Bio 301 - Ch 4 Book Notes
microbial nutrition
a limiting factor to microbial growth is the finite supply of nutrients in the environment
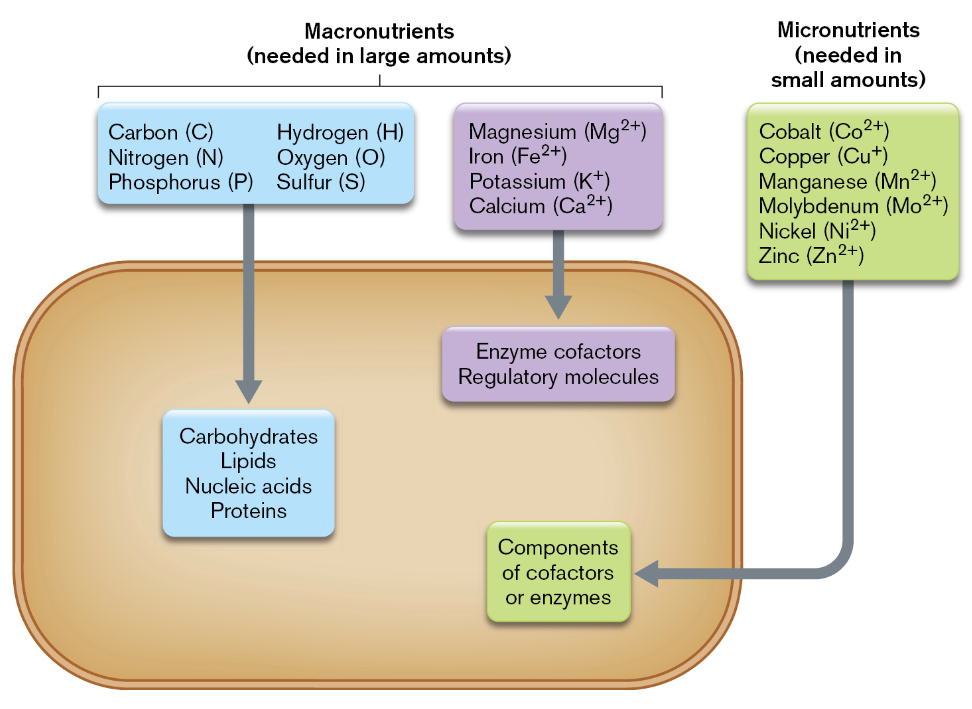
essential nutrients are compounds that microbes must have but cannot make
these nutrients must be found from the surrounding environment
if this nutrient is depleted, the microbe will not grow
all microorganisms require a minimum set of macronutrients and micronutrients
common macronutrients include C, N, P, H, O & S
there are also macronutrient cations that serve as cofactors: Mg2+, Fe2+, K+
Ca2+ acts as a regulatory signaling molecule
common micronutrients include Co, Cu, Mn, Mo, Ni & Zn
all cells require nutrients to increase in biomass and gain energy
microbes build biomass through autotrophy or heterotrophy
all Earth’s life forms are based on carbon which is a limited resource that must be recycled to maintain life
the survival and metabolism of any one group of organisms depend on the survival of other groups of organisms
the carbon cycle involves two counterbalancing metabolism: heterotrophy & autotrophy
heterotrophy breaks down multi-carbon nutrients and harvests the C to make cell constituents & some is converted into CO2 as a waste product
autotrophy takes the CO2 molecules discarded as waste by heterotrophs and reassembles them into multi-carbon nutrients like glucose
the glucose is then used as a C source for the heterotrophs
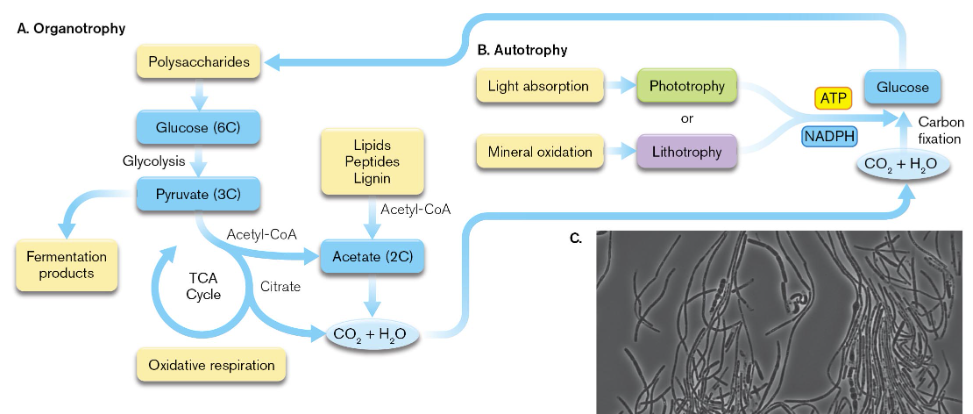
organotrophy
a form of heterotrophy in which organic sources are broken down in ways that harvest carbon for growth and give energy (oxidation)
