Establishing Pregnancy: Embryogensis, Maternal Recognition of Pregnancy, Implantation, and Placentation
Summary of Major Concepts
For a pregnancy to advance beyond blastocyst stage, the conceptus must provide a signal to maintain maternal production of progesterone and form a placenta with the maternal uterus
Development of the trophectoderm is a prerequisite for maternal recognition of pregnancy and implantation of the embryo into the uterus
Maternal recognition of pregnancy signals prevent/delay luteolysis by interfering with PGF2alpha-induced regression of the CL or by boosting the luteotropic signal
Placentas can be categorized based on the distribution of chorionic villi and by the histology of the interface between maternal-fetal tissues
Overview of Early Pregnancy
Pregnancy first needs to be successful within its environment and fertilization, but to continue, the zygote needs to develop into an embryo, and have a physiological relationship with the mother to live outside the body
Embryogensis: Zygote transforms into an embryo
In order for a fetus live outside the body:
The uterus remains in a state to support pregnancy due to a pregnancy recognition signal
Nutrients and wastes are exchanged due to the placenta interacting with the uterine endometrium
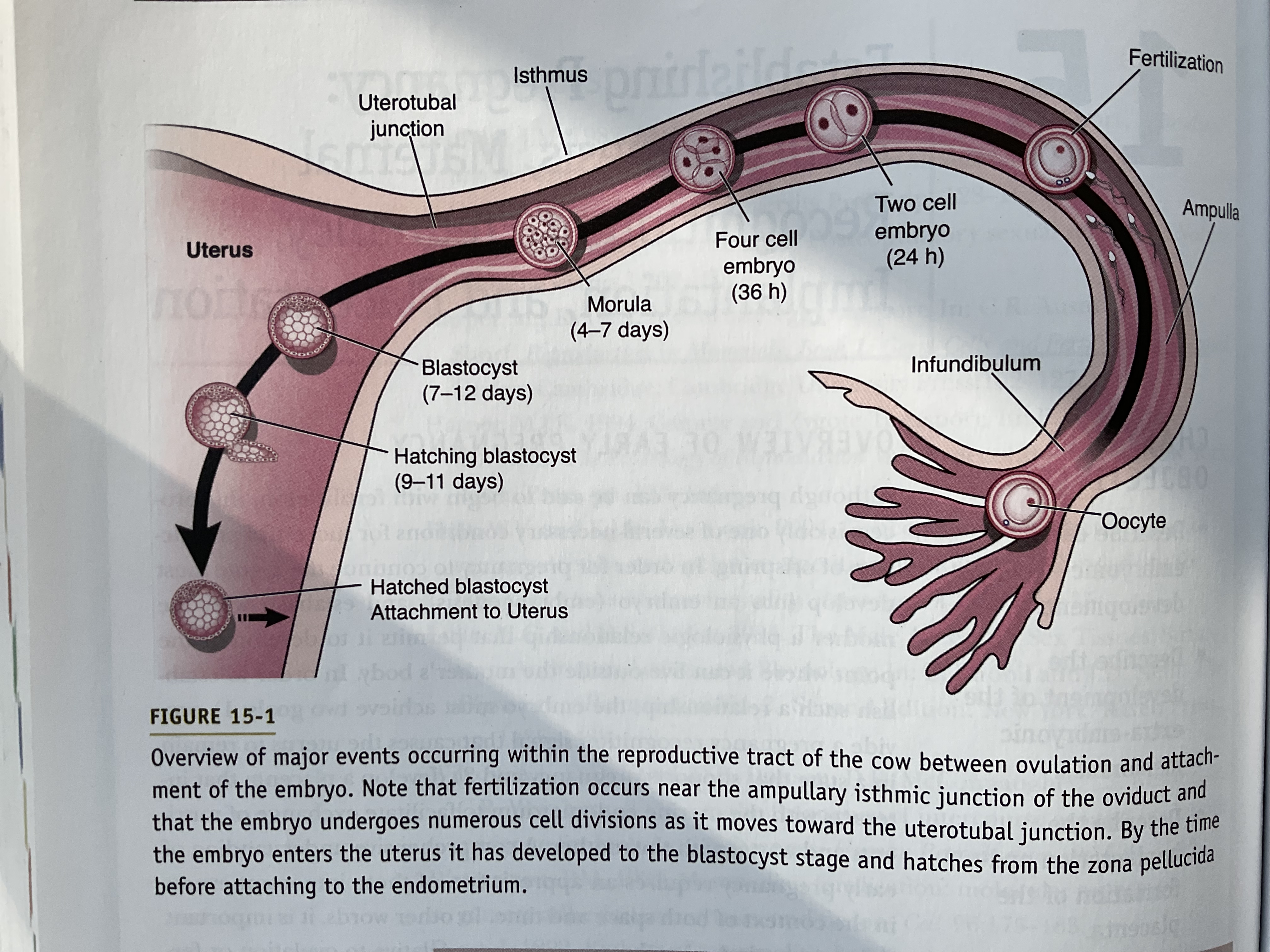
Early Embryogenesis
Zygote is present in the isthmus of oviduct
Embryo has the ability to develop a pregnancy-recognition signal and interact with the uterus
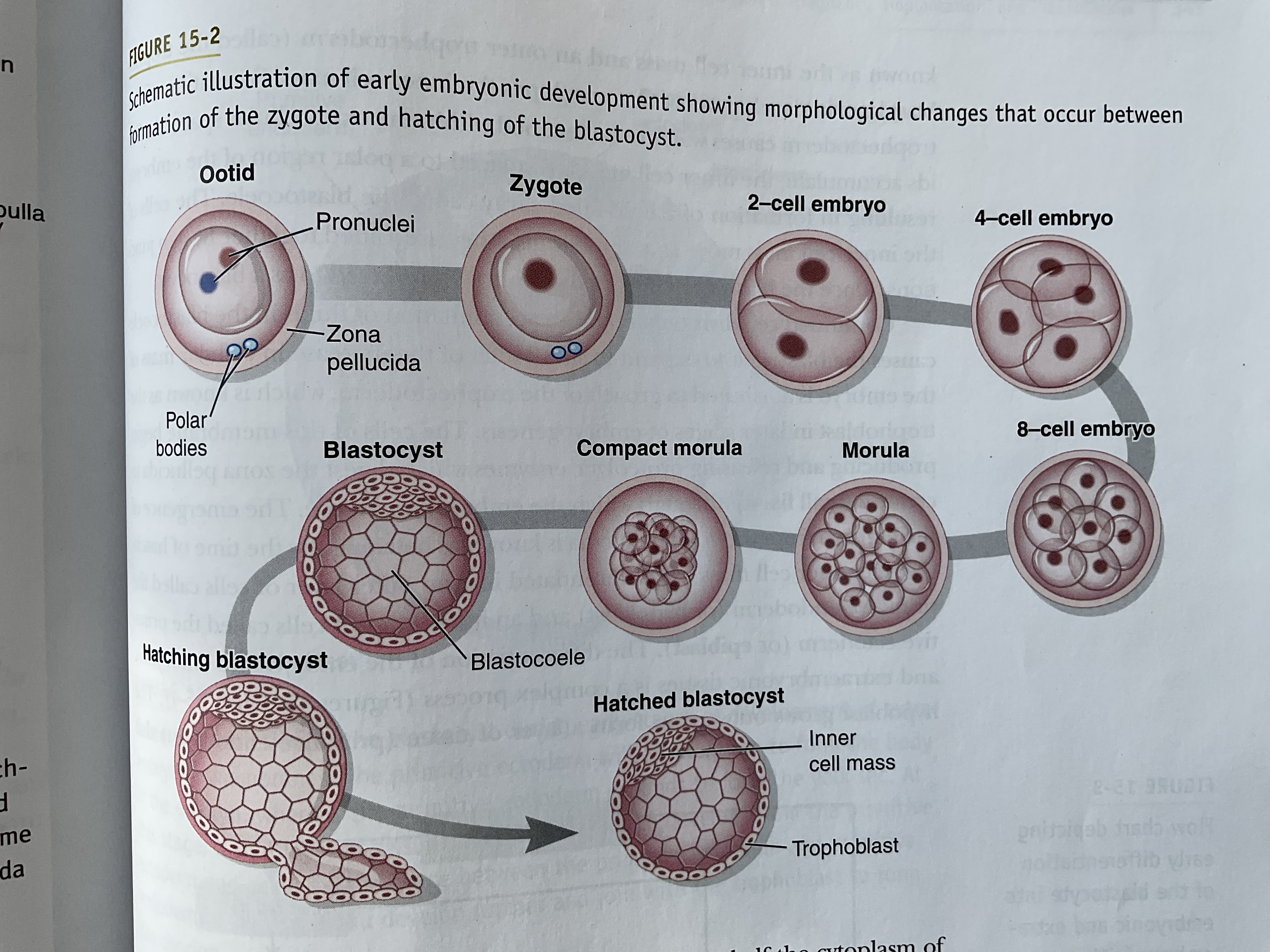
Zygote under goes cleavage division
The cells of the embryo (blastomeres) are totipotent up to the 16-cell stage
Totipotent: Each cell has the ability to give rise to an individual
Morula: At the 16-cell stage, embryo consists of densely packed cells called the morula and further compact into a 32-compact cell
A blastomere cell create tight junctions and create 2 major classes of cells:
Inner cell mass
Trophectoderm
Trophectoderm causes water to retain in the extracellular space called the blastocoele
As the trophectoderm grows, the cells of this membrane produce and release proteolytic enzymes which digest the zona pellucida to create a small fissure through which the embryo can escape
Primitive endoderm (hypoblast): When hatching, the inner cell mass has differentiated into an outer layer of cells, endoderm
Ectoderm: Inner cell mass when the blastocyst hatches
The hypoblast (endoderm) grows outward to form a layer of tissue that lines the blastocoele and eventually gives rise to the yolk sac
The epiblast (ectoderm) will form an extraembryonic membrane known as the amnion as well as the body of the embryo, which differentiates into the three embryonic germ layers:
Mesoderm
Endoderm
Ectoderm
Trophoblast remains on the exterior of the embryo and gives rise to the membranes that will become the placenta
Progesterone is ESSENTIAL for pregnancy
Progesterone sustains pregnancy in the uterus
