Cell Bio- Exam 2
Slide Set 5: Vesicular Traffic, Secretion, and Endocytosis
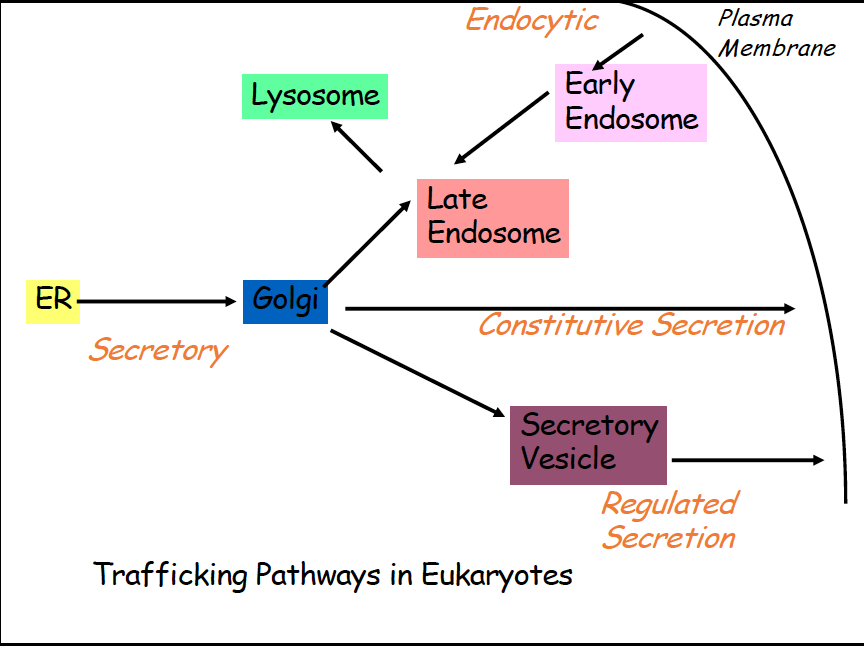
Vesicular Transport
- Proteins are synthesized in the ER, then are moved from ER to golgi, once mature proteins are formed, they need to leave the ER (Secretory)
- After golgi, they have multiple different pathways
- Constitutive secretion- constant secretion of proteins from cell, golgi to out of cell
- regulated secretion- secretory vesicle takes protein out of cell from golgi
- Endocytic- early endosome takes proteins from membrane to late endosome and then sometimes to lysosome
- Microscopy study with GFP
- studied
- use temperature,
- you can track proteins via fluorescent microscopy
- results:
- tracking total fluorescence signal over time
- Oligosaccaride modification
- mannose trimming occurs when oligosaccaride moves from ER to golgi
- treated with endoglycosidase D which cleaves sugar from protein
- Vesicle Budding and Fusion
- transport vesicle leaves donor compartment
- transport vesicle fuses with target compartment
- Coated Vesicle Budding
- SNARE protein helps transport vesicles recognize target membranes
- membrane cargo protein and soluble cargo protein bind together
- coat proteins surround vesicle
- Uncoated vesicle fusion
- V SNARE proteins will interact with T SNARE proteins on membrane
- , assists with docking
What is the mechanism by which vesicles are formed?
- Three types of coated vesicles
- Clathrin coated - helps with transport from trans golgi network to late endosome and helps transports obj entering the cell via endocytosis
- have heavy and light chains, as well as binding site for assembly particles
- soccer ball structure
- Functions:
- help form
- coat subunits bind to surface of donor membrane
- clathrin and other proteins help form bud/vesicle and help with the mechanical force of budding off
- capture membrane receptors
- adaptin helps transmem receptor bind to coating proteins
- certain aa are carried that signals adaptin to bind, these are then phosphorylated
- polymerizes around the neck and then hydrolyzes GTP, conformational change initiated in dynamin that stretches vesicle neck until the vesicle pinches off
- coatomer coated
- intra golgi traffic, golgi to ER
- plays a role in coat formation
- i
- coatomer coated
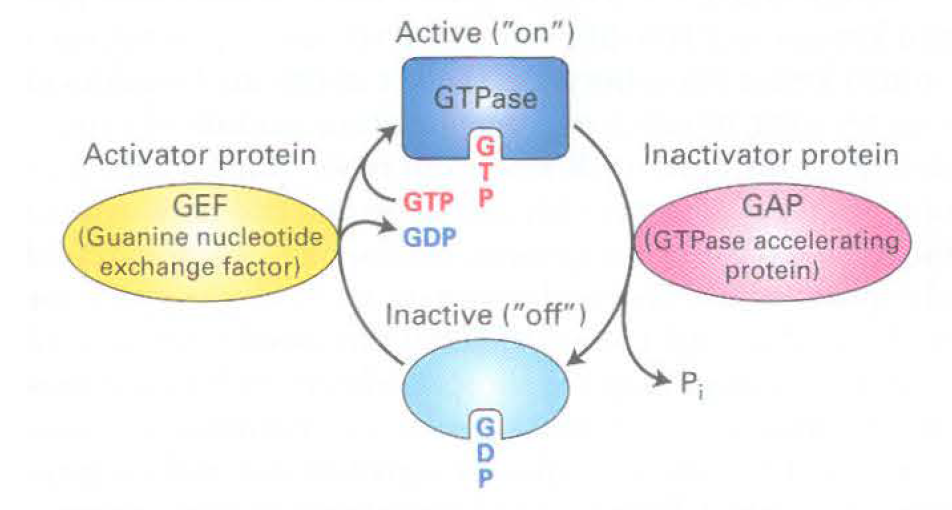
- GTPases
- Active- when protein binds to GTP
- Sar 1 initially binds to GTP, then binds to Sec 12 to hydrolyze GTP, then recruits COP2 components to have GTP bound to mem
- inactive- off, GDP bound
- - also a GTPase,
What are the molecular signals on vesicles that cause them to bind only to the appropriate target membrane?
