H17 DNA
Je kan de bouw van DNA beschrijven %%(BINAS 70A)%%****:
- De twee strengen van een ^^DNA-helix^^ hebben een tegenovergestelde richting.
- DNA bestaat uit vier deoxyribonucleotiden, bestaande uit een fosfaatgroep, een suikermolecuul (deoxyribose) en een ^^stikstofbase^^ (A&T, C&G). Aan het ^^5’-einde^^ zit de fosfaatgroep.
- Chromatine bestaat uit ^^nucleosomen^^, opgebouwd uit DNA en ^^histonen^^.
- Heterochromatine: plaats waar chromatine heel dicht opeengepakt is (inactief)
- Euchromatine: plaats waar chromatine veel minder compact is (actief), er vindt wel translatie plaats.
- Mitochondriën bevatten cirkelvorming ^^mtDNA^^ met genen voor verschillende RNA-moleculen en enzymen voor aerobe dissimilatie.
- Het mtDNA erft over van moeder op kind.
- Bij de mens is een groot deel van het ^^DNA niet coderend.^^
- ^^Repetitief DNA:^^ korte DNA-sequenties die vaak worden herhaald, bijv. bij ^^telomeren^^.
- Introns: DNA binnen een gen, wordt echter niet gebruikt om een eiwit te coderen.
Je kan beschrijven hoe cellen DNA kopiëren (****%%BINAS 71D%%):
- ^^Helicase^^: enzym dat ketens splitst, verbreekt de waterstofbruggen tussen twee strengen (C&G moeilijker uit elkaar dan A&T).
- Bij DNA-verdubbeling ontstaan replicatievorken, waar een ^^DNA-polymerase^^ start met de replicatie. Het enzym leest een DNA-streng in de 3’→5’ richting. Aan de leidende streng groeit het nieuwe DNA continu (5’→3’) en aan de volgende streng achterwaarts in ^^Okazaki-fragmenten^^.
- ^^DNA-ligase^^: verbindt de losse DNA-fragmenten aan elkaar. ^^Primers^^ worden uiteindelijk vervangen door DNA.
- DNA-replicatie is ^^semi-conservatief^^.
Je kan beschrijven hoe het DNA kopiëren in het laboratorium gebeurd (****%%BINAS 71M2%%):
^^PCR-methode:^^ cyclus van drie stappen, 30 keer herhaald.
- openritsen DNA-fragment; bij 94 graden laten de waterstofbruggen los.
- koppelen ^^primers^^ aan DNA
- synthese van een nieuwe DNA-streng
Je kan uitleggen hoe je DNA fragmenten van elkaar kan scheiden en zichtbaar kan maken:
^^Gelelektroferese^^: scheidingstechniek op basis van (DNA-)molecuulgrootte.

- Korte fragmenten DNA komen verder dan lange fragmenten.
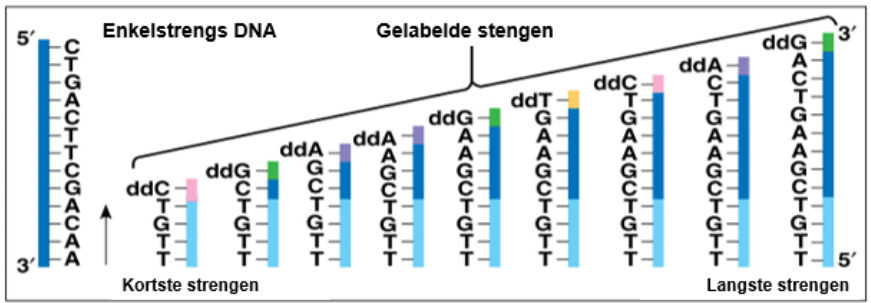
Je kan uitleggen op welke manieren de basenvolgorde in het DNA bepaald kan worden:
^^DNA-sequensen^^:
- DNA-molecuul wordt in stukjes verdeeld.
- Van het gewenste stuk wordt een groot aantal kopieën gemaakt.
- Strengen worden gescheiden en de ^^matrijsstreng^^ wordt gebruikt.
- Nieuwe DNA-strengen worden gemaakt door: een grote hoeveelheid nucleotiden en een kleine hoeveelheid fluorescerende nucleotiden. Na koppeling met een fluorescerende nucleotide stopt de keten.