Mammals rely on two strategies to ensure progesterone levels remain elevated until a conceptus (embryo in the uterus) is present in the uterus
Luteal Phase that covers the duration of gestation
Induced ovulators (rabbits and cats), copulation induces ovulation and development of the corpora lutea, which are function whether there are embryos or not
Mating, not the presence of embryos, prevents luteolysis
Length of luteal phase is shorter than the time required for the embryo to implant in the uterus
Pregnancy-recognition signal generated by the conceptus acts on the maternal reproductive system to prolong the life of the corpus luteum
In some species, CL life extends until placenta takes over the production of progesterone
In other cases, embryo generates signal that maintains the CL throughout entire pregnancy
Embryos typically generate signals a few days after hatching
Maternal Recognition of Pregnancy
Disrupts luteolysis to maintain the pregnant state
The goal of the conceptus is to derive nutrient support from the mother
The goal of the mother is to consume and conserve fuels
In order to survive in the maternal reproductive tract, the conceptus (essentially a foreign body) must invade the maternal body, intervene with the mothers ovarian cycle, and suppress maternal defensive mechanisms intended to resist the physiologic disruptions
Ruminants Maternal Recognition of Pregnancy
In cows, the pregnancy recognition signal is: bovine interferon-t (bIFN-t)
Ruminants rely on additional mechanisms to prevent the regression of the CL. Such as ovine embryos produce a protein that acts on large luteal cells to protect them from the luteolytic actions of PGF2-alpha
Ewes have a peptide, ovine interferon-t (oIFN-t), which is produced by trophoblasts and are detected 13-21 days after ovulation
This is the time in which the CL must be rescued in order for pregnancy to continue
oIFN-t prevents normal onset of luteolysis by interfering with release of PGF2-alpha by the endometrial epithelium. this involves blocking expression of receptors for oxytocin on these epithelial cells
Oxytocin is responsible for inducing the pulsation release of PGF2-alpha, required for luteolysis
Swine Maternal Recognition of Pregnancy
Pregnancy recognition signal disrupts the actions of PGF2-alpha
Swine signal of maternal recognition is estradiol
Porcine blastocyst produces large amounts of the steroid hormone estradiol between 10-12th day of pregnancy
Epithelial cells release the hormone PGF2-alpha into the extracellular space of the submucosa
PGF2-alpha then enters capillaries and travels to veins that drain the uterus
Luteolytic hormone is released into the lumen of the uterus; in an exocrine fashion
This is the primary way estradiol acts to lengthen the life of the CL
Another mechanism to lengthen the life of the CL is when estradiol appears to protect luteal cells from the luteolytic actions of PGF2-alpha
Horses Maternal Recognition of Pregnancy