these organisms convert a generous amount of organic carbon source to CO2 which is then released into the atmosphere
autotrophs assimilate CO2 gas as a carbon source via CO2 fixation which reduces CO2 to generate a complex, organic component of C, H, and O
autotrophs are classified as photoautotrophs or chemolithoautotrophs based on how they obtain their energy
photoautotrophs use light energy to fix CO2 into biomass but chemolithoautotrophs fix CO2 using chemical reactions
chemolithoautotrophs gain energy by oxidizing inorganic substance like iron or ammonia & they do not use light
microbes obtain energy through phototrophy or chemotrophy
macronutrients provide the essential building blocks that are needed to synthesize proteins & other cell structures
a source of energy is required — absorption of light or from redox reactions
chemotrophs
capture the energy difference to do work
two categories of chemotrophs use different sources of electron donors
lithotrophs: oxidize inorganic chemical for energy
organotrophs: oxidize organic compounds for energy
microorganisms that can use a mix of different sources of energy and carbon are mixotrophs
Rhodospirillum rubrum grows by photoheterotrophy when light is available and oxygen is absent, but it can switch to organotrophy and respiration, without absorbing light, when O2 is available
the amount of energy harvested from oxidizing a compound depends on the compound’s reduction state
the more reduced the compound is, the more electrons it has to give up and the higher potential energy yield
a highly reduced compound can donate electrons to a less reduced compound

energy is stored for later use
