


MCAT Biochemistry
Class 1 - 01/06/2024:
Macromolecules - are polymers made from monomers - enzymes that make polymers are polymerases via reactions called polymerization.
Proteins: made up of amino acids(20 kinds)
Have an N-C-C backbone, amine group, carboxylic group, and variable group
Bond together by a peptide bond(formed by dehydration synthesis - loss of H2O)
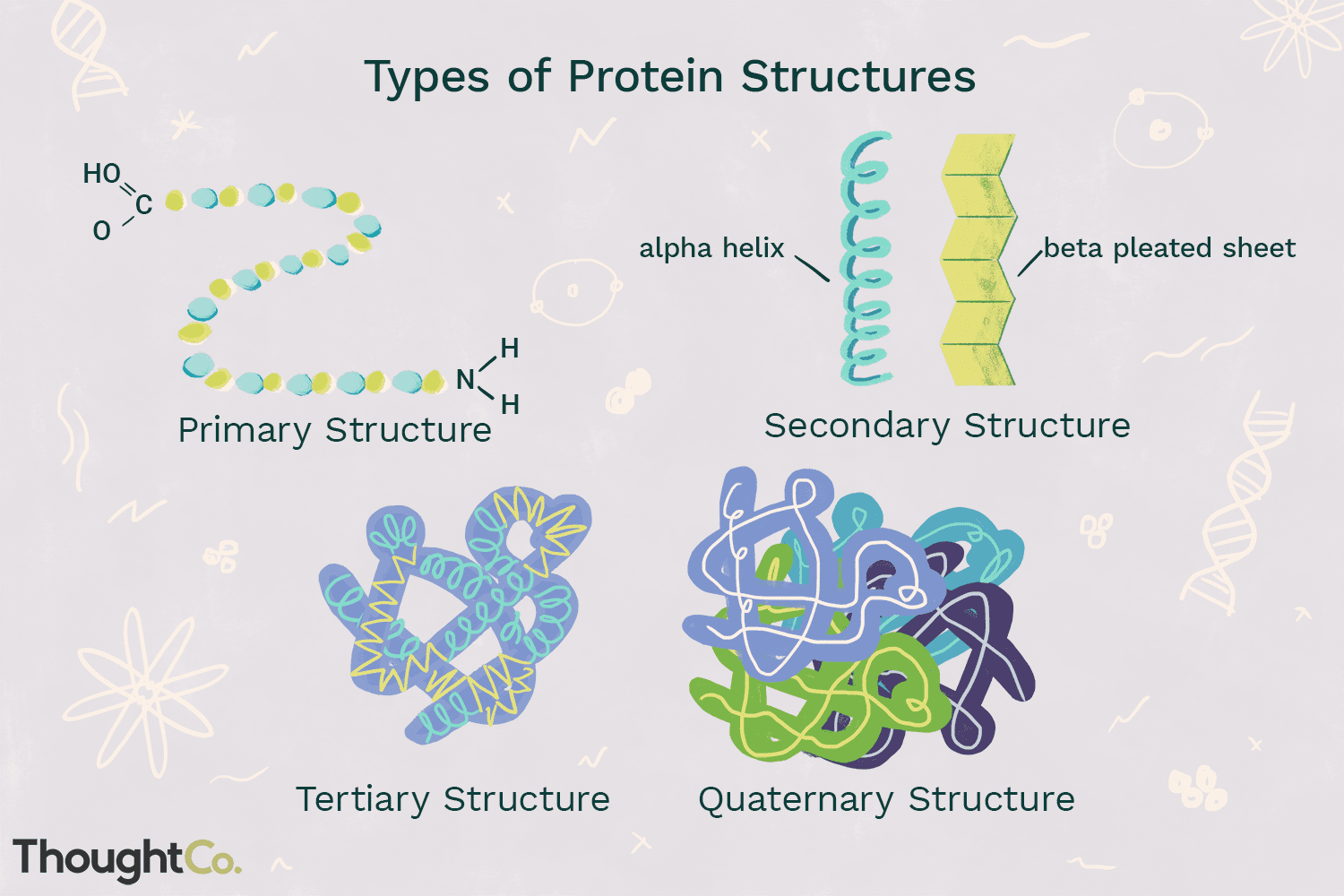
4 types of structures
Primary = amino acids joined
Secondary = alpha-helix or beta-sheets
Tertiary: formation of a polypeptide and side chain interactions - inner core becomes hydrophobic and outer becomes hydrophilic
Non-covalent interactions: non-polar/non-polar, polar neutral/polar neutral, acid/base(charged)
Covalent: disulphide bridges(harder to break)
Quaternary structure: side chain interactions between different polypeptides - subunits come to form larger units
Carbohydrates: from monosaccharides to disaccharides to polysaccharides;
monosaccharides - CnH2On - 3 common are glucose, fructose, galactose - ribose and deoxyribose
Disaccharides - 2 monosaccharides - 3 common = maltose, sucrose, lactose - C12H22O11 formula
Polysaccharides: many monosaccharides - 3 common are glycogen, starch, and cellulose - function as an energy source
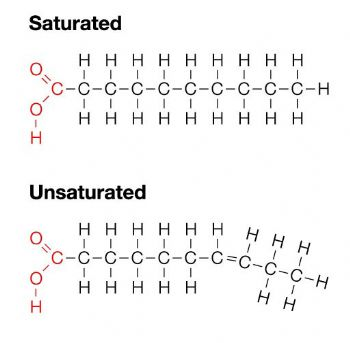
Lipids: the fats - made of a hydrocarbon structure(many C and H) - saturated fatty acids are solid at room temperature
Unsaturated are liquid at room temperature due to the double bond
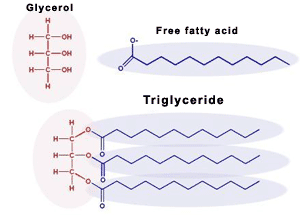
Triglyceride: 3 fatty acids combined
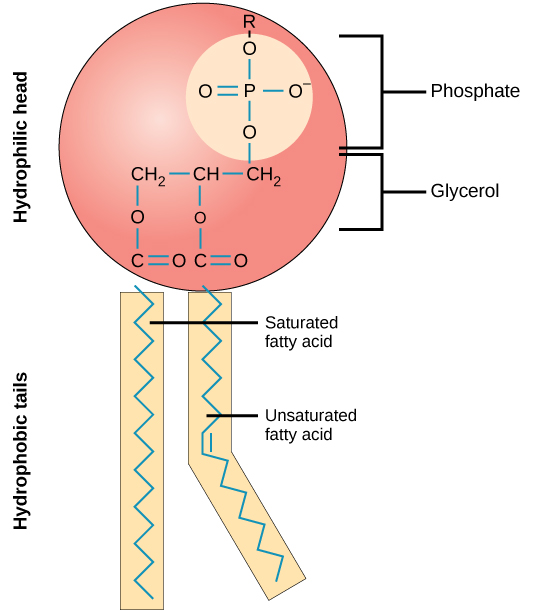
Phospholipids: 2 lipid structures and one phosphate - form the lipid bilayer due to having polar and non-polar sides
Terpenes: built from isoprene structures and need at least 2 of them - terpenes form waxes and lipid rings like vitamin A
Cholesterol and steroid hormones - 3 six-carbon rings and 1 five-carbon ring)
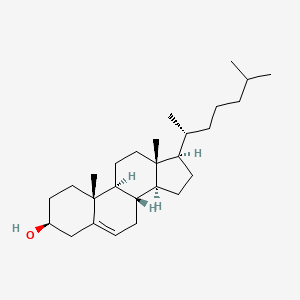
Thermodynamics: delta G = delta H - TdeltaS
G = Gibbs free energy
H = enthalpy (potential E)
T = temperature
S = Entropy (kinetic E)
When G<0 = negative G, spontaneous - gives E - exergonic
When G>0 = positive, non-spontaneous - needs E - endergonic
When G = 0, equilibrium
Reaction Coupling - using ATP as a source of energy - a very favourable reaction is used to drive an unfavourable one
ATP = ADP + Pi → very exergonic
Exergonic = giving off E
Endergonic = using up E
Chemical Kinetics: the study of reaction rates - all reaction rates proceed through a transition state which tends to be unstable
Activation E = is the required E to produce the TS
if Ea is High = slow rate
if Ea is low = faster rate
Reaction Coordinate Graph - shows the energy vs reaction coordinates over time - the smaller the Ea, the better
We can make the Ea smaller using catalysts - speeding the reaction up by stabilizing TS and reducing Ea
Enzymes: a physiological catalyst - works to speed up a reaction by increasing the rate of reaction, not used up in a reaction, and must be specific
Structure: an enzyme has an allosteric site and an active site - the active site is where the substrate binds(where the reaction occurs) and the allosteric site is another place for enzyme regulation(inhibition or activation)
