17.3 Photosynthesis
Light energy is transformed into chemical energy trapped in the bonds of the complex organic molecules produced. Organisms that photosynthesis, like plants are said to be autotrophic.
Heterotrophic organisms obtain complex organic molecules by eating other organisms.
Both autotrophic and heterotrophic organisms then break down complex organic molecules during the process of respiration to release the energy they need to drive metabolic processes.
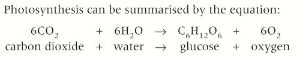 Structure and function of chloroplasts
Structure and function of chloroplasts
The network of membranes within chloroplasts provide a large surface area to maximise the absorption of light essential in the first step of photosynthesis.
The membranes form flattened sacs called thylakoids which are stacked to form grana. (singular granum)
The grana are joined by membranous channels called lamellae.
Light is absorbed by complexes of pigments such as chlorophyll which are embedded within the thylakoid membranes.
The fluid enclosed in the chloroplast is called the stroma and is the site of the many chemical reactions resulting in the formation of complex organic molecules.
Chlorophyll
Pigments molecules absorb specific wavelengths of light and reflect other and this is why they have different colours. The primary pigment in photosynthesis is chlorophyll, this absorbs mainly red and blue light and reflects green light.
The primary pigments is chlorophyll a. Other pigments like chlorophyll b, xanthophylls, and carotenoids absorb different wavelengths of light than those absorbed by chlorophyll a. Different combinations of pigments are the reason for different shades and colours of leaves.
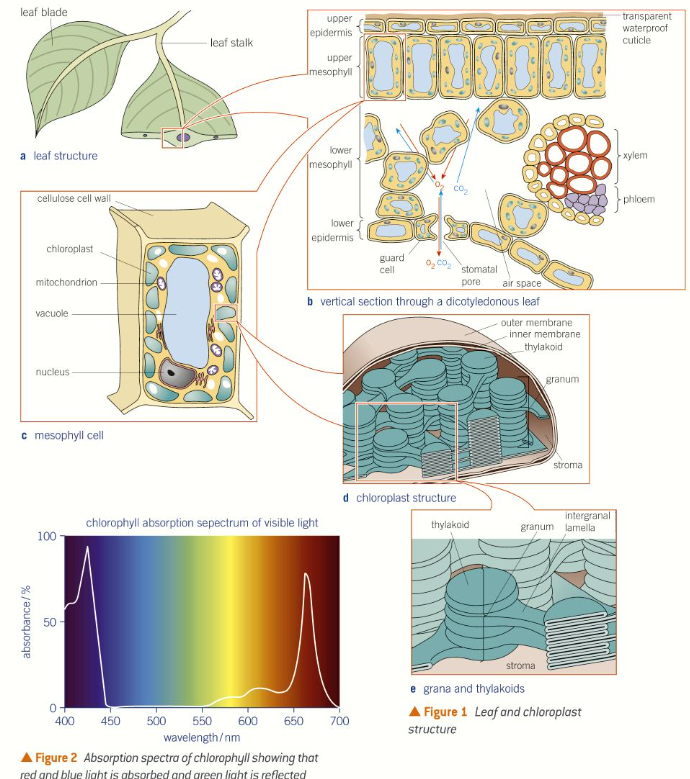
Chlorophyll b, xanthophylls and carotenoids are embedded in the thylakoid membrane of the chloroplast. These and other protein and pigments form a light harvesting system (also known as antennae complex) The role of the system is to absorb or harvest light energy of different wavelengths and transfer this energy quickly and efficiently to the reaction centre. Chlorophyll a is location in the reaction centre, which is where the reactions involved in photosynthesis take place.
The light harvesting system and the reaction centre are collectively known as a photosystem.
