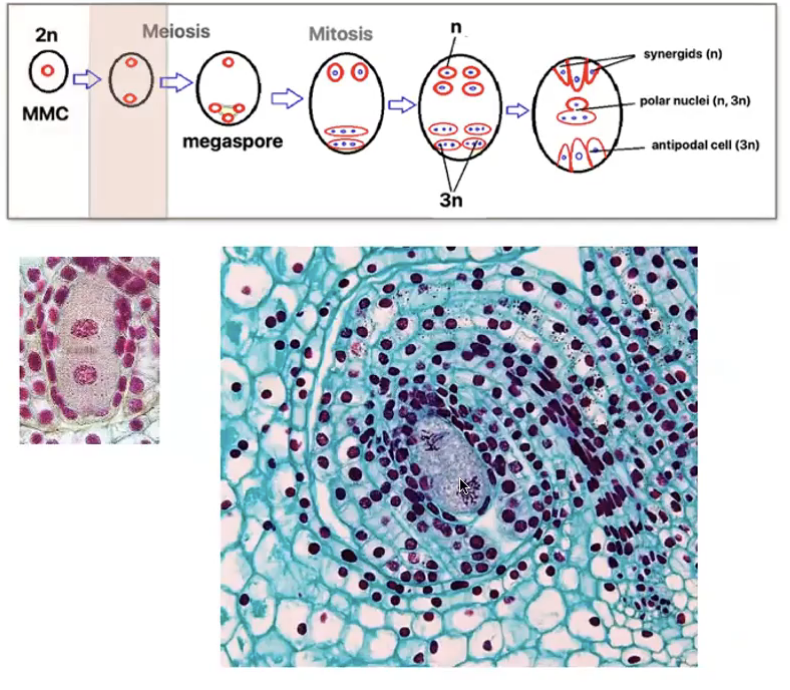
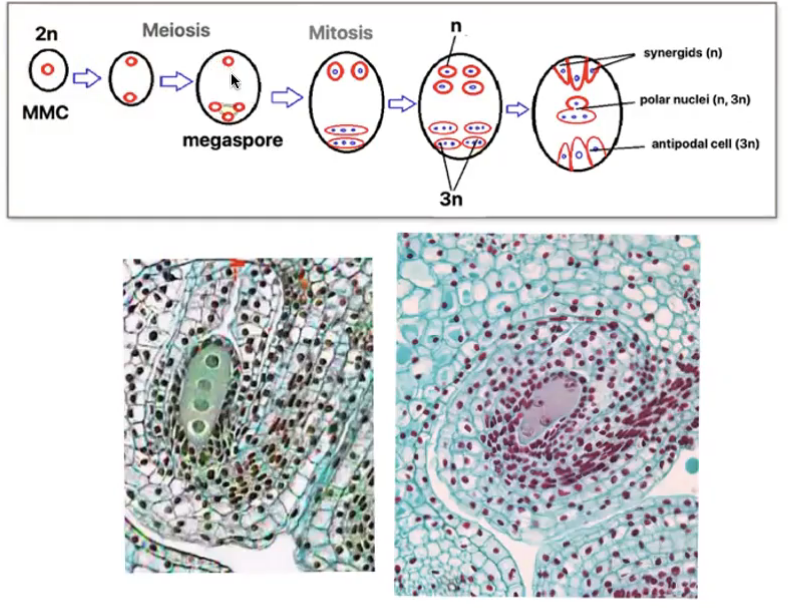
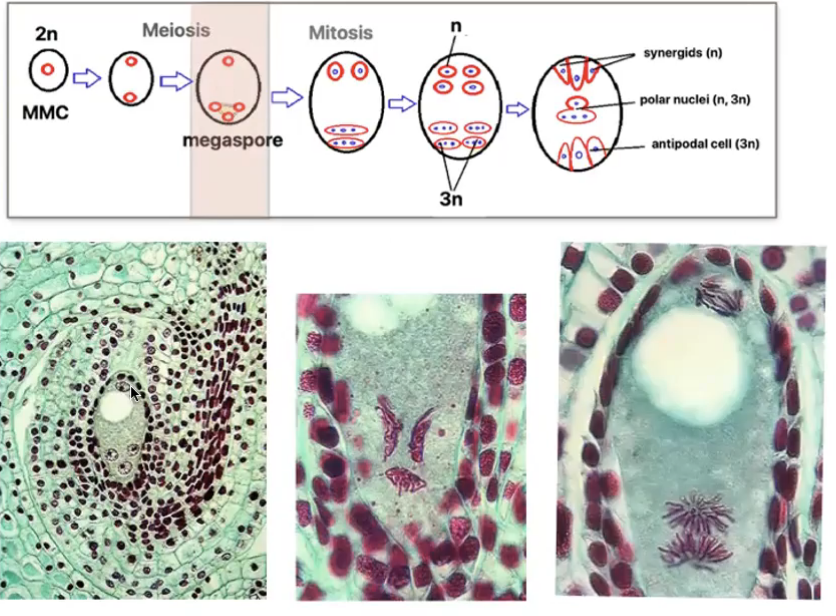
Angiosperm megasporogenesis and Megagametogenesis
Megasporogenesis
Megasporocyte (2n) in the nucellus of the megasporangium undergoes meiosis to produce 4 haploid megaspores (n) where in only 1 survives
Megagametogenesis forms the female gamete
Surviving megaspore divides by mitosis 3 times without cytokinesis to produce a huge cell with 8 nuclei
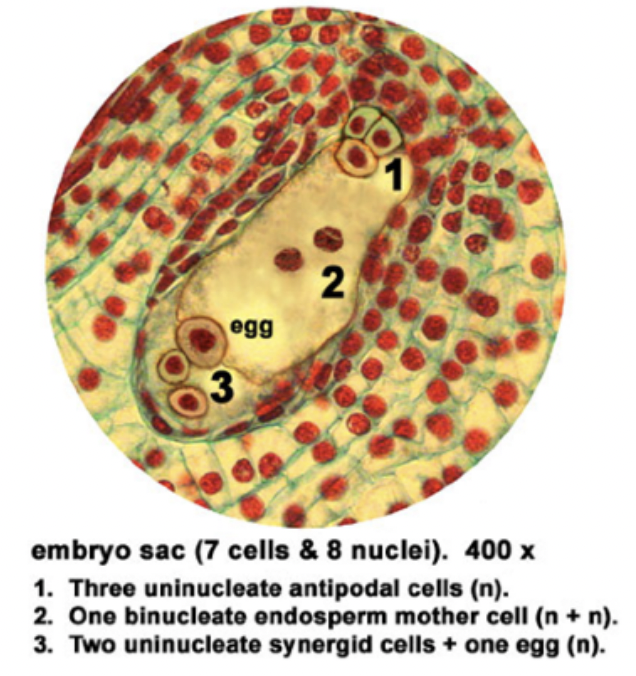
Multinucleate structure is divided to form the 7 celled embryo sac → female gametophyte
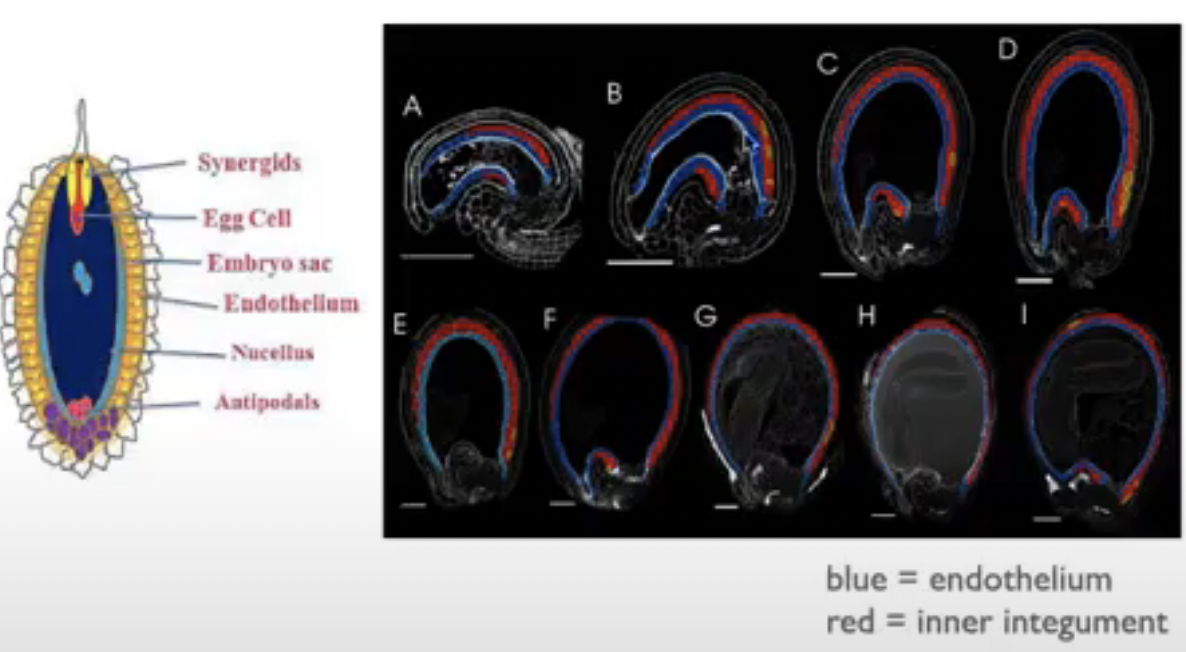
Cells of embryo sac
1 central cell (2n) → later fuses with sperm to form triploid endosperm
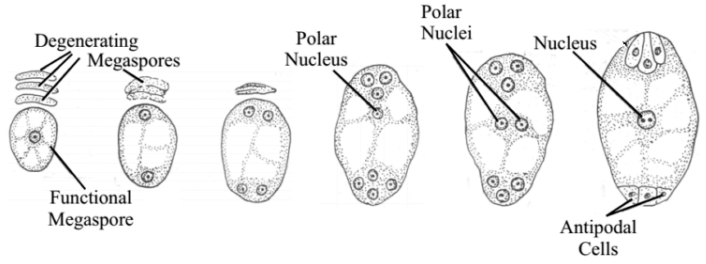
3 antipodal cells (n) → positioned at the ends of the embryo sac opposite the micropyle
1 egg cell (n) → positioned closest to the micropyle becomes the egg cell
2 synergid cells (n) → positioned at either side of the egg that help attract and guide the pollen tube for successful fertilization (egg apparatus)
Angiosperm Microsporogenesis and Microgametogenesis
Microsporogenesis
Anther of the stamen contains four microsporangia or pollen sac
Each pollen sac contains diploid microsporocytes (2n) that divide via meiosis to produce haploid microspores (n)
Each microsporocyte gives rise to four microspores that later develop into pollen grains
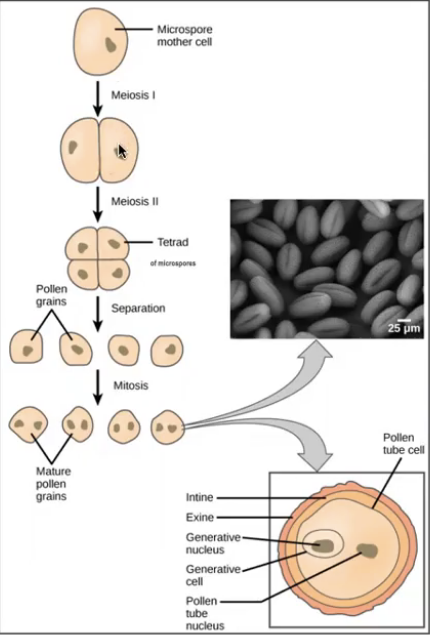
Pollen sac has a layer of cells, tapetum, that provides nutrition to microspores and contributes to pollen wall
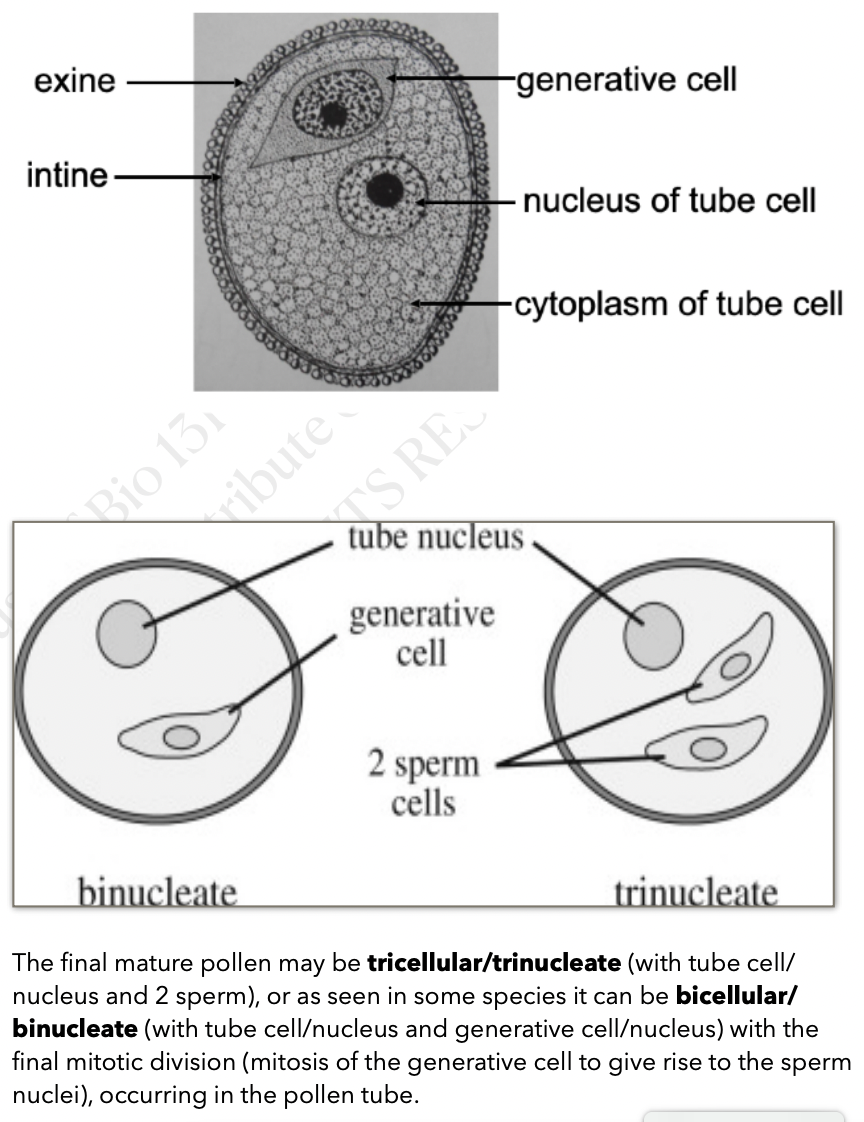
Each pollen grain has two coverings
Exine → thicker outerlayer that contains sporopollenin a complex waterproofing substance supplied by tapetal cells
Intine → inner latyer
Microsporogenesis
Microsporocyte (2n) divides via meiosis to produce 4 microspores (n) that initially occur in tetrads
Released from tetrads to form pollen grains
Haploid nuclei of pollen grain divides via mitosis
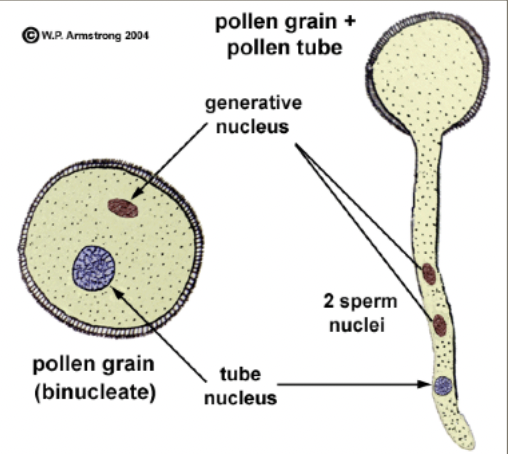
Mature pollen grains containing two cells the generative cell that is contained within the tube cell
Generative cell divides via mitosis to produce 2 sperm nuclei
Sperm delivery
When pollen grain lands on stigma, it absorbs water and germinates to produce a pollen tube which is an extension of the cytoplasm of the tube cell
No cell division as pollen tube elongates
Pollen tube grows via tip growth
Pollen tube elongates through style the generative cell divides by mitosis to produce 2 sperm nuclei
Tube nucleus leads ahead of two sperms as tube grows towards micropyle in response to chemical attractants produced by synergids
Arrival of pollen tube = death of one synergid = serving as a passage way into embryo sac
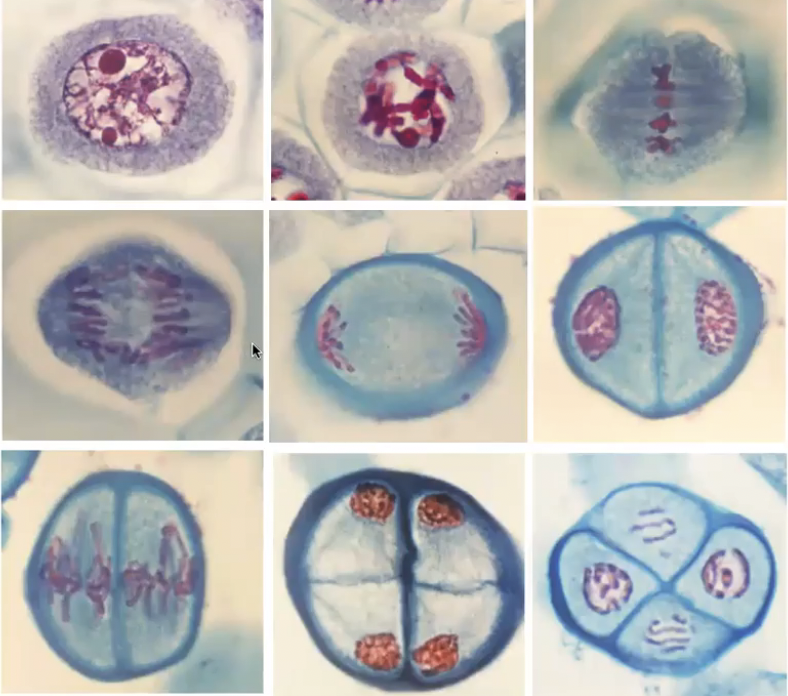
Meiosis in Angiosperms
Microsporogenesis and megasporogenesis are the two processes in plants that involve meiosis
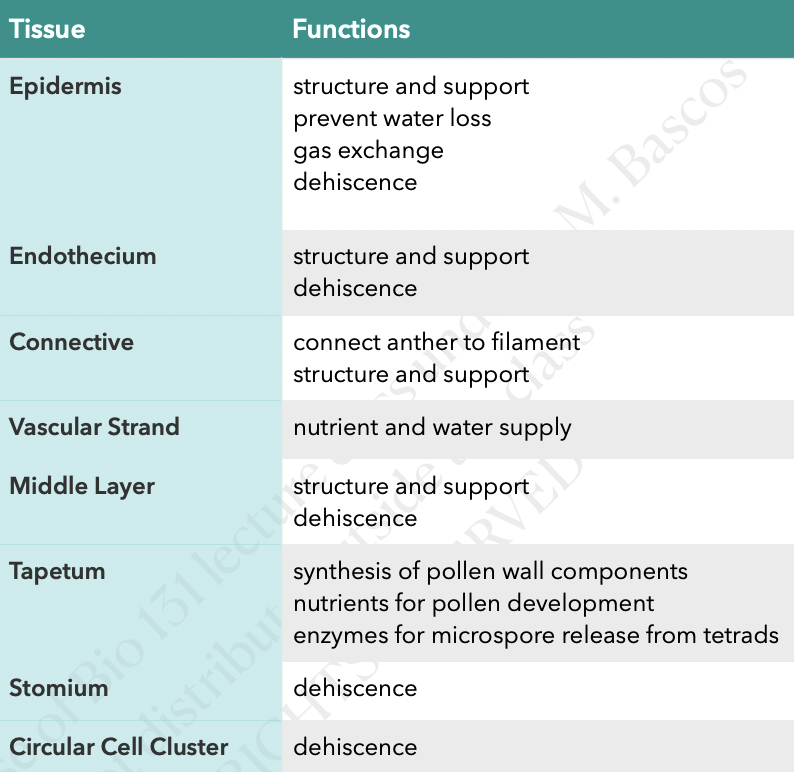
Stamen and Pollen Development

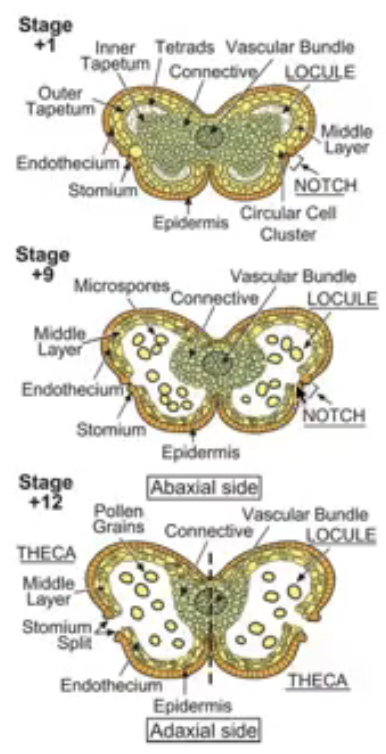
Section of a flower bud not a mature flower
6 developing stamens surrounding the developing ovaries
Stamen structure
Concerned with male sexual reproduction (anther is where microsporogenesis occurs)
Consists of the anther and filament
In gumamela, the filaments are fused to form the staminal tube
Androecium refers to all the 6 stamens collectively
Anther is a two-lobed organ with 2 locules in each lobe
Each locule has a microsporangium or pollen sac
Anther is a group of 4 microsporangia
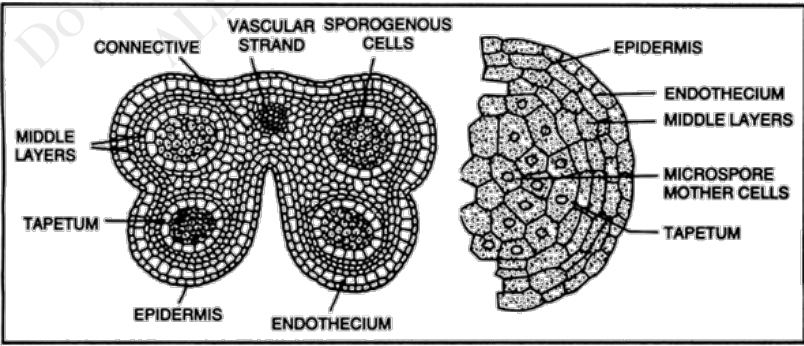
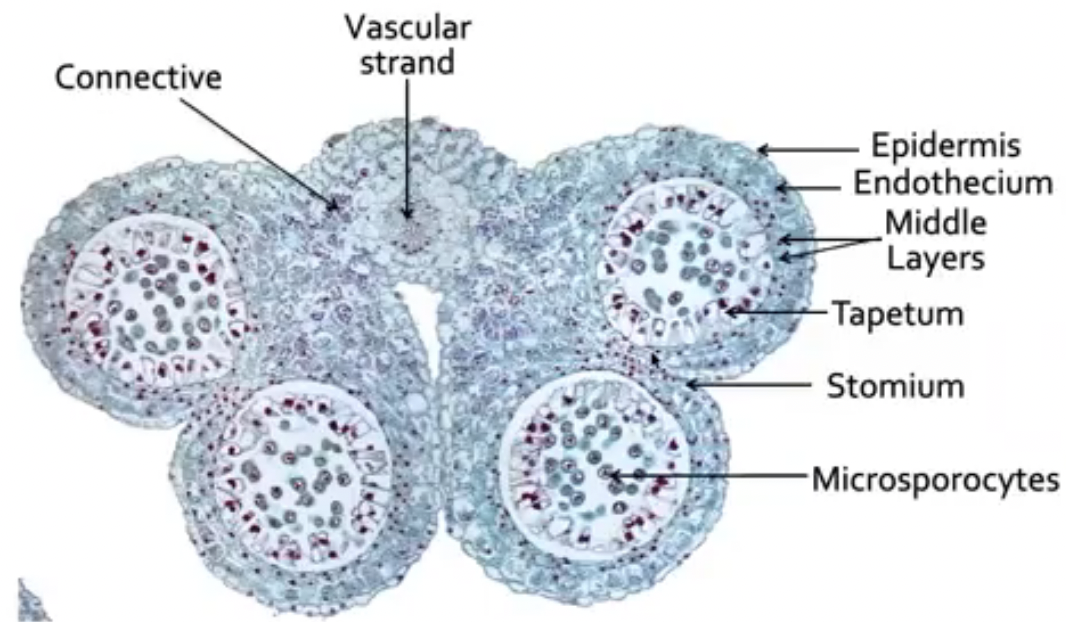
Outermost layer of cells is epidermis
Connective is the tissue found in between two lobes (joins the two lobes of the anther)
Vascular strand (with xylem and phloem tissue) is embedded in the connective
Endothecium is inner to epidermis with radially elongated cells
Endothecium cells develop fibrous thickening made of cellulose with pectin and lignin which helps in dehiscence of anther
Stomium are thin-walled cells in between the cells of the endothecium
Joins adjacent anther walls
Serves as final breakage site (opening site) for dehiscence
Middle layers is next to endothecium which become compressed and obliterated in mature anther
There are species without a middle layer
Tapetum circumscribes the locule and nearest to developing microspores
Cicular cell cluster can only be found in tobacco and other solanaceous plants
Composed of idioblast cells that accumulate calcium oxalate crystals
Cells undergo apoptosis before dehiscence