- De nieuwe DNA-stukjes worden gescheiden door gelelektroferese (korte stukjes dalen sneller)
- Een detector meet de fluorescentie van de passerende DNA-stukjes.
Je kan uitleggen hoe een cel informatie omzet naar eiwitten %%(BINAS 71E, J)%%:
^^Transscriptie^^ :
- Transscriptie start bij de ^^promotor^^; de promotor bevat een ^^TATA-box^^.
- Bepaalde eiwitten hechten zich aan de promotor: dit zijn de ^^transscriptiefactoren^^.
- Deze factoren maken de binding van ^^RNA-polymerase^^ mogelijk. Dit enzym maakt mRNA.
- De transscriptie stopt mij de ^^terminator^^.
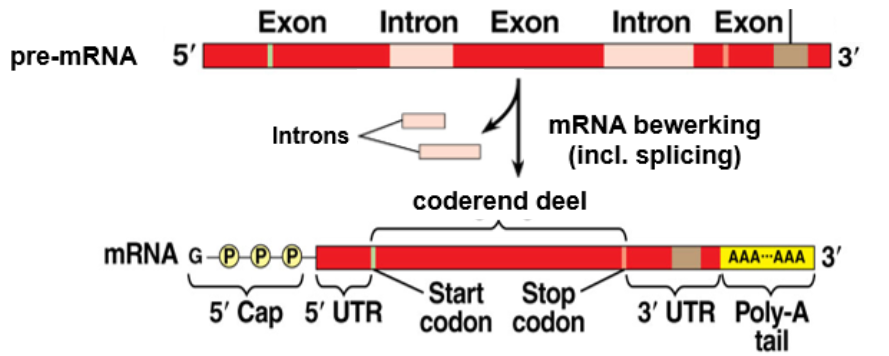
Daarna wordt het mRNA bewerkt. Tijdens ^^splicing^^ worden de ^^introns^^ (niet-coderend DNA) verwijderd, bij ^^capping^^ krijgt het mRNA een 5’cap, dit dient voor herkenning door ribosomen. Het molecuul krijgt ook een poly-A-staart, ter bescherming tegen exonuclease-enzymen.
^^Translatie :^^
tRNA koppelt aan een aminozuur om deze ‘af te leveren’ bij het ribosoom. Hier koppelt het tRNA aan het complementaire ^^startcodon^^ in het mRNA. Het rRNA katalyseert deze reactie.
Het aflezen van het mRNA gaat in ‘5→’3-richting. Translatie gaat door tot het ^^stopcodon^^ is afgelezen. Dan bind geen tR 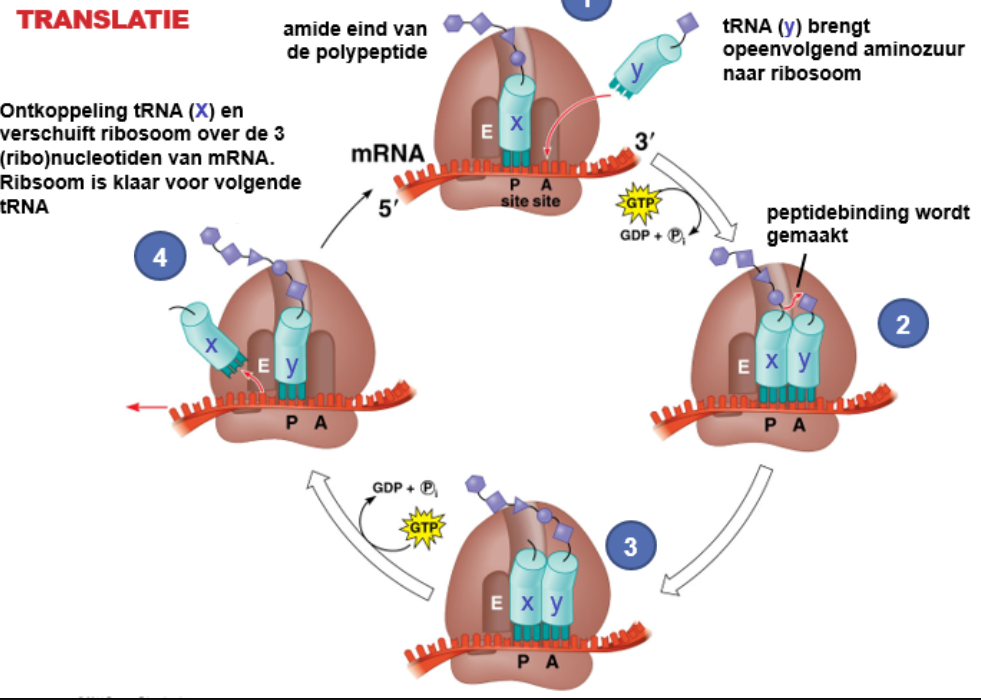 NA maar een releasefactor.
