Visual Systems II
LO: Describe the organization of retinal inputs to the LGN (p.g. 338-341) ✅
Segregation of Input by Eye & by Ganglion Cell Type
LGN neurons receive synaptic input from retinal ganglion cells
Geniculate neurons project an axon → primary visual cortex vis optic radiation
Segregation of LGN neurons means retinal info types are kept separate
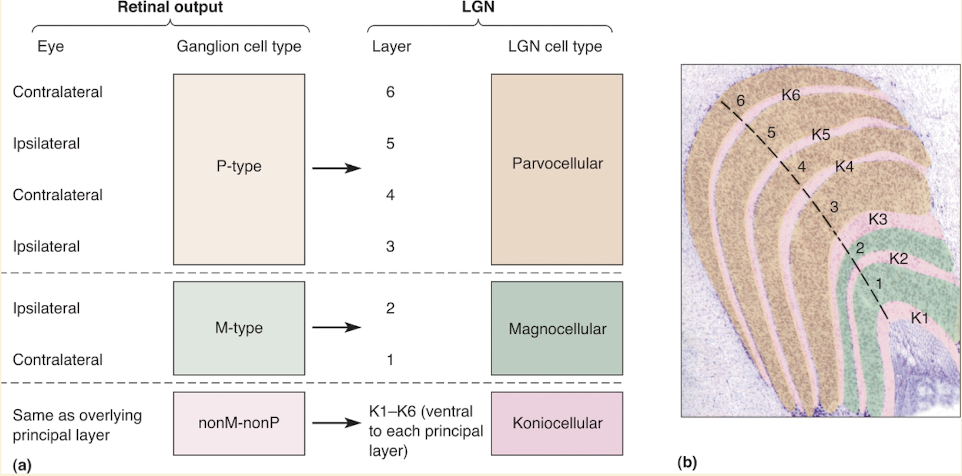
M-type, P-type, & nonM-nonP ganglion cell axons synapse on cells in diff LGN layers
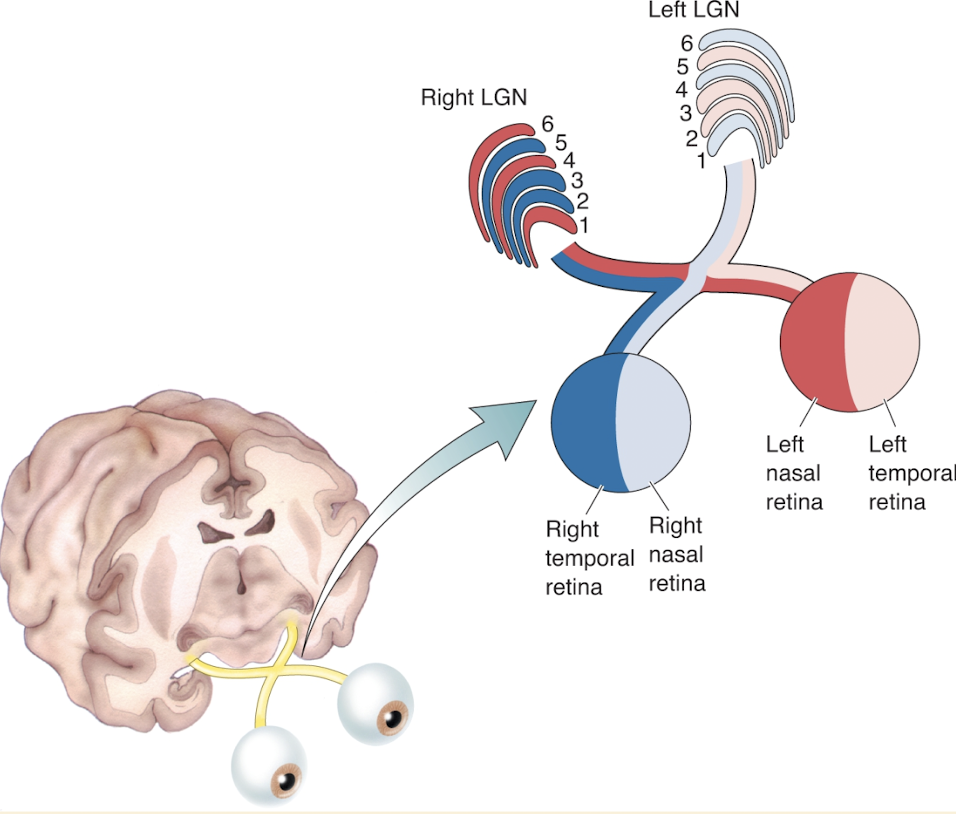
Right LGN receives info ab left visual field
Left visual field is viewed by nasal left retina & temporal right retina
@ LGN input from 2 eyes separate
In right eye axons synapse on LGN cells in layers 2, 3, & 5
Left eye axons synapse on cells in layers 1, 4, & 6
Ventral layes 1& 2 contain larger neurons
Called magnocellular LGN layers
Dorsal layers 3 through 6 contain smaller cells
Called parvocellular LGN layers
Koniocellular LGN Layers
Numerous tiny neurons lie ventral to each layer
Layers K1-K6
Receives input from nonM-nonP types of retinal ganglion cells
Each layer gets input from same eye as overlying M or P layer