Degeneration of ccc and connective allows 2 locules of each theca to become confluent to form a unified chamber so pollen grains can be released from stomium region
Degeneration of ccc breaks the septum between the two pollen sacs to make them continuous so when the stomium opens the spores from both sacs are released together
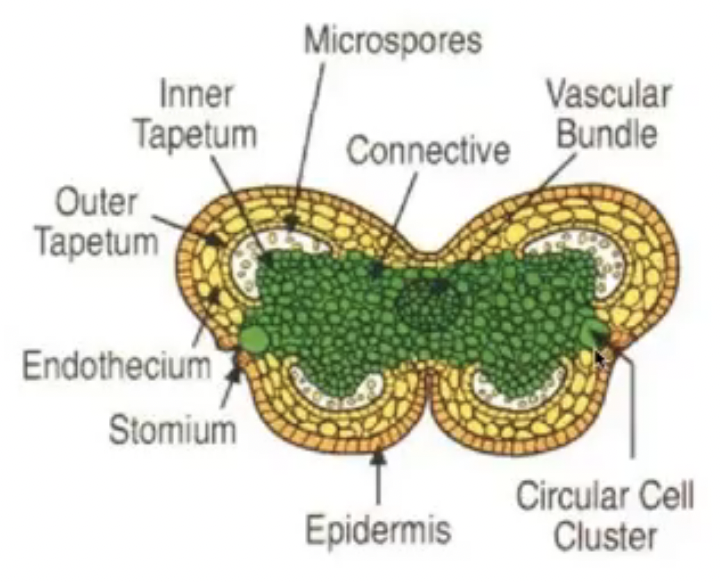
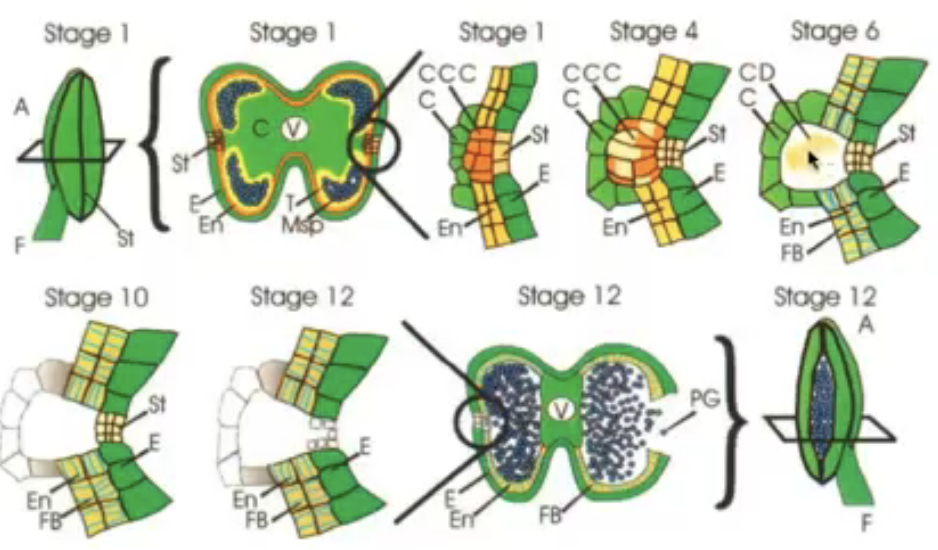
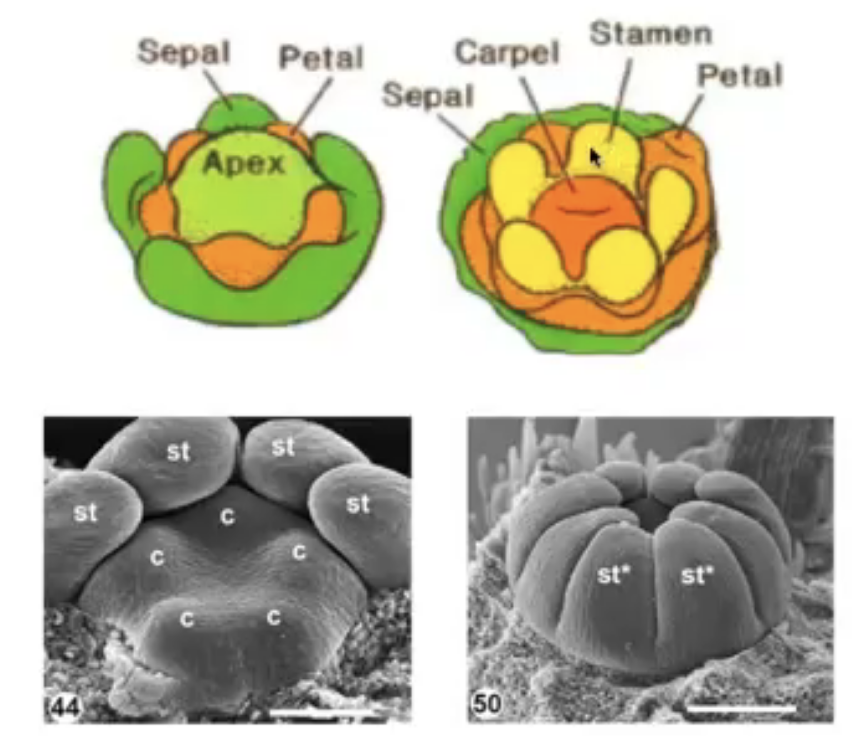
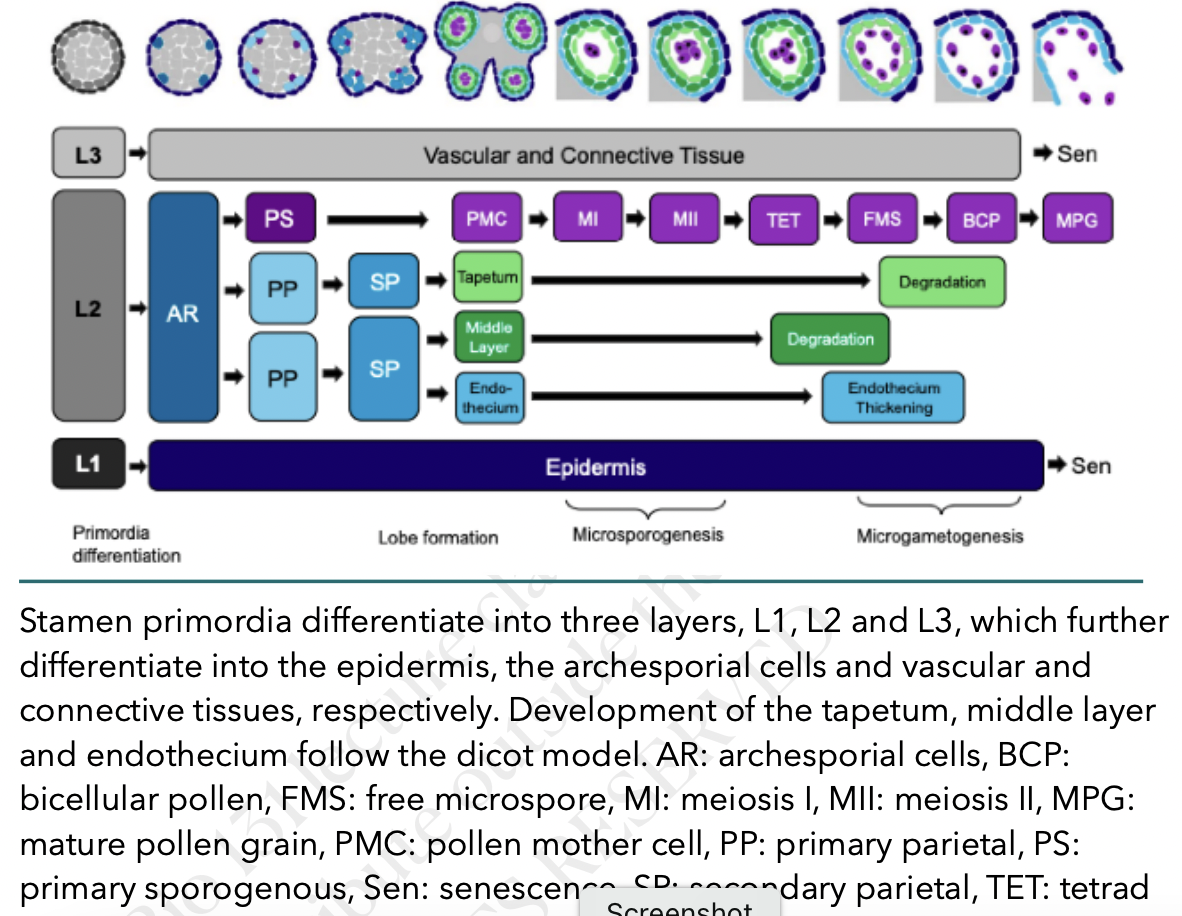
Anther development
All the structures of the flower develop from the floral meristem
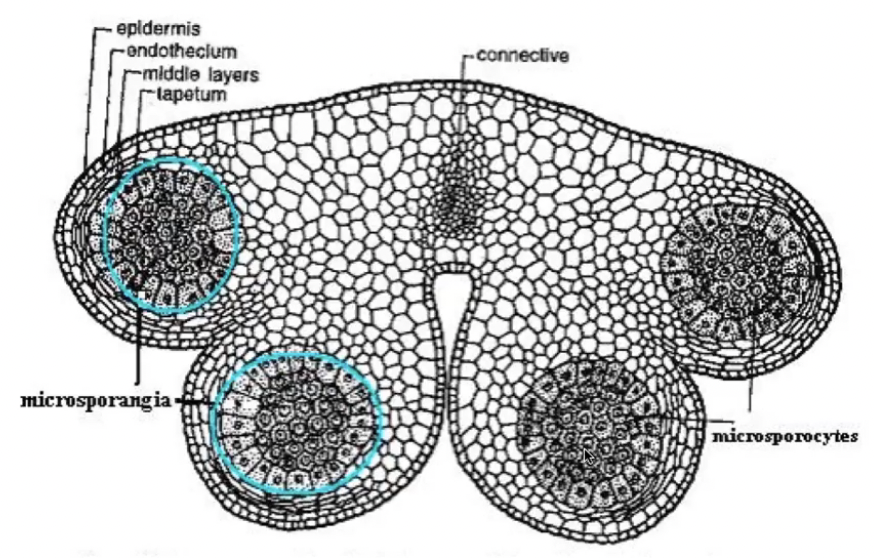
Anther primordium is seen as an oval structure in xs → mass of undifferentiated and homogenous meristematic cells
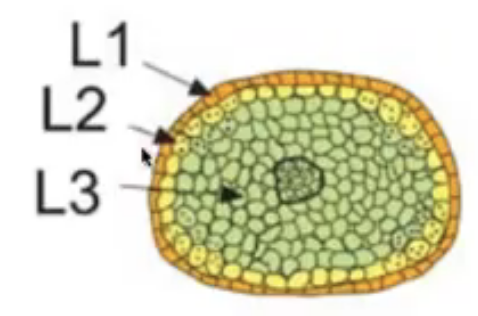
Consists of 3 germ layers
L1 → gives rise to outermost epidermis and stomium cell cluster
L2 → hypodermis and gives rise to archesporial cells
L3 → gives rise to connective tissue, vascular bundle, and circular cell cluster
Vascular tissue begins to form in the middle region
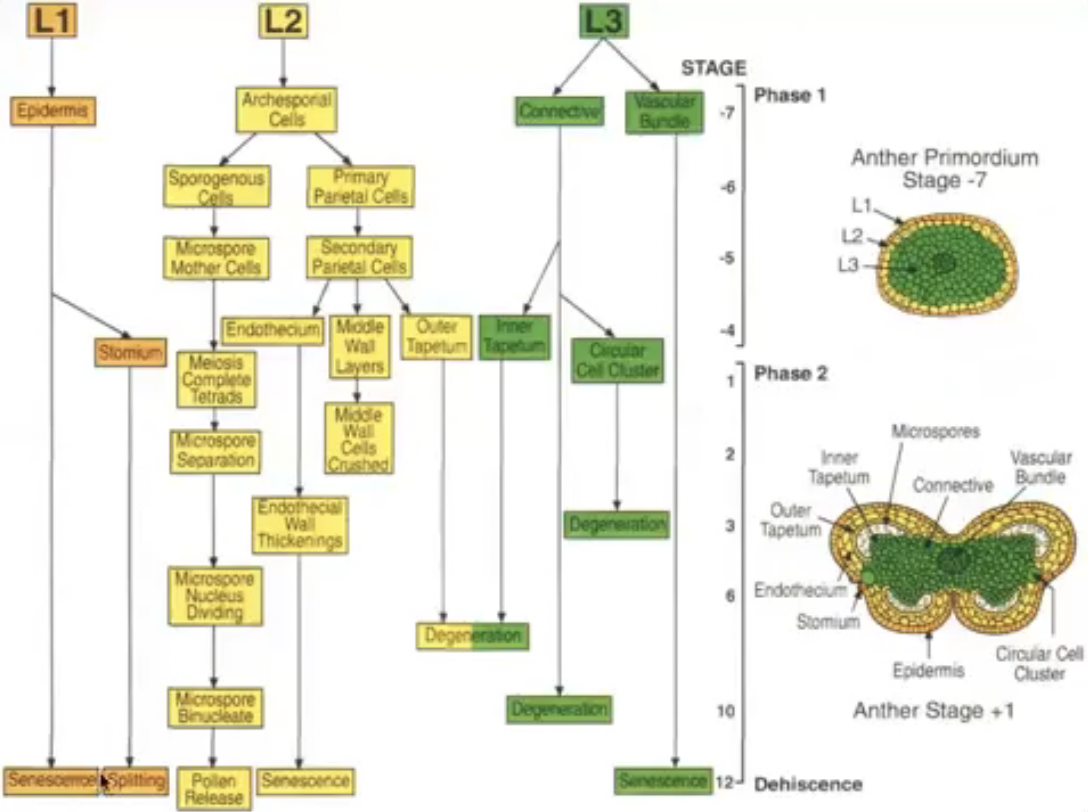
Tapetum has two origins for those species with more than one layer
Outer layer from L2
Inner layer from L3
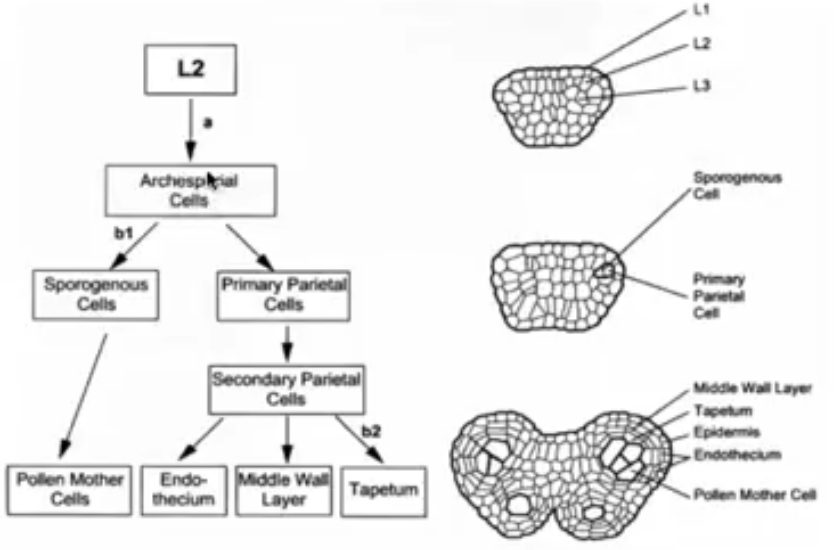
Derivatives of L2, the hypodermis
Archesporial cells formed from the periclinal division of L2 layer
Archesporial cells have large, prominent nuclei and dense cytoplasm
Periclinal division of archesporial cells form 2 cell layers
Inner layer → consists of primary sporogenous cells that become microsporocytes
Outer layer → consists of primary parietal cells that become secondary parietal cells that divide to become non-reproductive anther wall layers (endothecium, middle layer, and tapetum)
All wall layers except epidermis from L2 cells
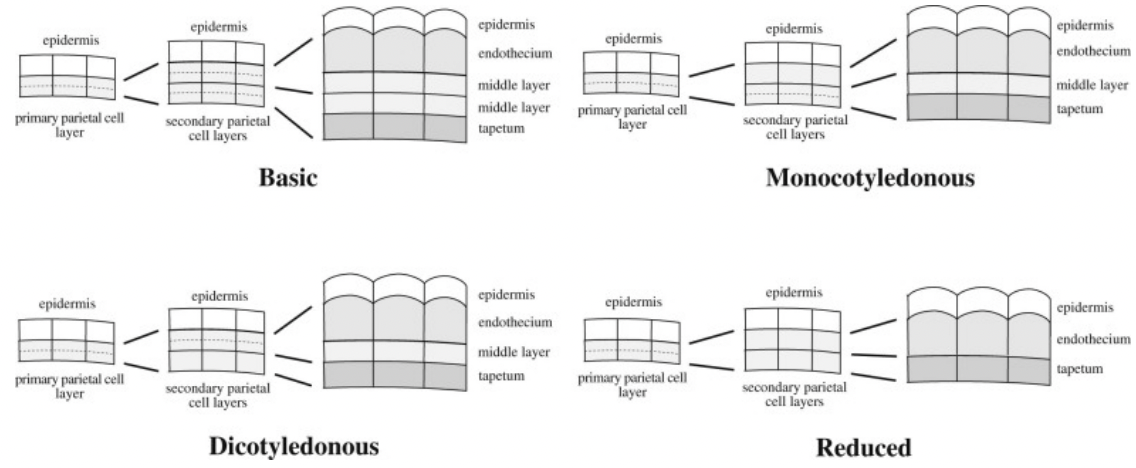
Derivatives of primary parietal cells
Divide tangentially (periclinal) to give rise to two layers of secondary parietal cells
Basic → both secondary parietal cell layers divide to yield 2 middle layers
Dicotyledonous → only outer secondary parietal cell layer divides to yield endothecium and single middle layer
Monocotyledonous → only inner secondary parietal cell layer divides to yield tapetum and single middle layer
Reduced → secondary parietal cells do not divide and develop into endothecium and tapetum respectively
Sporangium initiation is restricted to the four corners of the developing anther

Refer to days of flower opening where 0 represents day 0
All the days after day 0 is marked positive meaning it has been a few days since the flower opened and same for the negative numbers
The numbers correpond to a specific event
Example: stage 7 corresponds to day 0 which is whe nthe flower will open
Summary of stages of flower development in Arabidopsis

Stages in the development of the anther
Position dependent developmental pathway
Rare divisions enable L2 cells to penetrate L1 causing these cells to follow an epidermal pathway
Cells within each germ layer are conditionally specified and that the ultimate fate of these cells depends upon their position
Two phases of stamen development
Phase I → sporogenic cells engage in microsporogenesis while nonsporogenic cells form epidermis, tapetum, etc…
Growth, histodifferentiation, meiosis
Phase II → anther enlarges and filament elongates, pollen grains form, dehisce, and released
Tissue degeneration, dehiscence, pollen release
Schematic view of development of anther layers and microsporogenesis in Arabidopsis
Tapetum
Usually only a single layer that circumscribes the locule
Special features
ER-golgi complex
Numerous secretory vesicles toward side facing locule
Small vacuoles containing lipophilic substances
Loosening of cellulose structure or total dissolution of cells in certain areas which permit transfer of materials into anther locule
Exhibit endoreduplication and polyploidy → though cells are functionally differentiated and nondividing, they retain their mitotic potential resulting to multinucleate cells (no cytokinesis)
Attain max development when microspores are in tetrad stage
After tetrad stage, tapetal cells degenerate
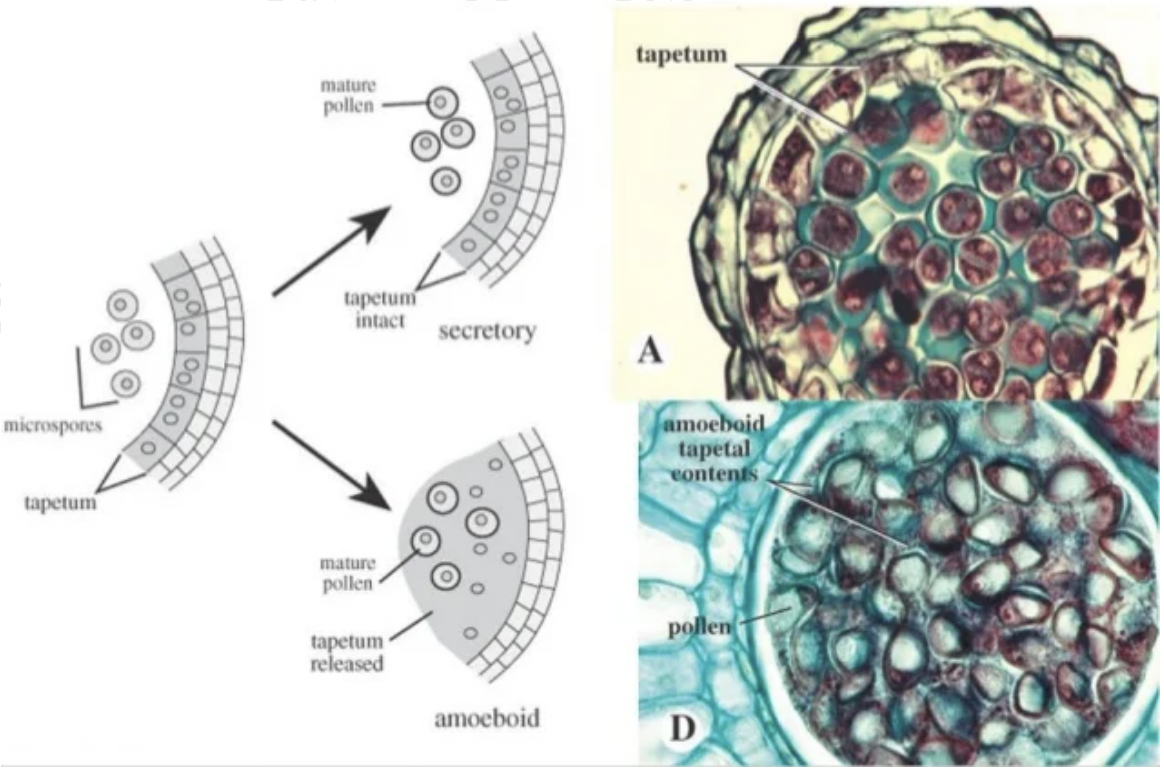
Two types of tapetum based on morphological changes during degeneration involving the remodelling of protoplasm and migration into locule
Amoeboid or periplasmodial tapetum → protoplasts lacking cell walls enlarge and fuse with one another and move into locule to surround developing pollen grains
Glandular or secretory tapetum → protoplast is stationary after cell walls lyse. Protoplasts break down and are resorbed
Substances produced by tapetum
Callase → digests the tetrad wall (made of callose) to release individual microspores