Two models - active site and induced fit; active is lock and key while the induced fit is when the enzyme needs to change shape to fit a substrate
Can perform both positive and negative feedback
Function: to speed up a reaction
Regulation: by many inhibitions ways, allosteric site, feedback inhibition
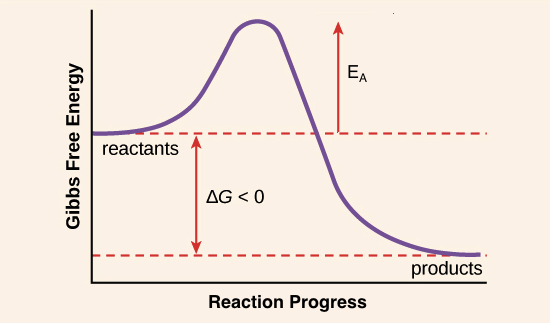
V vs. [S] Graph: reaction rate in Velocity vs the substrate concentration [S] → vmax is when the enzyme is saturated and depends on enzyme [C], and the [S] becomes constant - Vmax/2 is when the linear part of the graph is equal to [S]
Km is the substrate [S] required to reach ½ Vmax
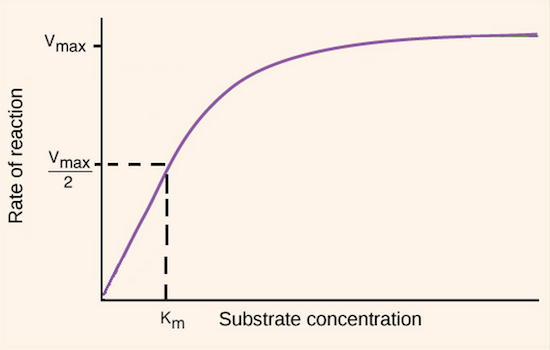
Enzyme Inhibition:
Competitive: compete for enzyme binding - same Vmax but the effect on Km is more since you need more substrate - a longer time to reach the same Km - binds at the active site - before substrate binds
Non-competitive: it affects the Vmax since we need more enzymes to deal with the substrate, but Km is unchanged since the active site is the same but prevents the activity of the enzyme- binds to the allosteric site before substrate binds
Un-competitive: it affects both the Vmax and Km since it binds to the allosteric site after the substrate is bound, which affects both enzyme performance and the amount of product being produced - binds to an allosteric site after the substrate binds
Mixed-Type Inhibition: binds at the allosteric site either when the enzyme is bound to the substrate or empty active site. Vmax will become lowered, but Km can vary whether enzyme bound or empty Active site
when bound to the substrate, Km decreases(like un-comp)
When empty active site, Km increases(like comp)
Lineweaver Burk Plots:
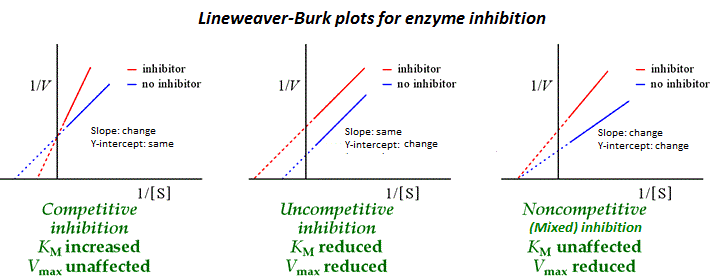
Class 2 - 08/06/2024:
Oxidation-Reduction Reactions - Redox
Oxidation is when you gain O, lose H and electrons
Reduction is when you lose O, gain H and electron
Cellular Respiration: When you convert sugar and O2 into carbon dioxide and water
a four-step process - glycolysis, PDC, Krebs, and electron transport
Glycolysis
processed in the cytoplasm and doesn’t need O2
all cells from all domains perform glycolysis → Sugar split into two pyruvate molecules and 4 ATP and 2 NADH formed at the end
Step 1: Got a phosphate from ATP and a glucose → Hexokinase → bam, G-6-P and ADP
Step 2: Got a G-6-P and a isomerization → Bam, F-6-P
Step 3: Got a F-6-P and an ATP → bam, F1-6-bp and ADP
Step 4: Split F1 into two to form 2×3CP
Step 5: add 2pi to 2×3CP → Form 2 PEP 2×3CP
Step 6: 2 PEP plus 2ADP → Pyruvate kinase → 2 Pyruvate
Mnemonic Glycolysis Products:
Gross Guys Favour Fat-Buns, Good Boys Prefer Pretty Pink Pyjamas
Glucose, G6P, F6P, F-1,6-Biphosphate; Glu-ald-phosphate, 1,3-Biphosphoglycerate, 3-Phosphoglycerate, 2-Phosphoglycerate, Phosphophenol pyruvate, Pyruvate
Mnemonic Glycolysis Enzymes:
Honestly, Ignoring Pencils And Getting Pens Make Everything Perfect
Hexokinase, Isomerase, PFK, Anolase, G3P dehydrogenase, Phosphogly-kinase, phosphoglycerate-Muctase, Enolase, Pyruvate Kinase
PDC - Pyruvate Dehydrogenase Complex: occurs in the matrix of the mitochondria and needs O2
