Human Evolution

Lineage + Taxa


Evolutionary Relationship between Humans and Apes
Overview
Humans and Apes: Humans are universally agreed to have evolved from within the apes.
Apes Classification:
Lesser Apes:
Gibbons (Hylobates): Southeast Asia
Great Apes:
Orangutan (Pongo pygmaeus): Southeast Asia
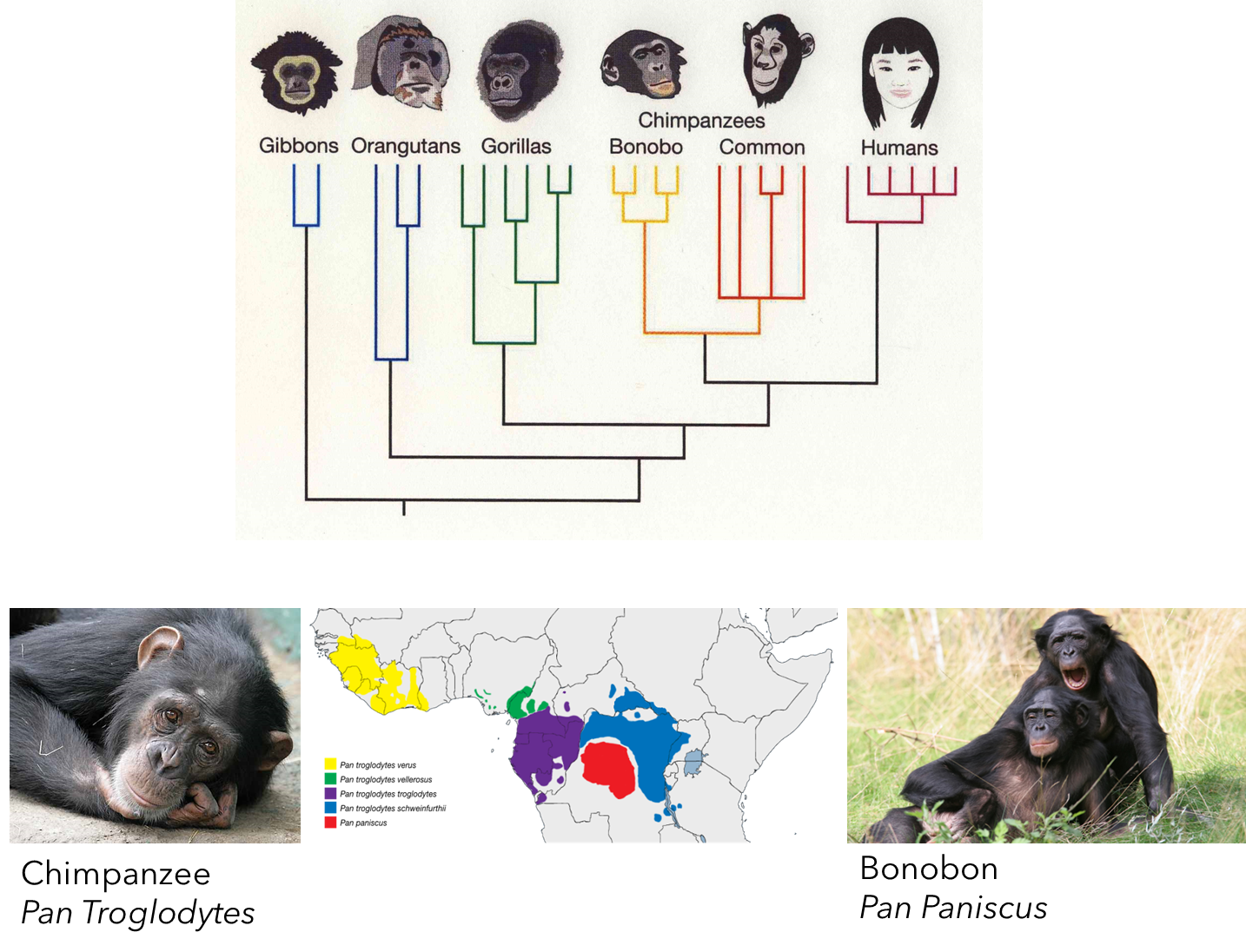
African Great Apes:
Gorilla (Gorilla gorilla)
Common Chimpanzee (Pan troglodytes)
Bonobo (Pygmy Chimpanzee) (Pan paniscus)
Shared Derived Characteristics of Apes
Brain Size:
Relatively large brains compared to other primates.
Absence of a Tail:
Unlike other primates, apes do not have tails.
Posture:
More erect posture than other primates.
Flexibility:
Increased flexibility in:
Hips and Ankles: Enhanced movement and posture.
Wrist and Thumb: Greater dexterity and range of motion.
Arm and Shoulder Structure:
Adaptations in the structure and use of arms and shoulders.
Evidence of Common Ancestry
Morphological Evidence:
Derived traits shared among apes indicate a common ancestor within the apes.
Molecular Analyses:
Provide unequivocal support for the evolutionary relationship between humans and apes.
Phylogenetic Relationships Among Apes
Overview
Reconstruction of Phylogeny:
Humans and Great Apes: Humans are placed within the great apes, specifically with the African great apes.
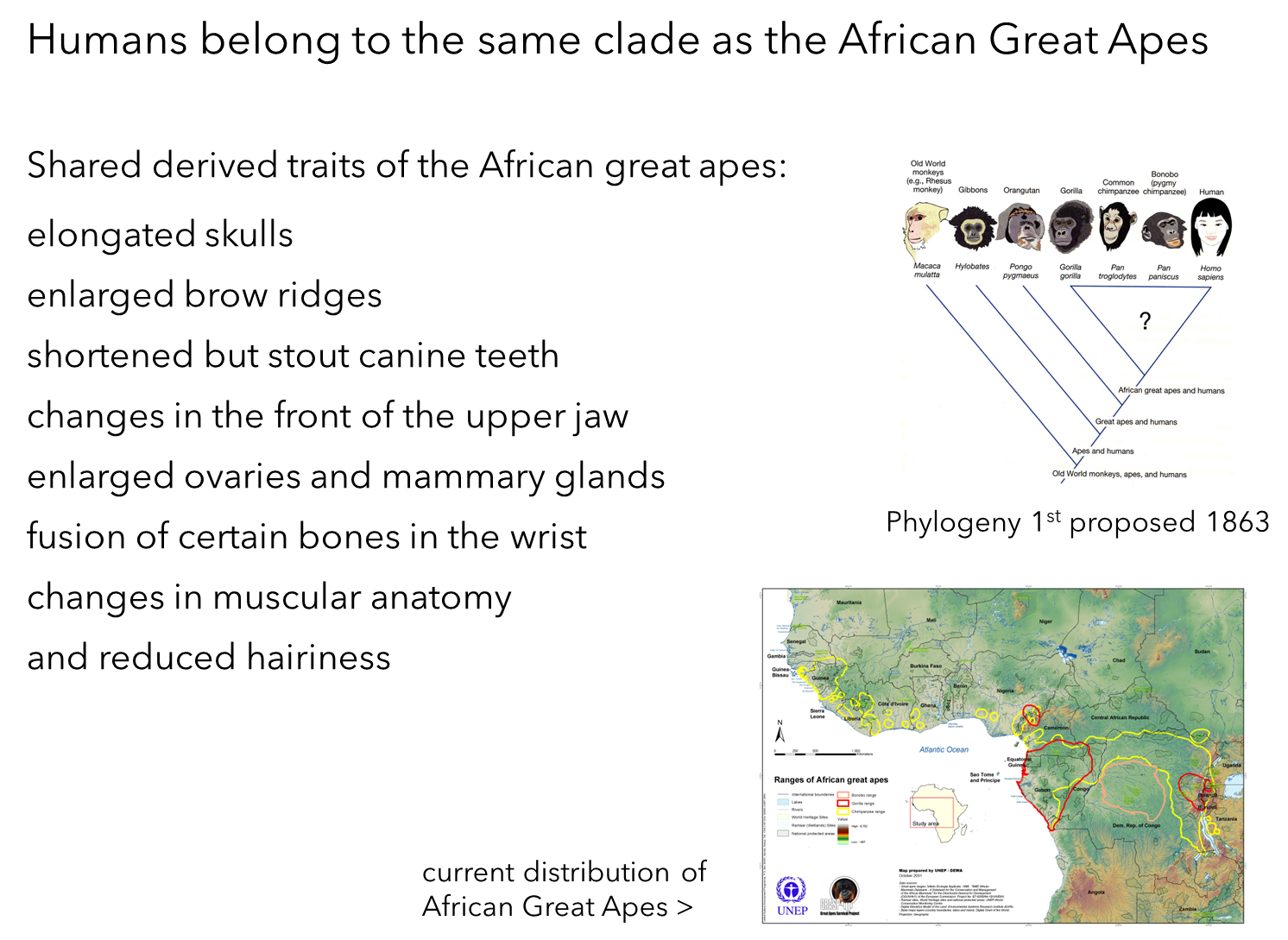
Origin of Phylogeny:
Thomas Henry Huxley (1863): First proposed the phylogenetic tree showing this relationship.
Initial Dispute: Huxley's proposal was controversial but has been accepted with more recent data.
Supporting Evidence
Cladistic Analyses:
Morphological Data: Supports the tree by highlighting shared derived traits between humans and African great apes.
Molecular Sequence Data: Also supports the tree, confirming the close relationship between humans and African great apes.
Derived Morphological Traits:
Elongated Skulls: Unique skull structure compared to other apes.
Enlarged Brow Ridges: Distinctive brow ridge features.
Shortened but Stout Canine Teeth: Adaptations in dental morphology.
Changes in Upper Jaw: Structural changes in the upper jaw.
Fusion of Wrist Bones: Unique wrist bone structure.
Enlarged Ovaries and Mammary Glands: Differences in reproductive anatomy.
Changes in Muscular Anatomy: Modifications in muscle structure.
Reduced Hairiness: Less hair compared to other apes.
Molecular Data:
Historical Agreement: Molecular sequence data has consistently shown a close relationship between humans and African great apes since the early days of molecular systematics.
Phylogenetic Trees: Early phylogenies identified humans, gorillas, and chimpanzees as close relatives but did not fully resolve their exact evolutionary relationships.
Scientific Effort:
Research Investment: Significant effort has been invested in resolving the evolutionary relationships among these species, possibly more than any other species-level problem in systematics.
Overview
Current Consensus:
Human and Chimpanzee Relationship: Humans and chimpanzees are more closely related to each other than to gorillas.
Historical Context
Decades of Debate:
The consensus on the evolutionary relationships took time to form due to several factors.
Reasons for Delay
Conflicts Among Molecular Data Sets:
Different molecular data sets initially produced conflicting results regarding the relationships among the great apes.
Conflicts Between Molecular and Morphological Evidence:
Discrepancies were found between molecular data and morphological evidence.
Convergent Evolution:
Both morphological and molecular data sets experienced convergent evolution, which led to conflicts.
Convergent Evolution: When different species evolve similar traits independently, this can obscure true evolutionary relationships.
Summary
Final Consensus:
Despite initial conflicts, researchers agree that humans and chimpanzees share a closer evolutionary relationship compared to their relationship with gorillas.
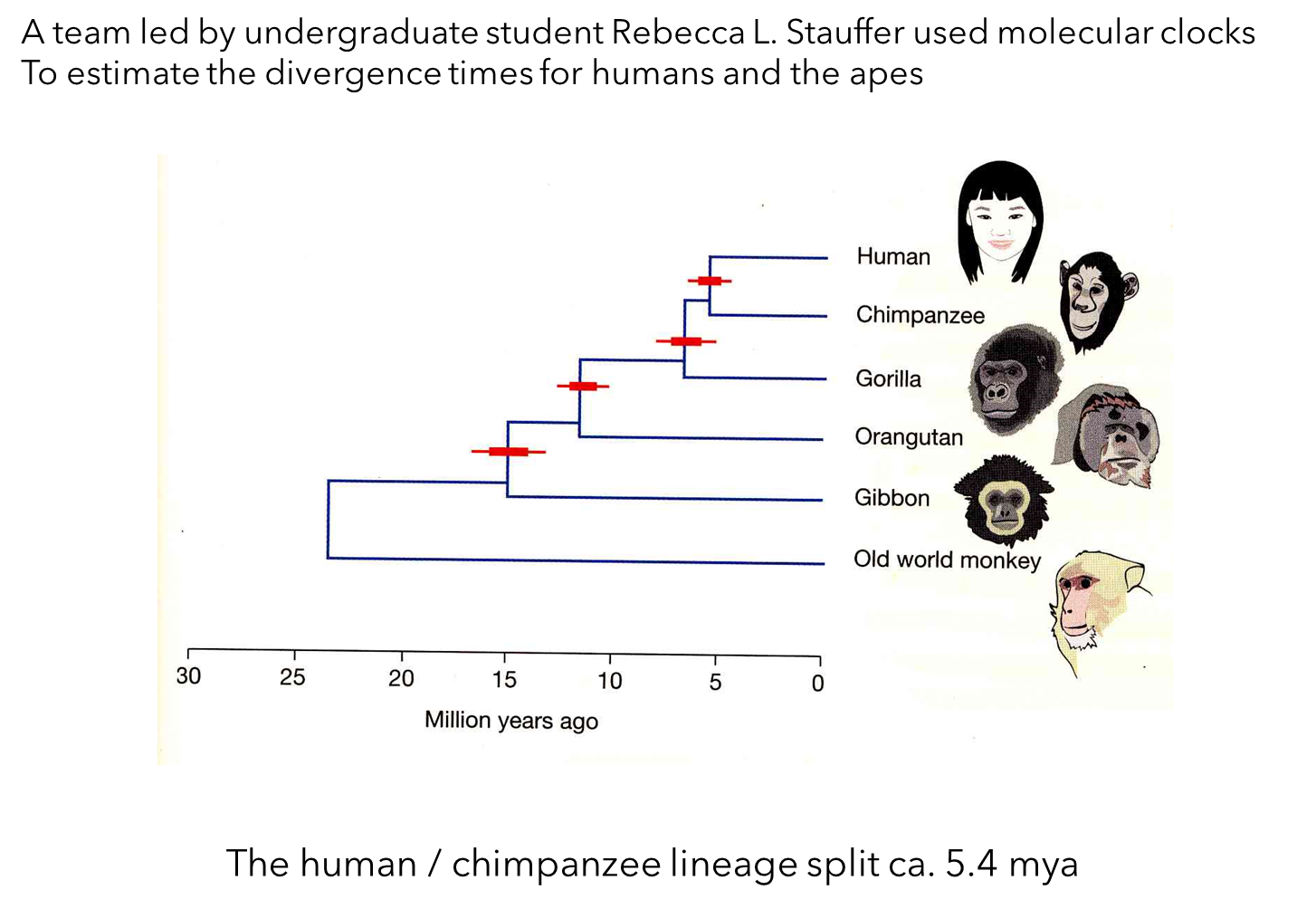
Estimating Divergence Times
Study Overview
Research Team:
Led by Rebecca L. Stauffer in the lab of S. Blair Hedges.
Published in 2001 (Stauffer et al. 2001).
Methodology
Molecular Clocks:
Used to estimate divergence times by comparing sequence differences in protein-coding genes.
Fossil Record:
Old World monkeys diverged from apes approximately 23.3 million years ago.
Gene Comparison:
Rate of Gene Evolution: Determined from differences between apes and Old World monkeys.
Divergence Estimation: Applied to humans versus chimpanzees to estimate divergence times among apes.
Data Integration:
Combined data from dozens of genes to improve accuracy.
Divergence Estimates
Gorillas:
Diverged from the lineage leading to humans and chimpanzees roughly 6.4 million years ago (± 1.5 million years).
Humans and Chimpanzees:
Split from each other approximately 5.4 million years ago (± 1.5 million years).
Genetic Differences
Time Span:
The divergence occurred between 5.4 to 6.4 million years ago.
Genetic Evolution:
Significant genetic differences have accumulated over this period.
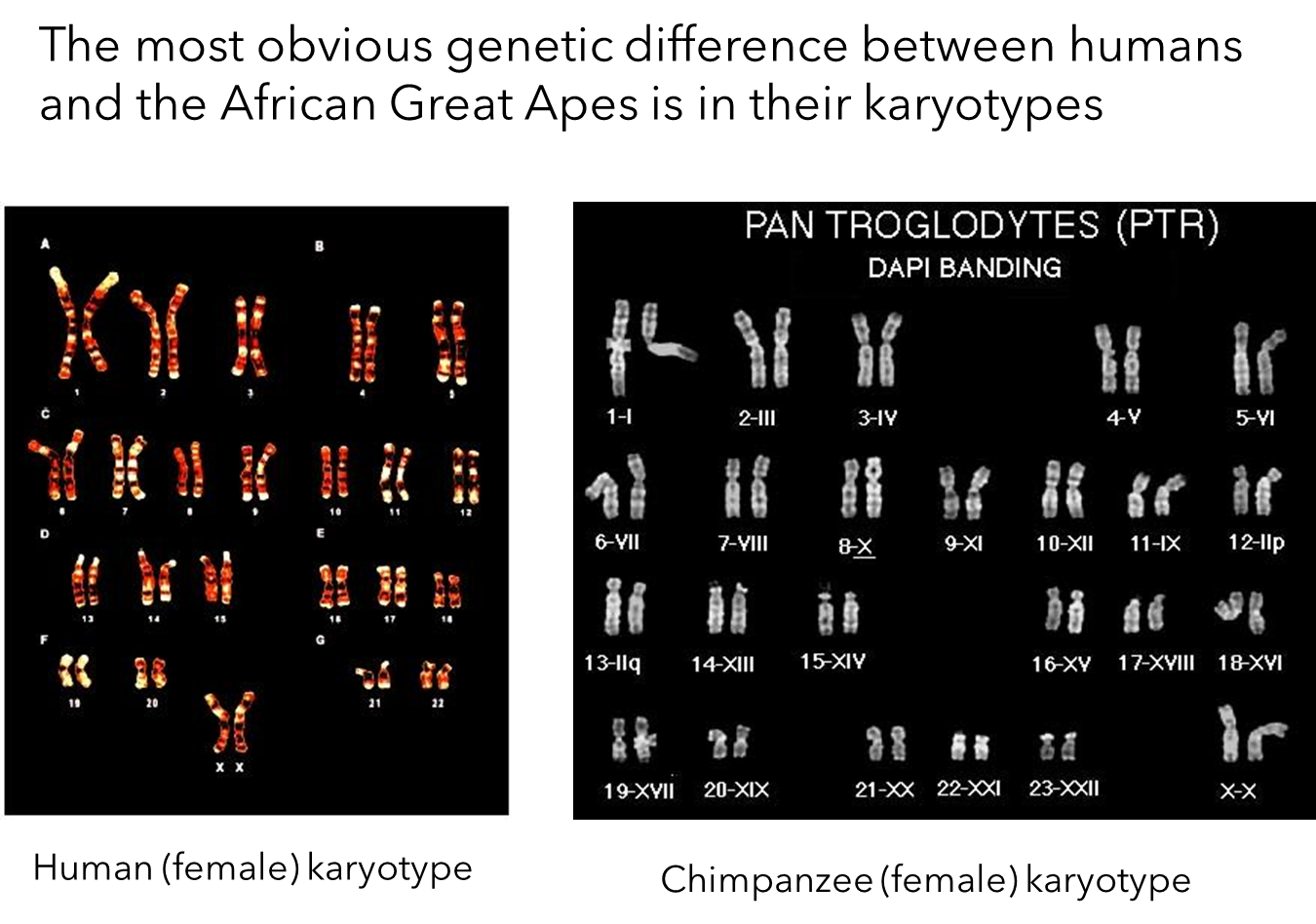
Genetic Differences in Karyotypes
Key Differences
Chromosome Number:
Gorillas and Chimpanzees: Have 24 pairs of chromosomes.
Humans: Have 23 pairs of chromosomes.
Explanation
Chromosome Fusion:
In the ancestors of humans, after the split from chimpanzees, two chromosomes fused to form what is now known as chromosome 2 in humans.
This fusion is the reason for the difference in chromosome numbers between humans and African great apes.
Question
What is the genetic difference in karyotypes between humans and the African great apes?
The genetic difference in karyotypes between humans and the African great apes lies in the number of chromosome pairs. Gorillas and chimpanzees have 24 pairs of chromosomes, while humans have 23 pairs. This difference is due to a chromosomal fusion event in the human lineage after it split from the lineage of chimpanzees. Specifically, two ancestral chromosomes fused to form what is now known as chromosome 2 in humans.