Pollenkitt → compounds imparting stickiness to the pollen grains usually composed of lipoidal compounds, carotenoids, and flavonoids
Tryphine → mixture of hydrophilic substances derived from tapetal cell debris that serves as the main encrustation of the pollen wall and aids in the adhesion of pollen grains to stigma
Anther dehiscence involves programmed destruction of specific cell types
Phase II in anther development is marked by a change from growth to degeneration of supporting tissue then dehiscence
Dehiscence begins after tetrad formation
Involves the stomium, endothecium, and circular cell cluster
Series of events:
Formation of fibrous band thickenings on endothecial cell wall
Degeneration of circular cell cluster and merging of the two pollen sacs in each theca into a single locule
Breakdown of the tapetum and connective
Rupture of anther at stomium and pollen release
Dehiscence requires gene activation of RNases, proteases, and cellulases
Onset of dehiscence results in transcriptional activation of genes that are inactive during Phase I of anther development
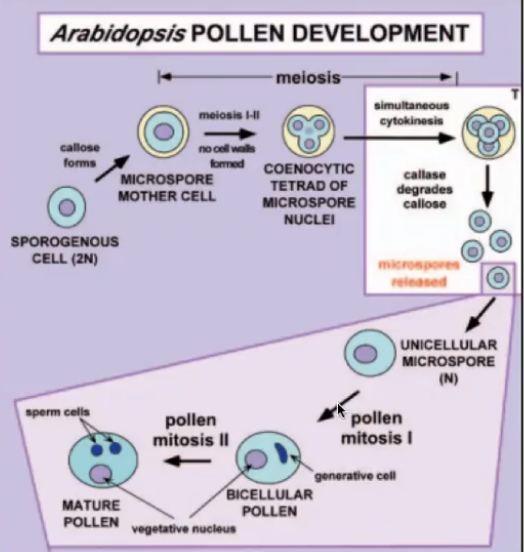
Pollen Development
3 stages
Microsporogenesis → differentiation of the sporogenous cells and meiosis
Occurs in microsporangia
Microsporocytes occur as a large mass
Callose separates microsporocytes from diploid tissue because will later on have haploid cells
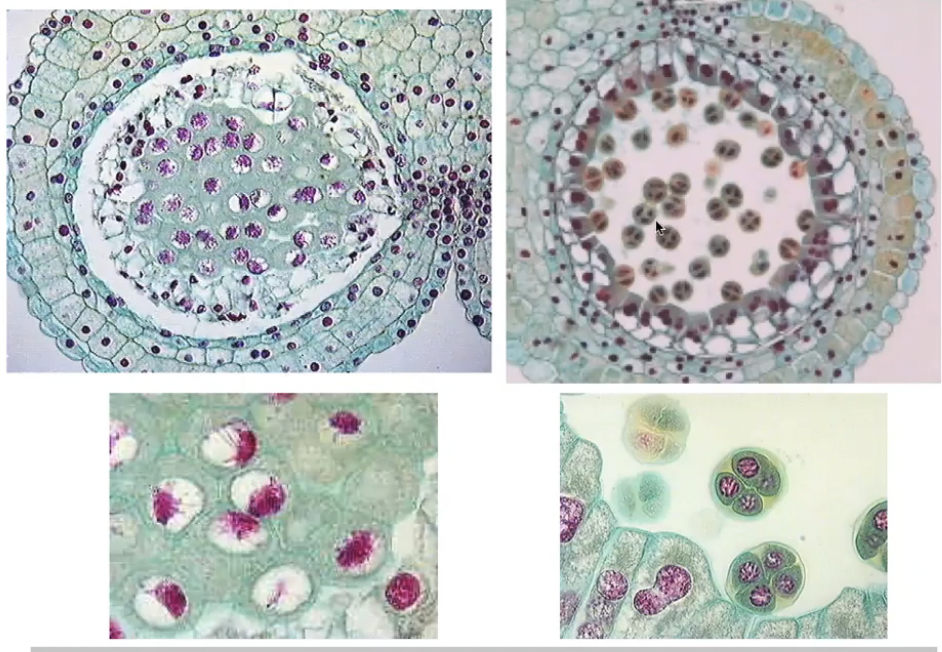
Post-meiotic development of microspores
Microspore mitosis
Sporogenous cells undergo 2-3 mitosis divisions to form microsporocytes within locules
Microsporocytes connected with each other and with tapetum via plasmodesmata
As microsporocytes enter meiosis, the plasmodesmatal bridges break and connections between microsporocytes are replaced with wide cytoplasmic channels
Microsporocytes are surrounded by primary cell wall composed of cellulose, hemicellulose, and pectin
Microsporocytes have a secondary cell wall composed of callose that acts as a protective seal
They undergo meiosis to form tetrads of microspores while encased in callose wall
As microspore matures, primary cell wall and callose layer degrade to release microspores into locules
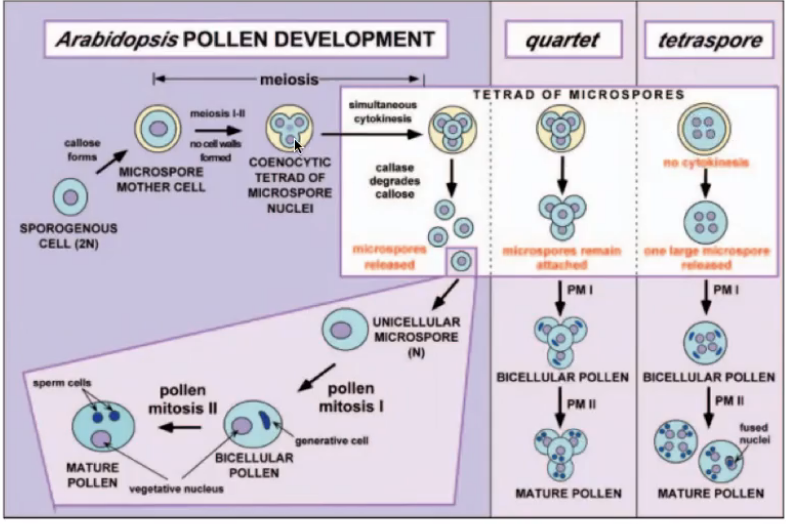
QRT mutants
QUARTET (QRT) genes are required for pollen separation
qrt mutants have four products of microsporogenesis fused and the pollen grains released as tetrads
QRT1, QRT2, QRT3 genes are required for proper degradation of dividing microsporocyte primary cell walls and subsequent separation of microspores resulting in the release of single pollen grains
qrt1 mutant degrades the callose layer but the primary sporocyte cell wall remains partially intact following meiosis
qrt2 mutant retains patchy callose around the microspore
qrt3 mutant fails to degrade the pectic polysaccharides of the walls which mechanically constrains the developing pollen grains leading to fusion of developing walls
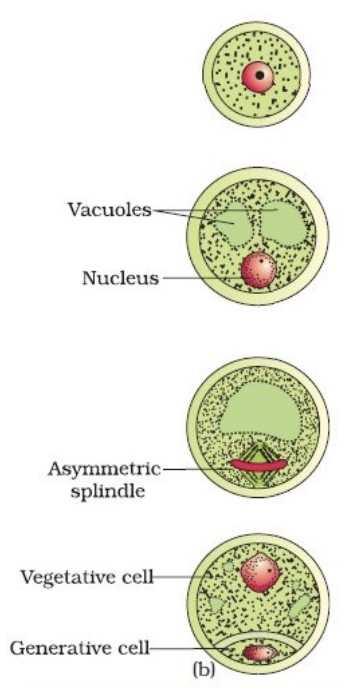
Microgametogenesis: Pollen Mitosis I and II
Events that lead to development of microspores into microgametophytes
Expansion of microspores with formation of large vauole
Displacement of microspore nucleus to an eccentric position against microspore wall
Nucleus undergoes pollen mitosis I resulting in formation of 2 unequal cells
Large vegetative cell with haploid nucleus
Small generative cell with haploid nucleus that lacks mitochondria and chloroplasts because cytoplasm was partitioned unequally (basis of maternal inheritance of chloroplast and mitochondrial genomes)
Generative cell detaches from the pollen grain wall and engulfed by vegetative cell forming a cell within a cell structure
Generative cell undergoes pollen mitosis II to form two sperm cells enclosed within the vegetative cell cytoplasm either before pollen is shed or within the pollen tube
Before pollen is shed → tricellular/trinucleate pollen
Within pollen tube → bicellular/binucleate pollen
Asymmetrical cell division is dependent on microtubules
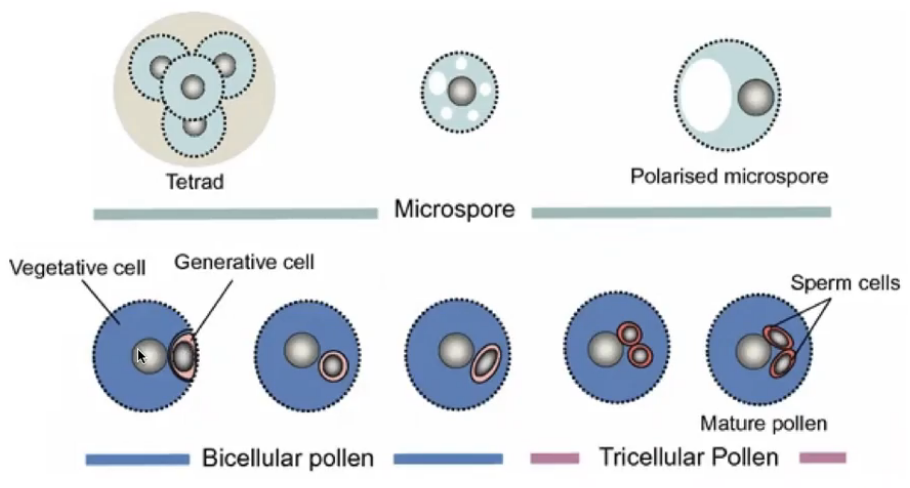
The pollen wall
Outer exine → sporopollenin found to be resistant to both physical and chemical decay
Does not develop over certain regions which define the positions of the pollen apertures or germ pores
Pollen aperture show wide variation
Exine sculpturing important in attachment to insect pollinators and adhesion to stigma
Inner intine → pectocellulosic (pectin + cellulose)
May also consist of hemicellulose and callose
Pollen wall synthesis
Exine → developed through contribution of microsporocyte cytoplasm and tapetum which play an important role in producing sporopollenin