Chlorophyll a - blue/green (found in all plants)
Chlorophyll b - yellow/green
Carotenoids - orange
Xanthophylls - yellow
Pheophytins - grey
Plants using sunscreen
Plants need light to photosynthesis but if the sunlight is too intense, chlorophyll is destroyed. Chlorophyll has to be continuously synthesised during the summer to maintain the level that is needed to photosynthesise at the required rate. Chlorophyll is not produced when there is little or no sunlight - this is why areas of grass that have been covered turn yellow.
Carotenoids are accessory pigments responsible for the yellow/orange colours seen in plant leaves. Orange carotene and yellow xanthophyll are two examples. These are not normally seen because they are masked by the green colour of chlorophyll. Carotenoids are not broken down unlike strong sunlight and are present through the whole growing season.
The shorter days and cooler nights of autumn cause changes in the pigment composition in leaves. Chlorophyll a is no longer needed and leaves turn yellow/orange as we see the carotenoids.
Anthocyanin is a red/purple pigment formed from a reaction between sugars and proteins present in cell sap. It is produced when the concentration of sugars is high. High light intensity also promotes the production of anthocyanins.
Anthocyanin produces the red skin of apples and the purple of black grapes. The colour of the anthocyanin pigments is pH dependant, leading to a range of different colour from red to purple.
Anthocyanins act as a sunscreen by absorbing blue-green and ultraviolet light thereby inhibiting the destruction of chlorophyll. In their role as pigments they help trees maximise production towards the end of the growing season as the weather changes in autumn.
The purple/red is also though to camouflage leaves from herbivores blind to red wavelengths.
Investigating photosynthetic pigments
Chromatography can be used to separate the different pigments in a plant extract. The mobile phase would be the solution containing a mixture of pigments and stationary phase a thin layer of silica gel applied to glass.
The different solubilities of the pigments in the mobile phase, and their differing interactions with the stationary phase, lead to them moving at different rates. This results in the pigments being separated as they move through the silica gel.
The retention value (Rf) for each pigments

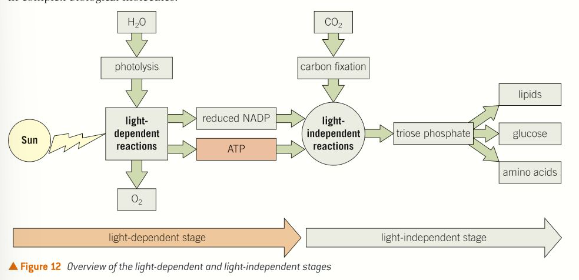
The two stages of photosynthesis
Light-dependant stage - energy from sunlight is absorbed and used to form ATP. Hydrogen and water is used to reduce coenzyme NADP to reduced NADP.
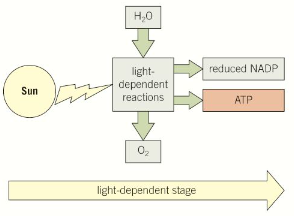
Light-independent stage - hydrogen from reduced NADP and carbon dioxide is used to build organic molecules such as glucose. ATP supplies the required energy.
The light-dependant stage of photosynthesis
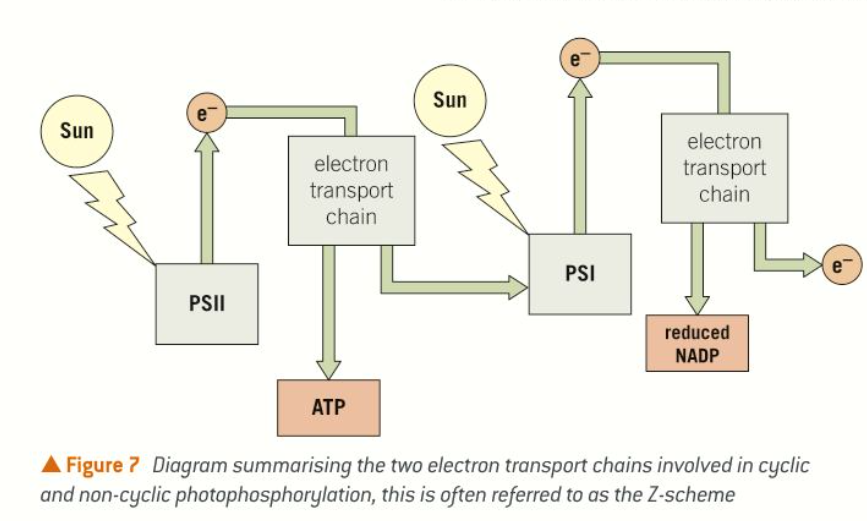
Non-cyclic photophosphorylation
Two photosystems are involved in non-cyclic photophosphorylation, PSII and PSI. The reaction centre of PSI absorbs light at a higher wavelength (700 nm) than PSII (680 nm). The light absorbs excites electrons at the reaction centres of the photosystems.