NA maar een releasefactor.
Je kan op basis van de relatie tussen tripletcode en aminozuur toelichten hoe eiwitten gevormd worden (****%%BINAS 71G%%):
Een triplet is een combinatie van achtereenvolgende 3 basen op het mRNA. Deze codeert voor een bepaald aminozuur. De derde base kan een ^^wiebelbase^^ zijn: een tRNA-molecuul kan op twee verschillende codes (bv. GCU en GCC) hetzelfde aminozuur koppelen.
Je kan toelichten dat een cel in staat is tot apoptose en dat deze een rol kan spelen tijdens de ontwikkeling van een meercellig organisme:
- ^^Apoptose^^ = geprogrammeerde celdood; bijvoorbeeld T-cellen die eigen weefsel aanvallen, vernietigen van ongewenste weefsels, versleten cellen, tumor cellen. Zie %%BINAS71L%%.
Je kan beschrijven welke typen mutatie er zijn:
- ^^Puntmutatie^^: verandering van één nucleotide.
- substitutie: één basenpaar is vervangen door een andere.
- deletie: één basenpaar is verdwenen
- insertie: één basenpaar is toegevoegd.
- ^^Chromosoommutatie^^: de bouw van een chromosoom wordt veranderd.
- inversie: omkering
- deletie: delen van chromosoom verdwijnen
- duplicatie: verdubbeling
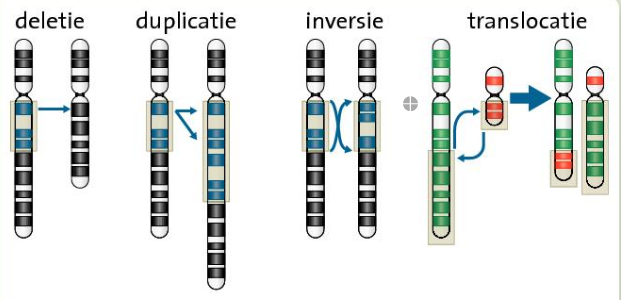
- ^^Genoommutaties^^: er ontstaan cellen met afwijkend aantal chromosomen. Oorzaak: afwijkende meiose (%%BINAS 76B%%).
Je kan uitleggen waardoor mutatie veroorzaakt kan worden:
Mutaties treden soms spontaan op. De volgende factoren vergroten de kans op mutaties: reactieve zuurstofverbindingen uit de mitochondriën of mutagene stoffen (uv- en radioactieve straling, andere stoffen uit omgeving).
Je kan uitleggen dat mutatie het fenotype kan beïnvloeden:
Mutatie heeft geen gevolg als…
- het gaat om een mutatie van een wiebelbase
- als het gaat om een mutatie op niet-coderend DNA of op een intron
Een mutatie kan wel de eiwitvolgorde veranderen als er een ^^leesraamverschuiving^^ ontstaat. Zo kan er een andere eiwitvolgorde ontstaan, of ketens worden korter of langer.
Je kan mechanismen voor genregulatie noemen en het belang ervan toelichten (****%%BINAS 71F%%):
Cellen kunnen cellen activeren of uitschakelen: zo heeft elke cel een eigen taak en een eigen eiwit(hormoon) productie.
- door middel van ^^chromatinestrucuur^^: milieufactoren (voeding/stress) beïnvloeden de mate van ^^methylering^^ van het DNA.
- ^^activering^^: door aanhechting van een acetylgroep verandert de chromatinestructuur naar los (^^euchromatine^^), waardoor transscriptie in dit deel van het DNA kan toenemen.