Under sporophytic control
Intine → under the control of the microspore cytoplasm and involves gametophytic gene expression from microspore nucleus
Hormonal regulation of pollen development and dehiscence
Gibberellins → associated with early filament elongation and tapetum development
Jasmonic acid → linked to later stages of pollen maturation, filament extension, and anther dehiscence
Acts by controlling water transport in anther
Auxin → important role in early and late pollen development by regulating entry into the cell cycle, controlling dehiscence, and controlling stamen filament growth
Principal source is local synthesis
yuc2yuc6 auxin biosynthesis double mutant has no pollen grain formation, no stamen elongation, and male flowers are sterile
Auxin receptor mutants exhibited premature anther development with early endothecium thickening, premature pollen mitotic divisions, stomium splitting, dehiscence
Mutants in stamen and pollen development
Three broad categories of male sterility in angiosperms
Structural male sterility due to morphological anomalies in stamens
Functional male sterility where viable pollen is produced but it is incapable of affecting fertilization
Sporogenous male sterility where pollen development is interrupted anywhere from the pre-meiotic formation of microsporocytes to the second mitotic division
Male sterile mutants provide a bsis for hybrid seed production
Possible: male sterile but female fertile → cannot self fertilize but can be fertilized using pollen from related plants to produce hybrids
Male sterility may result from mitochondrial mutations or nuclear mutations
Genes that cause male sterility
Maize mutant antherless (at) has normal filaments but lacks anthers
Gibberellic acid0deficient maize mutants d2, d3, d5 form small anthers and fail to produce pollen
Tomato stamenless-2 mutants have short stamens and nonfunctional microspores
Arabidopsis male sterile (ms1) mutants do not produce viable pollen, but are otherwise phenotypically normal. Degeneration of pollen occurs soon after microspore release from the tetrads, at which time the tapetum also appears abnormally vacuolated
Ovule Development and Embryo Sac Formation
Structure of the Ovule
Carpel → female reproductive organ
Pistil or gynoecium → collective term for all the carpels of a flower
Central most whorl of a flower
Carpel vs pistil
If there is only 1 carpel, that carpel serves as the pistil
Separated multiple carpes are equivalent to 3 pistils
Multiple carpels that are fused equals 1 pistil

Functional parts of the carpel at anthesis
Ovary → basally located
Ovules → found in the ovary
Stigma → terminally located where pollen grains land and germinate
Style → connects the stigma and ovary

Functional parts of the ovary
Placenta → specialized ridge in the ovary where ovules are attached
Locule → space where ovules are found which can hold just one or many ovules
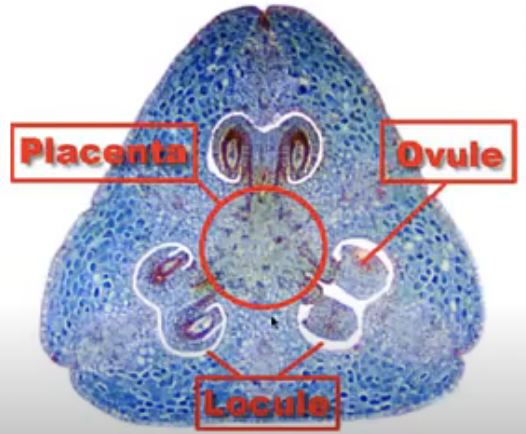
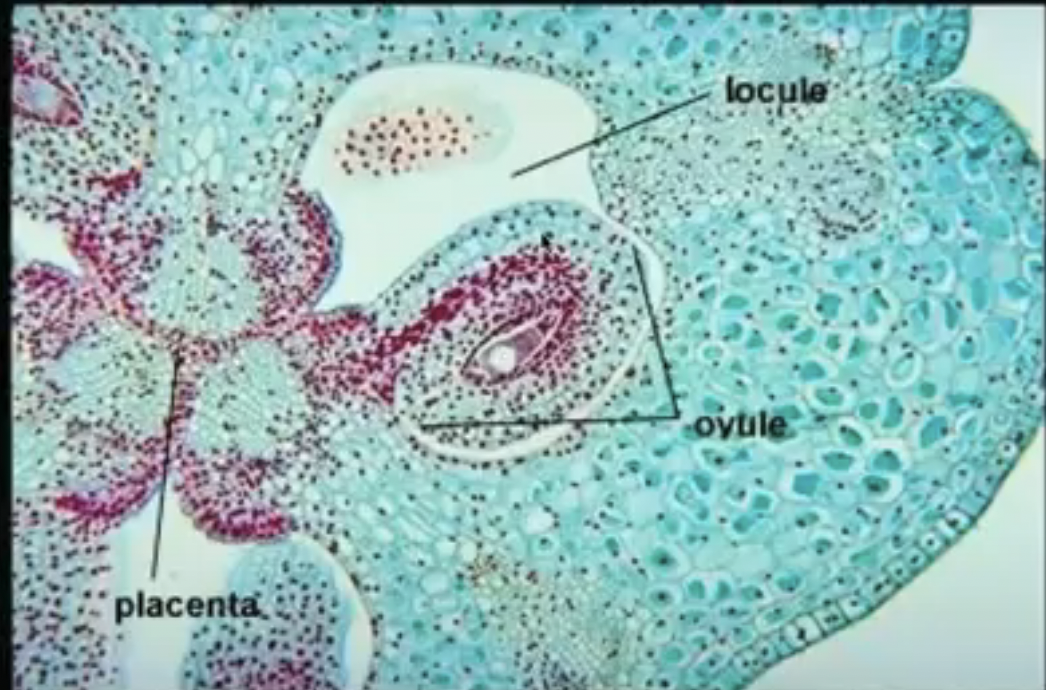
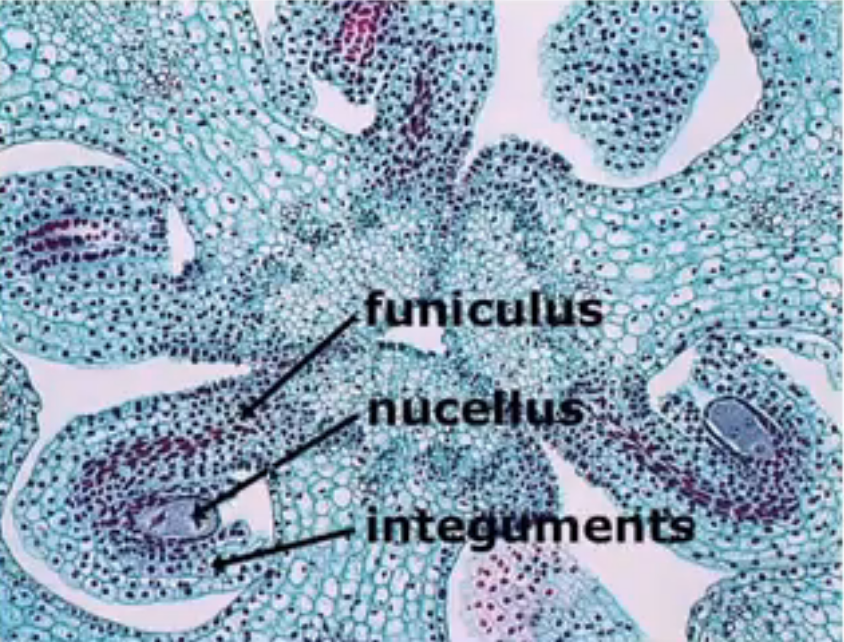
Funiculus → stalk where the mature ovule is raised to attach to the placenta
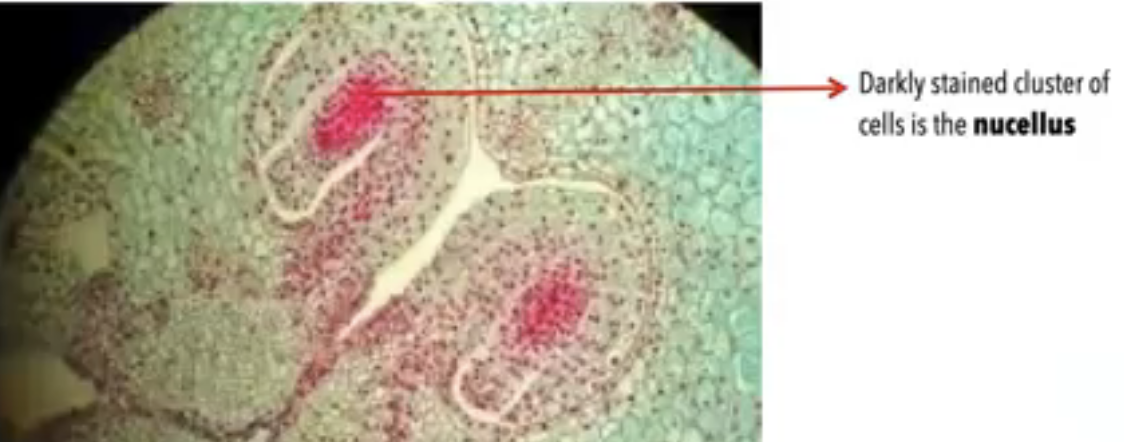
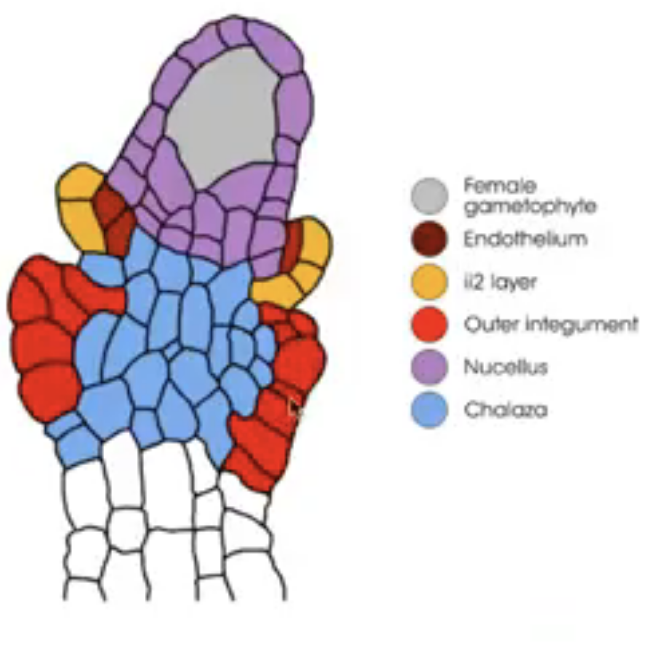
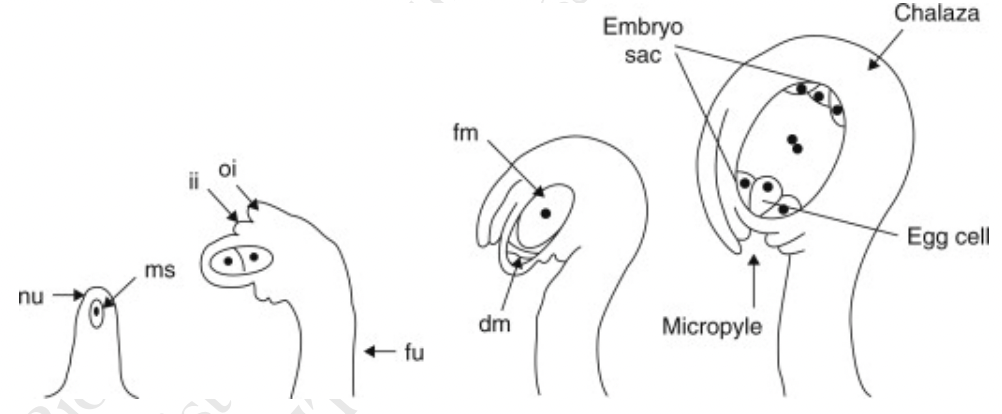
Nucellus → mass of homogenous cells in the mature ovule where megasporogenesis occurs (considered as the megasporangium)
Integuments → one or two multilayered covering outer to the nucleus
Micropyle → small opening along the integuments at the free end of the ovule
Chalaza → poorly defined region at the opposite pole of the ovule where the nucellus, integumants, and funiculus meet
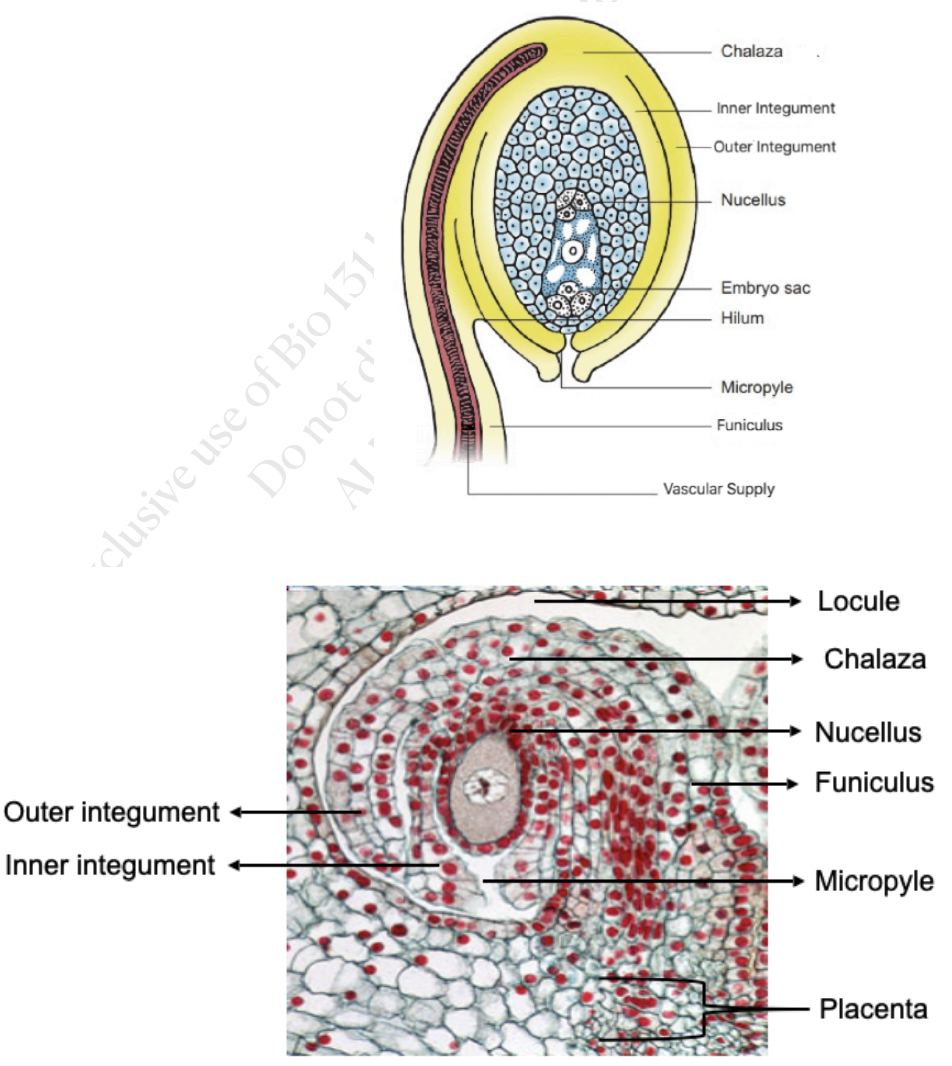
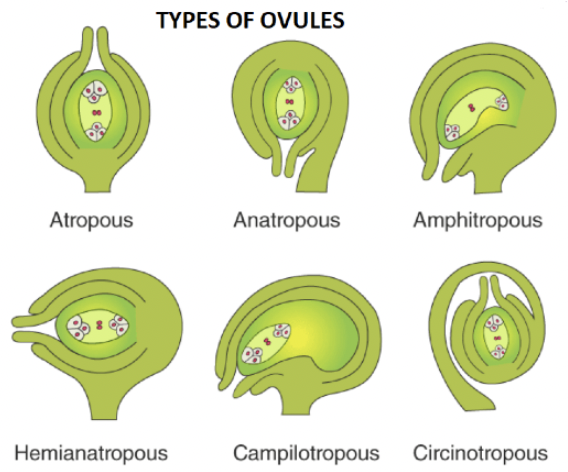
Degree of curvature of the ovule adds to the variation in its external morphology
Differential growth rates result in the curving or bending of ovules
Ovule Determination and Development
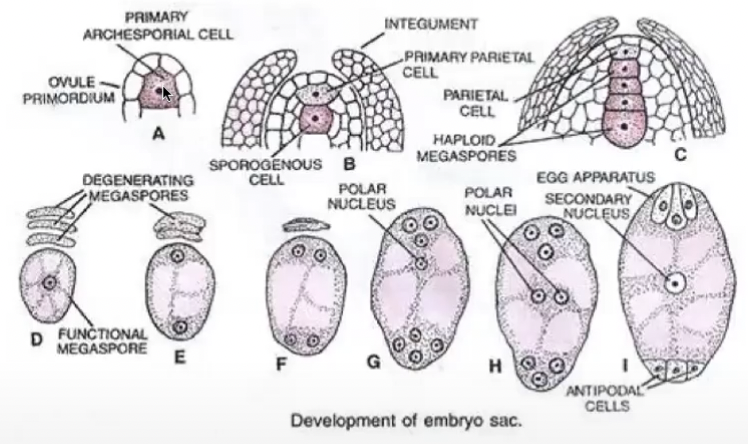
Ovule primordium is initiated by periclinal divisions from the sub-epidermal or hypodermal tissue of the placenta
L1 → outermost layer
L2 → sub-epidermal (or hypodermal) layer
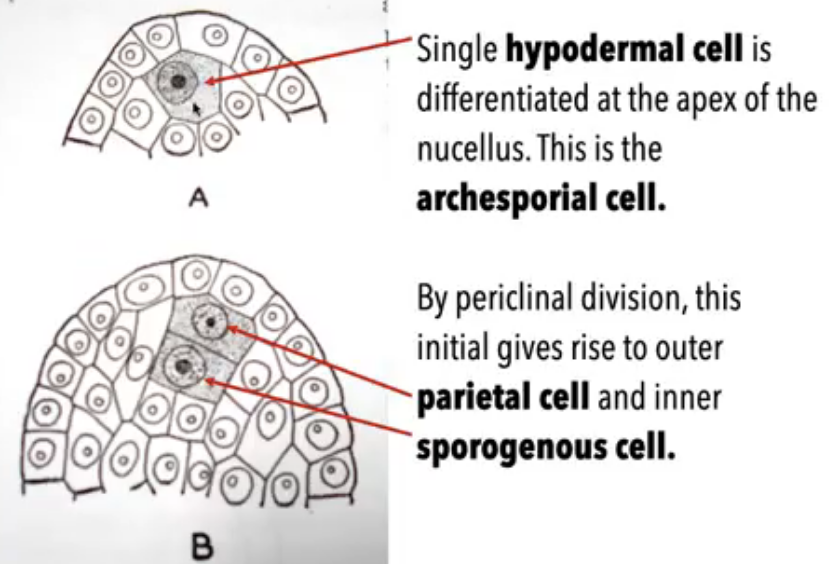
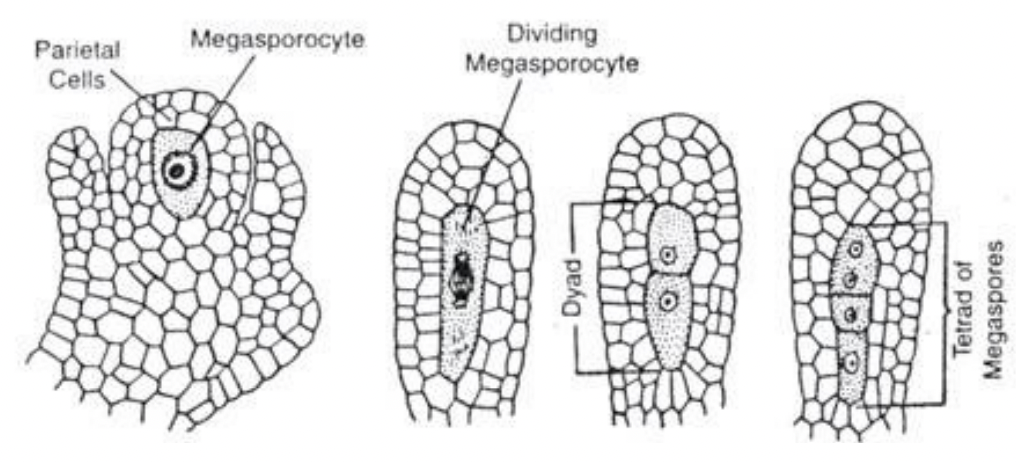
Single hypodermal cell enlarges to become the archesporial cell
Archesporial cell divides periclinally and gives rise to an outer parietal cell and inner sporogenous cell
Sporogenous cell functions as megasporocyte
Primary parietal layer divides anticlinally and periclinally to form parietal tissue
In some species, archesporial cells and primary parietal cells do not divide anymore.