Genetic Comparisons
Human and Chimpanzee Genomes
Genetic Similarity:
Average Genetic Difference: 1.23% (or 98.77% similarity) between comparable genes.
Variation:
Some genes are less similar, some are identical.
Certain genes are unique to humans or chimpanzees and cannot be compared.
Protein-Level Comparison:
Amino Acid Sequences: Only 29% of genes code for the same amino acid sequences in humans and chimpanzees.
Mouse and Human Genomes
Genetic Similarity:
Average Genetic Similarity: 85% among comparable genes.
Variation: Similar to the comparison between humans and chimpanzees, there is a lot of variation from gene to gene.
Unique Genes: Some genes are not common to both species and cannot be compared.
Context
Overall DNA Sequence Comparison:
Humans and chimpanzees are very similar.
Mouse and human genes also show significant similarity, reflecting our common ancestry.
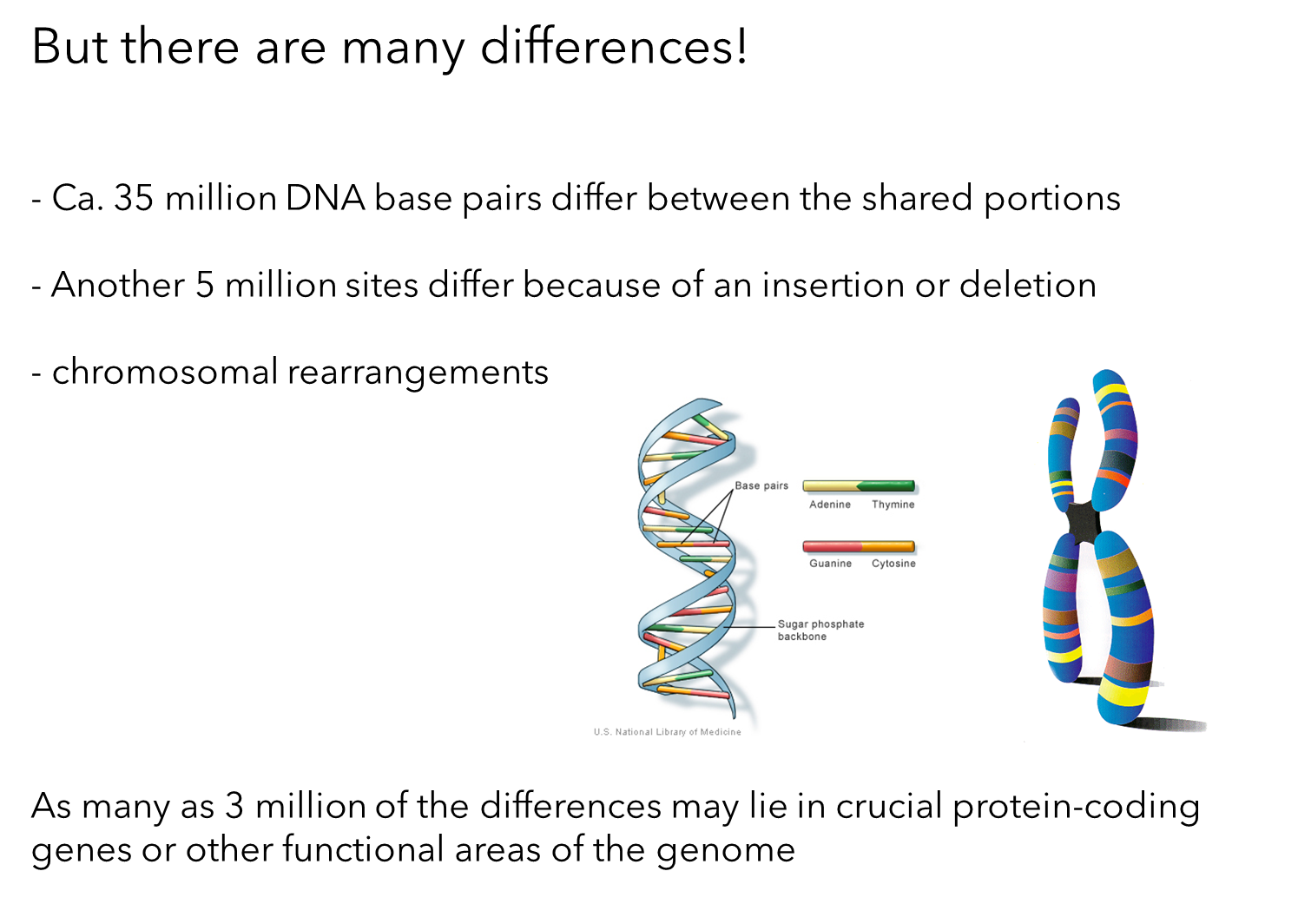
Genetic Differences between Humans and Chimpanzees
Key Differences
Base Pair Differences:
Approximately 35 million DNA base pairs differ between the shared portions of the human and chimpanzee genomes.
Insertion/Deletion Differences:
About 5 million sites differ due to insertions or deletions of DNA sequences in one of the lineages.
Chromosomal Rearrangements:
There are also a smaller number of chromosomal rearrangements.
Functional Implications
Non-Functional DNA:
Most of the differences lie in regions of DNA believed to have little or no functional impact.
Functional Areas:
As many as 3 million differences may be in crucial protein-coding genes or other functional areas of the genome.
Future Research
Emerging Data:
As the genomes of other mammals and primates are sequenced in the coming years, researchers will be able to identify DNA sequence changes specific to the human lineage.

Genetic Changes Related to Human-Specific Traits
Key Traits
Bipedalism: Walking upright on two feet.
Enlarged Brain: Significantly larger brain size compared to other primates.
Complex Tool Use: Ability to manufacture and use sophisticated tools.
Complex Language Skills: Advanced language and speech capabilities.
Research Focus
Gene Evolution:
Researchers are looking for genetic changes linked to these human-specific traits.
Some genes associated with these traits are evolving more rapidly in humans than in chimpanzees.
FOXP2 Gene:
Function: Regulates the activity of other genes and plays a key role in embryonic development.
Evolution:
Studies by Jianzhi Zhang and Svante Pääbo have shown that the FOXP2 gene has evolved at a higher rate since the human-chimpanzee split compared to before.
Role in Language:
Genes regulated by FOXP2 are involved in human language and speech.
Rare mutations in FOXP2 can lead to severe language and speech disorders in humans.
Future Prospects
Genome Sequencing:
With the human genome and those of other animals now sequenced, understanding of human evolution is expected to advance rapidly.