- play a role in vesicle traffic and fusion
- generate tight interactions, help vesicles fuse to the donor membrane
- RAB GTPase
- donor mem: RAB receptor, vesicle: RAB
- mediate diff transport vesicles fused to diff transport membranes
- many diff RABs in eukaryotic cells
How do transport vesicles and their target organelles fuse?
- , will help recognize membrane
- Vesicle Fusion Machinery
- Vesicle Docking: V SNARE and T SNARE associate, RAB binds to RAB receptor
- Assembly of
- SNAP 25- snare complex, includes
- generates strong force to help fusion to the membrane
- Membrane Fusion
- proteins work to untwist SNAP 25
- fusion of membranes occurs
- Disassembly of SNARE complexes
- SNARE complexes disassociate and are free for another round of vesicle fusion, RAB also disassociates from the RAB effector
Steps in Secretory Pathway cont
- Vesicular Transport from ER to Golgi
- protein always goes from cis to trans face of golgi
- cis cisterna→ medial cisterna → trans cisterna
- - four aa, KDEL; if added at c term of protein it will return to ER from cis golgi bc it will bind to place on cis golgi and be recognized
- and other mods in golgi
- removal of 3 mannose residues in cis golgi )
- protein moves to medial golgi by cisternal maturation
- 3 GlcNAc residues added , 2 more mannose removed, single fucose is added
- processing completed in trans golgi by addition of 3 galactose residues and linkage of N-acetylneuraminic acid residue to each galactose

- helps protein become hydrophilic→ aids in folding
- aid in transport (rarely- targeting to lysosome)
- resistance to proteases (stability)
- protein protein interactions
- Vesicular sorting at trans- golgi network
- Vesicular Trafficking to Final Destination (golgi to ___)
- Endosome
- Plasma Mem
- constitutive secretion- unregulated membrane fusion
- regulated secretion- regulated membrane fusion
- Lysosome
- some proteins go here
- very acidic environment
- v class pumps used with ATP to pump proton inside
- lysosomes form a functional hub for cellular trrafficking pathways
- ER→ Golgi→ lysosome
- Pinocytosis→ lysosome
- Phagocytosis→ lysosome
- autophagy→ lysosome
- receptor on trans golgi network that will bind to M6P and will incorporate into vesicle and then will go to late endosome
- if pH low in late endosome, M6P transferred to lysosome
- Lysosomal Storage diseases
- can be due to absence of 1 or more lysosomal hydrolases or the mistargeting of lysosomal hydrolases
- characterized by tissue destruction or accumulation of undigested macromolecules
- Endocytosis
- goes through plasma mem, through early endosome then late endosome, then lysosome
- -
- very tiny things; proteins, lipids. Goes through
- continuous process, rate depends on cell type
- pinocytotic vesicle forms from clathrin coated pits in plasma mem
- -
- large things like bacteria;
- feeding for lower single celled euks
- multi celled orgs- used as a defense against invading microbes
- requires surface receptors, triggered event
- -
- from ER, if we do not need certain organelles anymore,
- LDL Uptake
- LDL- byproduct of fat transport, have ApoB protein
- vesicle begins to form with help of clathrin coat
- transported to early endosome→ late endosome→ lysosome
- Disorders- LDL receptor missing, receptors do not associate with clathrin coat
- Fate of cell surface receptors after endocytosis
- recycling of receptor to same domain
- receptor transported back to surface of membrane and pH will change→ receptor ready to bind to another LDL particle
- degradation of receptor after endocytosis
- in lysosome
Slide Set 6: Microfilaments
The Cytoskeleton
- Functions of cytoskeleton
- cell shape, mvmt, and contraction
- organelle mvmt and organization
- cell division
- intracellular org and vesicle mvmt
- interacting with signaling pathways
- basically like the bones of the cell
- Components
- Microfilaments
- actin filaments, thinner
- Microtubules
- tubulin dimers, thicker
- Intermediate filaments
- various, diff proteins combined together
- Cell signaling
- signals tell cytoskeleton abt organization and mvmt of organelles as well as changes in cell shape, mvmt, and contraction
Actin Microfilaments
- Functions
- org of intracellular organelles and transport of vesicles (myosin)
- intracellular mobility (bacteria)
- cellular stability
- cellular motility
- muscle contraction
- Lamellipodium
- supported by growth of actin filaments, generates a protrusion structure to adhere to surface and move cell forward
- Polymerization and Dynamics
- 1 actin filament= 2 strands