- ^^silencing^^: door aanhechting van methylgroepen wordt het DNA compacter (^^heterochromatine^^)
- Tijdens de transcriptie:
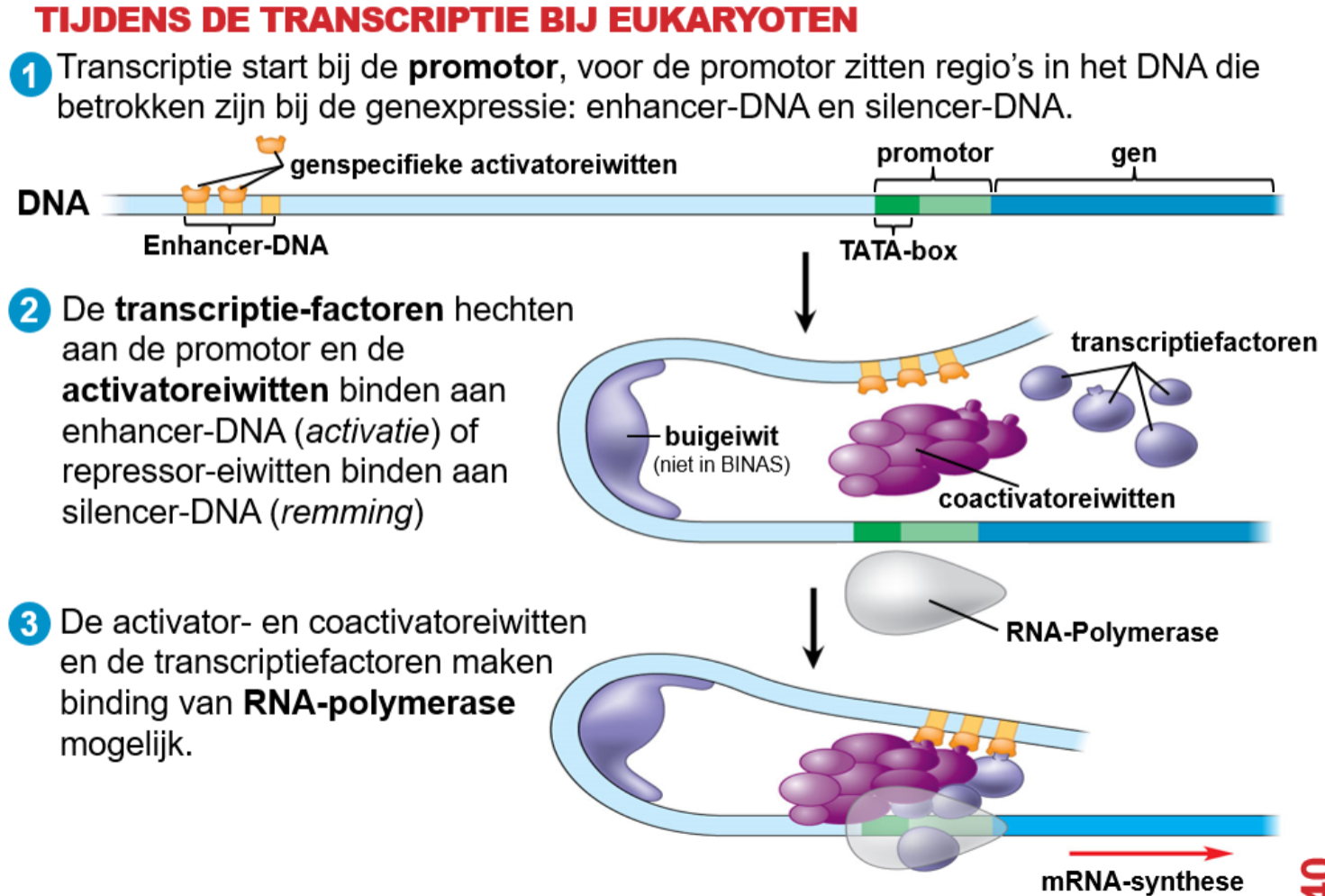 ^^Repressoreiwitten^^ koppelen aan ^^silencer-sequenties^^ en blokkeren het tot stand komen van de transcriptie.
^^Repressoreiwitten^^ koppelen aan ^^silencer-sequenties^^ en blokkeren het tot stand komen van de transcriptie.