HAR1 (Human Accelerated Region 1)
Function:
HAR1 is a non-coding RNA gene that plays a role in brain development.
Evolution:
HAR1 shows rapid evolution in humans compared to other primates.
Differences in HAR1 are associated with changes in the structure and function of the human brain.
Significance:
HAR1 is thought to be involved in the development of the cerebral cortex, which is crucial for higher cognitive functions.
HACANS2 (Human Accelerated Region 2)
Function:
HACANS2 is another non-coding region that influences brain development.
Evolution:
Like HAR1, HACANS2 has undergone significant evolutionary changes specific to humans.
Significance:
It is involved in regulatory networks that affect brain size and complexity.
FOXP2 (Forkhead Box P2)
Function:
FOXP2 is a transcription factor that regulates the expression of other genes.
It is critical for normal speech and language development.
Evolution:
FOXP2 has evolved rapidly in humans since the human-chimpanzee split.
Mutations in FOXP2 can cause severe language and speech disorders.
Significance:
The gene’s role in controlling vocalization and speech-related behaviors highlights its importance in human communication.
KRTHAPI (Keratin Gene, KRT6A)
Function:
KRTHAPI encodes keratin proteins that are essential for the structural integrity of skin and hair.
Evolution:
The gene has undergone changes in human lineage, affecting skin and hair characteristics.
Significance:
Variations in KRTHAPI can influence skin texture and hair types, which have adapted to different environmental pressures.
Fossils

Fossil Record of Hominids
Definition of Hominids
Hominids: Refers to any species more closely related to humans than to chimpanzees.
Chimpanzees: Considered humans' closest living relatives.
Fossil Discoveries
No chimpanzee fossils have ever been found.
A single fossil skull, Sahelanthropus tchadensis, dating back 6-7 million years, was discovered before the divergence of hominids and chimpanzees.
Discovery of Sahelanthropus tchadensis
Location: Found in the Djurab Desert of Chad.
Date of Discovery: July 2001.
Fossil Composition: Nearly complete cranium.
Characteristics of Sahelanthropus tchadensis
Braincase Size:
Small braincase (320-380 cm³)
Smaller than any known hominid and comparable to that of a chimpanzee.
Facial Structure:
Initially thought to have a relatively flat face, resembling hominids.
Later examination revealed that the front of the face was distorted during fossilization, leading to a superficial appearance of flatness.
Current Understanding
Most scientists now consider Sahelanthropus tchadensis to represent an ape, not a hominid.
The evolution of opinions reflects rapid advancements in research within human paleoanthropology, indicating that the textbook information may be outdated.
Misconceptions about Human Evolution
Common Misconception: "Humans are descended from apes."
Clarification:
Hominids are not direct descendants of any existing monkeys or apes.
Approximately 6-7 million years ago, humans and apes shared a common ancestor (potentially Sahelanthropus tchadensis).
Following this divergence, the ape lineage split into modern gorillas, chimpanzees, and orangutans, while the hominid line evolved separately.
Modern apes are evolutionary relatives, not ancestors, of humans.

True Hominid Fossils and Human Evolution
Importance of Fossils
Direct Evidence: Fossils provide the most direct glimpse into human evolutionary history.
Insight into Evolution: Comparison of available fossil hominids reveals that modern humans belong to a branch of the evolutionary tree that included other hominid species and even other human species.
Comparative Analysis of Fossils
Character Changes: Fossil comparisons offer unparalleled insight into the changes in characteristics within the human evolutionary lineage.
Major Trends:
Cranial Capacity:
There is a clear trend of increasing cranial capacity over time, which is a measure of brain size.
Representative fossil craniums demonstrate this increase.
Evolutionary Pathways
Intermediate Steps:
Modern humans evolved through many intermediate steps from ancestors that exhibited apelike anatomical features.
The overall picture of major evolutionary trends is well-documented and widely accepted among scientists studying the hominid fossil record.
Disagreements Among Scientists
Classification Issues:
There is ongoing disagreement regarding the number of distinct genera and species of hominids that have existed and their relationships to one another.
This disagreement arises from the sparse fossil record, which, while sufficient to document major trends, does not provide enough specimens for clear classification into distinct species and genera.
Challenges in Taxonomy
Sample Size Limitations:
With a limited number of fossils, it is challenging to determine whether variations between two specimens represent variation within a single species or differences between two separate species.
Taxonomist Experience:
The author draws a parallel to their own experiences as a taxonomist working with rare plants, where determining the significance of unique features in specimens is difficult.
Unique features may either indicate a true distinction for a particular species or may not be reliable for differentiating between two species, as they may not be consistent traits across a larger sample.
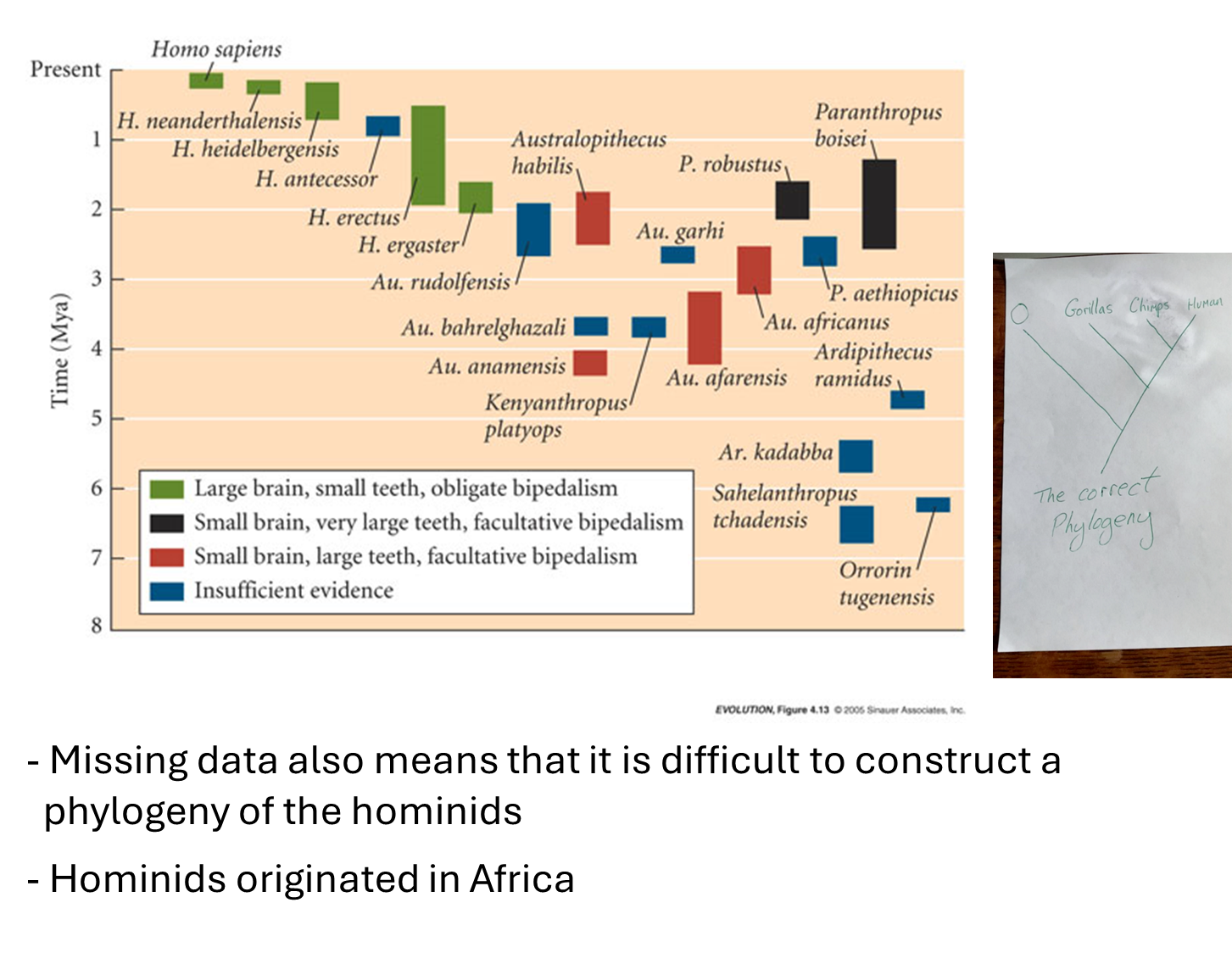
Challenges in Hominid Fossil Analysis
Limited Data Set
Comparative Difficulties: A limited data set makes it challenging to compare species and reconstruct their evolutionary history.
Some species are known from abundant material, including nearly complete skeletons.
Other species are represented only by a few fragments, hindering character comparison.
Temporal Evidence Abundance:
The fossil evidence diminishes as one goes further back in time due to natural geological processes that erode or damage fossils.
Human Factors in Fossil Data
Greed in Paleoanthropology:
Some researchers may attempt to restrict access to newly discovered fossils to gain credit for their work.
This behavior is detrimental to science, as different researchers bring varied skills and perspectives that can enhance understanding.
Negative Impact of Egos:
Personal conflicts and big egos within the field can adversely affect the comprehension of hominid evolution.
Focus on Skull and Tooth Characters
Data Completeness:
Analyses of hominid fossils predominantly rely on skull and tooth characters due to their more complete fossil record compared to other skeletal parts.
Collard and Wood's Analysis (2000)
Study Objective: Mark Collard and Bernard Wood aimed to reconstruct the phylogeny of living apes using a cladistic analysis based on skull and tooth characters similar to those used for hominids.
Methodology: The researchers used the established molecular phylogeny of apes as the baseline for accuracy.
Results:
Their analysis produced a phylogeny indicating that gorillas and orangutans are the closest relatives, followed by chimpanzees, with humans branching off first.
The analysis failed to align with the known molecular phylogeny of the apes, raising concerns about the reliability of skull and tooth character analyses for reconstructing phylogeny.
Cladistic Analysis Context:
This does not discredit cladistic analysis as a method for reconstructing phylogeny generally.
Cladistic analyses of soft tissue characters yielded phylogenies that matched molecular phylogenies accurately.
It suggests that skull and tooth characters may not be reliable for the specific question of hominid evolution.
Graphical Representation of Hominid Fossils
Current Best Approach:
Given the challenges, it may be more effective to categorize hominid fossils into species and depict them against a timeline rather than using a phylogenetic tree.
Recognized Hominids:
Approximately 20 hominid species are recognized, classified into six genera:
Genera:
Orrorin
Ardipithecus
Kenyanthropus
Australopithecus
Paranthropus
Homo (genus of modern humans)
Blue Boxes: Denote species with particularly sparse fossil evidence.
Green Boxes: Represent human species, with only one extant species: Homo sapiens.
Origin of Hominids
African Origin:
The claim that hominids originated in Africa is widely accepted among scientists.
All recognized fossils have been found exclusively in Africa, with only more recent hominids discovered in other parts of the world.
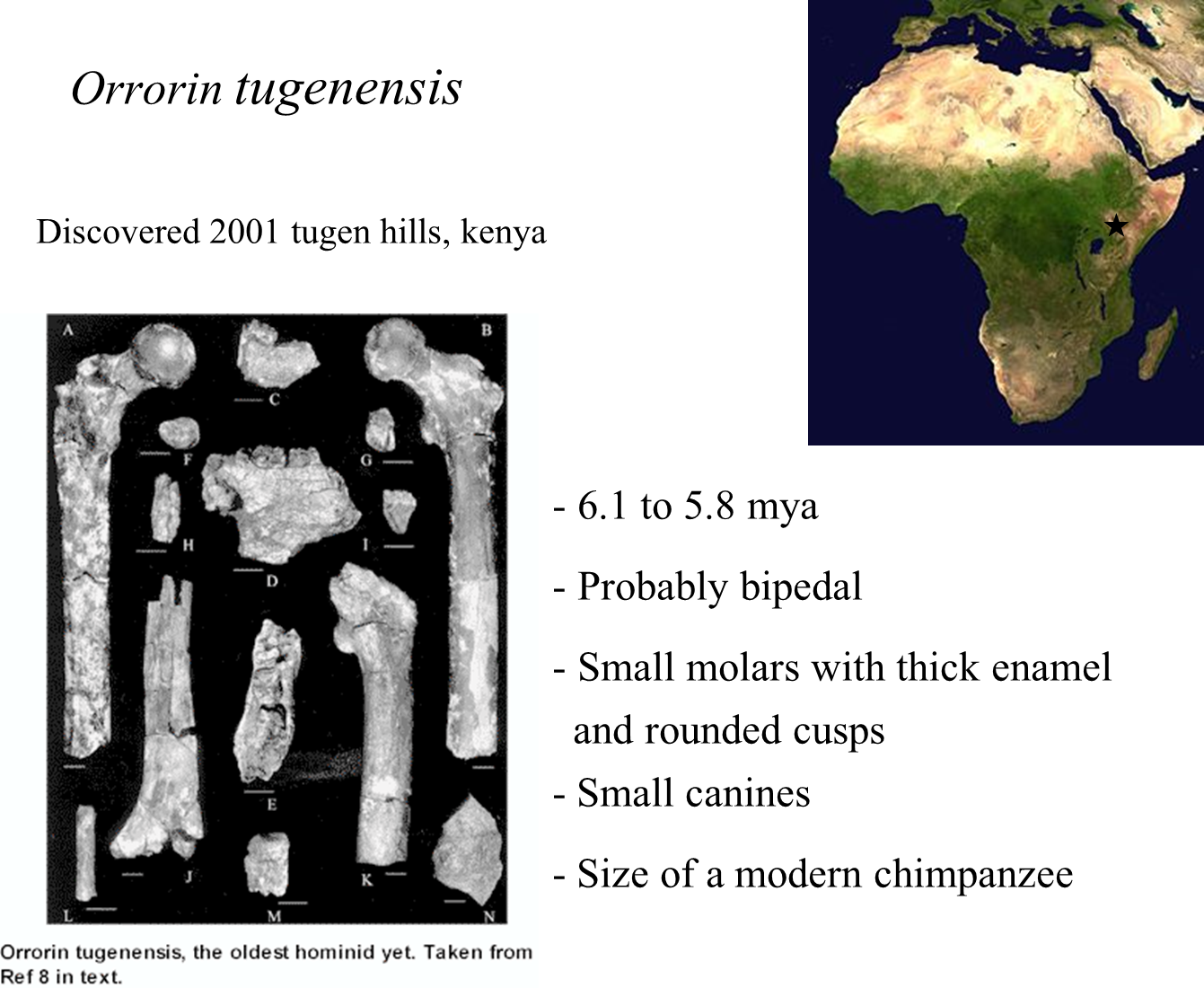
Orrorin tugenensis
Classification and Discovery
Genus: Orrorin
Orrorin tugenensis is the only species classified within this genus.
Fossil Evidence:
Fossils from at least five individuals have been discovered.
Found in the Tugen Hills of Kenya.
Dating and Geological Context
Rock Strata:
The fossils were discovered in rock strata dated between 6.1 and 5.8 million years ago.
Fossil Characteristics
Types of Fossils:
One of the fossils represents a jawbone.
Another fossil is a femur bone.
Bipedalism:
The structure of the femur bone suggests that Orrorin may have walked upright, indicating a probable bipedal habit.
However, the certainty of bipedalism is questioned due to the absence of many key bones typically associated with bipedal locomotion.
Dental and Dietary Characteristics
Teeth Structure:
The fossils feature small molars with rounded cusps and small canines.
These dental characteristics, along with thick enamel, are typical of hominids rather than apes.
Dietary inference suggests that Orrorin primarily consumed fruits and vegetables, with occasional meat.
Physical Characteristics
Size:
Orrorin was approximately the size of a modern chimpanzee.
Habitat and Environmental Context
Fossil Context:
Fossil leaves and other mammals found alongside Orrorin suggest it lived in a dry evergreen forest environment.
This habitat is surprising since it contrasts with the previous belief that bipedalism evolved in open savanna habitats.
Locomotion Insights
Movement:
The movement of Orrorin may have been similar to that of modern orangutans.
Orangutans exhibit bipedal movement over small, springy branches, maintaining a vertically oriented vertebral column while extending their arms for balance and keeping their legs straight.
This form of upright locomotion could have served as a means of navigation on the ground when gaps occurred in the forest canopy.
Future Research Needs
Fossil Evidence:
A better understanding of Orrorin's locomotion and behavior is necessary, highlighting the need for additional and more complete fossil discoveries.
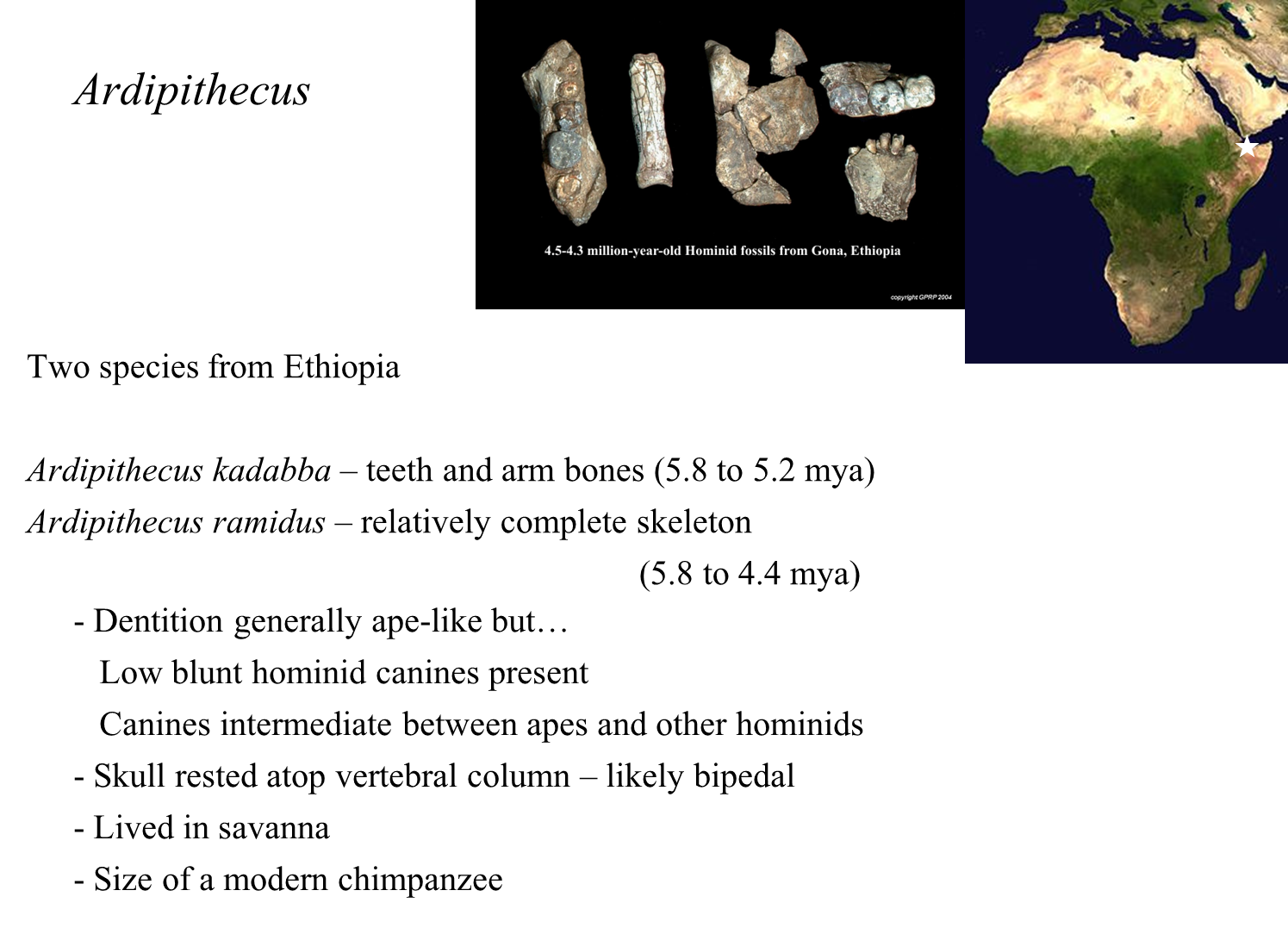
Ardipithecus
Overview
Genus Classification: The next oldest fossil hominids belong to the genus Ardipithecus.
Recognized Species: Two species are recognized within this genus; one is much better represented by fossils than the other.
Fossil Representation
Fossil Discovery: Both species were collected in the 1990s in Ethiopia.
Teeth Preservation: One species is primarily known from teeth, as the enamel on teeth is harder than other body parts, making it more likely to fossilize.
Evolutionary Significance
Primitive and Advanced Traits:
Ardipithecus exhibits a unique blend of primitive ape-like features and more advanced hominid characteristics.
The age and traits of Ardipithecus position it near the base of the hominid evolutionary tree.
Dental Characteristics
Teeth Structure:
The teeth show ape-like features, with enamel that is thin for a hominid but slightly thicker than that found in apes.
Molar shapes are also ape-like, with very asymmetric crowns.
Canines are short, blunt, and resemble those of slightly later early hominids.
Skull and Posture
Skull Positioning:
Fragments of the skull base indicate that the skull rested atop the vertebral column, a feature present in hominids but not in apes.
This positioning is crucial for facilitating prolonged bipedalism.
Bipedalism Evidence
Toe Morphology:
The toe structure suggests a bipedal habit, reinforcing the notion that Ardipithecus was capable of upright walking.
Habitat
Ecological Context:
Associated plant and animal fossils suggest that Ardipithecus species were forest dwellers.
Both species were approximately the size of modern-day chimpanzees.
A. ramidus: Discovery and Characteristics
Fossil Discovery
Announcement: On October 1, 2009, paleontologists announced the discovery of a relatively complete A. ramidus fossil skeleton, initially unearthed in 1994.
Nickname: The fossil is nicknamed "Ardi".
Fossil Composition
Fossil Features:
The remains include most of the skull, teeth, pelvis, hands, and feet.
The fossil represents a small-brained female weighing approximately 50 kilograms (110 lbs).
Location and Dating
Discovery Site: Found in Ethiopia's Afar desert at a site called Aramis in the Middle Awash region.
Radiometric Dating:
Radiometric dating of volcanic ash layers suggests Ardi lived about 4.4 million years ago.
Some researchers, including Beagle and Kappelman, argue for a revised date of 3.9 million years due to difficulties in radiometric dating of the region.
Evolutionary Insights
Bipedalism:
The form of Ardi's pelvis and limbs, along with the presence of an abductable hallux (big toe), indicates she was a facultative biped—bipedal on the ground but quadrupedal in trees.
This suggests a more primitive walking ability compared to later hominids, with limitations on walking or running long distances.
Dietary Implications
Teeth Characteristics:
The teeth of A. ramidus suggest an omnivorous diet and are more generalized than those of modern apes.