e.g. K1 receives input from contralateral eye like layer 1 neurons do
Projects to visual cortex
In LGN diff info from three categories of retinal ganglion cells from two eyes remain segregated
Receptive Fields
By inserting micro electrode into LGN can study AP discharges of geniculate neuron in response to visual stimuli & map receptive field
receptive field near identical to ganglion cells
Within all layers of LGN neurons are activated only by one eye & ON-center & OFF-center cells are mixed
Nonretinal Inputs to LGN
Retina is not main source of synaptic input to LGN
Also receives input from thalamus & brains stem
Neurons in brain stem are associated w/alertness & attenriveness
Flash of light when startled in dark room → result of activation of LGN neurons by pathway
Major input is primary visual cortex
80% of excitatory synapses
Role of PVC has not been indentified
One hypothesis: top down modulation from visual cortex to LGN gates subsequent bottom up input from LGN back to cortex
LO: Compare and contrast P-type and M-type ganglion cells ✅
P-Type
Project exclusively to parvocellular LGN
Parvocellular LGN cells have
Small center surround receptive fields
Respond to stimulation of receptive field centers w/ increase in frequency of APs
Color oppocency
M-Type
Project entirely to the magnocellular LGN
Magnocellular LGN neurons have
Large center-surround receptive fields
Respond to stimulation of receptive field centers w/burst of APs
Insensitive to differences in wave length
Like M-type ganglion cells
Non-M-nonP Type (Koniocellular Layers)
Have either light/dark or color opponcency
LO: Predict the site of a lesion in the retinofugal pathway based upon the visual field deficit
In class
LO: List the structures of the retinofugal pathway including non-thalamic targets and their purpose
LO: Explain retinotopy (p.g. 342) ✅
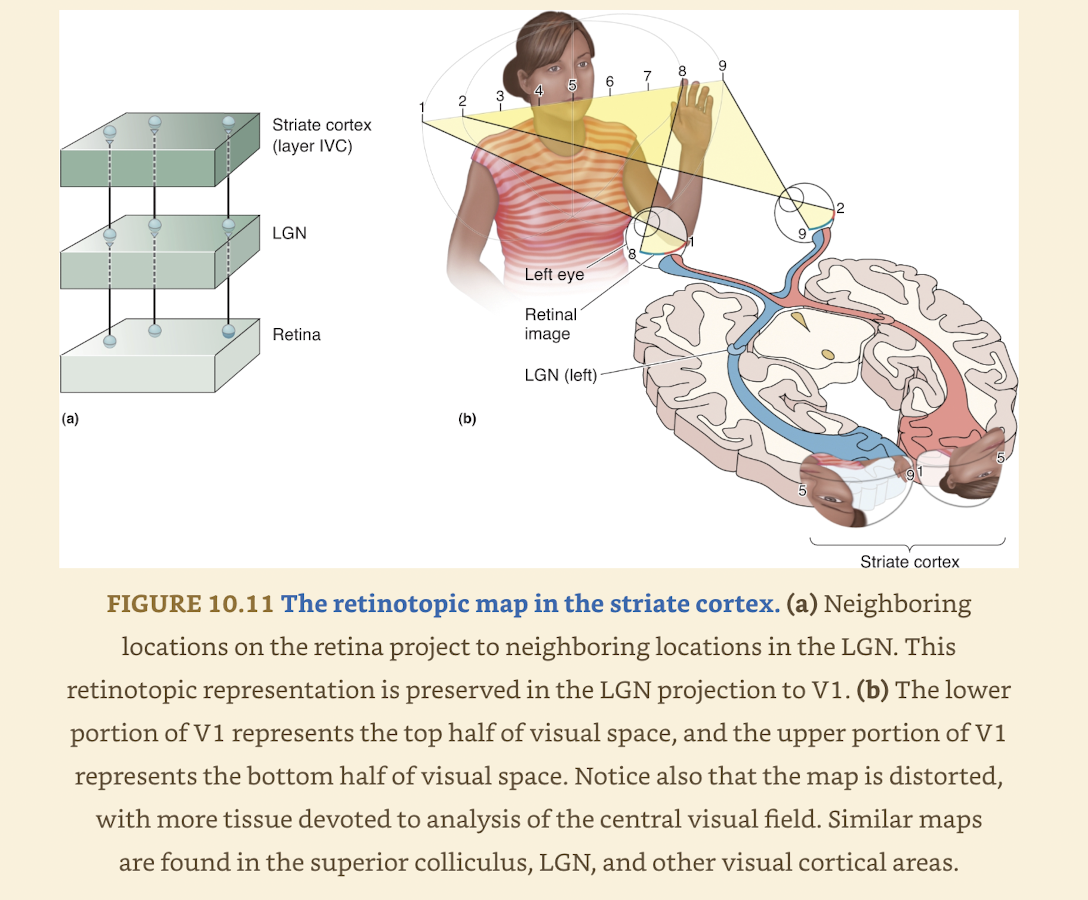
Projection starting in retina → LGN & V1 illustrates Retinotopy
Retinotopy: organization where neighboring cells in retina feed info to neighboring places in target structures
In this case LGN & striate cortex
In this way 2D surface of retina is mapped onto 2D surface of next structures
Three important things to remember:
Mapping of visual field is often distorted bc visual space is not sampled uniformly by cells in retina
Central few degrees of visual field are overrepresented (magnified)