Though their is a clear signal that conceptus generates that prolongs the life of the CL, it is unclear on what the exact mechanism (or hormone) causes this
Equine conceptus migrates through the uterus, distributing a blocker of PGF2-alpha, reducing the concentration of PGF2-alpha found in the uterine vein until 35 days after pregnancy
Whatever the signal is, it is not effective enough to extend the life of the corpus luteum to the point where the placenta takes over the production of progesterone (160 days after ovulation)
The chorion of the conceptus produces an LH-like hormone: equine chorionic gonadotropin (eCG)
Induces luteinization of follicles
These accessory corpora lutea produce enough progesterone to maintain pregnancy until the placenta takes over
Primates Maternal Recognition of Pregnancy
Involves a signal that extends the life of the CL until the placenta begins to produce enough progesterone to maintain pregnancy
Luteal-placental shift: Progesterone produced by the corpus luteum eventually declines, but by this time the production from the placenta is adequate to maintain the uterus in a pregnant state
Suppression of the Maternal Immune System
Successful pregnancy depends on the embryo’s ability to sustain maternal production of progesterone and to counteract mechanisms that defend the maternal system from invasion of foreign bodies
Although the uterus is an immunocompetent organ (capable of mounting an immunologic attack), and generates antibodies in response to embryo-specific antigens which circulate in maternal blood, the conceptus somehow counteracts these maternal defense mechanisms
The conceptus uses diversionary tactics to evade destruction by immune responses (behaves as a invading parasite)
Development of the Placenta
Development of the placenta (placentation) is a critical prerequisite for development of the embryo past the blastocyst stage
2 major steps
Trophectoderm of the embryo attaches to the luminal epithelium of the uterus
Embryo develops extraembryonic (or fetal) membranes that become intimately associated with maternal tissue to form the placenta
In species such as humans and rodents: The fetal membrane invades into the submucosa of the uterine epithelium
In species like horses, swine, and ruminants: Little to no invasion of the fetal membrane and implantation is synonymous with attachment
Implantation depends on two conditions:
Development of the embryo to the blastocyst stage
Development of the uterus to a state that favors implantation
The latter process is dependent on progesterone, but can require estradiol in some cases
Attachment
Attachment of the embryo to the uterus involves the direct interaction of the trophoblast with the uterine epithelium
Contains two stages: Apposition and adhesion
Apposition: Development of intimate contact between the embryonic and maternal tissues
Begins when the embryo becomes immobilized within the uterus (2 mechanisms)
Carnivores, ruminants, horses: extraembryonic membranes expand to fill the lumen of the uterus causing membranes to become apposed to the uterine epithelium