- one + end (0.12 M), one - end (0.6)
- g actin is monomer, microfilament polymer of actin
- ATP binding cleft in actin structure
- alpha, gamma, and beta actin: all associated with diff structures
- Actin Binding Proteins
- Polymerization- Profilin and Thymosin B4
- of ATP
- Crosslinking- Filamin
- Motor Proteins- myosin
- stability/cap end of filaments- capz and tropomodulin
- Actin based Motility
- will form
- actin binds to structure and elongation commences
- and from cell to cell→ hijack actin machinery and polymerize it to move around
- new filament assembly from preexisting filaments
- until capped by Cap Z
- which enhances depolymerization at the - end of the filaments
- this process
- Toxins that perturb pool of actin monomers
- stabilizes and binds actin dimers, r
- types of lateral attachment of microfilaments to membranes
- ankyrin- binds to Band 3 and then spectrin, forms network
- band 4.1
- Actin Motor Proteins
- can bind to actin and help generate contraction in muscle cells
- diff myosin has diff amts of each
- myosin heads can bind to ATP and actin
- works with contractions
- , head grp rotated into position to bind,
- power stroke occurs,
- Step size vs neck length
- is myosin step size/velocity proportional to neck length?
- contractile ring
- myosin 2 takes a large part in forming when cells are splitting, myosin 1 is on outside of cells
- Sarcomere (not protein, just structure of skeletal muscle)
- vertical component is Z band, in between is A band, myosin in between actin filaments
- actin end facing inside is - end
- sarcoplasmic reticulum- specialized region of the ER, regulates and stores Ca (Ca helps muscle cells to contract)
- Cap Z- binds to + end of actin
- Tropomodulin- binds to - end of actin
- Nebulin- binds to side of actin filaments
- Titin- binds to myosin and Z disk proteins
- dominant active rho- always keep making actin
- - filopodia formation
- guys see a Rac and are activated
- - lamellipodia formation
- Stress fiber formation
- leads to
Slide Set 7: Microtubules and Intermediate filaments
MIcrofilaments vs Microtubules vs Intermediate filaments
- microfilaments
- actin binds ATP
- form rigid gels, networks, and bundles
- tracks for myosin
- contractile machinery and network at cell cortex
- rigid and not easily bent
- organization for long range organelles
- Intermediate filaments
- great tensile strength, less dynamic, unpolarized
- no motors
- cell and tissue integrity
Microtubules
- play a role in….
- organization of organelles and transport of vesicles
- mvmt of cilia and flagella
- nerve cell, RBC, and flagellar structure
- alignment and separation of chrom during mitosis
- Two populations of microtubules
- Unstable short lived- assembles and disassembles rapidly
- stable and long lived- remain polymerized for a long time (sperm flagella, RBC, nerve cells)
- Polymerization and Structure of Microtubules
- Structure
- tubulin has alpha and beta parts
- microtubules made up of 13 protofilaments → singlet
- can have doublets (cilia/flagella) and triplets (basal bodies and centrioles) as well
- Polymerization
- MTOC-any structure used by cells to nucleate and organized microtubules
- centrosome falls into this category
- gamma tubulin ring nucleates microtubule assembly
- Dynamics of Microtubules
- Length over time: Assembly stage→ Catastrophe stage→ Disassembly stage→ Rescue Stage
- assembly
- then form
- GDP microtubule is the rest
- GTP cap bc
- \
- end more smooth (assembly), - end more rough (disassembly)
- Disassembly and reassembly of microtubules
- cool to 4 deg, microtubule will disassemble
- warm to 37 deg the microtubule will repolarize
- Drugs that disrupt microtubule dynamics
- causes depolymerization
- - stabilize the microtubule structure
- Binding Proteins
- similar to taxol
- MAP2- longer
- Tau- shorter
- +TIPS
- can regulate + end of microtubules
- Motors
- ferry cargo around the cell
- have light chain, bind to ATP for energy resource
- bind to microtubule with head groups, bind to vesicle via kinesin receptor
- hydrolyze ATP to drive mvmt
- Kinesin 1 and 2- organelle, mRNA, and chromosome transport
- Process
- first head group, no ATP,
- and then process restarts
- Power stroke of dynein- ATP hydrolysis causes change in orientation of head→ mvmt of MT
- LIS1 protein- interact with ATPase domain of dynein to elongate power stroke
Intermediate filaments
- heterogeneous
- great tensile strength
- no known motors use them as tracks
- more stable than filaments or tubules
- no intrinsic polarity
- made up of protofilaments that can form diff structures