Discrete point of light can activate many cells in retina & more cells in target structure
Not real map based on brains interpretation of distributed patterns of activity
LO: Explain the characteristics of receptive fields in the striate cortex
Anatomy of the Striate Cortex
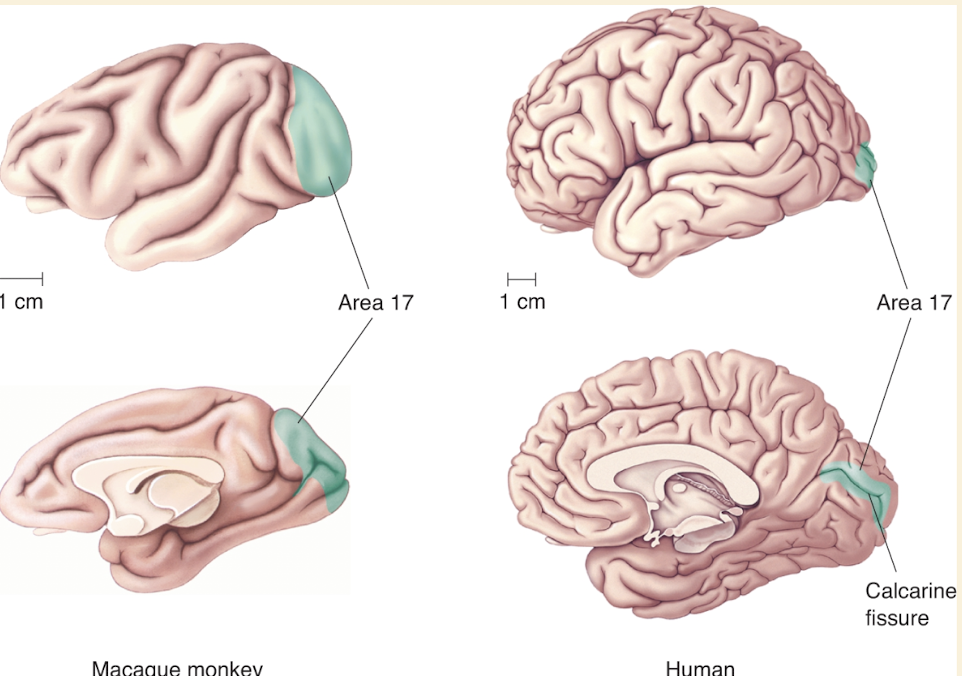
Primary visual cortex = area 17
Other terms: V1 & striate cortex
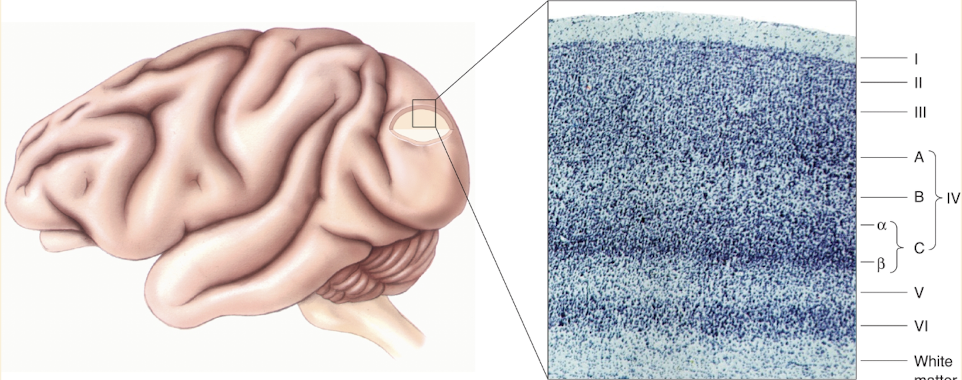
Lamination of Striate Cortex
Cell layers named by: VI, V, IV, III, & II
Layer I
under pia matter
devoid of neurons, mainly axons & dendrites of cells in other layers
9 distinct layers of neurons (even tho photo shows 6 ish)
Suggests division of labor in cortex
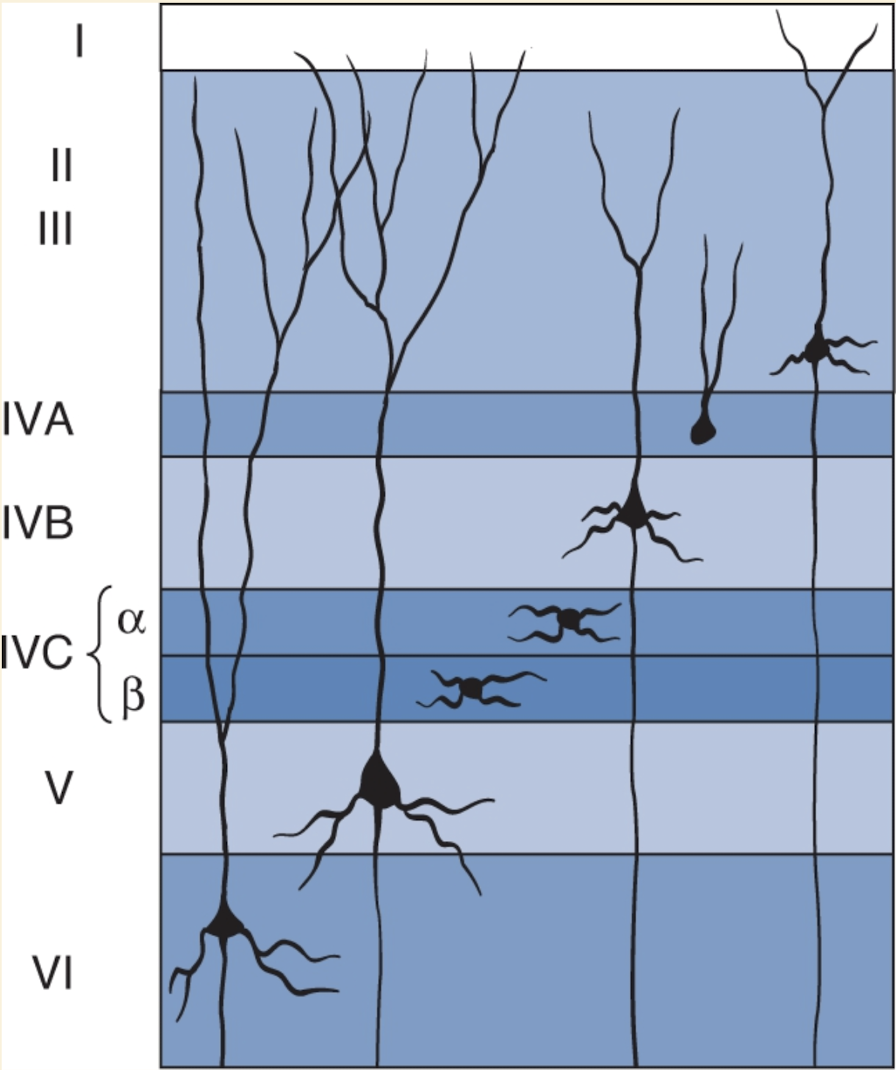
Spiny stellate cells: small neurons w/spine covered dendrites that radiate from cell body
Seen in two layer IVC
Local connects (except layer IVB)
Pyramidal cells
Outside IVC
Also covered w/spines & have thick apical dendrite ascending toward pia matter
Only send axons out to form connects w/other parts of brain
Inputs & Outputs
The striate cortex has distinct layers (lamination) similar to the LGN, but each layer has a different role.