L3 → innermost cells
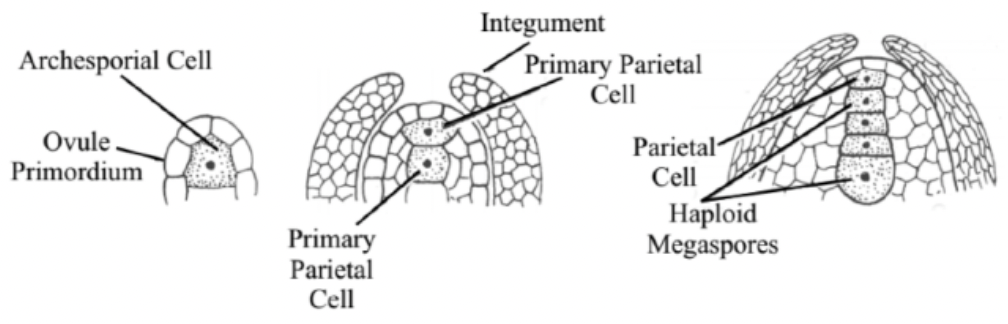
Relatively homogenous mass of cells of primordium will be organized into 2 different regions
Funiculus
Chalaza
Nucellus
Arabidopsis → archesporial cell developing in the hypodermis becomes the megasporocyte (no periclinal division)
Ovule is polyclonal → derived from more than one cell layer
Megasporocyte derived from L2
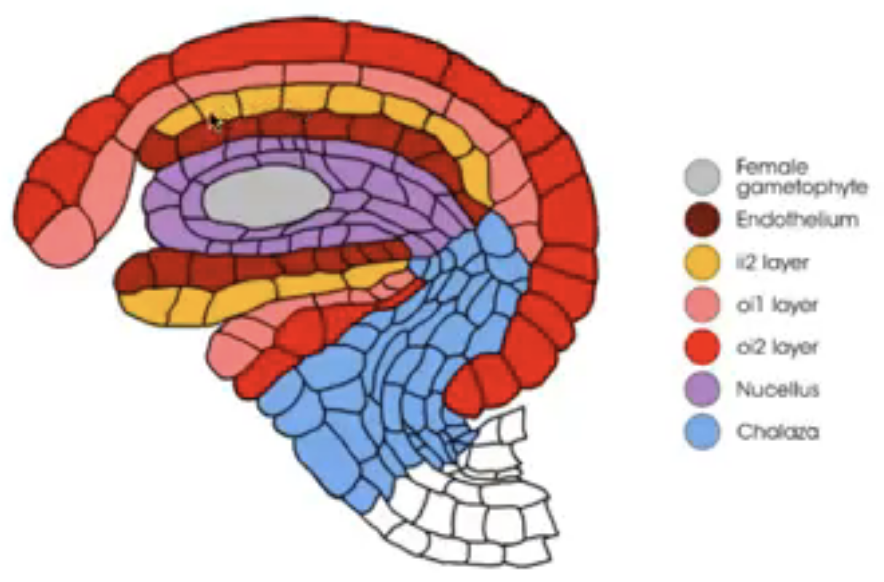
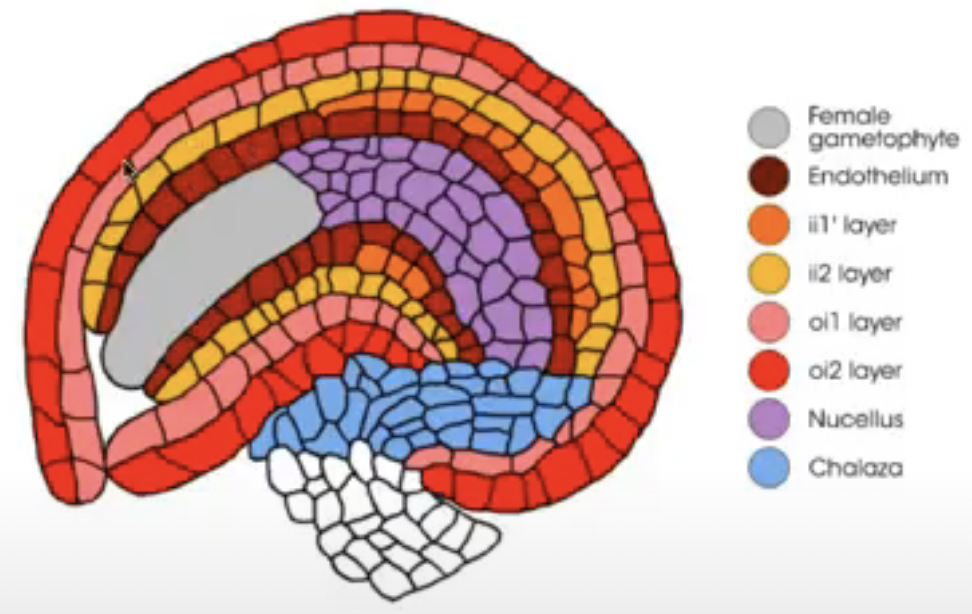
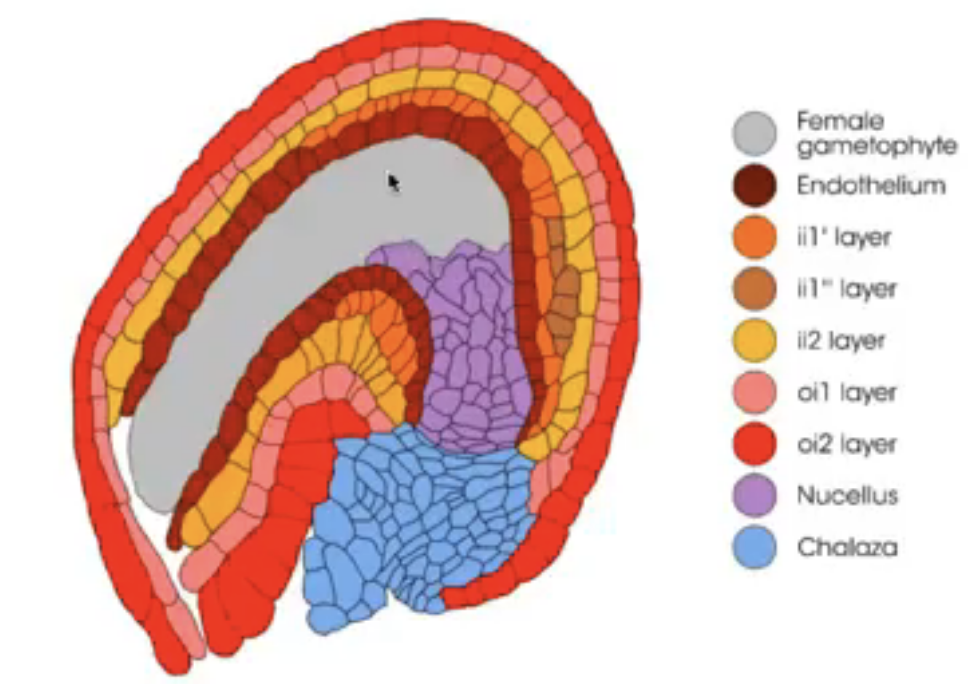
Integuments arise from the tissue surrounding the nucellus (i.e. chalaza)
Dicots and monocots have 2 integuments
Inner integument from L1
Outer integument from L1 and L2
bell mutant | bel1 | Ovule lacks an outer integument |
aberrant testa shape mutant | ats | No clear distinction between inner and outer integument |
inner no outer | ino | Outer integument does not proceed beyond its initial state so ovule only has inner integument |
aintegumenta and huellenlos | ant and hll | Extreme types wherein no integuments are formed at all |
Before embryo sac matures, nucellus degenerates in many species, leaving the embryo sac in direct contact with the inner integument cell layers that may differentiate into endothelium
Endothelium → similar to tapetum of anther and may function in the production and secretion of substances for the developing reproductive cells
During enlargement, ovule bends so the micropyle lies close to the placenta along which the pollen tubes grow
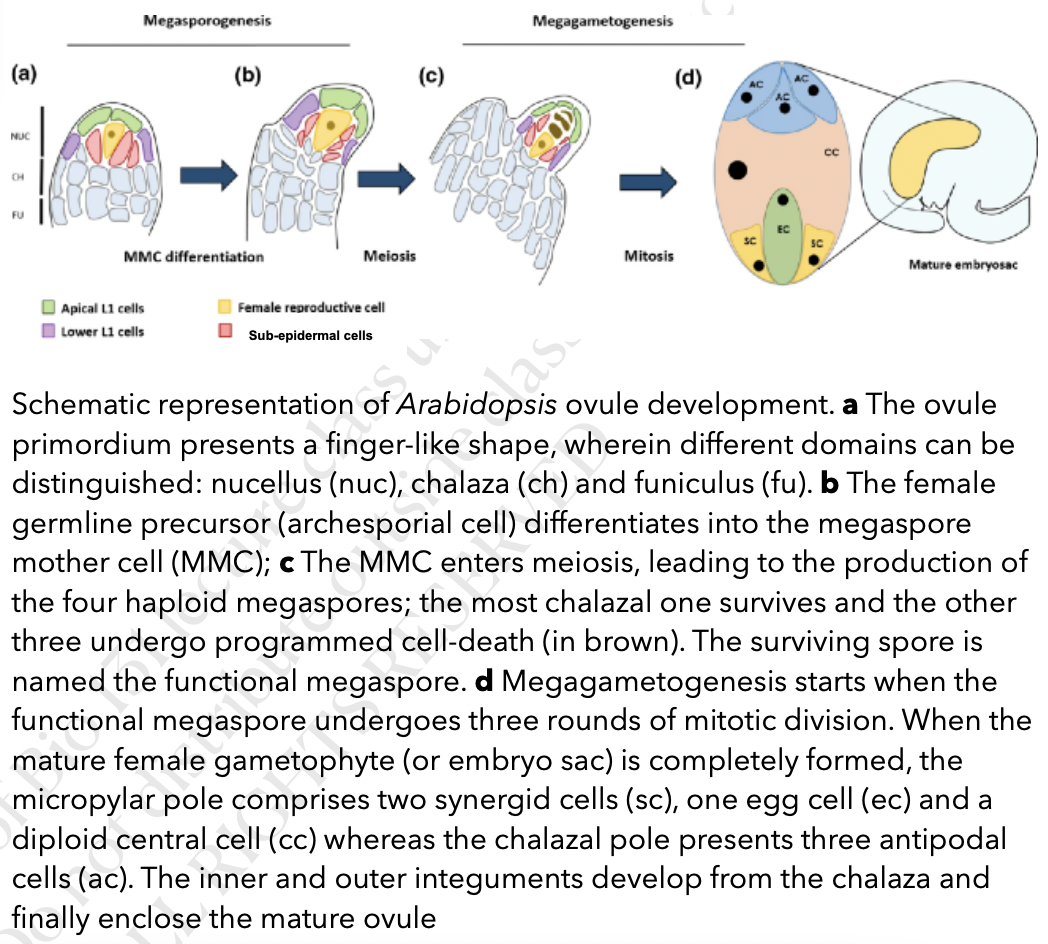
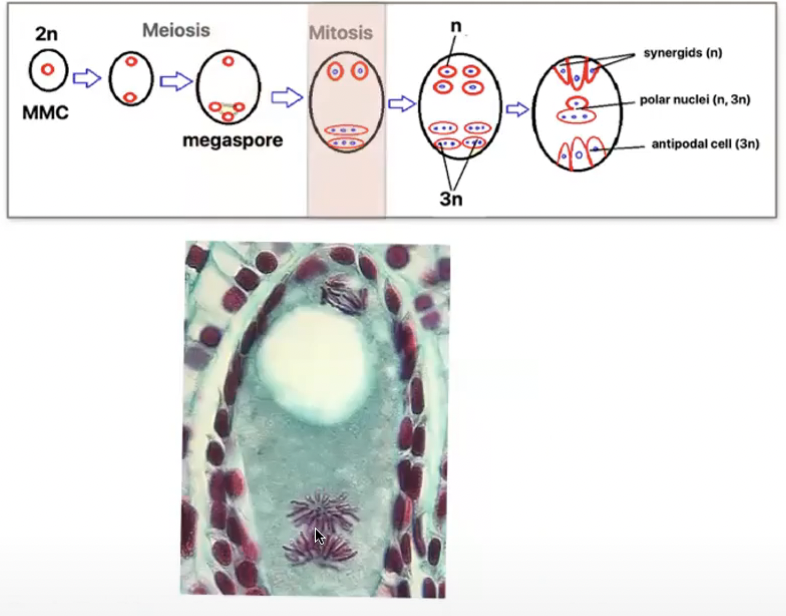
Megasporogenesis
Where megasporocyte (2n) undergoes meiosis I to form a dyad and then meiosis II to form 4 haploid megaspores arranged in linear tetrad (separated by cell walls)
Megaspore nearest the chalaza remains functional out of the tetrad and other 3 degenerates
Functional megaspore produces female gametophyte via mitosis
Megagametogenesis (no cytokinesis) → Monosporic or polygonum type
Megaspore grows and gets nutrients from nucellus
Megaspore nucleus divides mitotically to form 2 nuclei
Each nuclei moves towards the opposite pole and once again divide twice mitotically
Each pole now has 4 nuclei (8 in total)
Embryo sac is not coenocytic during development
Callose shell surounds the entire embryo sac during development
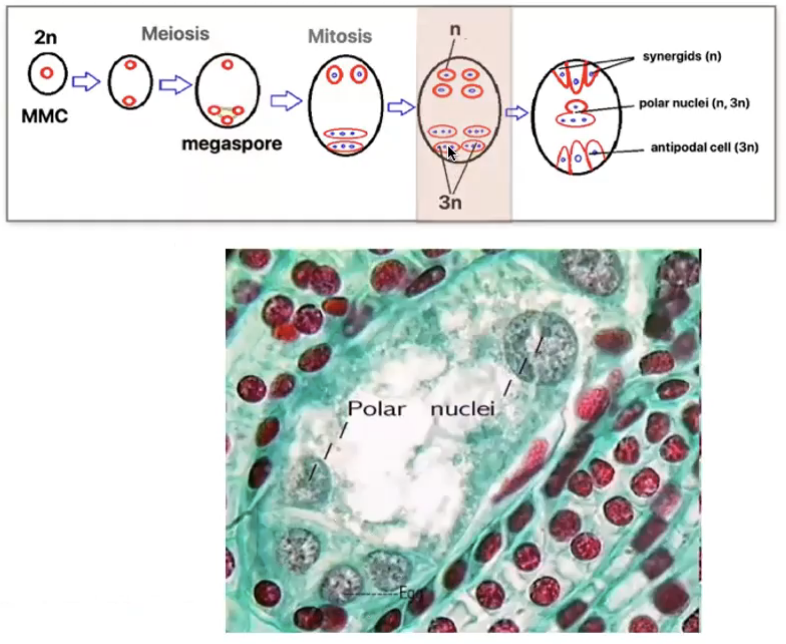
1 nuclei at each pole migrates towards center becoming the polar nuclei of central cell
3 remaining nuclei at each pole is surrounded by cytoplasm and membranes to form cells via cytokinesis
Movement of nuclei due to remnants of spindle fibers
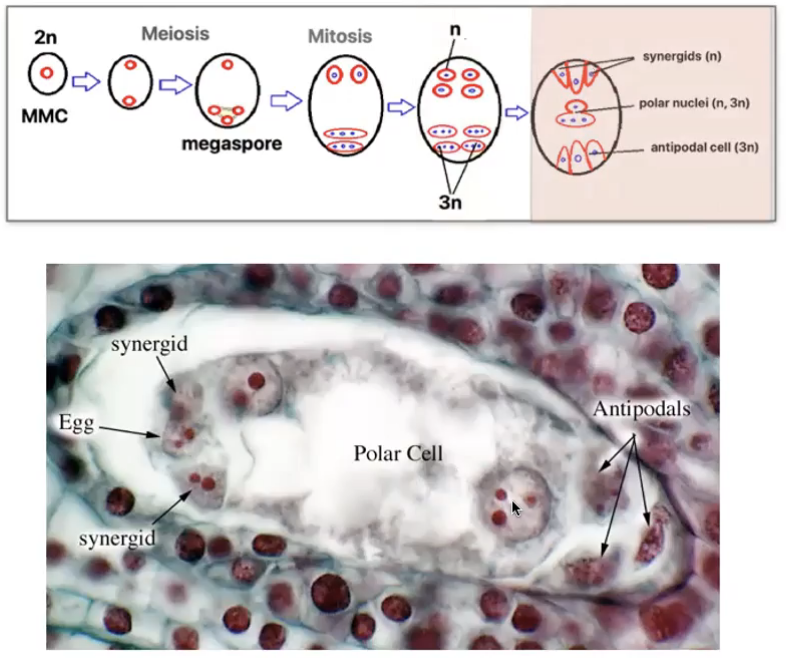
3 cells towards micropyle → egg-apparatus
Larger cell → Egg cell
2 smaller cells → synergids
3 cells towards the chalaza → antipodal cells
8 nucleated and 7-celled structure is the female gametophyte or mature embro sac
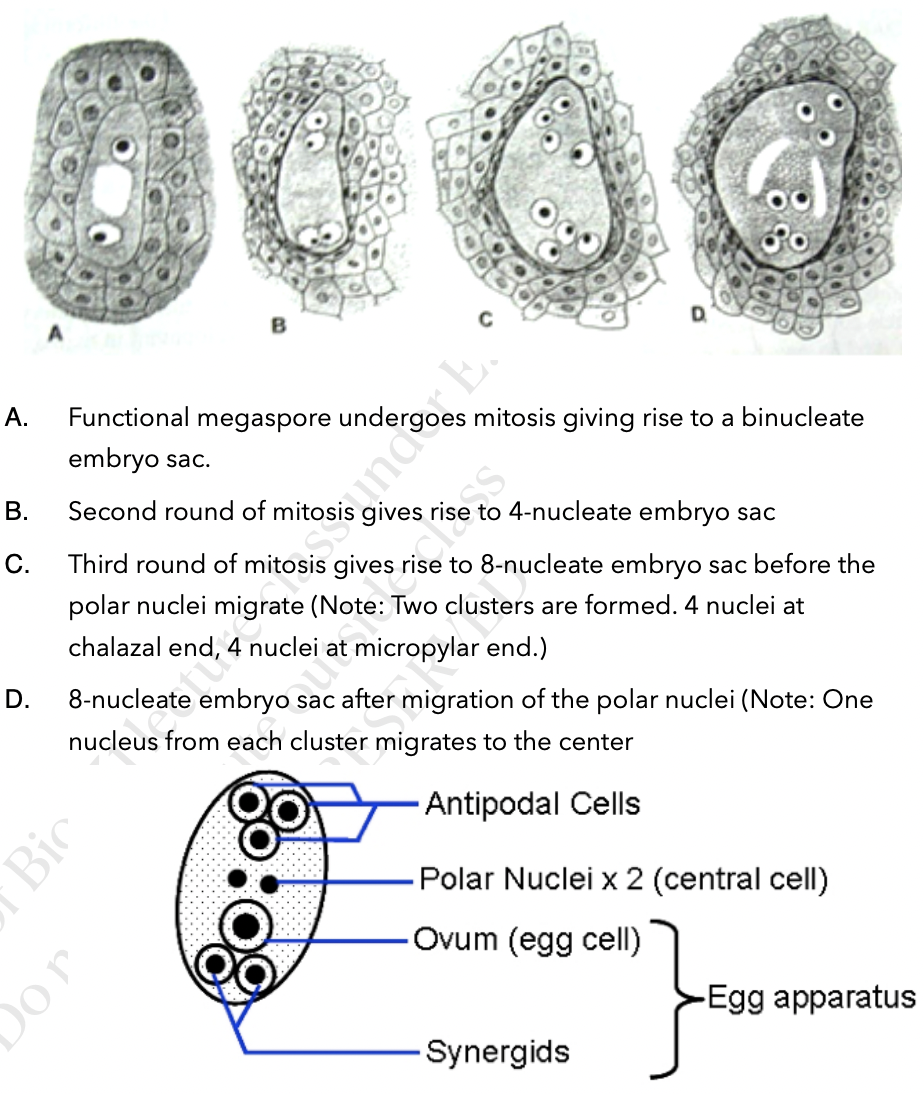
Polar nuclei may or may not fuse before fertilization
If they fuse → polar nuclei form a secondary nucleus (2n)
If they dont fuse → becomes a single cell with 2 nuclei (n+n)
Filiform apparatus → finger like processes produced from the outer wall of the synergids
help synergids absorb food from the nucellus and transfer to the embryo sac
May secrete chemicals which attract the growing pollen tube
Characteristics of the Cells of the Embryo Sac
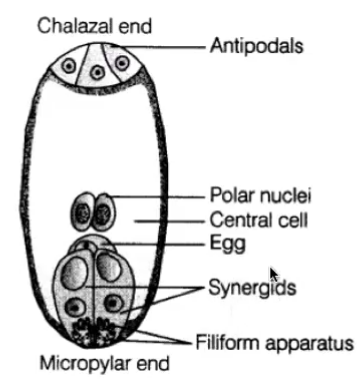
Egg cell (n)
Highly vacuolated
Amount of cytoplasm is limited and is spread as a thin layer surrounding the vacuole
Cytoplasm with little ER, limited number of plastids, mitochondria, and dictyosomes, but high number of ribosomes that are randomly distributed
Cell wall does not extend over the entire cell
Strongly polarized where micropylar end has a large vauole and chalazal end has most of the cytoplasm
Antipodals (n)
Transient existence
Minimal cytoplasmic organelles
May have nuclear abnormalities like endoreplication
Polar nuclei (n)
Metabolically active
Extensive ER, numerous plastics, mitochondria, dictyosomes, and polysomes
Large quantities of starch, proteins and lipids
Synergids (n)
Limited life span, degenerate after fertilization
Probably involved in nutrition of the egg cell
Has extensive wall ingrowth at micropylar region called filiform apparatus
Produce chemicals that attract pollen tube
Variations in gametophyte development
Deviations from monosporic megagametophyte development
Number of megaspores or megaspore nuclei that participate in the formation of the embryo sac
Total number of divisions that take place during the formation of the megaspore and gametophyte
Number and arrangement of the nuclei and their ploidy level in the mature embryo sac
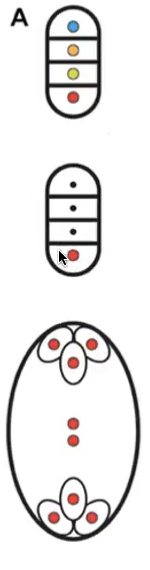
Monosporic trimitotic embryo sac
Meiosis of megaspore mother cell (2n in nucellus produces 4 megaspores (n)
3 undergo apoptosis
All 8 nuclei are genetically identical → products of mitosis of the megaspore nucleus
Polygonum type
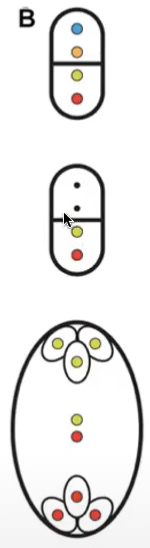
Bisporic bimitotic development
Results from failure of cytokinesis after meiosis II
2 binucleate cells are produced after megasporogenesis
Bisporis bimitotic embryo sacs → allium-type
Micropylar binucleate cell is suppressed while chalazal binucleate cell undergoes development
2 nuclei in functional megaspore contain different genetic combinations due to being products of meiosis thus the nuclei of mature embryo sac will not all be genetically identical
Only 2 mitosis divisions are involved in the formation of mature embryo sac
Tetrasporic bimitotic development
Associated with suppression of cytokinesis after both meiosis I and II
Four-nucleate megaspore
Produces a chimeric embryo sac after mitosis of 4 genetically different nuclei
2 mitotic divisions of the 4 nuclei = 16 nucleate embryo sac
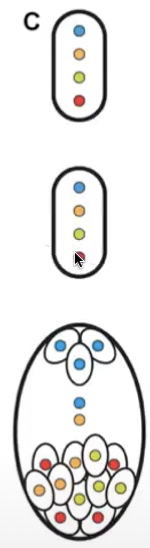
Tetrasporic bimitotic ‘Fritillaria-type’
Where three somatic spores of megaspore tetrad fuse to form a triploid nucleus
Egg cell and synergids are haploid and antipodal cells are triploid
One polar nucleus is haploid and the other is triploid
Gene regulation of ovule formation
ANT transcription factor → clear role in ovule primordia formation
Expressed in the placenta and in the integuments of the developing ovules
ant mutant plants → ovules do not develop integuments and megasporogenesis is blocked at the tetrad stage → female sterility
ant-9 mutant → number of ovules per carpel is reduced by more than half in respect to the wild type
HUELLENLOS (HLL) → encodes a mitochondrial ribosomal protein
hll mutants → ovule do not develop integuments
hll-1 and hll-3 → reduction of 10% in number of ovules and display smaller gynoecia
Double mutant hll ant → more severe at the level of primordia outgrowth
short integument 2 (sin2) mutants
Arrest in cell division in both ovule integuments
Shorter pistils bearing less ovules than wild type
Double mutant sin2 ant-5 → same with ant-5 single mutant
ANT is epistatic to SIN2 with respect to ovule development
sin2 hll-1 double mutant → stronger effect on ovule development than their single mutants
ANT plays a master role, SIN2 and HLL contribute to ovule primordia formation