Pyruvate is changed from being a 3C molecule to a 2 C molecule and CO2 and NADH are produced; this is from taking pyruvate and coenzyme A to make acetyl-coA
Oxidative Decarboxylation does this: release of CO2 and make NADH
Krebs Cycle: Occurs in the matrix of the Mitochondria and needs O2- Takes 2xacetyl-coA with oxaloacetate to make 2xCO2, NADH and FADH2
In order:
2C + Oxaloacetate → citrate(6C) → NADH + CO2 → 5C → NADH and CO2 → 4c → GTP → succinate → fumarate → malate → oxalacetate
Krebs mnemonic Substrates:
Can Interests Kindly Send Some Free Money Oop
Citrate, Isocitrate, α-Ketoglutarate, Succinyl-CoA, Succinate, Fumarate, muctase, Oxcaloacetate
Kreb’s mnemonic Enzymes:
Can Anyone Interested Kindly Send Some Free Money
Citrate synthase, Aconitase, Isocitrate dehydrogenase, α-ketoglutarate dehydrogenase, Succinyl synthase, Succinate dehydrogenase, Fumarase, Malate dehydrogenase
ETC and Oxidative Phosphorylation: occurs in the inner membrane of the mitochondria and needs O2
OP is the oxidation of NADH and FADH2 to make ADP into ATP → This makes energy move e- transport chain and pumps protons out of the mitochondria
ETC is a chain of 5 e- carriers that perform redox roles(cytochromes)
Step 1: NADH dehydrogenase → reduce NADH, pass e
Step 2: coenzyme Q → release FAD from FADH, pass e
Step 3: cytochrome C reductase → passes e to next
Step 4: cytochrome C oxidase → makes H2O and sends to ATP synthase to make ATP
ETC Mnemonic:
Norway’s Queens’ CyteR Can’t Oxidize ATP
NADH Reductase, CoQ, Cytochrome Reductase, CoC, Cytochrome Oxidase, ATP Synthase
Total ATP of respiration = 30 ATP made in Euk, and 32 in Prok
Fermentation: when no oxygen is available - no ETC, No oxidative phosphorylation, no Krebs, no PDC, no glycolysis
Take Pyruvate and form ethanol yeasts) and lactic acid(in muscles, other cells)
Gluconeogenesis: the formation of glucose from 2 pyruvates (reverse of glycolysis but some different unique enzymes) - Happens in the body when glycogen stores are depleted in the liver
2 pyruvate with pyruvate carboxylase forms 2 oxaloacetate
PEP carboxykinase makes 2 PEP
2 PEP is turned into 2×3CP to then F16CP
F16CP is then turned into F6CP to G-6-P
G-6-P with glucose-6-phosphatase to make glucose
Glycogenesis: the formation of glycogen from glucose
made by using G-6-P to G-1-P by phosphoglucomutase and then using UDP to make UDP-glucose which is turned into glycogen by glycogen synthase
Glycogenolysis: breakdown of glycogen to glucose
Here glycogen is phosphorylated into G-1-P and then isomerized to G-6-P to make glucose again
happens in the liver
Insulin increases when glucose is high to make glycogen - stores energy for later
Pentose Phosphate Pathway: takes G-6-P to form NADPH and ribose-5-phosphate. NADPH is important in its role of reducing the power of fatty acid synthesis and helps neutralize reactive O2 species as well as make the building block for nucleotides
Fatty Acid Oxidation: the fat digestion - the removal of 2C units as acetyl-CoA from a fatty acid and makes 1 FADH2 and 1 NADH - the acetyl is then moved to Krebs or ketone bodies
dietary fat chylomicrons move from the lymph system to the liver and organs and then undergo beta-oxidation which then turns the fatty acid into acetyl-coA