Only some layers of the striate cortex receive input from the LGN or send output to other brain areas.
Most LGN axons end in layer IVC of the striate cortex.
Within layer IVC, there are two sublayers:
IVCα receives input from magnocellular LGN neurons.
IVCβ receives input from parvocellular LGN neurons.
These two sublayers form separate but overlapping visual maps of the visual field.
Koniocellular inputs from the LGN mainly go to layers II and III of the striate cortex.
Innervation of other Cortical layers from IVC
Most intracortical connections extend perpendicular to the cortical surface along radial lines running from the white matter to layer I.
These radial connections preserve the retinotopic organization established in layer IV (cells aligned vertically process information from the same part of the retina).
Example: a cell in layer VI receives input from the same retinal location as a cell above it in layer IV.
Some layer III pyramidal cells send horizontal collateral branches within layer III, creating lateral (horizontal) connections.
Radial connections and horizontal connections serve different functions in visual processing.
After information leaves layer IV, there is continued segregation of the magnocellular and parvocellular pathways:
Layer IVCα (receiving magnocellular LGN input) projects mainly to layer IVB.
Layer IVCβ (receiving parvocellular LGN input) projects mainly to layer III.
In layers III and IVB, axons can form synapses with pyramidal cell dendrites from all cortical layers.
Ocular Dominance Columns
Research Question: How are left and right eye LGN inputs arranged in the striate cortex — intermixed or segregated?
Researchers: David Hubel and Torsten Wiesel (Harvard Medical School, early 1970s).
Method:
A radioactive amino acid was injected into one eye of a monkey.
The amino acid was incorporated into proteins by retinal ganglion cells.
These proteins were anterogradely transported down the ganglion cell axons into the LGN.
Postsynaptic LGN neurons (receiving input from the injected eye) absorbed the radioactive proteins.
These LGN neurons then transported the proteins to their axon terminals in layer IVC of the striate cortex.
Visualization Technique:
Thin sections of striate cortex were covered with photographic film and developed using autoradiography.