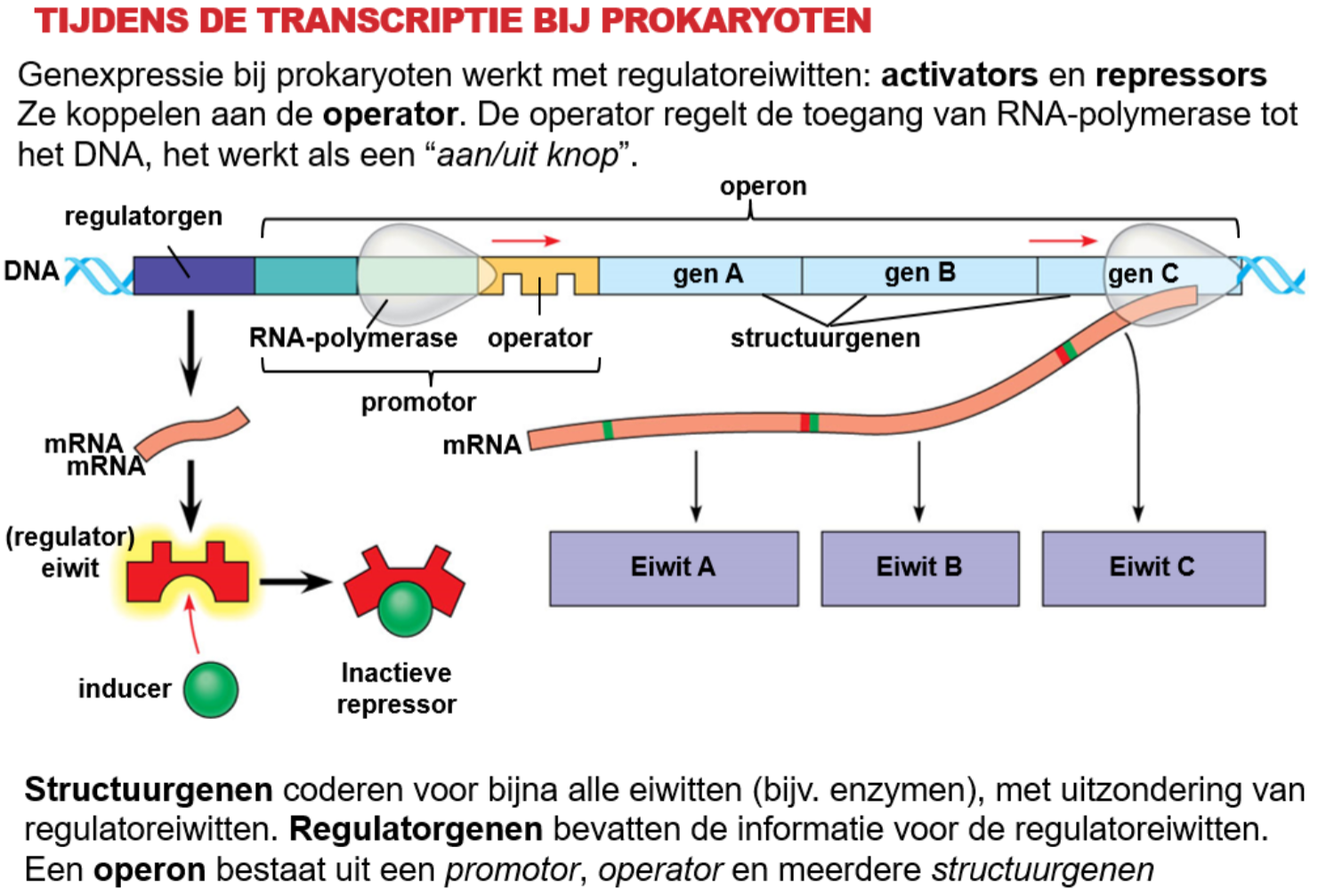 b.
b.
Een ^^correpressor^^ activeert de repressor, een ^^inducer^^ deactiveert de repressor.
- na de transcriptie:
- ^^splicing^^: vanuit het pre-mRNA ontstaat een variatie aan mRNA-ketens.
- vertragen van het aanhechten van de poly-A-staart.
- lengte van de poly-A-staart bepalen en zo de levensduur van het mRNA.
- ^^micro-RNA^^ kan translatie van bepaald mRNA blokkeren.