Australopithecus
Overview
Genus Classification: Australopithecus is an early hominid genus that appears slightly later in the fossil record and exhibits more advanced characteristics compared to earlier hominids, though it remains close to the base of the hominid evolutionary tree.
Species Recognition: Approximately seven species within the genus Australopithecus are recognized by paleontologists.
Fossil Preservation: Most species are known from several well-preserved fossils, which provide a clearer understanding of their morphology and behavior.
Geographical Distribution: All recognized Australopithecus species have been found exclusively in Africa, highlighting the continent's significance in the evolutionary history of humans.
Notable Fossil: Lucy
Fossil Identification: The most famous fossil of this genus is an almost complete skeleton of a female known as Australopithecus afarensis, nicknamed Lucy.
Discovery Significance: Lucy's discovery was crucial for understanding the evolution of early hominids and has become iconic in paleoanthropology.
Physical Characteristics of Australopithecus afarensis
Primitive Traits:
Facial Structure: Early Australopithecus species, including A. afarensis, exhibit a lower face that projects beyond the eyes, similar to modern apes. In contrast, more advanced hominids, such as modern humans, have flatter faces.
Canine Size: Lucy has relatively large canines—larger than those of modern humans but smaller than those of apes, indicating a transitional morphology.
Proportional Limbs: The arm length of Lucy is relatively long compared to her legs, resembling the proportions seen in apes, suggesting adaptations for climbing.
Brain Size: Lucy possessed a smaller brain compared to later hominids, reflecting the earlier stage of hominid evolution.
Bipedal Adaptations:
Pelvis and Leg Structure: Despite retaining some primitive traits, the structure of Lucy's pelvis and legs clearly indicate bipedal locomotion, suggesting she walked upright.
Curved Finger and Toe Bones: The curved bones in Lucy's fingers and toes imply she engaged in tree climbing, pointing to a dual adaptation for both arboreal and terrestrial lifestyles.