The silver grains on the film marked the locations of radioactive LGN axon terminals.
Findings:
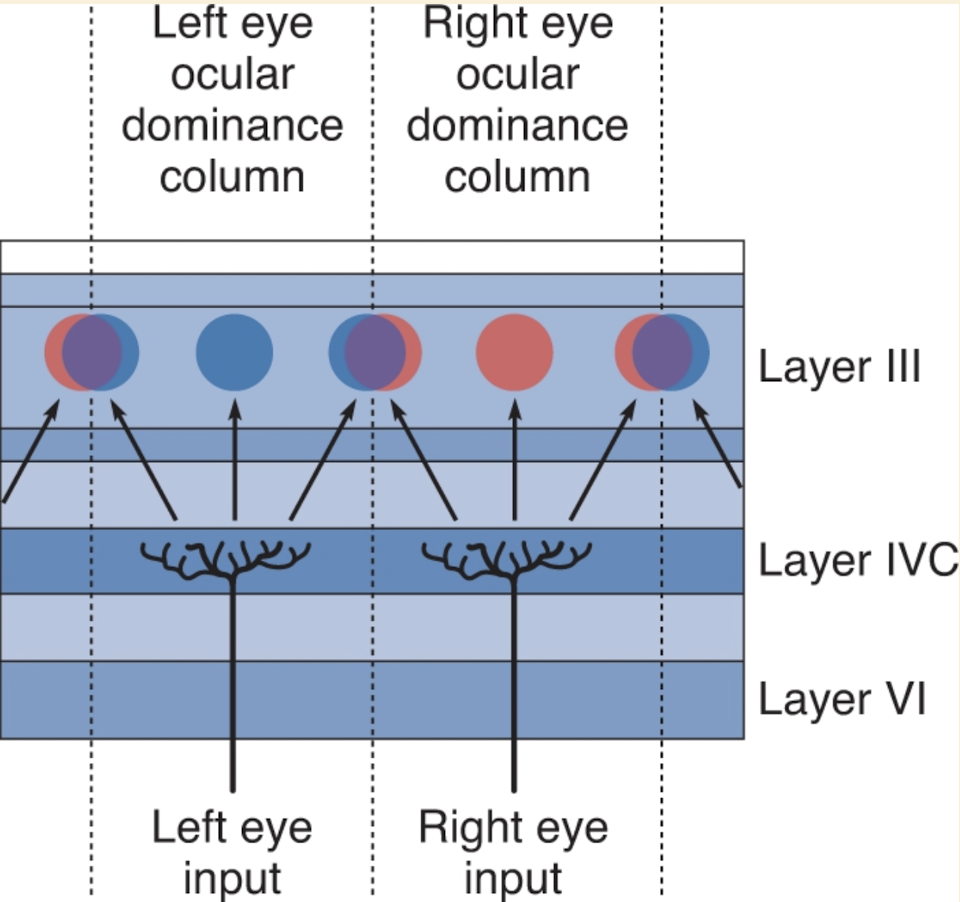
In cross-sections (perpendicular to cortex), radioactive terminals appeared in equally spaced patches (~0.5 mm wide) within layer IVC, rather than in a continuous spread.
In tangential sections (parallel to layer IV), left and right eye inputs appeared as alternating bands — resembling zebra stripes.
Thus, inputs from the two eyes remain segregated in layer IV, just as in the LGN.
Integration of Inputs:
Layer IVC stellate cells send radial axons mainly to layers IVB and III.
In layers II, III, V, and VI, neurons begin to receive input from both eyes.
A neuron may receive input from both eyes but is usually “dominated” by one (receiving stronger input from one eye).
Example: a neuron above a left-eye patch in layer IVC receives input from both eyes but is left-eye dominant.
Ocular Dominance Columns:
Alternating bands of neurons dominated by left or right eye extend through the full thickness of the striate cortex.
These bands are called ocular dominance columns.
Cytochrome Oxidase Blobs
Layers II and III of the striate cortex (V1) play a major role in visual processing, sending most of the output from V1 to other cortical areas.
Anatomical studies show that V1 output originates from two distinct populations of neurons in these superficial layers.
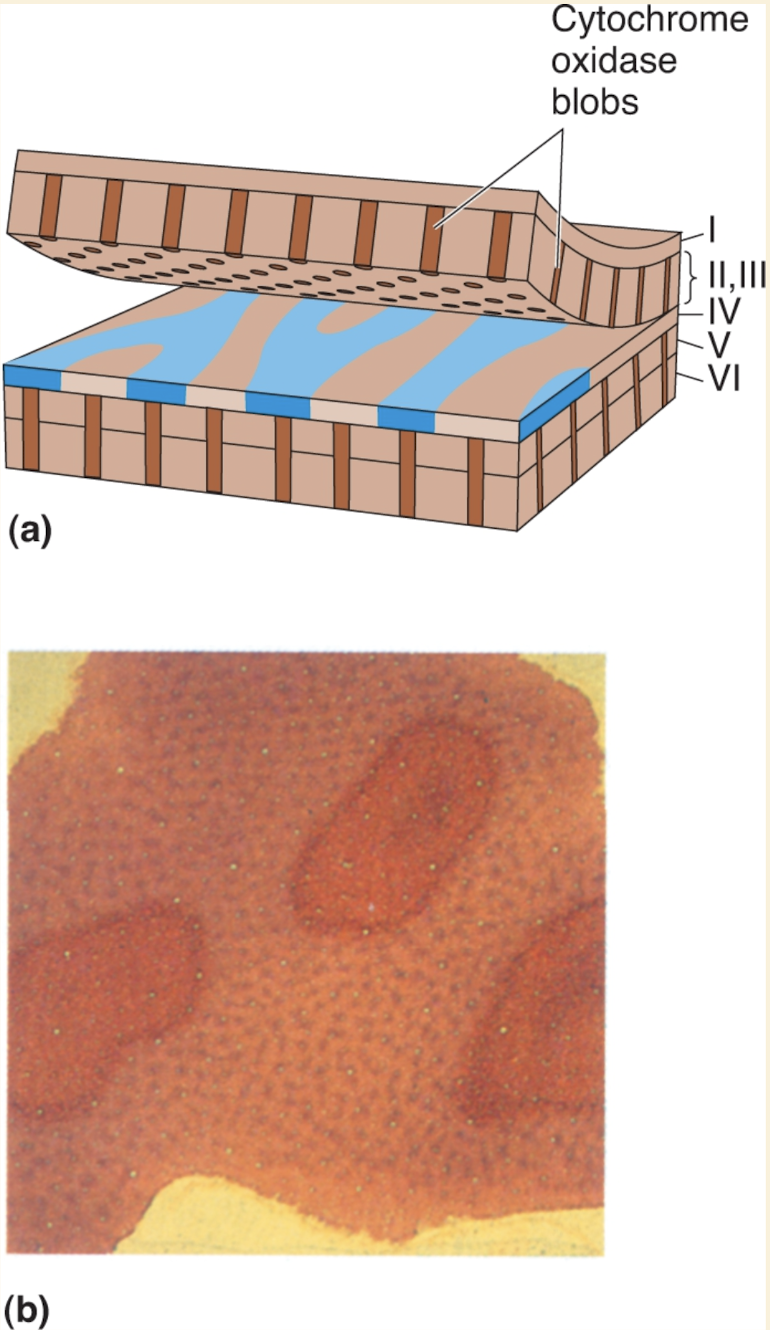
When V1 tissue is stained for cytochrome oxidase (a mitochondrial enzyme involved in metabolism), the stain appears non-uniformly distributed in layers II and III.
In cross sections, the cytochrome oxidase stain forms vertical columns (colonnades) that run through layers II and III, and also appear in layers V and VI.
In tangential sections (sliced parallel to layer III), these columns appear as spots, resembling the spots of a leopard.
These cytochrome oxidase–rich columns are called blobs.
Blobs are organized in rows, each one centered on an ocular dominance stripe in layer IV.
The regions between blobs are known as interblob regions.
Blobs receive direct LGN input from the koniocellular layers, and also receive parvocellular and magnocellular input indirectly from layer IVC of the striate cortex.