once energy is obtained, it must be converted to a useful form that the cell can use; this might be electrochemical or chemical energy
energy stored by an electrical potential across the membrane is called membrane potential
this is generated when chemical energy is used to pump protons, Na+, or K+ to the outside of the cell & makes the cation conc greater on the outside of the cell compared to the inside
an example is membrane proteins such as cytochrome oxidases use energy from respiration to pump protons across the cell membrane and out of the cell, generating a proton gradient
the proton gradient (delta P) plus the charge difference (voltage potential) across the membrane forms an electrochemical potential
when this electrochemical potential includes a proton gradient, it also has proton potential or proton motive force
the energy stored in the PMF can be used by specific transport proteins to move nutrients into the cell, to directly drive motors that rotate flagella & to drive the synthesis of ATP production by ATP synthase
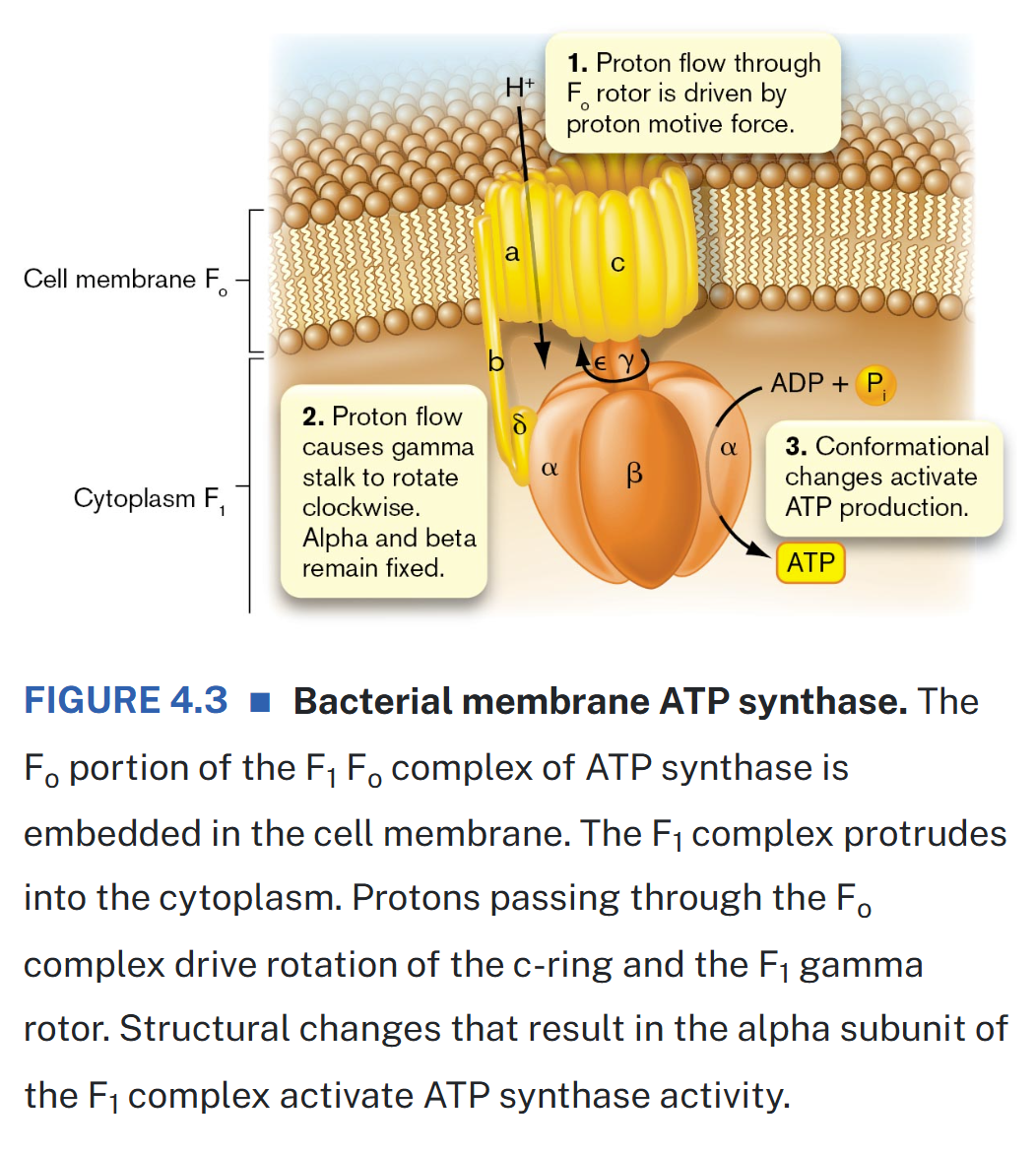
ATP synthase components & function mechanism
the membrane-embedded F-ATP synthase, also called F1Fo ATP synthase, provides most of the ATP for aerobic respiring cells
ATP synthase is a complex of many different proteins
the enzyme includes a channel (Fo) which allows protons to move across the membrane & drive the rotation of the ATP c-ring
the rotation of the c-ring causes changes in the F1 complex that mediate formation of ATP
some bacteria living in alkaline environments use a different form of ion motive form involving Na+ (sodium motive force)