Evolutionary Context
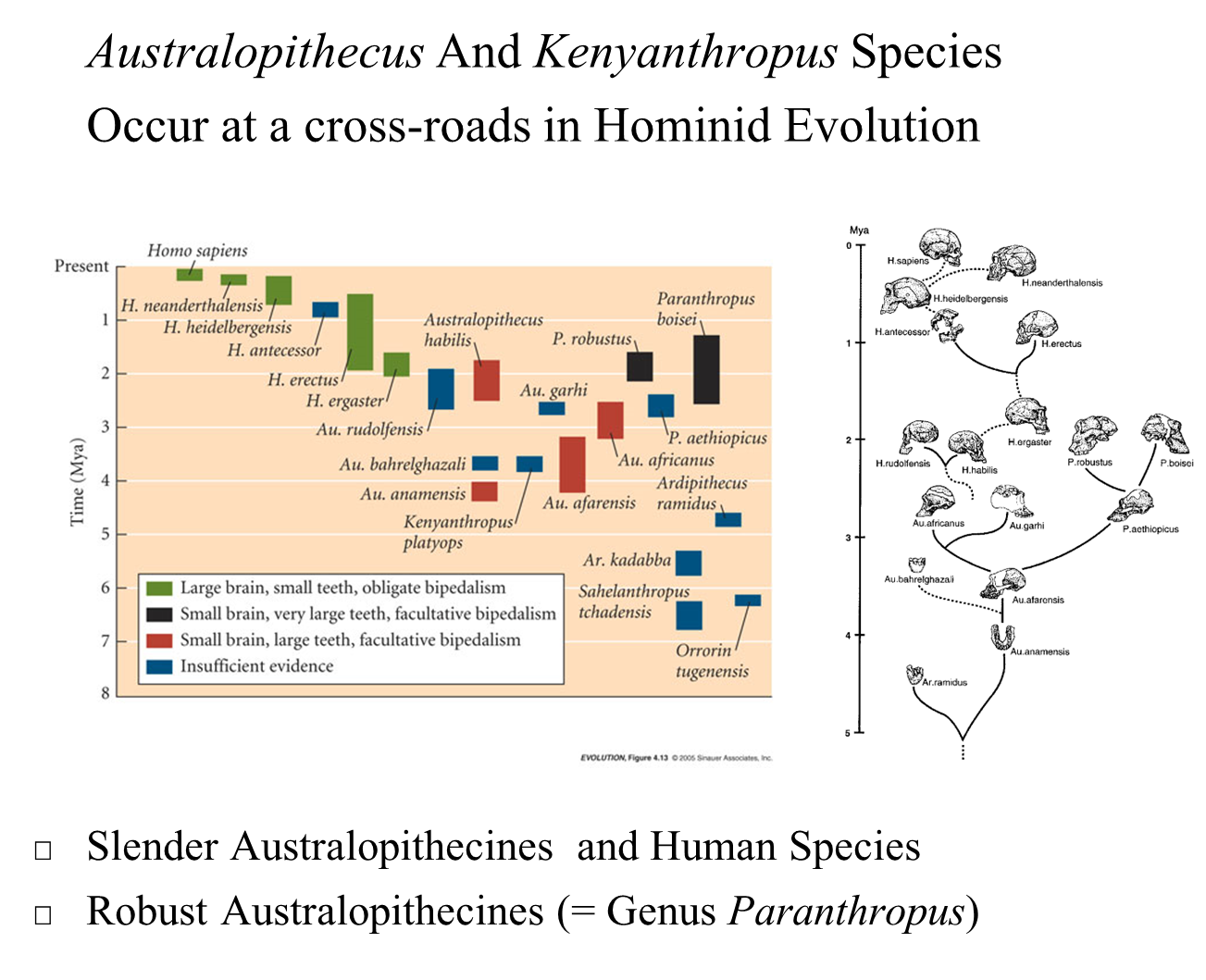
Crossroads of Hominid Evolution: The genus Australopithecus, along with Kenyanthropus, occupies a crucial position in the evolutionary history of hominids. They represent an evolutionary stage where several key adaptations are evident, bridging the gap between earlier hominids like Orrorin and Ardipithecus and later species that are more closely related to modern humans.
Characterization
Early Features: The first Australopithecus species and Kenyanthropus display characteristics that are only slightly more advanced than those found in Orrorin and Ardipithecus. These include adaptations for bipedalism and a mix of primitive and derived traits.
Later Species Development: Some of the later Australopithecus species show morphological features that approach those of the early members of the genus Homo, indicating a significant evolutionary transition.
Lineage Split
Post-Australopithecus afarensis Divergence: A significant evolutionary split occurred after Australopithecus afarensis. This split led to two distinct lineages:
Slender Australopithecines: This lineage eventually gave rise to the human species, characterized by traits that support bipedalism and other adaptations conducive to life on the ground.
Robust Australopithecines: This lineage includes species classified in the genus Paranthropus. These robust forms exhibited adaptations such as larger teeth and strong jaw structures, likely associated with a diet that required processing tough plant materials.
Intermediate Species
Transition Between Genera:
Three species—Homo ergaster, Homo rudolfensis, and Homo habilis—are considered intermediates between the genera Australopithecus and Homo. These species exhibit a blend of traits from both genera and have been variously classified in either group throughout research history.
Fossil Comparisons:
Older Fossils: Fossils of Homo habilis are often compared to Australopithecus africanus, sharing several morphological characteristics.
Later Fossils: The more recent fossils from Homo ergaster and Homo rudolfensis show a gradual evolution toward traits seen in Homo erectus, indicating an evolutionary trend toward more advanced hominid features.
Morphological Traits and Adaptations
Bipedalism: Both Australopithecus and Kenyanthropus exhibit adaptations for bipedal locomotion, which is a defining characteristic of hominids. However, their bipedalism is still considered primitive compared to that of later hominids.
Dentition and Diet: The dental morphology of these species indicates an omnivorous diet, which likely included a variety of fruits, vegetables, and possibly meat, reflecting a significant adaptability to different ecological niches.
Conclusion
The Australopithecus and Kenyanthropus genera mark an important juncture in the evolution of hominids. They not only show the transitional traits that bridge the gap between earlier hominids and the genus Homo, but they also highlight the complexity of human evolution, including the divergence of lineages leading to modern humans and robust forms like Paranthropus. Continued research and fossil discoveries in this area are essential for understanding the nuanced paths of human ancestry.
Evolutionary Context
Crossroads in Hominid Evolution:
Both Australopithecus and Kenyanthropus represent a significant point in the evolutionary tree of hominids.
They exhibit a combination of primitive and advanced traits, positioning them closer to modern humans compared to earlier hominids like Orrorin and Ardipithecus.
Characteristics of Australopithecus and Kenyanthropus
Advanced Yet Primitive Traits:
The initial Australopithecus species and Kenyanthropus show anatomical features that are only slightly more advanced than those of Orrorin and Ardipithecus.
However, certain later species within these genera begin to display characteristics approaching those found in early members of the genus Homo.
Evolutionary Split After Australopithecus afarensis
Lineage Divergence:
After Australopithecus afarensis, a significant evolutionary split occurred in the hominid lineage:
Slender Australopithecines:
This lineage eventually leads to the development of the human species, characterized by adaptations favoring bipedalism and other traits supportive of a terrestrial lifestyle.
Robust Australopithecines:
This lineage includes species classified in the genus Paranthropus, which exhibit adaptations suited for a diet requiring robust dental structures.
Intermediate Species Between Genera
Transitional Forms:
Three species—Homo ergaster, Homo rudolfensis, and Homo habilis—are viewed as intermediate between Australopithecus and Homo.
These species have characteristics that bridge the anatomical and functional gaps between the two genera and have been classified within both groups at various times.
Morphological Comparisons
Fossil Relationships:
Older Fossils:
The fossils of Homo habilis are often compared to Australopithecus africanus, sharing several morphological traits, including aspects of dental structure and cranial features.
Later Fossils:
More recent fossils from Homo ergaster and Homo rudolfensis show progressive traits that align more closely with Homo erectus, indicating an evolutionary trend toward more modern hominid features.
Key Takeaways
Significance in Hominid Evolution:
The study of Australopithecus and Kenyanthropus offers critical insights into the evolutionary history of hominids, illustrating how early species exhibited a mix of adaptations that facilitated both arboreal and terrestrial lifestyles.
Understanding the evolutionary split and the characteristics of intermediate species enhances our comprehension of human origins and the complex paths taken in hominid evolution.

Genus: Homo
Species: Homo ergaster
Time Period: Lived throughout eastern and southern Africa between 1.9 to 1.4 million years ago.
General Trends:
Cranial capacity serves as a measure of brain size and has shown changes as subsequent Homo species evolved.

Species: Homo heidelbergensis
Relation to Modern Humans: Closely related to Homo sapiens.
Key Characteristics:
Use of Fire: H. heidelbergensis was the first species to use fire.
Hunting Tools:
The first to use stone-tipped wooden spears for cooperative hunting.
Shelter Construction:
The first to build shelters.
Fossil Evidence:
Fossils of H. heidelbergensis merge into the earliest fossils of Homo sapiens.
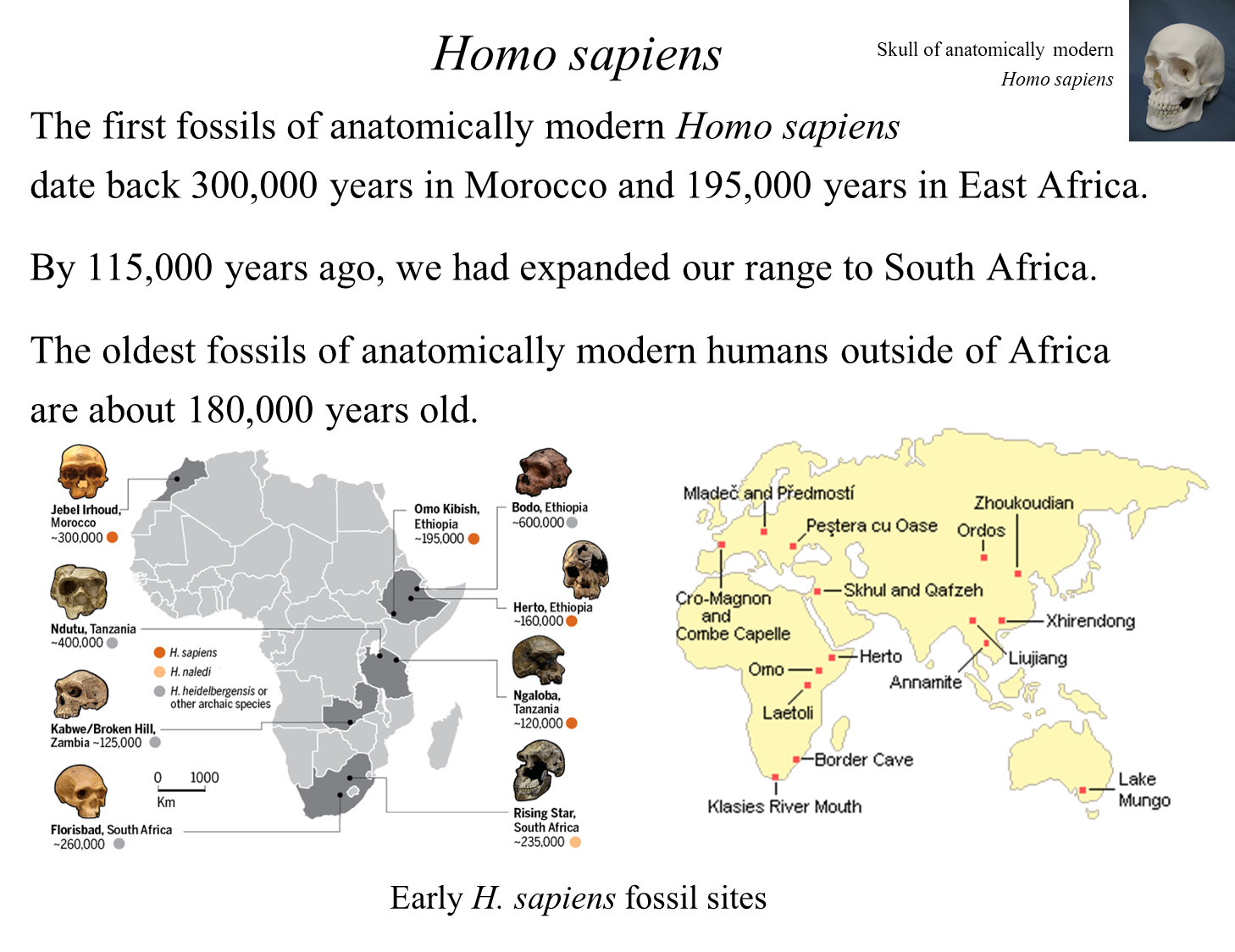
Time Period: Approximately 200,000 years ago during a period of dramatic climate change.
Location: Originated in Africa.
Survival Behaviors:
Engaged in gathering and hunting for food.
Developed behaviors to respond to challenges of survival in unstable environments.
Anatomical Characteristics:
Anatomically Modern Humans: Refers to early fossils exhibiting skeletal characteristics similar to those of modern humans.
Brain Size:
Relatively larger brains compared to Homo heidelbergensis and transitional fossils.
Brain housing required skull reorganization.
Skull Features:
Thin-walled, high vaulted skull.
Flat and near-vertical forehead.
Reduced heavy brow ridges compared to earlier human species.
Jaw Structure:
Less heavily developed jaws.
Smaller teeth.
Projecting chin.
Skeletal Build:
Lighter skeletal build compared to earlier humans.
Fossil Evidence:
Key Fossil: A 195,000-year-old fossil from Ethiopia.
Represents early changes in skull structure associated with modern humans, including:
Rounded skull case.
Projecting chin.
Expansion of Range:
By 115,000 years ago, early modern humans had spread to South Africa.
By 180,000 years ago, migrated to Israel in Southwest Asia.
Lack of Evidence Elsewhere:
No reliable evidence of modern humans in other regions of the Old World until 60,000-40,000 years ago, coinciding with a temperate period during the last ice age.