Binocularity
In V1, there is a direct correspondence between the anatomical arrangement of connections and neuronal responses to light in the two eyes.
Neurons in layers IVCα and IVCβ receive afferents from LGN layers representing either the left or right eye.
Physiological recordings show these neurons are monocular, responding to light from only one eye.
Axons leaving layer IVC diverge to more superficial layers, mixing inputs from both eyes.
Microelectrode recordings confirm that most neurons in layers superficial to IVC are binocular, responding to light from either eye.
Ocular dominance columns correlate with neuronal responses:
Neurons above centers of ocular dominance patches in layer IVC are dominated by the same eye represented in IVC, even though they are binocular.
In regions with more equal mixing of left and right eye inputs, superficial neurons respond equally to both eyes.
Binocular receptive fields: neurons in superficial layers have two receptive fields, one in each eye.
Retinotopy is preserved: the two receptive fields are aligned to the same point in the contralateral visual field.
Functional significance: binocular receptive fields are essential for depth perception and stereoscopic vision, allowing humans to form a single coherent image and perform fine motor tasks requiring depth judgment (e.g., threading a needle).
Orientation Selectivity
Receptive fields in retina, LGN, and layer IVC are mostly circular, responding best to a spot of light matching the receptive field center.
Outside layer IVC, many V1 neurons respond better to elongated bars of light, rather than small spots.
Orientation of the bar is critical:
Each neuron has a preferred orientation that elicits the strongest response.
Bars perpendicular to the preferred orientation produce weaker responses.
Orientation selectivity: the property of neurons responding maximally to bars at a particular angle.
Most V1 neurons outside IVC (and some within) are orientation selective.
Optimal orientation can be any angle around 360°.
Radial organization (orientation columns):
As a microelectrode moves perpendicular to the cortical surface, the preferred orientation of neurons remains constant across layers II–VI.
Such vertically aligned neurons form an orientation column.