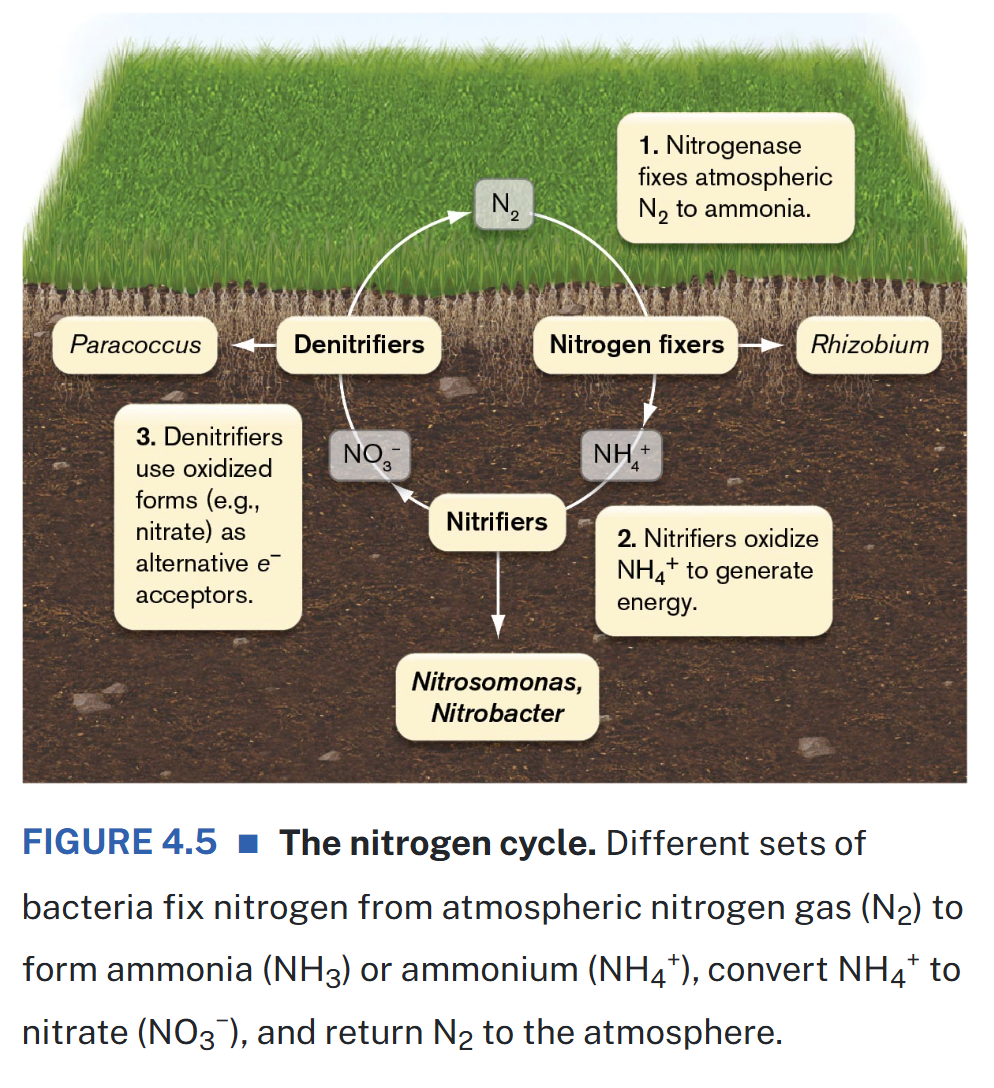
The Nitrogen Cycle
N is an essential component of proteins, nucleic acids, and other cell constituents
79% of the Earth’s atmosphere is composed of N2 gas which is an inaccessible form of N for most organisms
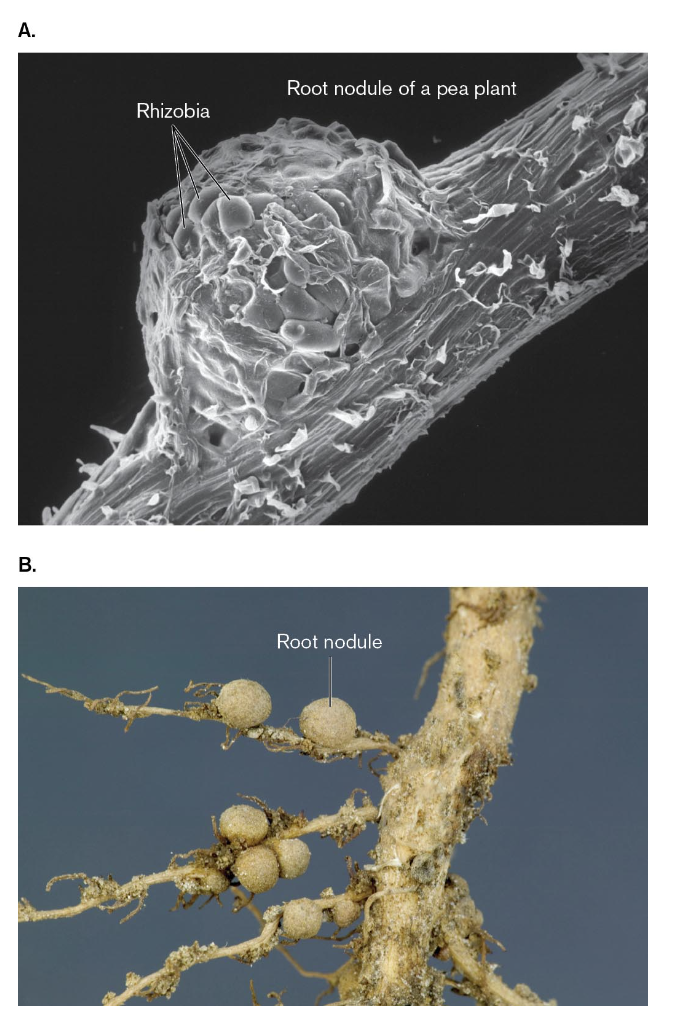
nitrogen-fixing bacteria have the ability to convert diatomic nitrogen into ammonium
these bacteria may be free-living in the soil or in water or they may form symbiotic associations with plants
the product ammonium is then used by all microbes to make amino acids and other nitrogen-containing compounds
nitrogen-fixing bacteria that form symbiotic relations include the following:
Rhizobium, Sinorhizobium, and Bradyrhizobium species are nitrogen-fixing symbionts of leguminous plants such as soybeans, chickpeas, and clover
although symbionts are most widely known nitrogen-fixing bacteria, the majority of nitrogen in soil & aquatic environment is fixed by free-living bacteria and archaea
various groups of organisms collaborate to recycle ammonium ions and nitrate ions into nitrogen gas & collect energy in the process of converting reactants to products and biologically useful forms of N
in nitrification, nitrifiers gain energy by converting or oxidizing ammonia in two steps to form nitrate
this process is a form of lithotrophy
denitrification is a process that uses nitrate and related inorganic forms of nitrogen as terminal electron acceptors for certain electron transport chains
these bacteria reduce nitrate to N2 which sends N gas back into the atmosphere which roughly balances the amount removed by nitrogen fixation
Nutrient Uptake
