Chapter 12 - Transport Across Cell Membrane
Membranes have various proteins, which allow for the transport of different molecules that cannot traditionally pass through the membrane to pass through
Ion concentration makes a gradient across a cell membrane, particularly useful in specific cells such as nerve cells. They are created by the unequal distribution of an ion between sides, which creates an electrochemical gradient across the membrane. This is mostly done with cations, but also with the chloride anion.
With Na+ and K+, there is always more Na+ outside and always more K+ than inside. Calcium is stored in the ER, and servers as am important cofactor and has a low concentration in the cytosol.
Simple diffusion is transport of small non-polar uncharged molecules along their concentration gradient. Channel mediated and transporter mediated transportation is passive, both going down their concentration gradients (no energy required). Active transport goes against the concentration gradient, and requires energy.
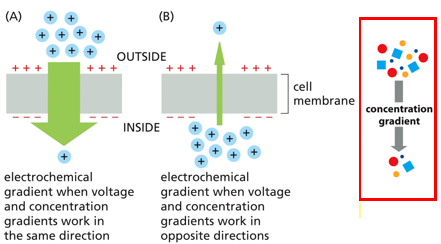
The electrochemical gradient has 2 parts, molecular concentration and charge. If both parts work together, the gradient is very strong. When they oppose each other, the effect is smaller
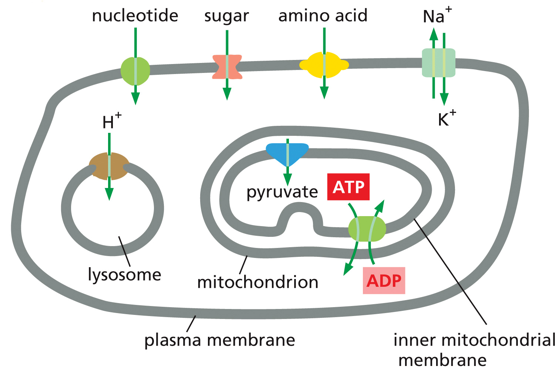
In the plasma membrane, different nutrients get imported in (nucleotide/sugar/amino acid) and Na+/K+ are important for action potential in neural cells. Energy conversion in the mitochondria requires the inputs of pyruvate and ADP, and releases an ADP, all of which have transport proteins. The lysosome has a low pH and needs protons transported in to help maintain this.
Different ways things transport across the membrane includes simple diffusion, channel-mediated methods, transporter-mediated methods, and pumps. These are often driven by gradients, light, or ATP.
Primary active transport makes ion gradients, which are used to transport other things against their transport gradients (secondary active transport). Energy sources include ATP, photons, and electrons.
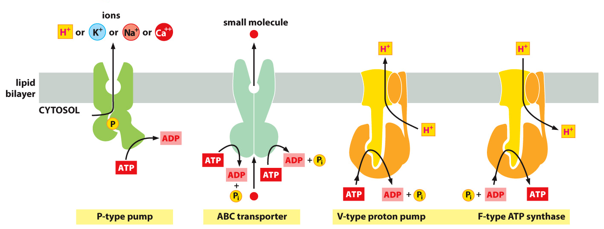
Primary active transport utilizes ATP, phosphorylating the protein. There are 3 types of ATP pumps. P-type pumps, ABC transporters, and V-type pumps/P-type ATP synthase. P-type pumps use phosphorylation from ATP to change conformation. V-type pumps are eukaryotic, and F-type pumps are prokaryotic. V and F use the energy from ATP to power a change, rather than the phosphate itself causing it.
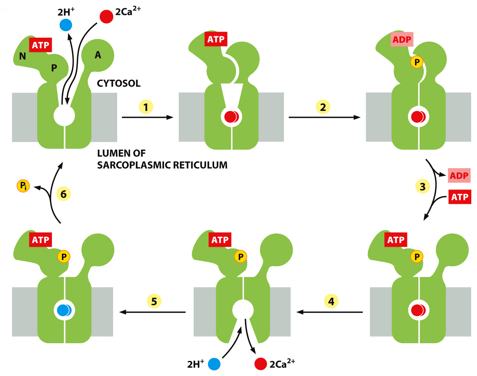
P-type pumps have calcium binding sites. They are found between the lumen of the sarcoplasmic reticulum and cytosol. One example is The Na+/K+ pump, along with the for calcium. Phosphorylation doesn’t give it energy, rather causes a conformational change.