Fatty Acid Synthesis: uses high amounts of ATP and NADPH where 2C units are added to the chain until 16C fat is made
using acetyl-coA to make malonyl-CoA
Ketogenesis: during starvation, acetyl-coA turns into ketone bodies and can supply energy to the brain and lower blood pH
Protein Catabolism: break down of protein by proteases to amino acids
Metabolism: when the body is fed, glycolysis, glycogenesis, and fatty acid synthesis are favoured. When the body is starved, glycogenolysis, glucogenesis, and fatty acid oxidation are favoured.
Class 3 - 15/06/24:
Nucleotide: made up of sugar, base, and phosphates
Sugar = deoxyribose or ribose
Base = ACTG
Phosphates = 3 linked together
Nucleic Acid Structure: 5’-3’ linkage, antiparallel and complementary, phosphodiester bonds
Pyrimidines = U, T, C(smaller)
Purines = A, G(bigger)
A-T, C-G, A-U(in RNA)
DNA structure:
in prokaryotes, circular DNA genome, formed by methylation, and supercoiling
in eukaryotes, several linear chromosomes → (biggest)chromosome, to chromatin, to histones bound to make nucleosomes, to make smaller DNA strands(smallest)
Centromere: the middle of the chromosome where the spindle fibres attach to - made of heterochromatin and repetitive DNA sequences - short sequences repeat - both single and double-stranded DNA which can loop to form a knot at the end of the chromosome to stabilize it
Telomere: the ends of a eukaryotic linear chromosome - also has a hand in aging
DNA protection: the tighter it is, the less likely it will be uncoiled
The Central Dogma: DNA leads to transcription to RNA that leads to the translation of proteins - the unidirectional flow is a fundamental law - genetic code is by the use of Codons
3 nucleotides = 1 amino acid → 3 bases make a codon
Codons = words of amino acids
Start: AUG
Stop = UAA, UGA, UAG
U Are Annoying, U Go Away, U Are Gone
Humans have 24 chromosomes(including sex), three billion nucleotides, 21000 genes, and large intergenic regions → Everyone is unique
Mutations: Can be fatal, silent, inside or outside damages
Polymerase errors = point mutations, small repeats, insert/deletion, frame-shifts
Endogenous damages(physical, reactive O2 species) = oxidized DNA, cross-linked bases, double/single-strand breaks
Exogenous damage(radiation, chemicals) = UV, x-rays, chemicals
Transposons = large insertions/deletions, inversions, duplications
Point Mutation: missense, nonsense, silent
Transposons: mobile genetic elements - old and defective
They can cut and paste by transposase enzyme and can paste it somewhere else
if inserted in the intergenic region, it has no effect. if inserted in the coding region, it can become mutagenic
Mutation repair:
Bad bases: mismatch pathway, nucleotide repair
Mismatch: during or after replication - parent strand is methylated, but the daughter is not → can identify parent-daughter
Nucleotide Excision repair: can happen at any time in the cell cycle - removes the bad base and replaces it with a good one(ideally before replication)
Broken chromosome: homology-directed repair, non-homologous joining
Homology-directed: must happen after replication when a sister chromatid is present and must use an identical sister chromatid as a template to fix the broken chromosome