Evolutionary Background:
Homo neanderthalensis evolved from a common ancestor shared with Homo sapiens.
The "th" in "Neanderthal" is pronounced as a "t".
DNA comparisons indicate that the evolutionary split between Neanderthals and Homo sapiens occurred between 400,000 and 500,000 years ago.
Both species evolved from Homo heidelbergensis.
Physical Characteristics:
Fossil Evidence:
Numerous fossils and preserved bones of Neanderthals have been discovered.
DNA has been extracted from some preserved bones and compared with that of Homo sapiens.
Skeletal Features:
Neanderthals had denser bones.
Thicker skulls with projecting brow ridges.
Distinct bump on the back of the skull.
Overall physical similarities to modern humans.
Interbreeding:
Neanderthals and Homo sapiens were similar enough anatomically to interbreed.
DNA evidence suggests that interbreeding occurred, but it was not a frequent occurrence.
Behavioral and Cultural Aspects:
Fire Control: Neanderthals demonstrated the ability to control fire.
Burial Practices:
They buried their dead, indicating an understanding of ritual.
Some graves were adorned with flowers, suggesting early forms of ritualized burial.
Tool Use:
Manufactured and utilized complex stone tools.
Clothing: Evidence suggests they wore clothing.
Coexistence with Neanderthals:
Homo sapiens coexisted with Neanderthals, a different human species, in parts of Europe for several thousand years.
Neanderthals went extinct approximately 30,000 years ago.
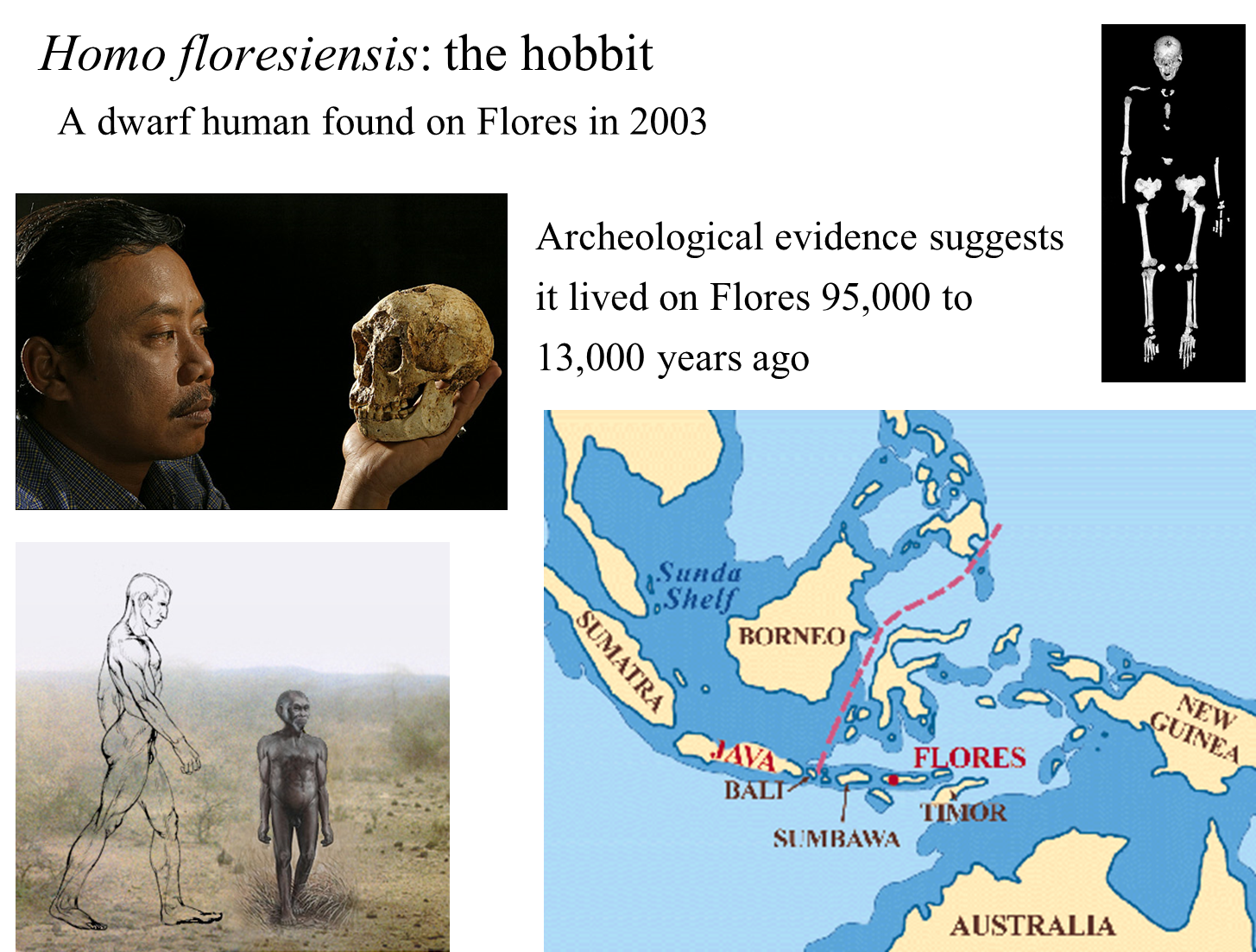
Discovery and Characteristics of Homo floresiensis:
Discovery:
Fossils of H. floresiensis were first discovered in 2003 in a cave on the island of Flores, Indonesia.
The fossils represent six individuals and were classified as a new species of human.
Physical Characteristics:
H. floresiensis is unique in stature, measuring about 3 feet 6 inches in height.
It has a very small brain, earning it the nickname "the hobbit."
The skull features a receding forehead and lacks a chin.
Distinctive wrist bones further differentiate it from other hominids.
Archaeological Evidence:
Evidence indicates that H. floresiensis used stone tools and controlled fire.
They hunted now-extinct pygmy elephants and giant rats.
Dating of Fossils:
Fossils date from approximately 38,000 to 18,000 years ago.
Archaeological evidence suggests that this species lived on Flores from at least 95,000 to 13,000 years ago, indicating their recent presence.
Phylogenetic Placement:
The exact placement of H. floresiensis in the human phylogeny remains uncertain.
It may have descended from Homo erectus, which is the ancestor of Homo heidelbergensis, possibly representing an ancient human lineage.
Cultural Folklore:
On the island of Flores, local folklore speaks of an ancient small human-like creature called Ebu gogo.
When the Portuguese first visited Flores in the 17th century, Ebu gogo was thought to still exist, but there have been no reported sightings since then.
Taxonomic Issues:
Many generic and species-level taxonomic issues in human evolution remain unresolved.
Evolution of Modern Humans:
The overarching narrative that modern humans evolved gradually from ape-like ancestors is well-established.
Brain Size Evolution:
The increase in brain size did not occur uniformly along the hominid lineage.
There were periods when changes in brain size occurred more rapidly than during other times.
Evolution of Bipedalism:
Bipedalism is suggested to have evolved to free the arms for carrying food back to social units, particularly to an individual's mate and offspring.
Selective Pressures for Brain Size:
The increase in brain size may have been selected for due to its advantages in tool use.
Social interactions, such as forming cooperative and mutually beneficial bonds and detecting cheaters in social exchanges, may also have been significant selective forces.
Evolution of Teeth:
Smaller teeth of a more even size evolved in response to a changing diet.
Ancient Human Migration and our modern diversity