Primates + rodents: Embryo is small, so membranes don’t fill uterus
Adhesion: Involves the intertwining of microvilli on the maternal and embryonic epithelial cells (makes it difficult to dislodge)
Occurs once the trophoblast is stabilized against the uterine epithelium
Invasion
In primates and rodents, the trophectoderm penetrates the basal lamina of the uterine mucosa to establish connections with the maternal vasculature
Once the conceptus adheres to the maternal cells, it induces decidualization of stromal cells (fibroblast-like cells in the submucosa) and endothelial cells
These cells are transformed into decidual cells, which form the maternal portion of the placenta in these species
Formation of Extra-Embryonic Membranes
Embryogenesis continues during implantation
The epiblast differentiates into three majors layers (Ectoderm, mesoderm, and endoderm) after blastocoele formation to form the yolk sac from the hypoblast
Mesoderm grows outward and fuses with trophectoderm to form the chorion
Projects bilaterally to form amniotic folds
Amniotic folds will meet to form the amniotic cavity that surrounds the embryo
When this is occurring, the yolk sac regresses and the allantois enlarges to fill the cavity between the chorion and amnion
Allantois: A vascularized pouch from the hindgut
The allantois will fuse to the chorion to form the allantochorion (chorioallantoic membrane)
Chorioallantoic Membrane: Becomes the fetal component of the placenta
Chorion is avascular
Allantois: Supply’s the fetal blood supply to the placenta
Classification of Placentae
One way to classify the placenta is based on the origin of fetal blood vessels that vascularize the chorion
Another way to classify is based upon their gross morphologies
Contact between maternal and fetal tissues is the result of interaction between microvilli of the maternal and fetal epithelium
Distribution of the microvilli of the chorion can be diffuse (covering the available surface) or restricted to specialized regions
Pigs: villi are scattered across the surface
Horses: villi are in small clusters called microcotyledons
Cotyledonary: distribution of microvilli (ruminants)
Cotyledons: Cotyledonary placenta consists of discrete zones of microvilli called cotyledons
Derived from the trophoblasts and have capillary beds that are supplied by and drained by blood vessels of the fetus
Each cotyledon establishes a physical interaction with a maternal caruncle forming a placentome
Zonary placentas: A band of microvilli encompasses the equatorial region of the allantochorion
This type of placenta fills the lumen of the uterine horn so the fetal membrane makes contact with the maternal tissue along the entire circumference (transfer zone)
Pigmented due to small blood clots
Maternal caruncle tissue and chorionic cotyledon are attached through chorionic villi interdigitate with folds in the mucosal epithelium of the caruncle