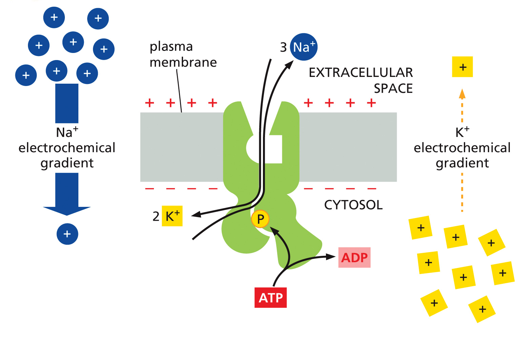
Na+/K+ pumps exist to help regulate the electrochemical gradient, especially in the brain. Across the membrane, the concentration of Na+ is higher outside and the concentration of K+ is higher inside. The charge is more negative inside than out. When activated by ATP, it will bring in 2 K+ and send out 3 Na+, which helps keep the more negative charge inside.
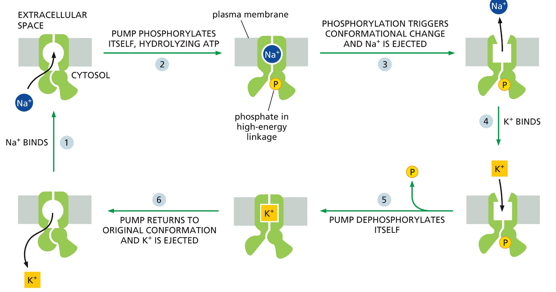
The Na/K pump will hydrolyze ATP to activate itself, which triggers a conformational change releasing Na+. This allows for K+ to ender the protein. Then, it is dephosphorylated and K+ is released into the cytosol
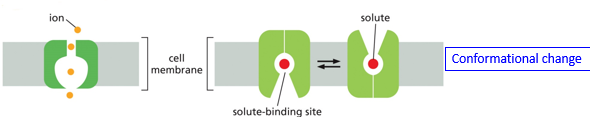
There is also passive transport, which may go along with a gradient. This may still use transporters and channel proteins. A conformational change may occur when a certain ion in bound, switching which side of the protein is open. Other names for this are transporter mediated diffusion, facilitated transport, and facilitated diffusion.
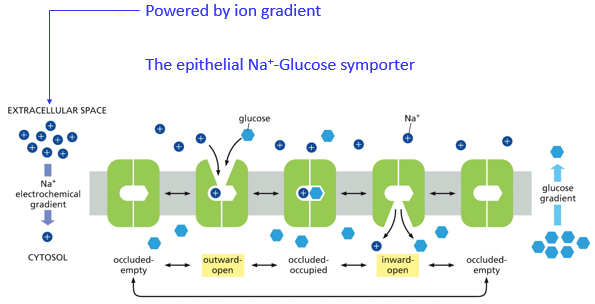
Passive transport is requires for molecules that are too large/charged to cross the semipermeable membrane on their own, such as glucose, pyruvate, and amino acids. An eversion is when one end of the pump is open, open to the outside is usually eversion 1, and open inside is eversion 2. The occlusion is when the transporter is closed with molecules inside, between eversions.
Blood cells have a low Km for glucose for their passive transport uniporters, as they only do glycolysis for ATP. The liver has a lower Km, as it can do neoglucogenesis.
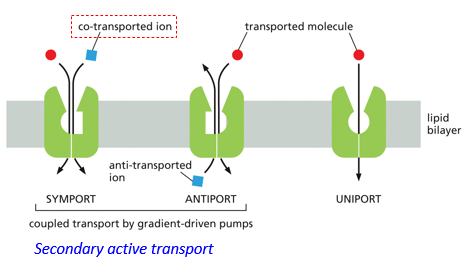
Gradient driven transport (secondary active transport) utilizes symports and antiports. Symports transport a molecule and an associated ion in the same direction. Antiports will take one molecule in and remove the other. There also uniports, which will only transport the target molecule. Both symports and antiports are coupled transport by gradient-driven pumps.
Glucose transport in an epithelial cell is regulated by transporters. Typically, they will diffuse across a membrane, but here molecular fences called tight junctions restrict their movement, to maintain function of the epithelial cell, which may line an intestine.
The mitochondria doesn’t have any P-type pumps, as it produces ATP rather than uses it. This requires the mitochondria to rely solely on secondary active transport.