Geographic Distribution:
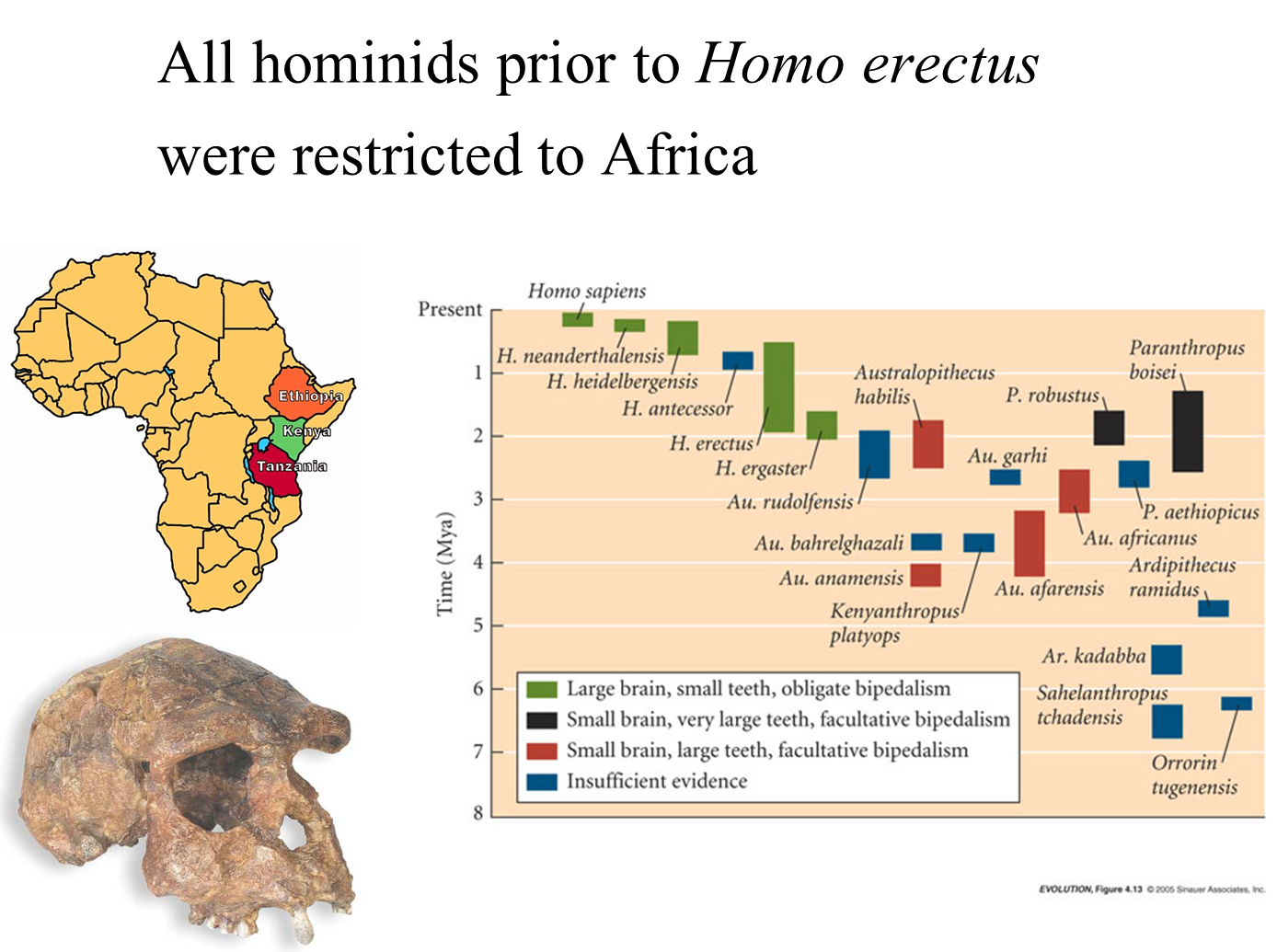
All hominoid species up to Homo erectus were primarily restricted to Africa.
The center of diversity for many African hominoid species was located in the African Rift Valley area, encompassing present-day Ethiopia, Kenya, and Tanzania.
Environmental Conditions:
During the time of these early hominoids, the African Rift Valley region was wetter than it is today.
This area likely had a greater forest cover, providing diverse habitats for early hominins.
Homo Erectus:
The skull depicted is that of Homo erectus, which signifies a notable evolutionary advancement in hominid development.
First Homo Species Outside Africa:
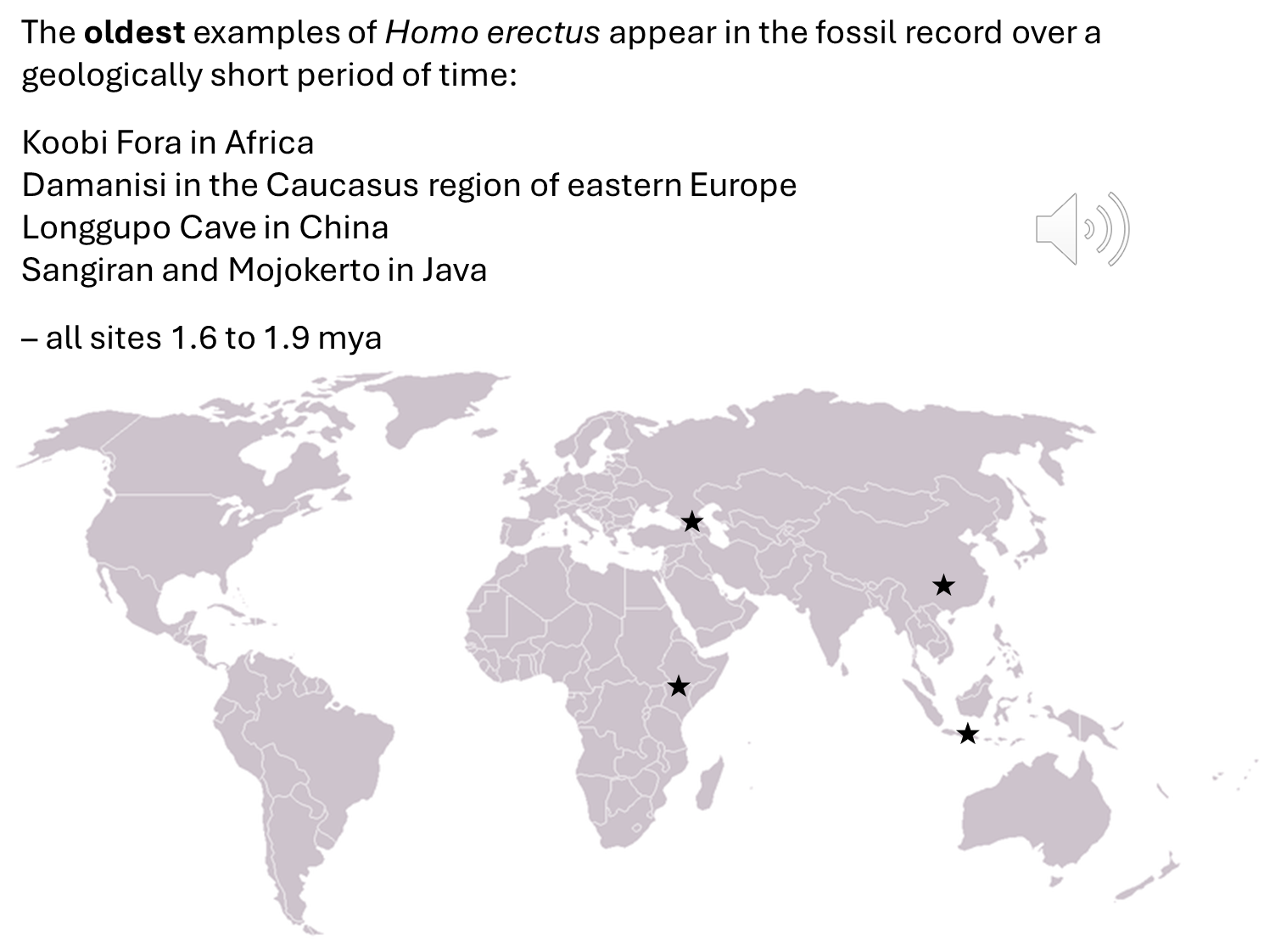
Homo erectus is recognized as the first Homo species to appear in the fossil record outside of Africa.
Fossil Distribution:
The oldest fossils of Homo erectus emerged almost simultaneously in three regions:
Africa
Europe
Asia
Timeline:
The appearance of these fossils occurred over a short time frame, approximately 1.9 to 1.6 million years ago.
Significant Archaeological Sites:
The five earliest sites where Homo erectus fossils have been found include:
Koobi Fora in Africa
Dmanisi in the Caucasus region of Eastern Europe
Longgupo Cave in China
Sangiran in Java
Mojokerto in Java
Evolutionary Significance:
Homo erectus is considered a key ancestor in the evolutionary line leading to Homo sapiens.
Evolutionary Lineage:
Homo erectus is considered a direct ancestor of Homo sapiens.
Homo heidelbergensis and Homo neanderthalensis are often included in the classification of archaic Homo sapiens.
Some scientists view Homo heidelbergensis and Homo neanderthalensis as evolutionary stages between Homo erectus and anatomically modern Homo sapiens.
Others propose that Homo neanderthalensis represents a side branch in the hominid lineage, not a direct ancestor of Homo sapiens.
Geographic Distribution:
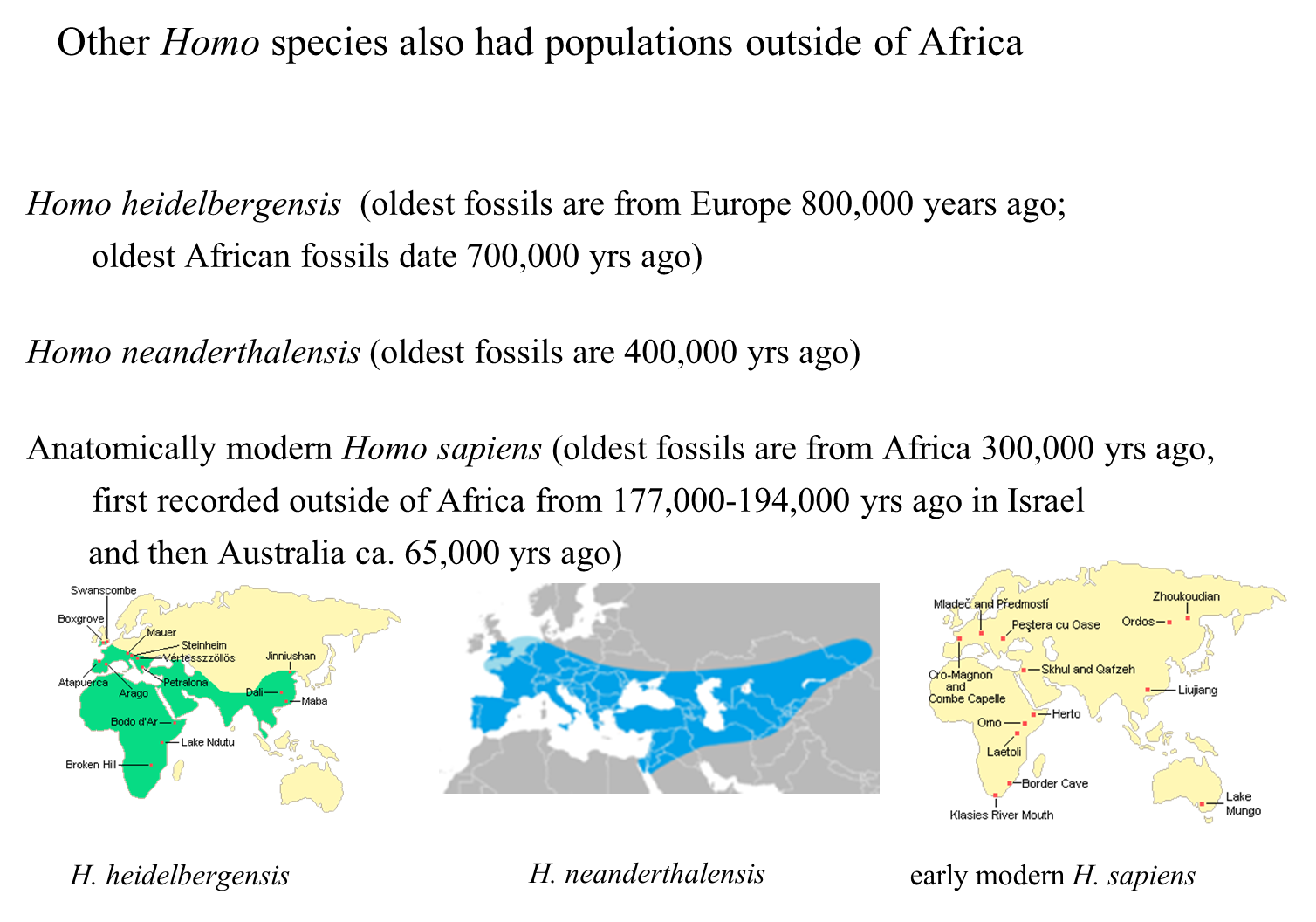
All three species—Homo erectus, Homo heidelbergensis, and Homo neanderthalensis—had populations outside of Africa.
Homo heidelbergensis:
Found in both Africa and Europe/Asia.
Homo neanderthalensis:
Dispersed from Israel to Western Asia and Southern England.
Homo sapiens Distribution:
The earliest anatomically modern Homo sapiens fossils date back to approximately 130,000 years ago and were found in Africa.
The first population of anatomically modern humans to migrate outside of Africa is evidenced in Israel, dating to around 100,000 years ago.
The next oldest fossils of Homo sapiens were discovered in Australia, dating to about 42,000 years ago.
Controversy in Origins
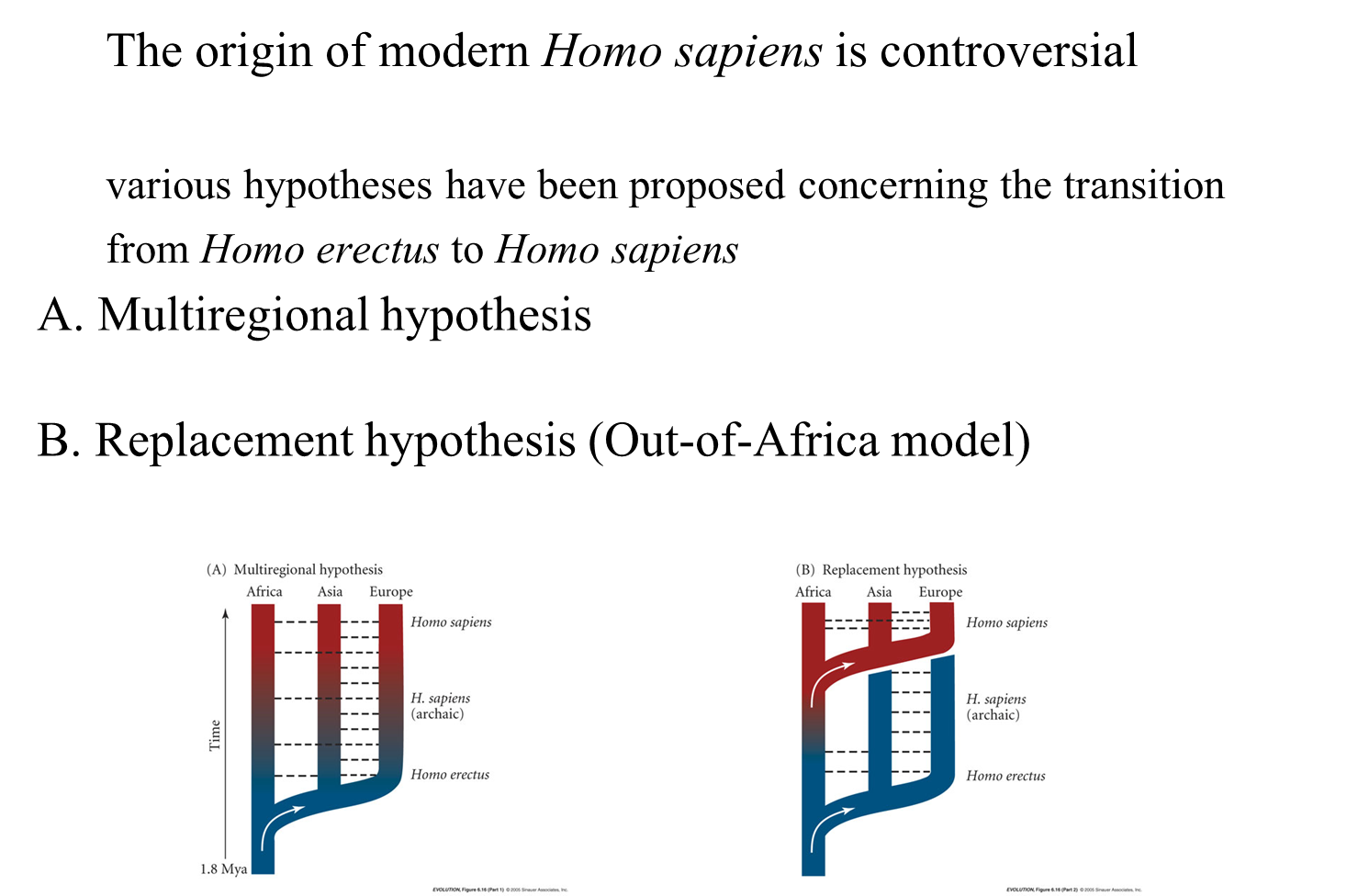
The origin of modern Homo sapiens is a subject of debate, with various hypotheses proposed regarding the transition from Homo erectus to Homo sapiens.
Main Hypotheses
Out of Africa Model (also known as the replacement hypothesis, single origin hypothesis, or Recent African Origins Model):
Timeframe: Suggests that anatomically modern humans originated in Africa between 200,000 and 130,000 years ago.
Migration: A branch of this population left Africa around 60,000 years ago, spreading across the globe.
Replacement: These emigrants replaced other Homo species already present in other regions, such as Neanderthals and Homo erectus.
Multiregional Hypothesis:
Origin: Proposes that anatomically modern Homo sapiens emerged around 1.25 million years ago from four separate lineages of Homo erectus:
One lineage in Africa.
One lineage in Europe.
One lineage in Asia.
One lineage in Australia.
Hybridization: Suggests there was significant hybridization between these four lineages, which maintained them as a single species.
Local Evolution: Some modern human populations, according to this model, are directly derived from local archaic humans, such as Homo neanderthalensis and Homo heidelbergensis.
Implications of the Models
Geographical Races:
Out of Africa Model: Present-day racial variation is seen as the result of recent geographic differentiation within the last 100,000 to 200,000 years after anatomically modern humans left Africa.
Multiregional Hypothesis: Present-day racial differences are attributed to a combination of recent and ancient geographic differentiation, starting as early as 1.25 million years ago.
Conclusion
The majority of scientists currently support the Out of Africa Model due to the diverse evidence that aligns more closely with this hypothesis. Further slides will present evidence supporting both models and explain the prevailing scientific consensus.
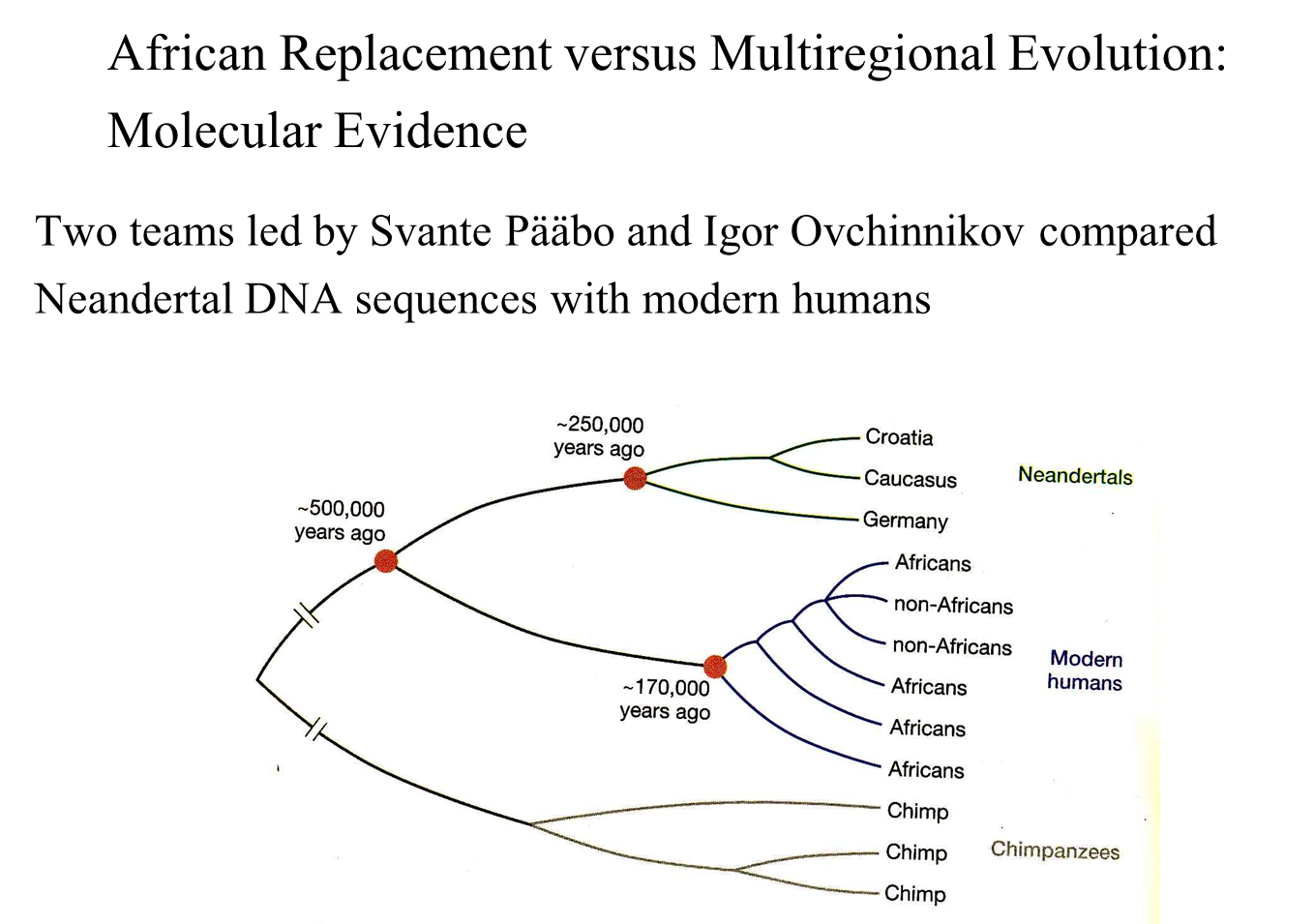
Overview of Sequencing Studies
Two teams have sequenced genes from ancient Neanderthal skeletons collected from:
Croatia
Caucasus
Germany
The genetic sequences from these Neanderthals were compared with those of modern humans from various regions:
Europe
Africa
Asia
America
Australia
Oceania
Key Findings
Genetic Relationship:
Modern humans from all regions are more closely related to each other than to the three Neanderthals.
The three Neanderthals are more closely related to each other than to any modern humans.
Sequence Differences:
On average, the three Neanderthals exhibit three times as much genetic sequence difference from modern humans than is found among modern human populations.
Common Ancestors:
The last common ancestor of modern humans is estimated to have lived 170,000 years ago, consistent with fossil evidence.
The last common ancestor of Neanderthals is estimated to have lived 250,000 years ago, aligning with Neanderthal fossil data.
Modern humans and Neanderthals share a last common ancestor dating back 500,000 years.
Implications of Findings
The genetic data suggest that modern humans and Neanderthals are distinct species, which aligns with the Out of Africa Model:
This model posits that modern humans evolved as a distinct lineage separate from Neanderthals and supplanted them without significant interbreeding.
Limitations of the Study
The conclusions are based on only three Neanderthal individuals.
There is a significant challenge in extracting usable DNA from ancient specimens like Neanderthals, which may limit the comprehensiveness and reliability of the findings.
Context of Cladistic Analysis
Cladistic analysis of modern human gene sequences is utilized to test the Out of Africa Model and the Multiregional Hypothesis.
DNA extraction and sequencing from living humans is straightforward, allowing for comparative studies.
Challenges in Testing Theories
Several studies have been conducted; however, both models are similar genetically, leading to difficulties in differentiation:
Both models describe a species originating in Africa.
Both models depict a spread through Europe and Asia.
Both models highlight the differentiation into regionally distinct populations while maintaining gene flow.
Key Difference Between Models
The primary distinction lies in the timeline of population divergence:
Multiregional Model: Suggests human differentiation began 1.8 million years ago.
Out of Africa Model: Proposes this process began around 200,000 years ago.
Expected Differences
The expected differences between the two models are quantitative rather than qualitative:
Predicted variations in gene frequencies instead of distinct genetic differences.