The excited electrons are released from the reaction centre PSII and are passed to an electron transport chain. ATP is produced by the process of chemiosmosis.
The electrons lost from the reaction centre at PSII are replaced from water molecules broken down using energy from the Sun.
Excited electrons are released from the reaction centre at PSI, passed to another electron transport chain, and ATP is again produced by chemiosmosis. The electrons lost from this reaction centre are replaced by electrons that have just travelled along the first electron transport chain after being released from PSII.
The electrons leaving the transport chain following PSI are accepted, along with a hydrogen ion, by the coenzyme NADP, forming reduced NADP. Reduced NADP provides the hydrogen or reducing power in the production of organic molecules, such as glucose in the light-independent stage which follows.
Photolysis
Water molecules are split into hydrogen ions, electrons, and oxygen molecules using energy from the Sun in a process called photolysis.
The electrons released replace the electrons lost from the reaction centre of PSII. This is why water, along with light and carbon dioxide is a raw material of photosynthesis.
The oxygen evolving complex which forms part of PSII is an enzyme that catalyses the breakdown of water. Here water molecules are being split into hydrogen ions, electrons, and oxygen molecules using energy from the Sun in process called photolysis.
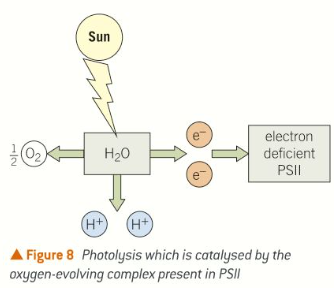
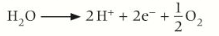 Oxygen gas is released as a by-product. The protons are released into the lumen of the thylakoids, increasing the proton concentration across the membrane. As they move back through the membrane down a concentration and electrochemical gradient, they drive the formation of more ATP.
Oxygen gas is released as a by-product. The protons are released into the lumen of the thylakoids, increasing the proton concentration across the membrane. As they move back through the membrane down a concentration and electrochemical gradient, they drive the formation of more ATP.
Once the hydrogen ions are returned to the stroma, they combine with NADP and an electron from PSI to form reduced NADP. → this is used in the light-independent reactions. This process removes the hydrogen ions from the stroma so it helps to maintain the proton gradient across the thylakoid membrane.
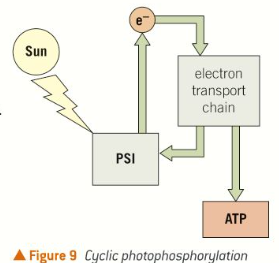
Cyclic photophosphorylation
The electrons leaving the electron transport chain after PSI can be returned to PSI, instead of being used to form reduced NADP, leading to cyclic photophosphorylation. This means that PSI can still lead to the production of ATP without any electrons being supplied from PSII.
Reduced NADP is not produced when this happens.
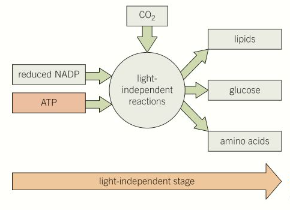
The light independent stage of photosynthesis
This takes place in the stroma of chloroplasts and uses carbon dioxide as a raw material. The products from light-dependant stage (ATP and reduced NADP) are required. Organic molecules are produced by a series of reactions known as the Calvin cycle.