Ion channels form hydrophilic channels across the membrane, they do not open/close but rather allow only charged ions through. They are all highly specific for a particular kind of ion. Occasionally water will slip through but water is an exception to almost every rule in biology
The bacterial K+ channel opens like a pinwheel, it attracts positive ions and repels negative ions due to the negative protein charge, allowing them into the vestibule space. They will be pushed out when another ion enters the channel and “knocks” the original ion out into the side of the membrane with the lower concentration
There are carbonyls coming from the carbon backbone (in the peptide bond) that indicate the specificity from Na+ and K+. Each ion is surrounded by water due to its positive charge. The orientation of these water molecules varies from ion to ion, due to cation size. It takes energy to remove these. The carbonyls attract the ion in the same manner as the water molecules, think of it like the spider wall in ANW
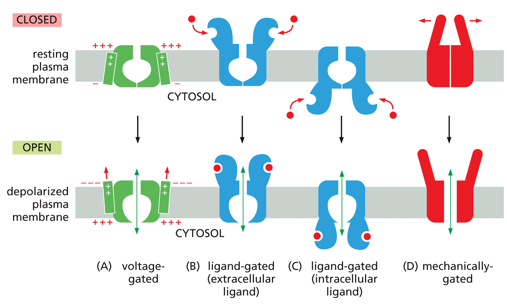
Gated ion channels have various different ways of channel regulation (opening/closing). Some are voltage gated, ligand gated, or mechanically gated. Voltage gated open with charge, ligand gated open when bound by a particular molecule, and mechanically gated open/close with physical touch (like in the ear, physical senses)
K+ leak channels allow for K+ to move. When closed, the plasma membrane is a potential of 0 mV. When open, the membrane potential balances the tendency of K+ to leave. There is no way for the ion gradient and chemical gradient to both be 0 at once.
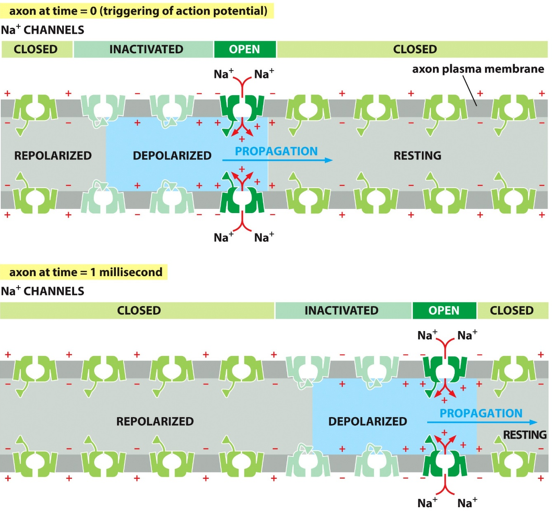
The neurons use electric gradients to send signals, of more positive depolarizations and resting/repolarized negativity. These are known as action potentials (APs). They will go through 3 conformations: open, plugged, and closed. The plug is what causes the refractory period.
For a voltage gated channel, the initial conformation is closed, with the plug out. They have helices to sense voltage changes caused by APs, which open the channel to let in a given ion, like Na+. Since closing takes to long, they plug themselves and enter the refractory period and enter the inactive conformation. Once they close, they can send another AP.
The change in membrane potential due to a specific channel is sensed by the nearby channel, causing the AP to travel. It doesn’t go backwards due to the refractory period
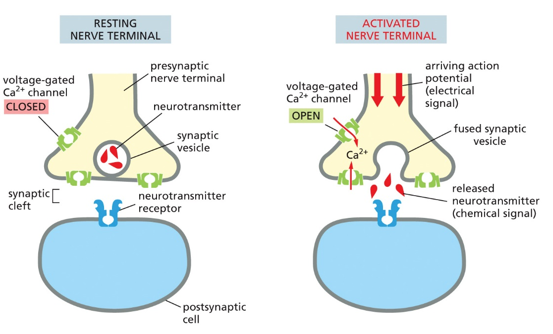
When an action potential reaches the end of the nerve cell, known as the axon terminal, certain voltage-gated ion channels are activated and open, allowing in Ca2+. These calcium ions allow for synaptic vesicles to burst open, and release certain chemicals to the postsynaptic neuron. Only chemicals, called neurotransmitters, can jump the synaptic cleft gap. The AP reaching the axon terminal on the presynaptic neuron releases the neurotransmitter from a synaptic vesicle, which jump the gap and bind to postsynaptic neurons, opening them and restarting a signal
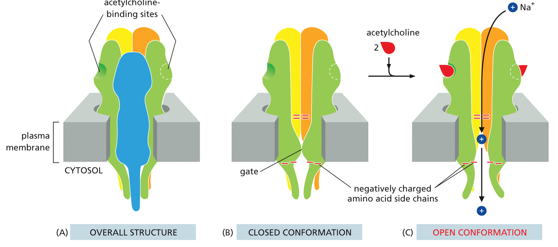
The acetylcholine receptor is closed when not bound to by Ach. When it is, the channel opens and allows in Na+ ions, which depolarizes the postsynaptic cell, or makes it have a more positive charge. It has 5 subunits
There are certain channels knows as aquaporins, which consist of many alpha helices and allow water into the cell. This makes water transport faster than it would be if it only relied on its gradient. It does cross on its own, but also uses these.