Role of hormones in ovule primordium formation
Auxin
Responsible for the correct apical basal patterning of the gynoecium
Auxin gradient hypothesis supports
high levels of auxin in gynoecium apical regions control stigma and style formation
Medium levels direct ovary formation
Low levels for the gynophores at the gynoecium base
yucca1 yucca4 (yuc1 yuc4) and weak ethylene insensitive8 tryptophan aminotransferase related2 (wei8 tar2) double mutants → severe gynoecium defects lead to a pistil with a reduction or complete absence of ovules → complete sterility
Cytokinins → activate ovule primordia formation
Brassinosteroids → involved in the control of the initiation and formation of reproductive organs
BR-deficient and BR-insensitive mutants → smaller and less seeds
BR-enhanced → more seeds
Play a role in ovule initiation
Pollination and Double Fertilization
Pollination in Angiosperms
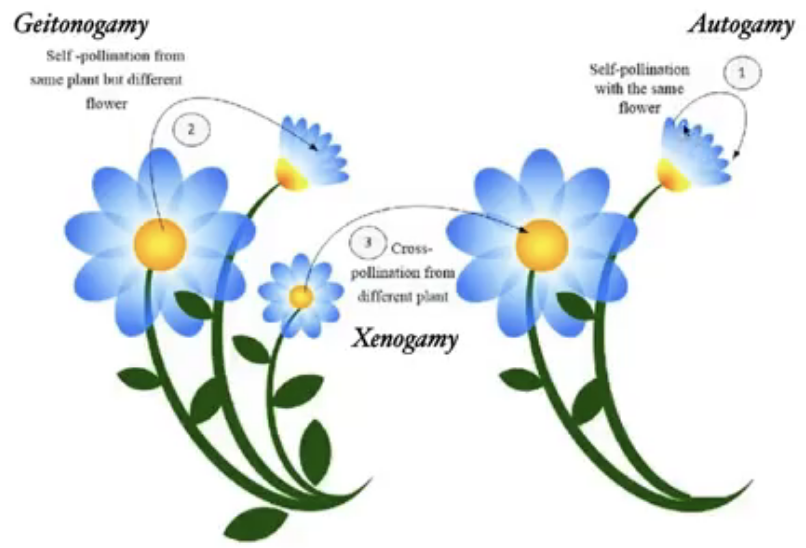
Pollination → transfer of pollen grains from anther to the stigma of the same or different flower of the same species
Self pollination → same flower or stigma of another flower in the same plant
Autogamy → transfer of pollen from anther to stigma of same flower
Possible when flower is bisexual and male and female parts mature at the same time
No need for external agents of pollination
Geitonogamy → transfer of pollen from anther to the stigma of another flower of same plant
Unisexual or bisexual flower
All flowers of same plant are genetically identical so still considered self pollination
Ecologically considered cross pollination
Needs external agents of pollination
Geitonogamy genetically similar to autogamy and functionally involves a pollinating agent in cross-pollination
Cross pollination → transfer to another plant
Xenogamy
Requires external agents
Only type that brings genetically different pollen grains to the stigma
Biotic or abiotic external agencies are okay
Wet vs Dry stigma
Wet stigma → consists of a loose aggregate of secretory cells that produce a fluid rich in glycoproteins, mucilages, and nutrients for pollen germination
Dry stigmas → do not secrete large quantities but are are highly specialized tissue that allow pollen germination
Pollen growth involves formation of the pollen tube that emerges from the pollen pore which grows down through the stigma and into the style carrying sperm to egg
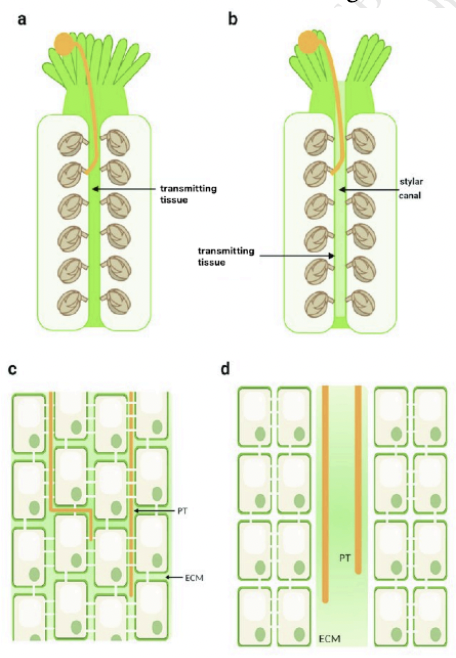
Open vs Closed styles
Open styles → central cavity whose inner epidermal surface is coated with mucopolysaccharides, lipoproteins, and glycoproteins
Serves as a nutrient medium for pollen tube
Play a role in directing growth of pollen tube
Continuous stylar canal lined with secretory epidermis
Epidermal layer of secretory cells lining a canal with extracellular matrix
Closed styles → cells are embedded in an extracellular matrix similar to the inner surface of the open style
Pollen tubes growth through the extracellular matrix, deriving both guidance and nutrition from matrix molecules
Continuous strand of transmitting tissue inside pistil
Presence of substantial intercellular spaces filled with extracellular matrix
Elongated cells connected via plasmodesmata
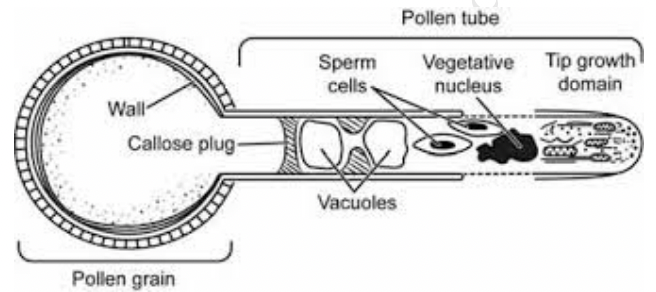
Pollen germination
Does not involve cell division
Pollen tubes are extensions of the tube cell
Tip growth
Cell wall of pollen tube has callose
Callose → synthesized by golgi apparatus and transported to the extreme tip by golgi derived vesicles
Fusion of vesicles with plasma membrane expands cell membrane of elongating tube while contents of the vesicle expand the wall
Membrane of these vesicles coated with myosin
Vesicles are transported to the tip via actin filaments
Total cytoplasmic volume does not increase as pollen tube grows
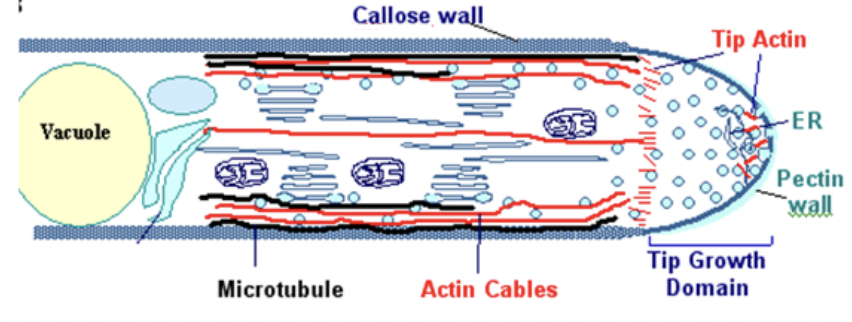
Bulk of cytoplasm is in close proximity to growting tip and continues to move with tip
Distal vacuole expands as the tube elongates which pushes cytoplasm towards tip
Elongating cell forms periodic callose cross walls or callose plugs at distal region of cytoplasm that seals off newer portions of the tube
Only terminal portion of tube has living cytoplasm
Cytoskeleton of pollen tube continually transports organelles generative nucleus and vegetative nucleus to growing tip
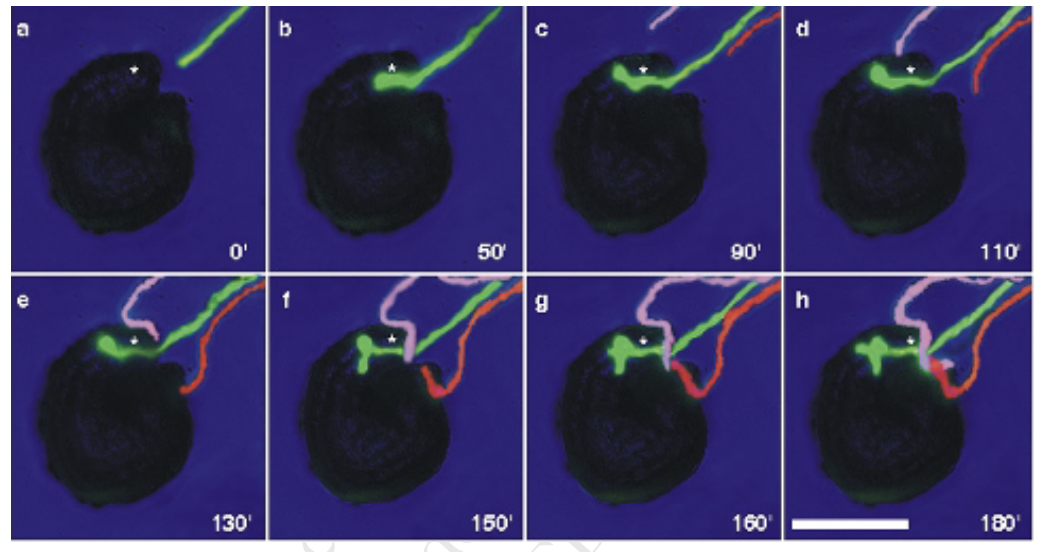
Pollen tube guidance
Chemcal attractant released by ovule prior to fertilization is developmentally regulated only occurring when the flower reaches a certain stage
Underaged ovules cannot attract pollen tubes because the structures that produce the chemical attractants are not yet there
Proposed source of chemical attractant is the synergids
This stage coincides with synergid development
Guidance signals are species specific where ovule signals of a certain plant species will not attract pollen tubes from another plant species
Only one pollen tube can gain access to each micropyle and other approaching tubes turn sharply away once a tube has entered
Chemical repellant serves as a block to polyspermy
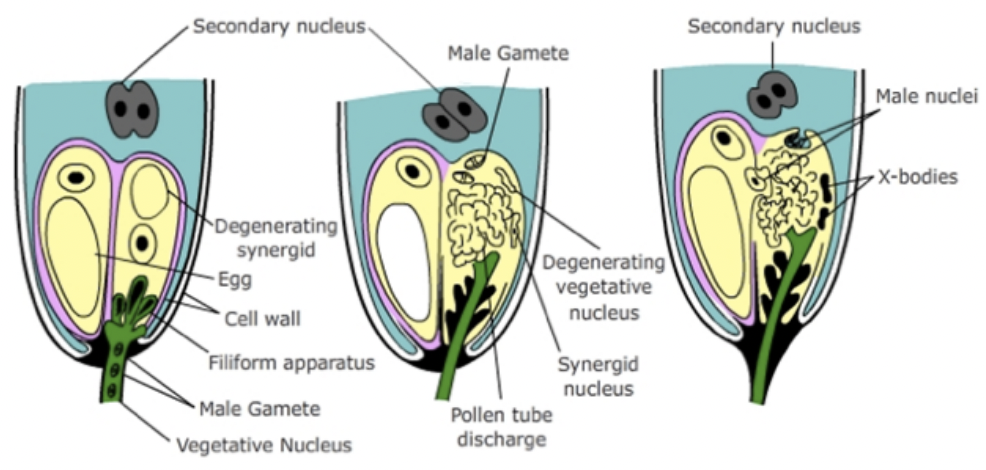
Double fertilization
Pollen tube enters one of the synergids at the base of embryo sac
Inside cytoplasm of synergid, pollen tube ruptures to release the tube nucleus and 2 sperm
1 sperm fuses with egg → zygote
1 sperm fuses with 2 polar nuclei → triploid endosperm
Embryogenesis and Endosperm Development
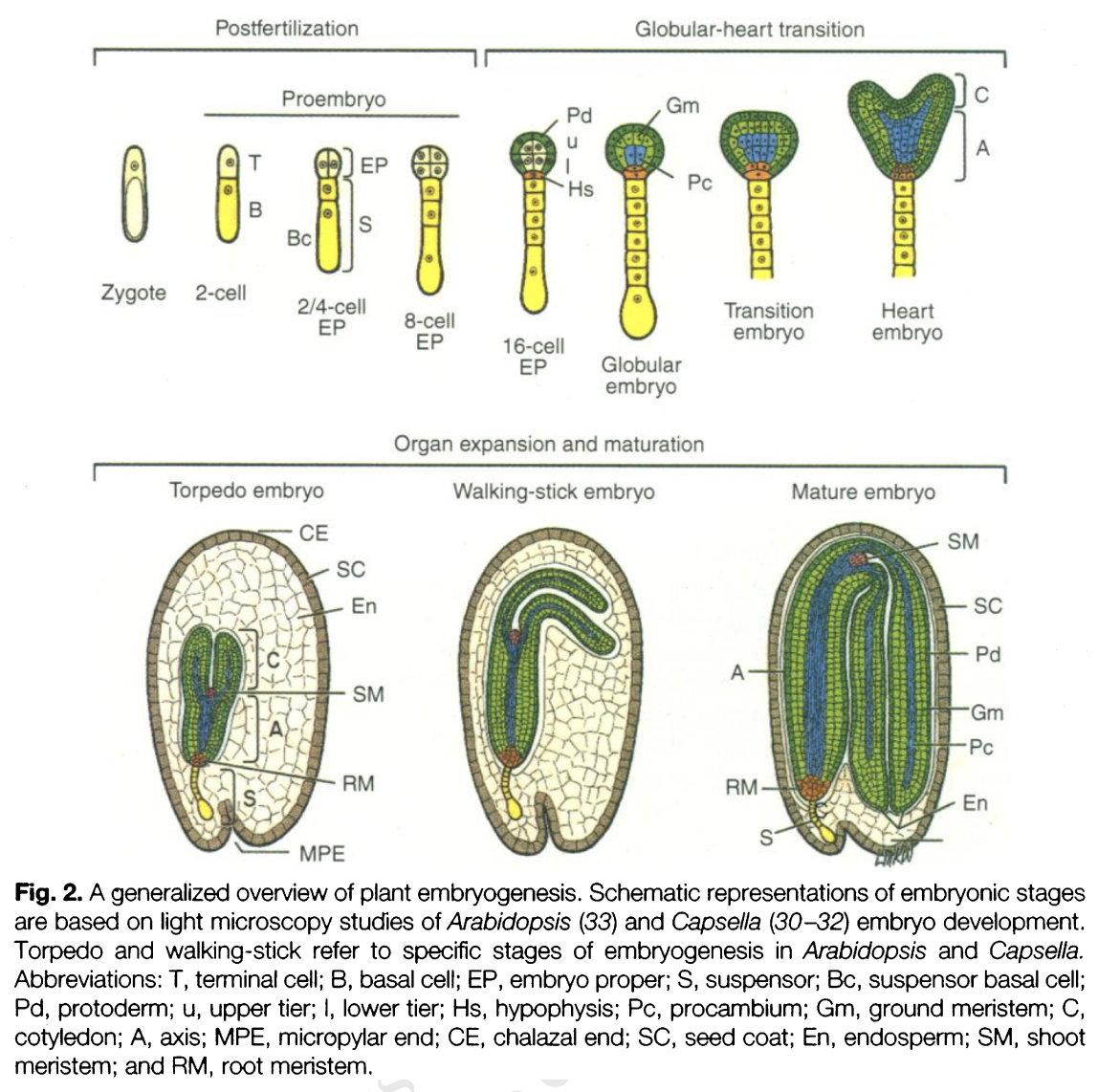
Embryogenesis
Establishes the axis of the plant, with RAM and SAM at opposite ends, and the basic pattern of tissues within axis
Accompanied by the growth and development of the endosperm
Organs only formed after seed germination → post embryonic
Angiosperm Embryogenesis
Develops at the micropylar end of the embryo sac where zygote is situated
Zygote divides to form embryo via mitosis
Division of zygote only begins when ample endosperm has been formed
Eudicot embryogenesis
Egg cell → polarized structure with
large central vacuole asymmetrically positioned micropylar end of the egg
Nucleaus and cytoplasm opposite end
Zygote undergoes unequal transverse division to form 2 cells
Basal cell → larger cell at micropylar end that inherits the vacuole
Apical cell → smaller cell at chalaza that inherits most of cytoplasm
Basal cell divides transversely to form the suspensor
Suspensor → attaches the embryo to the embryo sac
Hypophysis → refers to the uppermost cell of the suspensor
Descendants of hypophysis forms the quiescent center or primary root meristem and columella initial
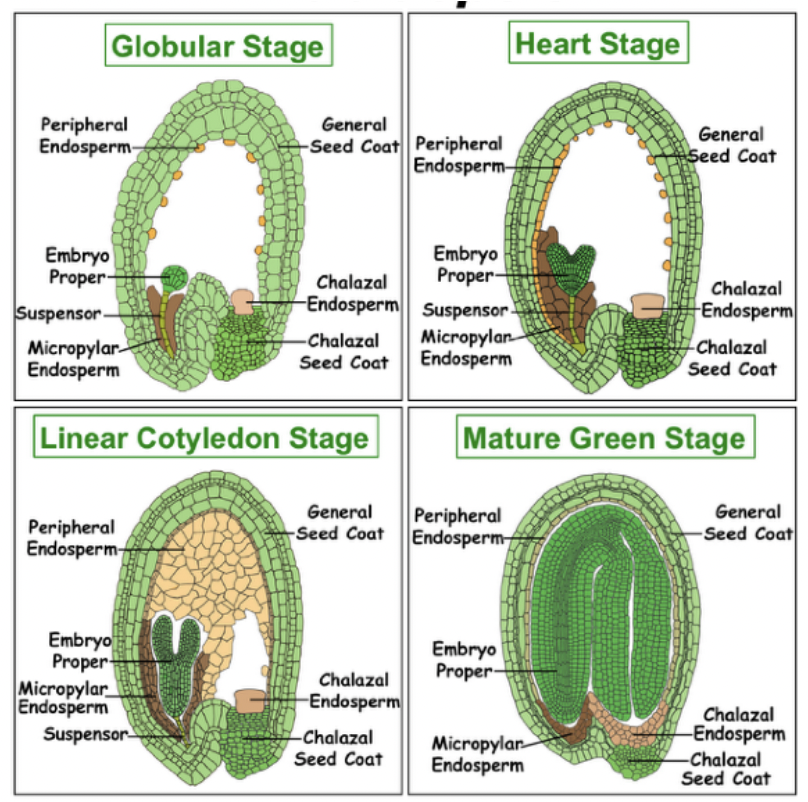
Apical cell becomes the embryo proper → GLOBULAR STAGE
2-celled spherical embryo → longitudinal division
Quadrant (4-celled) → 2-celled embryo divides longitudinally
Octant (8-celled) → 4-celled embryo divides transversely
Dermatogen stage (16-celled) → each of 8 cells produce a surface layer of 8 cells to cover itself
Surface layer → becomes protodern
First sign of tissue differentiation
32-celled and 64-celled embryo → protoderm and internal cells continue to divide
Radial symmetry is established
Triangular stage → transition between late globular and early heart stage