Tangential organization (across the surface):
As a microelectrode moves parallel to the cortex, the preferred orientation gradually shifts.
Optical imaging reveals a mosaic-like pattern of orientation preferences across the striate cortex.
Depending on the angle of traversal, preferred orientation can rotate smoothly (like a clock sweep) or shift abruptly.
Spatial scale: a complete 180° shift in preferred orientation typically occurs over ~1 mm in layer III.
Key insight: V1 is organized into orientation columns and a systematic mosaic, allowing neurons to represent all possible orientations in the visual field.
Simple and Complex Receptive Fields
LGN neurons have antagonistic center-surround receptive fields, responding best to small spots in the center versus larger spots that also cover the surround.
Binocularity in V1 neurons arises because binocular neurons receive afferents from both eyes.
Orientation and direction selectivity are more complex and involve specific receptive field arrangements:
Many orientation-selective neurons have elongated receptive fields along a specific axis.
These fields have ON-center or OFF-center regions flanked by antagonistic areas, analogous to LGN center-surround fields.
Cortical neurons appear to receive convergent input from multiple LGN cells aligned along the same axis.
Simple cells:
Defined by segregated ON and OFF regions in their receptive fields.
This linear arrangement makes them orientation selective.
Example: a bar of light in the middle evokes an ON response, flanking positions evoke OFF responses.
Complex cells:
Lack distinct ON and OFF regions.
Respond to ON and OFF stimuli anywhere in their receptive field.
Likely constructed from multiple like-oriented simple cells, though this is debated.
Both simple and complex cells are typically:
Binocular
Orientation selective
Sensitive to stimulus direction and various color inputs.
Blob Receptive Fields
Blob vs. interblob regions in striate cortex: distinct cytochrome oxidase labeling suggests potential functional differences between neurons in these regions.
Interblob neurons:
Exhibit binocularity, orientation selectivity, and direction selectivity.
Include simple and complex cells.
Some are wavelength sensitive, some are not.
Blob neurons:
Receive direct input from koniocellular LGN layers and magnocellular/parvocellular input via layer IVC.
Early studies suggested they are generally wavelength sensitive and monocular, lacking orientation and direction selectivity, resembling LGN input.
Receptive fields can be:
Circular, with color-opponent center-surround (like parvocellular/koniocellular LGN).
Red–green or blue–yellow center only.
Double-opponent cells: color-opponent center and surround.
Recent studies:
Both blob and interblob neurons show selectivity for orientation and color, suggesting more overlap than previously thought.
Blob cells have higher average firing rates than interblob cells, corresponding to greater cytochrome oxidase activity.
Conclusion:
Despite anatomical differences, there is no simple distinction between receptive field properties of blob vs. interblob neurons.
Neurons sensitive to wavelength are important for color perception, but the exact role of cytochrome oxidase blobs in color vision remains uncertain.
NOTE: Did not read section parallel pathways and cortical modules
LO: Describe the hierarchy of receptive fields in the visual system - idek
LO: Describe the functional differences between the dorsal stream and the ventral stream ✅
Dorsal Stream
Area MT
Specialize processing of object motion
Receives retintopically organized input from V1, V2 & V3
Mainly V3, associated with V2 & V1
Innervated by cells in layer IVB
Large receptive fields
All the cells are direction selective
Neurons respond to types of motion we perceive (optical illusions)
Dorsal Area & Motion Processing
Specialized movement sensitivity
e.g. MST has cells selective for linear motion, radial motion, and circular motion
Three roles of MST proposed
Navigation
Directing eye movement
Motion perception
Ventral Stream
Progression of area form V1, V2, and V4
Mainly V4, associated with V2 & V1
V4 receives input from blob & inter blob via relay in V2
Neurons have larger receptive fields, cells are both orientation selective & color selective
Achromatopsia: partial or complete loss of color vision despite presence of normal functional cones
Associated w/cortical damage in occipital & temporal lobes
Area IT
Output of V4
Appears to be farthest extent visual processing in ventral stream
Variety of colors & abstract shapes are good stimuli for cells in IT
Output from IT → temporal lobe structures
May be important for visual perception & visual memory
Fusiform face area
Crucial for face recognition
Face-selective cells
Prosopagnosia: difficulty of recognizing faces even though vision is normal
Results from stroke, damage to extra-striate visual cortex