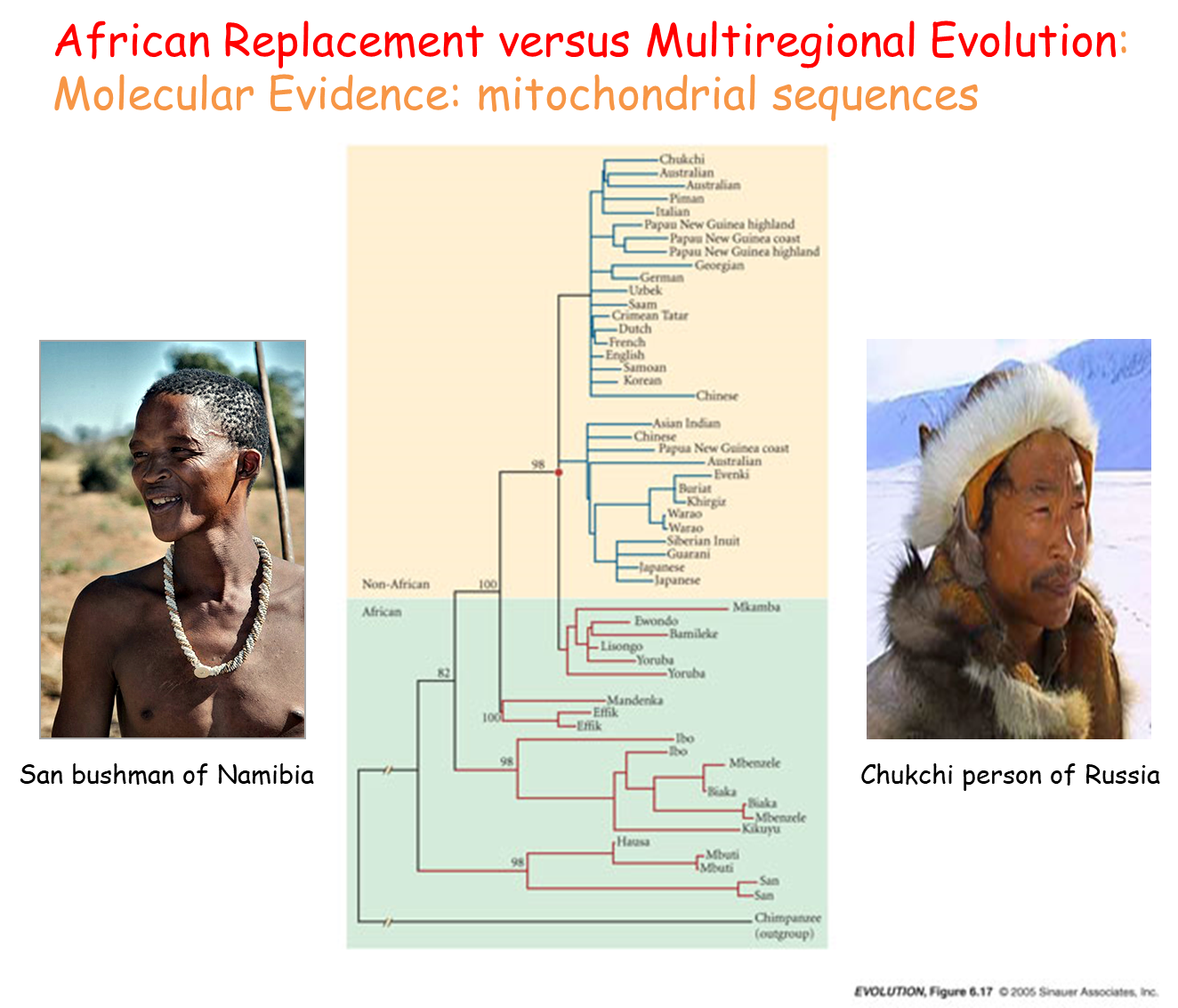
Overview of the Evolutionary Tree
The tree is constructed from complete mitochondrial DNA sequences of 53 modern humans.
Each branch tip represents an individual identified by their race.
The tree is rooted using chimpanzee DNA.
Numbers on branches indicate branch support, with higher numbers indicating better statistical support for that part of the tree.
The tree shows strong statistical support.
Key Findings
Branching Pattern:
All non-African races branch from within African races.
African races occupy a basal position in the tree, supporting the idea that human lineage evolved in Africa before colonizing other regions.
This aligns with predictions from both models and fossil data.
Evidence Supporting the African Replacement Model
Genetic Diversity:
Current human genetic diversity is greater in Africa than in other regions.
This supports the notion that humans lived in Africa for an extended period, accumulating mutations, before a single lineage migrated out to populate the rest of the world.
Under the Multiregional Hypothesis, genetic diversity would be expected to be roughly equal across all regions due to the longer time frame of human existence outside of Africa.
Genetic Differences Between Populations:
Overall genetic differences among human populations outside of Africa are not very large.
This suggests that Europe and Asia were colonized by a small number of individuals in the relatively recent past, leading to limited time for mutations to accumulate.
Dating the Last Common Ancestor
Using the number of sequence changes, the last common ancestor of both African and non-African populations is dated to approximately 52,000 years ago.
This date aligns with archaeological and paleontological evidence, providing strong support for the African Replacement Model.
Limitations of Current Analysis
The segment of the tree relating to non-African populations shows minimal sequence change, offering little information about their interrelations.
To investigate more recent migration patterns, more variable genetic techniques are necessary.
Overview of the Africa Replacement Hypothesis
Proposes that Homo sapiens migrated out of Africa and replaced existing species without breeding with them.
Recent evidence suggests that there was limited interbreeding between Homo sapiens and Neanderthals where their distributions overlapped.
Evidence of Interbreeding
Trinkaus's Findings:
Identified mixed Neanderthal-modern human characteristics in two specimens (~30,000 years old):
One specimen from France.
One specimen from Portugal (almost complete skeleton of a 4-year-old boy).
Characteristics:
Prominent chin and small lower arms typical of anatomically modern humans.
Large jaw, stocky body, short legs, and a bump on the back of the skull characteristic of Neanderthals.
However, the evidence from only two fossils is limited.
Genome Comparison Breakthrough
Paabo's Research Group:
Analyzed the complete genomes of Neanderthals and modern humans to compare allele frequencies.
Hypothesis: If Neanderthals share more alleles with humans descended from populations that overlapped with them (Europe/Asia) than with those from Africa, this supports the interbreeding hypothesis.
Findings:
Non-African humans share 1-4% of their genome with Neanderthals, indicating limited interbreeding.
This genetic contribution is thought to have occurred in the Near East within the last 100,000 years.
Insights into Neanderthal Appearance
Molecular data has improved the understanding of Neanderthal physical characteristics.
Although no fossilized soft tissue remains have been found, genomic analysis has provided clues about their appearance:
Neanderthals likely had pale skin and red hair.
Pale skin would have been advantageous for Neanderthals in Europe, aiding in vitamin D synthesis due to reduced sunlight exposure.
Conclusion
While the Africa Replacement Hypothesis is supported by both molecular and other data, the discovery of interbreeding with Neanderthals adds complexity to our understanding of human evolution.

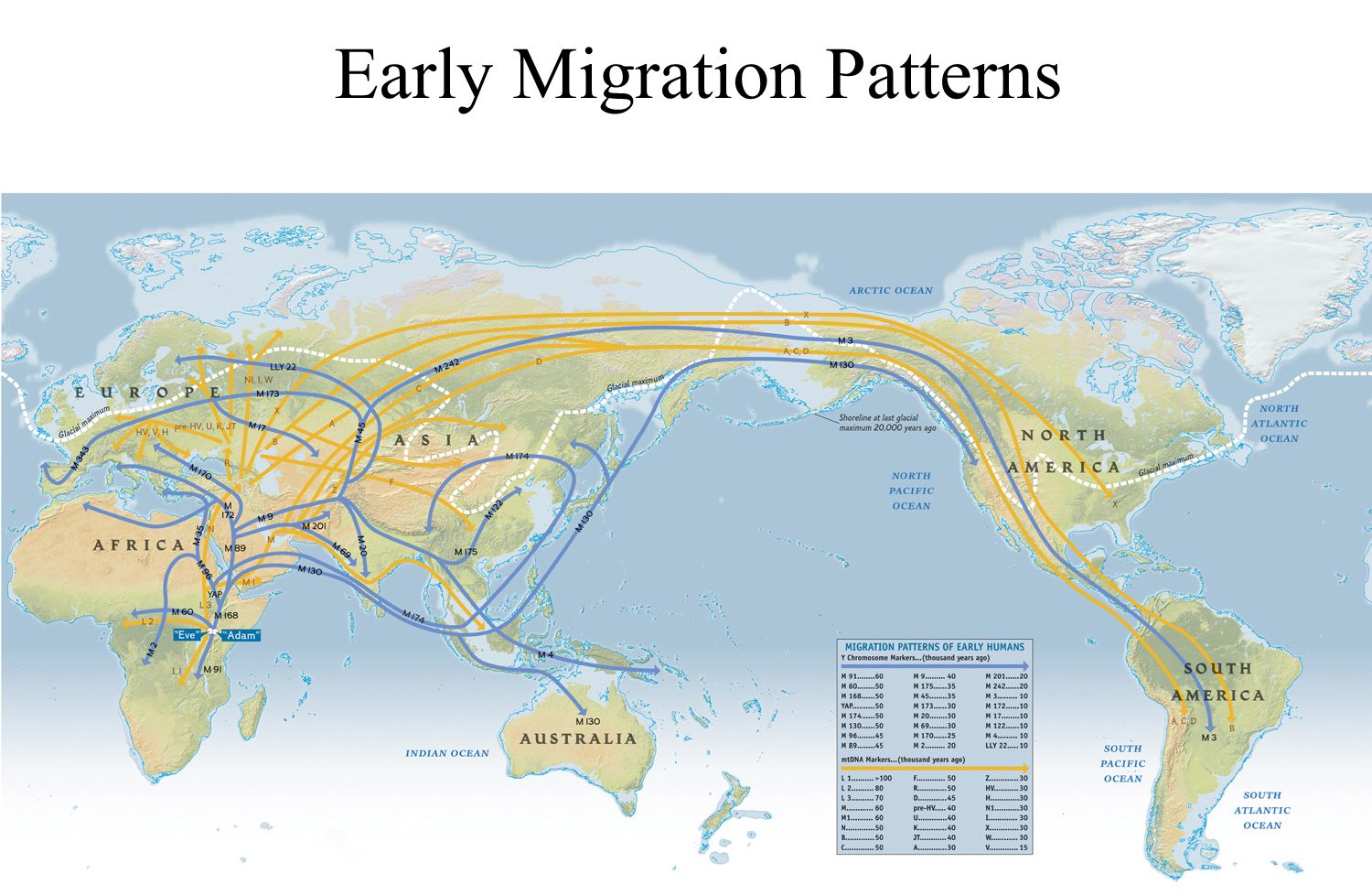
Key Findings from Molecular Data:
The migration map illustrates the pathways taken by ancient humans as they dispersed across the globe.
Pathways of Migration:
Colonization of the Americas:
Route: From Siberian Russia across the Bering Strait.
Timing: Multiple migration events occurred, particularly during the last great ice age when water was locked in glaciers, exposing a land bridge between Siberia and Alaska.
Routes Out of Africa:
The map identifies two main routes of migration from Africa:
The book's version may show only one route, whereas this detailed version captures the complexity of early human dispersal.
These routes indicate the diversity of migratory patterns and how humans adapted to various environments as they spread.
Significance:
The migration pathways reflect not only human adaptation to changing climates and landscapes but also the rich tapestry of human history as groups migrated, settled, and interacted with one another across continents.
Understanding these routes helps in tracing the genetic and cultural heritage of modern populations today.