Non-homologous end-joining: happens anytime in the cell cycle and ligate ends are broken together - can be mutagenic since this causes loss of some bases or translocations
DNA rearrangement by transposons: can’t repair
DNA Replication:
4 General rules: semiconservative, 5’-3’, requires RNA primer, and needs a template
5 Main enzymes of replication:
Helicase - unwinds DNA
Topoisomerase - cuts DNA and relaxes the supercoiling
Primase - synthesizes RNA primer
DNA polymerase - replicates the DNA and proofreads
Ligase - Links Okazaki fragments
Replication starts at the ORI - and starts to go from the 5’ end to 3’, both sides in opposite directions
In Eukaryotes, many replication bubbles formed(many ORI)
Ends of the chromosomes become shortened after replication - shorter telomeres
Prokaryotic DNA Polymerases:
Pol. 2: 5’-3’ AND 3’-5’ exonuclease
back-up for DNA Pol.3 and repairs DNA
Pol. 1 and 3: are more error-prone 5’-3’ and repair DNA
Telomerase: elongate the telomeres on the parent strand of the DNA - cells that express telomerase are known as immortal cells ex. spermatogonia, stem cells, cancer cells
Has RNA primers and reverse transcriptase enzyme
DNA vs. RNA:
DNA is double-stranded, has thymine, deoxyribose sugar, double helix, one type
RNA is single-stranded, has uracil, ribose sugar, many 3D shapes, many types
Types of RNA - rRNA, tRNA, mRNA, hnRNA, miRNA, siRNA
Replication vs. Transcription:
Replication has a start site. is in the 5’-3’ direction, has a DNA template
Transcription has a stop site, no primer, and no editing - the start for translation
Regulated by a promoter - higher affinity for RNA polymerase to get a lot of RNA, has DNA binding proteins, repressors and enhancers
Transcription in Prox. vs Euk:
Prokaryotes: transcription and translation at the same time, no mRNA processing, polycistronic, 1 RNA polymerase
Eukaryotes: transcription and translation separate, has mRNA processing(poly-A tail, 5’ G cap, splicing), monocistronic(one RNA, one protein), 3 RNA polymerases
tRNA and Wobble Pairing:
tRNA: transfer RNA - responsible for translocation → has an anticoding region to pair with RNA to code amino acids using codons - needs two ATP to load amino acids
The first two bind by Watson-Crick pairing - the third is more flexible, and adenine can be converted into I for more flexibility
Wobble base pairing: makes it such that the first two must be the same, but the third, Wobble area, can be flexible - allows for non-traditional pairing
Ribosomes for Translation: have a large subunit and a small subunit
Euk: 60s and 40s → 80s total
Prok": 50s and 30s → 70s total
In translation, RNA enters the A site and the new-forming amino acid is added to the P site - it stops when a release factor binds and breaks the bond between the final tRNA and the final amino acid
Energy Count: translation uses a total of 200 ATP, and is most used in tRNA loading
the # of amino acids x 4 = # ATP needed
Post-translational Modification:
Protein folding - by chaperonins
Covalent modification - disulphide bridges, phosphorylation, etc
Processing - cleavage to form active protein
MCAT Biochemistry
Class 1 - 01/06/2024:
Macromolecules - are polymers made from monomers - enzymes that make polymers are polymerases via reactions called polymerization.