Rapid cell divisions occur leading to formation of 2 cotyledon primordia → HEART STAGE
RAM and SAM are established in the embryo
Procambium can be distinguished in late heart stage
Bilateral symmetry is established → axial polarity
Elongation of the embryo axis → TORPEDO STAGE
Hypocotyle and radicle recognized
Vascular tissue differentiation within begins
Suspensor deteriorates
Cotyledon → functions in food storage, food absorption, and/or photosynthesis
Cotyledons fold over assuming the WALKING STICK STAGE
MATURE EMBRYO
Radicle → embryonic root
Embryo is dormant
Seed is ready for dispersal
Endosperm
Triploid tissue formed when a sperm fertilized 2 polar nuclei
Some eudicots → endosperm divides and fill portion of mature seed
Function → stores nutrients
Non-endospermic eudicots → endosperm is digested and nutrients moved to 2 cotyledons
Suspensor
Structure formed by the larger basal cell after division of zygote
Range from single to massive collection of cells
Can contain tiers of multi-nucleated cells forming a syncytium
Have basal cells at micropylar end → site of max metabolic activity
Can be polyploid and/or undergo endoreduplication
Legumes → presence of giant polytene chromosomes
Suspensor pushes embryo proper into endosperm cavity and connects embryo proper to surrounding maternal and endosperm tissues
Serves as conduit for nutrients and growth regulators
Have structures that enhance ability to transfer molecules
Cell-wall ingrowths
Haustorial outgrowths
Numerous plasmodesmata
Lacks a cuticle layer
Organelles present: mitochondria, ER, specialized plastids
In early development stages, suspensor cells have higher RNA and protein synthesis levels than embryo proper
Hormones present: GA, auxin, cytokinin, abscisic acid
Programmed cell death upon entering maturation
Chief events of embryogenesis
Establishment of the precursors/initials for dermal, ground, and vascular tissues
Differentiated in a radial pattern
By globular stage (sometimes during octant stage)
Establishment of apical-basal polarity
By transition from globular to heart stage
Establishment of RAM and SAM
Heart stage
Monocot Embryogenesis
More complex than eudicots
Early embryo development is similar
Proembryo stage
First cell division is asymmetrical (in various planes)
Apical cell → divides faster to become embryo
Basal cell
Globular stage
Suspensor is not a single or double row of cells and is less differentiated
Late globular → outer epidermal layer is evident + group of cells on one side of proembryo divides faster to produce embryo axis
Scutellar stage
Remnant of cotyledon can be seen
Scutellum → single modified cotyledon that acts as a conductive tissue between endosperm and embryo axis
Coleoptilar stage
Embryo axis differentiates into plumule and radicle
Coleoptile → specialized tissue protecting the shoot
Coleorhiza → specialized tissue protecting the root
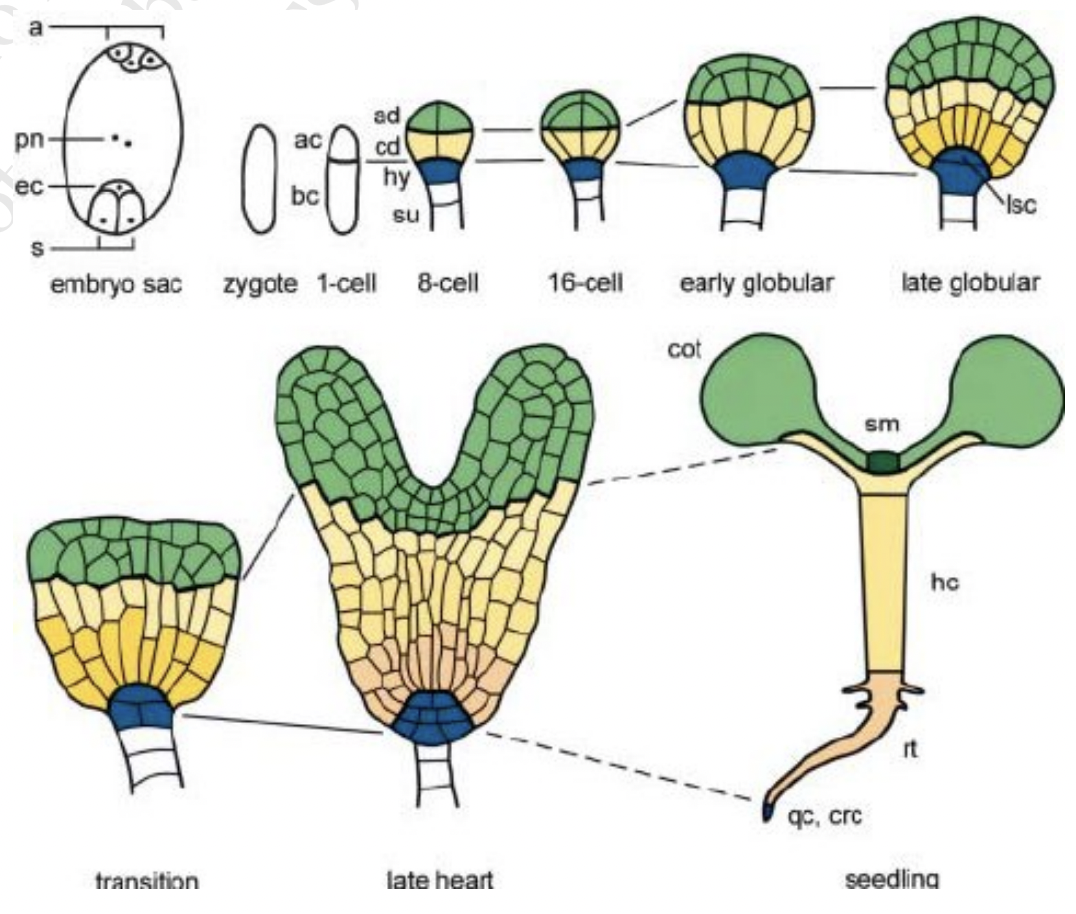
Plant embryos form from regions that develop autonomously
8cell stage has four regions with different developmental fates
Domains | Composition | Fates |
Apical | 4 most apical cells | SAM Most cotyledons |
Central | 4 lower cells | Hypocotyl Root Contributes to cotyledon and RAM |
Basal | Hypophysis of suspensor | Distal parts of RAM Quiescent center Stem cells of central root cap |
Extra suspensor | (non embryonic) | Pushes embryo into ovule lumen and provides connection to mother tissue |
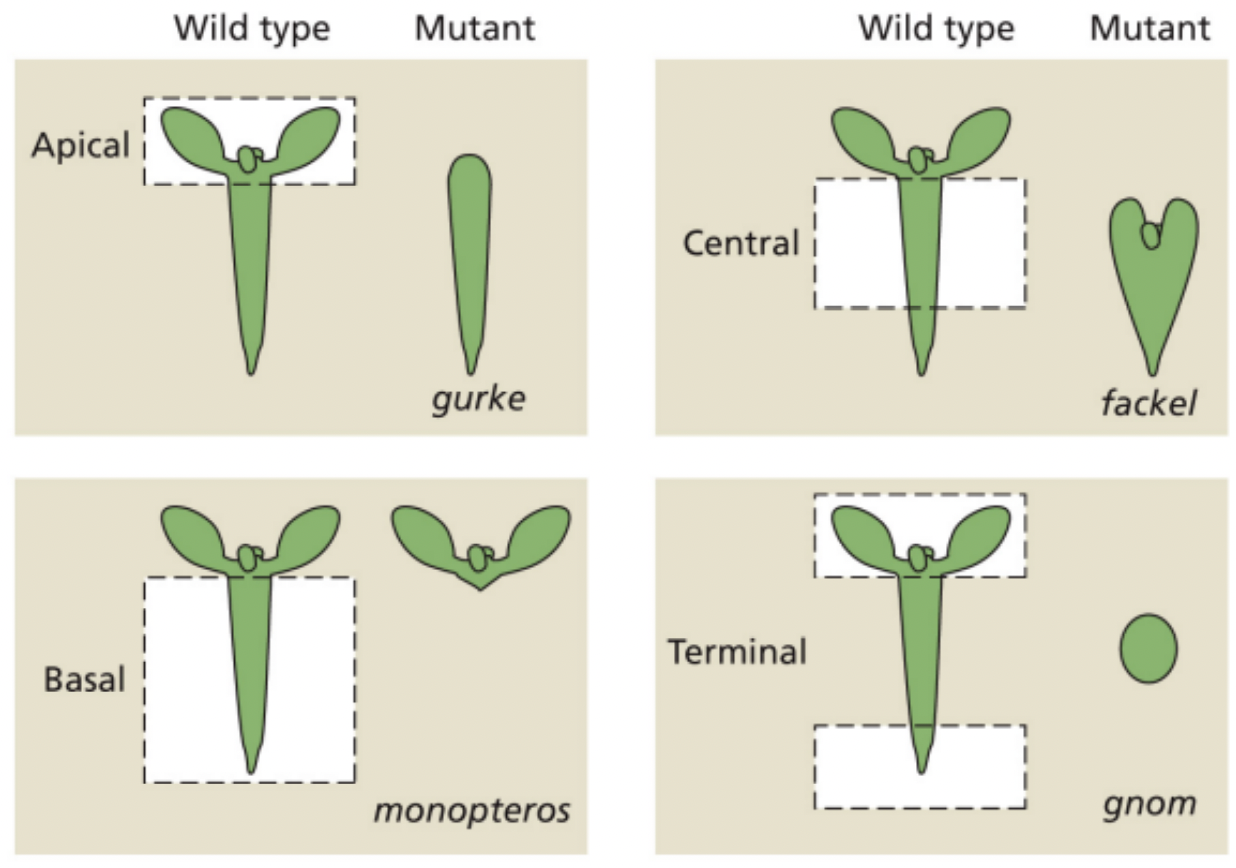
4 regulatory genes affect aspects of the apical-basal pattern
Mutations in these genes result in deletion of specific embryonic regions
Mutations | Region deleted | Fates |
gurke | Apical region | No Cotyledons No SAM |
fackel | Central region | Cotyledon attached to root directly |
monopteros | Central region Basal region | No hypocotyl No root |
gnom | Apical region Basal region | No root No cotyledon Extreme: spherical and no axial polarity |

Mutants | Phenotype | involved |
knolle, keule |
|
|
lec (leafy cotyledon) |
|
|
Endosperm Development
Seed development initiated by double fertilization
Fertilization of haploid egg cell → diploid embryo
Fertilization of diploid central cell → triploid endosperm
Function
Nourish and support embryo by delivering nutrients acquired from mother plant
Protects embryo from mechanical injury
Fates
Consumed by developing embryo before maturation
Persist in mature seed and used up during seed germination
Gymnosperm endosperm is haploid and formed before fertilization
Angiosperm endosperm is triploid and formed after fertilization
Absent in : Orchidaceae, Podostemaceae, and Trapaceae
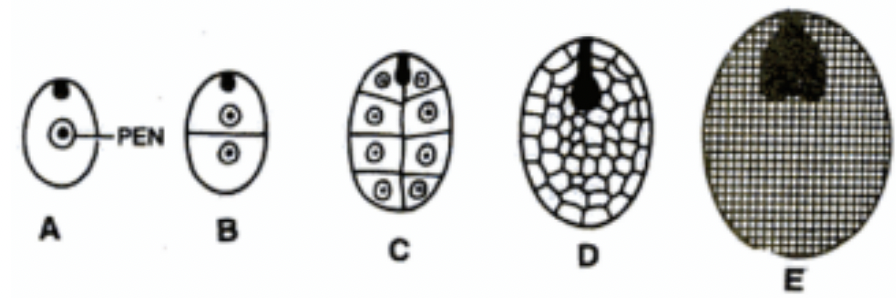
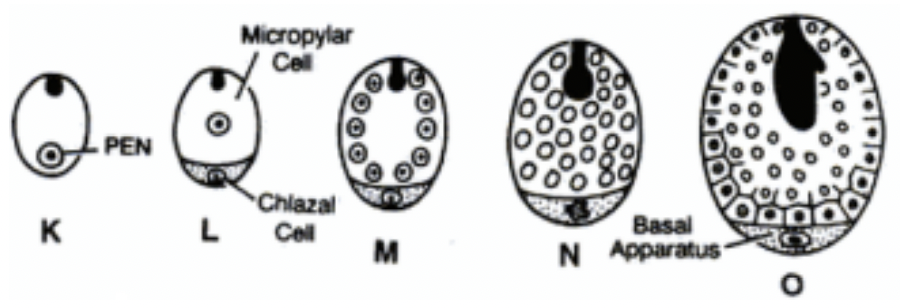
Primary endosperm nucleus (PEN) → where endosperm develops from as a result of triple fusion
Triploid (fusion of one male gamete with 2 polar nuclei)
Free nuclear proliferation without cytokinesis (syncytial.coenocytic phase) → cellularization phase initiated in a region surrounding embryo → outer to inner region of endosperm
Arabidopsis, endosperm cellularization during early heart stage
Endosperm cellularization failure → embryo arrest and seed abortion
Early stages of seed germination
When seed dormancy is broken, embryo starts to produce GA
GA triggers aleurone cells within the seed to start releasing amylase
Amylase will hydrolyze starch in endosperm into maltose
Cotyledons absorb the maltose from the endosperm and give it to the embryo
Types of Endosperm
Cellular endosperm (advanced)
PEN division → cell wall formation
First division = 2 equal sized cells: chalazal and mycropylar cells
Subsequent divisions followed by cell wall formation
Thus, endosperm is cellular from the beginning
Nuclear Endosperm (primitive)
Most common in angiosperms
PEN division = many free nuclei → Coenocytic stage
Division not accompanied by wall formation
Free nuclei arrange towards periphery of cytoplasm → wall formation starts from periphery towards center
Cell plate formation centripetally
Arabidopsis and Capsella
Liquid endosperm of coconut
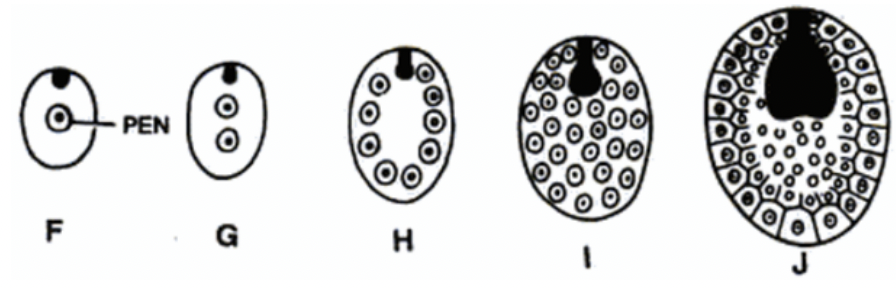
Helobial endosperm (advanced)
Intermediate between cellular and nuclear types
PEN division → large micropylar cell and small chalazal cell
Nucleaus of micropylar divides freely without cell wall formation and cell wall forms from periphery to inward
Nuclaeus of chalazal cell remains undivided or divides for few times (basal apparatus)
Helobial endosperm
WEEK 1
All the cells in the plant body will have the exact same copy of the DNA of the genes. Different gene expression is the reason for the different cell types
Plants: Formed after germination. Dependent on apical meristems. Indeterminate growth.
Plants are sedentary, instead they alter its development and morphology to help them survive.
Cell division in plants are concentrated in the meristems. In animals, it happens everywhere
Indeterminate: shoot and root
Determinate: flower meristem,
Developmental plasticity → effect of environment to the development of plants
Unlike animals. By the time they get to their environment, they are already fully formed