Calvin cycle
Carbon dioxide enters the intercellular spaces within the spongy mesophyll of leaves by diffusion from the atmosphere through stomata.
It diffuses into cells and into the stroma of chloroplasts where it is combined with a 5 carbon molecule called Ribulose bisphosphate (RuBP)
The carbon dioxide is therefore fixed, meaning it is incorporated into an organic molecule.
The enzyme ribulose bisphosphate carboxylase (RuBisCO) catalyses the reaction and an unstable 6 carbon intermediate is produced.
RuBisCO is the key enzyme in photosynthesis. It is a very insufficient enzyme as it is competitively inhibited by oxygen so a lot of it is needed to carry out photosynthesis successfully. Biologists estimate that is it probably the most abundant enzyme in the world.
The unstable 6 carbon compound formed immediately breaks down, forming 2 3-carbon glycerate 3-phosphate (GP) molecules.
Each GP molecule is converted to another 3 carbon molecule, triose phosphate (TP), using a hydrogen atom from reduced NADP and energy supplied through ATP, both supplied from the light-dependant reactions of photosynthesis.
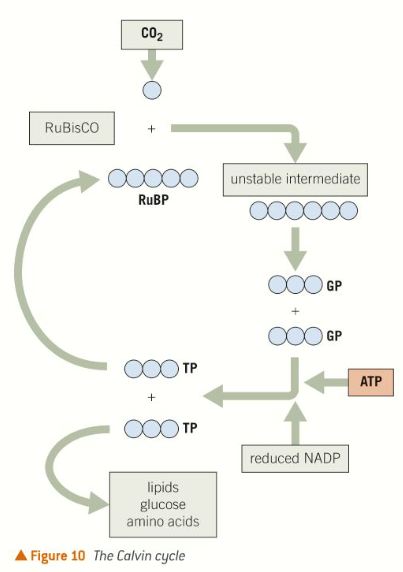
Triose phosphate is a carbohydrate, 3 carbon sugar, the majority of which is recycled to regenerate RuBP so that the Calvin cycle can continue. It is a starting pointing for the synthesis of many complex biological molecules, including other carbohydrates, lipids, proteins and nucleic acids.
The Calvin cycle can be summarised in 3 steps
Fixation - carbon dioxide is fixed (incorporated into an organic molecule) in the first step.
Reduction - GP is reduced to TP by the addition of hydrogen from reduced NADP using energy supplied by ATP.
Regeneration - RuBP is regenerated from the recycled TP
Regeneration of RuBP
For one glucose molecule to be produced 6 carbon dioxide molecules have to enter the Calvin cycle, resulting in 6 full turns of the cycle.
This will result in the production of 12 TP molecules, 2 of which will be removed to make the glucose molecule.
This means that 10TP molecules are recycled to regenerate 6 RuBP molecules.
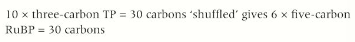

Photorespiration
Stomata need to be open for plants to obtain carbon dioxide for photosynthesis. Water vapour leaves plants through open stomata by the process of transpiration. When the temperature is high and humidity of the atmosphere is low plants can lose too much water. To prevent excess water loss, stomata close.
This prevents the entry of carbon dioxide into the leaves of the plant. The plant will still be photosynthesising and so the carbon dioxide levels fall and oxygen levels increase.
Oxygen is a competitive inhibitor of the enzyme RuBisCO, leading to the production of phosphoglycolate and reducing the production of GP. This only happens when the concentration of carbon dioxide becomes very low.
Phosphoglycolate is a toxic 2 carbon molecule that needs to be removed. It is converted by the plant into other organic molecules and energy from ATP is needed for the conversion.
RuBisCO has a higher affinity for co2 than oxygen and approximately 25% of the products of the Calvin cycle are lost in photorespiration, reducing the efficiency of photosynthesis.
Summary of photosynthesis