Proteins: made up of amino acids(20 kinds)
Have an N-C-C backbone, amine group, carboxylic group, and variable group
Bond together by a peptide bond(formed by dehydration synthesis - loss of H2O)
4 types of structures
Primary = amino acids joined
Secondary = alpha-helix or beta-sheets
Tertiary: formation of a polypeptide and side chain interactions - inner core becomes hydrophobic and outer becomes hydrophilic
Non-covalent interactions: non-polar/non-polar, polar neutral/polar neutral, acid/base(charged)
Covalent: disulphide bridges(harder to break)
Quaternary structure: side chain interactions between different polypeptides - subunits come to form larger units
Carbohydrates: from monosaccharides to disaccharides to polysaccharides;
monosaccharides - CnH2On - 3 common are glucose, fructose, galactose - ribose and deoxyribose
Disaccharides - 2 monosaccharides - 3 common = maltose, sucrose, lactose - C12H22O11 formula
Polysaccharides: many monosaccharides - 3 common are glycogen, starch, and cellulose - function as an energy source
Lipids: the fats - made of a hydrocarbon structure(many C and H) - saturated fatty acids are solid at room temperature
Unsaturated are liquid at room temperature due to the double bond
Triglyceride: 3 fatty acids combined
Phospholipids: 2 lipid structures and one phosphate - form the lipid bilayer due to having polar and non-polar sides
Terpenes: built from isoprene structures and need at least 2 of them - terpenes form waxes and lipid rings like vitamin A
Cholesterol and steroid hormones - 3 six-carbon rings and 1 five-carbon ring)
Thermodynamics: delta G = delta H - TdeltaS
G = Gibbs free energy
H = enthalpy (potential E)
T = temperature
S = Entropy (kinetic E)
When G<0 = negative G, spontaneous - gives E - exergonic
When G>0 = positive, non-spontaneous - needs E - endergonic
When G = 0, equilibrium
Reaction Coupling - using ATP as a source of energy - a very favourable reaction is used to drive an unfavourable one
ATP = ADP + Pi → very exergonic
Exergonic = giving off E
Endergonic = using up E
Chemical Kinetics: the study of reaction rates - all reaction rates proceed through a transition state which tends to be unstable
Activation E = is the required E to produce the TS
if Ea is High = slow rate
if Ea is low = faster rate
Reaction Coordinate Graph - shows the energy vs reaction coordinates over time - the smaller the Ea, the better
We can make the Ea smaller using catalysts - speeding the reaction up by stabilizing TS and reducing Ea
Enzymes: a physiological catalyst - works to speed up a reaction by increasing the rate of reaction, not used up in a reaction, and must be specific
Structure: an enzyme has an allosteric site and an active site - the active site is where the substrate binds(where the reaction occurs) and the allosteric site is another place for enzyme regulation(inhibition or activation)
Two models - active site and induced fit; active is lock and key while the induced fit is when the enzyme needs to change shape to fit a substrate
Can perform both positive and negative feedback
Function: to speed up a reaction
Regulation: by many inhibitions ways, allosteric site, feedback inhibition
V vs. [S] Graph: reaction rate in Velocity vs the substrate concentration [S] → vmax is when the enzyme is saturated and depends on enzyme [C], and the [S] becomes constant - Vmax/2 is when the linear part of the graph is equal to [S]
Km is the substrate [S] required to reach ½ Vmax
Enzyme Inhibition:
Competitive: compete for enzyme binding - same Vmax but the effect on Km is more since you need more substrate - a longer time to reach the same Km - binds at the active site - before substrate binds
Non-competitive: it affects the Vmax since we need more enzymes to deal with the substrate, but Km is unchanged since the active site is the same but prevents the activity of the enzyme- binds to the allosteric site before substrate binds
Un-competitive: it affects both the Vmax and Km since it binds to the allosteric site after the substrate is bound, which affects both enzyme performance and the amount of product being produced - binds to an allosteric site after the substrate binds
Mixed-Type Inhibition: binds at the allosteric site either when the enzyme is bound to the substrate or empty active site. Vmax will become lowered, but Km can vary whether enzyme bound or empty Active site
when bound to the substrate, Km decreases(like un-comp)
When empty active site, Km increases(like comp)
Lineweaver Burk Plots:
Class 2 - 08/06/2024:
Oxidation-Reduction Reactions - Redox
Oxidation is when you gain O, lose H and electrons
Reduction is when you lose O, gain H and electron
Cellular Respiration: When you convert sugar and O2 into carbon dioxide and water
a four-step process - glycolysis, PDC, Krebs, and electron transport
Glycolysis