Totipotent → ability to become any cell type in the body of that organism
Zygote can give rise to any other structure later on → naturally totipotent
Even if it is a mature cell type it can be induced to become totipotent. This is not always natural so it needs to be induced
Callus
Animals → Cells of the blastula
Youngest cells of the meristems are still naturally totipotent
Pluripotent → lesser ability because fates are determined, slightly determined fates
Protoderm cannot give rise to a xylem and phloem but it can develop into any epidermis cell type
Animals → cells of the gastrula
Embryogenesis
Embryo formation in animals, all organs have been formed already
Development during embryogenesis
Plant embryogenesis is just one small portion of the entire plant life.
Only meristems are established
No organs in plant embryogenesis
Development happens post embryonic
Plant development has no cell migration
Anticlinal → perpendicular, all in one row, for wide organs
Periclinal → parallel, all in one column, for elongated organs
Depends on where cell plate is located
Model organisms
Arabidopsis → eudicots
Zea mays → monocot
Tobacco
Rice
Characteristics
Short life cycle to look at progeny
With high seed production = more offspring = more replicates
Self-fertilization = to look at homozygosity and heterozygosity of gene
WEEK 1
Development vs growth
Development → differentiation, maturation
Growth → increase in number or size of cells
Differentiation → normal process,
Meristem cell becomes cell of stomata then guard cell
Proplastids → chloroplast
Dedifferentiation → mature to immature
Phloem cell → procambium
Mature leaf → callus
Chloroplast → proplastid
Rediffirentiation → Mature cell to nother mature cell type
Chloroplast → chromoplast, vice versa
Callus → shoot or root
Do they need to dedifferentiate to differentiate?
Can occur directly
Mesophyll cells → tracheary element without reversion to undifferentiated state
Pattern formation
Asymmetric cell division
Apical → embryo proper, transversely and longitudinally
Basal → suspensor, longitudinal
Lateral inhibition → prevents cells beside it from becoming the same cell type as them
Programmed cell death → holes in leaves
Plane of cell division very important → determines plant morphology
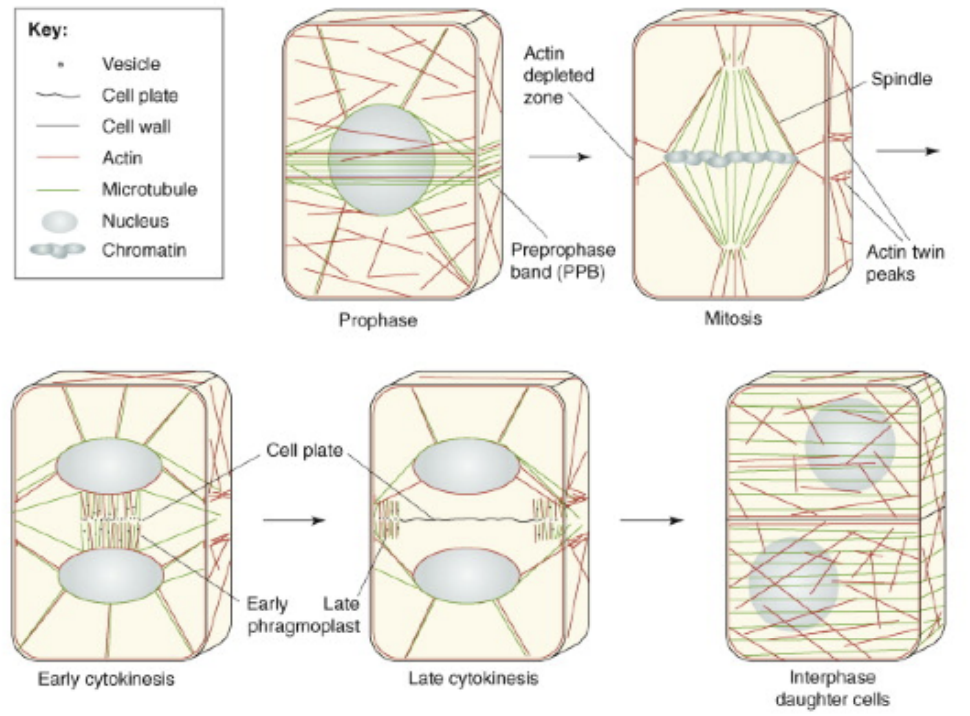
Preprophase band and pragmoplast → both composed of microtubules and actin but they both appear in different stages
Preprophase occurs prior to actual mitosis, during interphase specifically G1 phase
Preprophase disappears and leaves behind a signal telling the phragmoplast where to form
Phragmoplast appears during telophase of mitosis
Phragmoplast tells the golgi derived vesicles where to go
Cell plate formation form the inside going out starting from the middle spreading outward
CDZ is part of cytoplasm where phragmoplast is formed → just a region
Centrifugally
Microtubules are the tracks of the train
Cellulose microfibrils deposited the same way the microtubules are laid
Cellulose microfibrils also provide guidance
Dual guidance model by microtubules and existing cellulose microfibrils
Auxin
Auxin is the hormone
Presence of auxin
TIR1 will be able to mark Aux/IAA for degradation
ARFs will be free to induce changes in transcription
Change in transcription happens
Absence
TIRI is not able to mark Aux/IAA for degradation
AFFs are not free to induce changes in transcription
Communication
Apoplast → cell walls
Symplast → plasmodesmata
Week 2
Plant life cycle
Gametophyte generation → haploid cells that function to produce gametes via mitosis
Haploid and multicellular
Different genetic composition compared to sporophyte due to meiosis
Fern → prothallus
Spore formation via meiosis (haploid and unicellular)
Megasporogenesis
Microsporogenesis
Fern → sporogenesis only
Sexual reproduction → genetic variation
Pistil and stamen structures all part of sporophyte
Embryo sac and pollen grain are gametophyte
Stamen → within anther → in pollen sacs (microsporangium) → meiosis for microspore formation
Dehiscence → anther opened and pollen grains are released
Microspores are in tetrads → released from tetrads become pollen grains but whether they are not they are mature we are not sure
Mature → two or more cells inside
Vegetative + generative = bicellular
2 sperm cells + vegetative = tricellular
Not mature → only one cell
Microsporocytes are not in tetrads
Pollen grains are smaller than microsporocytes
Microsporocytes are larger
Pollen grain not completely round
Microsporocyte are very round
Microsporocytes have walls that connect them so that they undergo meiosis at the same time
L3 → connective and vascular + inner tapetum
LI → outer → epidermis and stomium
L2 → middle → primary parietal (outer), sporogenous cells (inner)
Hypodermis
Middle wall layer, tapetum, pollen mother cells, endothecium → all diploid
Microsporocytes → haploid
Degeneration → complete deterioration of the structure
Senescence → related to aging, meaning the structure aged and stops to divide completely
Stomium is the point where anthers dehisce
CCC → inner to stomium composed of large cells sometimes with crystals that degenerate and connect the two pollen sacs
Week 3
Pistil → stigma, style ovary
Compound pistil = fusion of carpels (typically fusion in the ovary like lily)
Many separate carpels = 1:1 ratio
All pistils are gynoecium
Ovule contains the cells that become the embryo sac later on
Nucellus surrounds the embryo sac → megasporangium
Megaspore mother cell via meiosis = megaspore = 3 degenerate, 1 functional
3rd mitosis then cytokinesis
Synergids accept the pollen tube
Monosporic
All resulting cells are geentically identical
Bisporic, bimitotic
1 binucleate cell → 2 different haploid nuclei divide → cells of embryo sac have different genetics
Tetrasporic, bimitotic
1 tetranucleate cell → 4 different haploid nuclei divide → cells of embryo sac have different genetics
Lily
Nuclei fusion → 3n
Antipodal are 3n
Synergids are n
Polar nuclei 1 is n
Polar nuclei 2 is 3n
Endosperm is 5n (4n + haploid sperm)
Pollination
Resulting offspring will never be the same as the parent plant even if self pollination
Never be a clone because of meiosis
Clone only for asexual reproduction
Spore formation = sexual reproduction = no clones
Embryogenesis
Shoot and root apical meristem establishment = embryogenesis
Monocot → coleoptile, coleorhiza, scutellum
Suspensor is multiseriate
Scutellum doesnt have a storage function, it is an absorptive structure
Coleorhiza protects the RAM
Coleoptile protects the SAM
1 cotyledon
Most of the seed is full of endosperm
Eudicot
Dermatogen stage → 16 cell stage is where tissue differentiation can be observed
Outer 8 become protoderm
Hypophysis is the only cell that becomes part of the RAM
Heart stage → cotyledon primordia
Torpedo stage → elongation of cotyledon
Walking stick stage → cotyledons fold over to fit inside the seed
Mature
Go (GURKE) For (FACKEL) More (MONOPTEROS) Gold (GNOM)
Endosperm development → all become cellular at the end
Nuclear → outermost to innermost cytokinesis
Helobial → Chalazal cell does not divide, upper cell like nuclear