a microbe must find means of motility to access nutrients and move those nutrients across the membrane into the cytoplasm
the cell membrane is selectively permeable to the nutrients that the cell can use
selective permeability can be achieved in the following ways:
membrane-spanning protein channels or pores that differentiate between substrates of certain sizes or chemical composition; permits the entry of those substances (facilitated diffusion)
substrate-specific carrier proteins, permeases, that span the cytoplasmic membrane and transport substrates into the cell
nutrient-binding proteins that sample the external environment (Gram-positive bacteria) or the periplasmic space (Gram-negative bacteria) for specific nutrients & then those pass those nutrients to specific permeases in the membrane
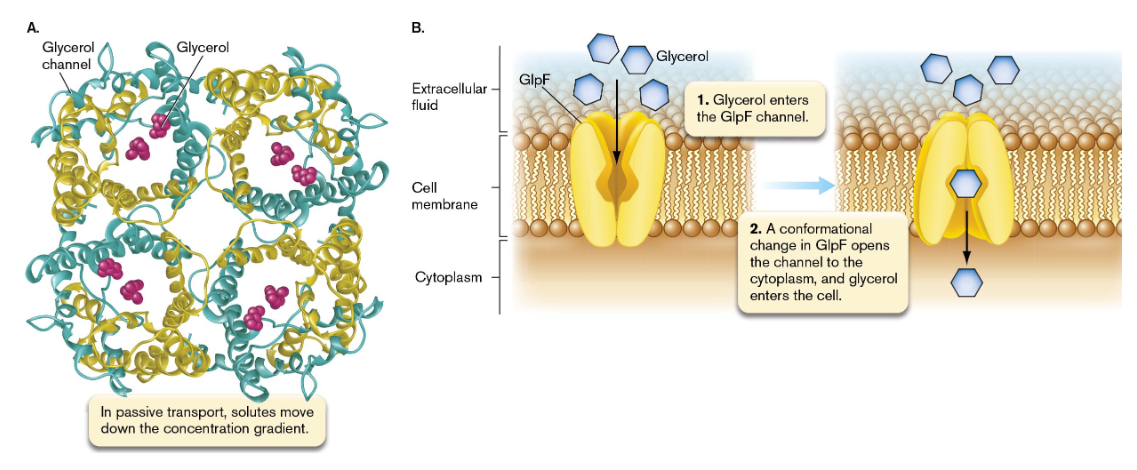
microbes must overcome the issue of low nutrient concentrations in the natural environment
if the intracellular concentration of a nutrient never rose higher than the extracellular concentration, the cell would starve in low-nutrient environments
to surpass this issue, most organisms have evolved efficient transport systems that concentrate nutrients inside the cell relative to outside
moving molecules against a concentration gradient requires some form of energy input
environments where nutrients are available but exist at low concentrations & are composed in form that cannot be transported into the cell
to combat this, many microbes unlock the nutrients by secreting digestive enzymes that break down the complex carbs or other molecules into smaller compounds that are easier to transport
facilitated diffusion