processed in the cytoplasm and doesn’t need O2
all cells from all domains perform glycolysis → Sugar split into two pyruvate molecules and 4 ATP and 2 NADH formed at the end
Step 1: Got a phosphate from ATP and a glucose → Hexokinase → bam, G-6-P and ADP
Step 2: Got a G-6-P and a isomerization → Bam, F-6-P
Step 3: Got a F-6-P and an ATP → bam, F1-6-bp and ADP
Step 4: Split F1 into two to form 2×3CP
Step 5: add 2pi to 2×3CP → Form 2 PEP 2×3CP
Step 6: 2 PEP plus 2ADP → Pyruvate kinase → 2 Pyruvate
Mnemonic Glycolysis Products:
Gross Guys Favour Fat-Buns, Good Boys Prefer Pretty Pink Pyjamas
Glucose, G6P, F6P, F-1,6-Biphosphate; Glu-ald-phosphate, 1,3-Biphosphoglycerate, 3-Phosphoglycerate, 2-Phosphoglycerate, Phosphophenol pyruvate, Pyruvate
Mnemonic Glycolysis Enzymes:
Honestly, Ignoring Pencils And Getting Pens Make Everything Perfect
Hexokinase, Isomerase, PFK, Anolase, G3P dehydrogenase, Phosphogly-kinase, phosphoglycerate-Muctase, Enolase, Pyruvate Kinase
PDC - Pyruvate Dehydrogenase Complex: occurs in the matrix of the mitochondria and needs O2
Pyruvate is changed from being a 3C molecule to a 2 C molecule and CO2 and NADH are produced; this is from taking pyruvate and coenzyme A to make acetyl-coA
Oxidative Decarboxylation does this: release of CO2 and make NADH
Krebs Cycle: Occurs in the matrix of the Mitochondria and needs O2- Takes 2xacetyl-coA with oxaloacetate to make 2xCO2, NADH and FADH2
In order:
2C + Oxaloacetate → citrate(6C) → NADH + CO2 → 5C → NADH and CO2 → 4c → GTP → succinate → fumarate → malate → oxalacetate
Krebs mnemonic Substrates:
Can Interests Kindly Send Some Free Money Oop
Citrate, Isocitrate, α-Ketoglutarate, Succinyl-CoA, Succinate, Fumarate, muctase, Oxcaloacetate
Kreb’s mnemonic Enzymes:
Can Anyone Interested Kindly Send Some Free Money
Citrate synthase, Aconitase, Isocitrate dehydrogenase, α-ketoglutarate dehydrogenase, Succinyl synthase, Succinate dehydrogenase, Fumarase, Malate dehydrogenase
ETC and Oxidative Phosphorylation: occurs in the inner membrane of the mitochondria and needs O2
OP is the oxidation of NADH and FADH2 to make ADP into ATP → This makes energy move e- transport chain and pumps protons out of the mitochondria
ETC is a chain of 5 e- carriers that perform redox roles(cytochromes)
Step 1: NADH dehydrogenase → reduce NADH, pass e
Step 2: coenzyme Q → release FAD from FADH, pass e
Step 3: cytochrome C reductase → passes e to next
Step 4: cytochrome C oxidase → makes H2O and sends to ATP synthase to make ATP
ETC Mnemonic:
Norway’s Queens’ CyteR Can’t Oxidize ATP
NADH Reductase, CoQ, Cytochrome Reductase, CoC, Cytochrome Oxidase, ATP Synthase
Total ATP of respiration = 30 ATP made in Euk, and 32 in Prok
Fermentation: when no oxygen is available - no ETC, No oxidative phosphorylation, no Krebs, no PDC, no glycolysis
Take Pyruvate and form ethanol yeasts) and lactic acid(in muscles, other cells)
Gluconeogenesis: the formation of glucose from 2 pyruvates (reverse of glycolysis but some different unique enzymes) - Happens in the body when glycogen stores are depleted in the liver
2 pyruvate with pyruvate carboxylase forms 2 oxaloacetate
PEP carboxykinase makes 2 PEP
2 PEP is turned into 2×3CP to then F16CP
F16CP is then turned into F6CP to G-6-P
G-6-P with glucose-6-phosphatase to make glucose
Glycogenesis: the formation of glycogen from glucose
made by using G-6-P to G-1-P by phosphoglucomutase and then using UDP to make UDP-glucose which is turned into glycogen by glycogen synthase
Glycogenolysis: breakdown of glycogen to glucose
Here glycogen is phosphorylated into G-1-P and then isomerized to G-6-P to make glucose again
happens in the liver
Insulin increases when glucose is high to make glycogen - stores energy for later
Pentose Phosphate Pathway: takes G-6-P to form NADPH and ribose-5-phosphate. NADPH is important in its role of reducing the power of fatty acid synthesis and helps neutralize reactive O2 species as well as make the building block for nucleotides
Fatty Acid Oxidation: the fat digestion - the removal of 2C units as acetyl-CoA from a fatty acid and makes 1 FADH2 and 1 NADH - the acetyl is then moved to Krebs or ketone bodies
dietary fat chylomicrons move from the lymph system to the liver and organs and then undergo beta-oxidation which then turns the fatty acid into acetyl-coA
Fatty Acid Synthesis: uses high amounts of ATP and NADPH where 2C units are added to the chain until 16C fat is made
using acetyl-coA to make malonyl-CoA
Ketogenesis: during starvation, acetyl-coA turns into ketone bodies and can supply energy to the brain and lower blood pH
Protein Catabolism: break down of protein by proteases to amino acids
Metabolism: when the body is fed, glycolysis, glycogenesis, and fatty acid synthesis are favoured. When the body is starved, glycogenolysis, glucogenesis, and fatty acid oxidation are favoured.
Class 3 - 15/06/24:
Nucleotide: made up of sugar, base, and phosphates
Sugar = deoxyribose or ribose
Base = ACTG
Phosphates = 3 linked together
Nucleic Acid Structure: 5’-3’ linkage, antiparallel and complementary, phosphodiester bonds
Pyrimidines = U, T, C(smaller)
Purines = A, G(bigger)
A-T, C-G, A-U(in RNA)
DNA structure:
in prokaryotes, circular DNA genome, formed by methylation, and supercoiling
in eukaryotes, several linear chromosomes → (biggest)chromosome, to chromatin, to histones bound to make nucleosomes, to make smaller DNA strands(smallest)
Centromere: the middle of the chromosome where the spindle fibres attach to - made of heterochromatin and repetitive DNA sequences - short sequences repeat - both single and double-stranded DNA which can loop to form a knot at the end of the chromosome to stabilize it
Telomere: the ends of a eukaryotic linear chromosome - also has a hand in aging
DNA protection: the tighter it is, the less likely it will be uncoiled
The Central Dogma: DNA leads to transcription to RNA that leads to the translation of proteins - the unidirectional flow is a fundamental law - genetic code is by the use of Codons
3 nucleotides = 1 amino acid → 3 bases make a codon
Codons = words of amino acids
Start: AUG
Stop = UAA, UGA, UAG
U Are Annoying, U Go Away, U Are Gone
Humans have 24 chromosomes(including sex), three billion nucleotides, 21000 genes, and large intergenic regions → Everyone is unique
Mutations: Can be fatal, silent, inside or outside damages
Polymerase errors = point mutations, small repeats, insert/deletion, frame-shifts
Endogenous damages(physical, reactive O2 species) = oxidized DNA, cross-linked bases, double/single-strand breaks
Exogenous damage(radiation, chemicals) = UV, x-rays, chemicals
Transposons = large insertions/deletions, inversions, duplications
Point Mutation: missense, nonsense, silent
Transposons: mobile genetic elements - old and defective
They can cut and paste by transposase enzyme and can paste it somewhere else
if inserted in the intergenic region, it has no effect. if inserted in the coding region, it can become mutagenic
Mutation repair:
Bad bases: mismatch pathway, nucleotide repair
Mismatch: during or after replication - parent strand is methylated, but the daughter is not → can identify parent-daughter
Nucleotide Excision repair: can happen at any time in the cell cycle - removes the bad base and replaces it with a good one(ideally before replication)
Broken chromosome: homology-directed repair, non-homologous joining
Homology-directed: must happen after replication when a sister chromatid is present and must use an identical sister chromatid as a template to fix the broken chromosome
Non-homologous end-joining: happens anytime in the cell cycle and ligate ends are broken together - can be mutagenic since this causes loss of some bases or translocations
DNA rearrangement by transposons: can’t repair
DNA Replication:
4 General rules: semiconservative, 5’-3’, requires RNA primer, and needs a template
5 Main enzymes of replication:
Helicase - unwinds DNA
Topoisomerase - cuts DNA and relaxes the supercoiling
Primase - synthesizes RNA primer
DNA polymerase - replicates the DNA and proofreads
Ligase - Links Okazaki fragments
Replication starts at the ORI - and starts to go from the 5’ end to 3’, both sides in opposite directions
In Eukaryotes, many replication bubbles formed(many ORI)
Ends of the chromosomes become shortened after replication - shorter telomeres
Prokaryotic DNA Polymerases:
Pol. 2: 5’-3’ AND 3’-5’ exonuclease
back-up for DNA Pol.3 and repairs DNA
Pol. 1 and 3: are more error-prone 5’-3’ and repair DNA
Telomerase: elongate the telomeres on the parent strand of the DNA - cells that express telomerase are known as immortal cells ex. spermatogonia, stem cells, cancer cells
Has RNA primers and reverse transcriptase enzyme
DNA vs. RNA:
DNA is double-stranded, has thymine, deoxyribose sugar, double helix, one type
RNA is single-stranded, has uracil, ribose sugar, many 3D shapes, many types
Types of RNA - rRNA, tRNA, mRNA, hnRNA, miRNA, siRNA
Replication vs. Transcription:
Replication has a start site. is in the 5’-3’ direction, has a DNA template
Transcription has a stop site, no primer, and no editing - the start for translation
Regulated by a promoter - higher affinity for RNA polymerase to get a lot of RNA, has DNA binding proteins, repressors and enhancers
Transcription in Prox. vs Euk:
Prokaryotes: transcription and translation at the same time, no mRNA processing, polycistronic, 1 RNA polymerase
Eukaryotes: transcription and translation separate, has mRNA processing(poly-A tail, 5’ G cap, splicing), monocistronic(one RNA, one protein), 3 RNA polymerases
tRNA and Wobble Pairing:
tRNA: transfer RNA - responsible for translocation → has an anticoding region to pair with RNA to code amino acids using codons - needs two ATP to load amino acids
The first two bind by Watson-Crick pairing - the third is more flexible, and adenine can be converted into I for more flexibility
Wobble base pairing: makes it such that the first two must be the same, but the third, Wobble area, can be flexible - allows for non-traditional pairing
Ribosomes for Translation: have a large subunit and a small subunit
Euk: 60s and 40s → 80s total
Prok": 50s and 30s → 70s total
In translation, RNA enters the A site and the new-forming amino acid is added to the P site - it stops when a release factor binds and breaks the bond between the final tRNA and the final amino acid
Energy Count: translation uses a total of 200 ATP, and is most used in tRNA loading
the # of amino acids x 4 = # ATP needed
Post-translational Modification:
Protein folding - by chaperonins
Covalent modification - disulphide bridges, phosphorylation, etc
Processing - cleavage to form active protein