Plant Physiology 3
Plant Respiration
Ch. 12
In photosynthesis, CO2 is reduced to carbohydrate and the goal is to store the energy. In respiration, reduced carbon is oxidized to CO2 and water and the goal is to release the energy stored in carbon compounds for cellular use. It will release lots of energy, energy in the form of ATP. Electrons drive ATP synthesis.
C12H22O11 + 12O2 → 12CO2 + 11H2O
Gibbs free energy is about -5760 kJ per mole; much energy is released.
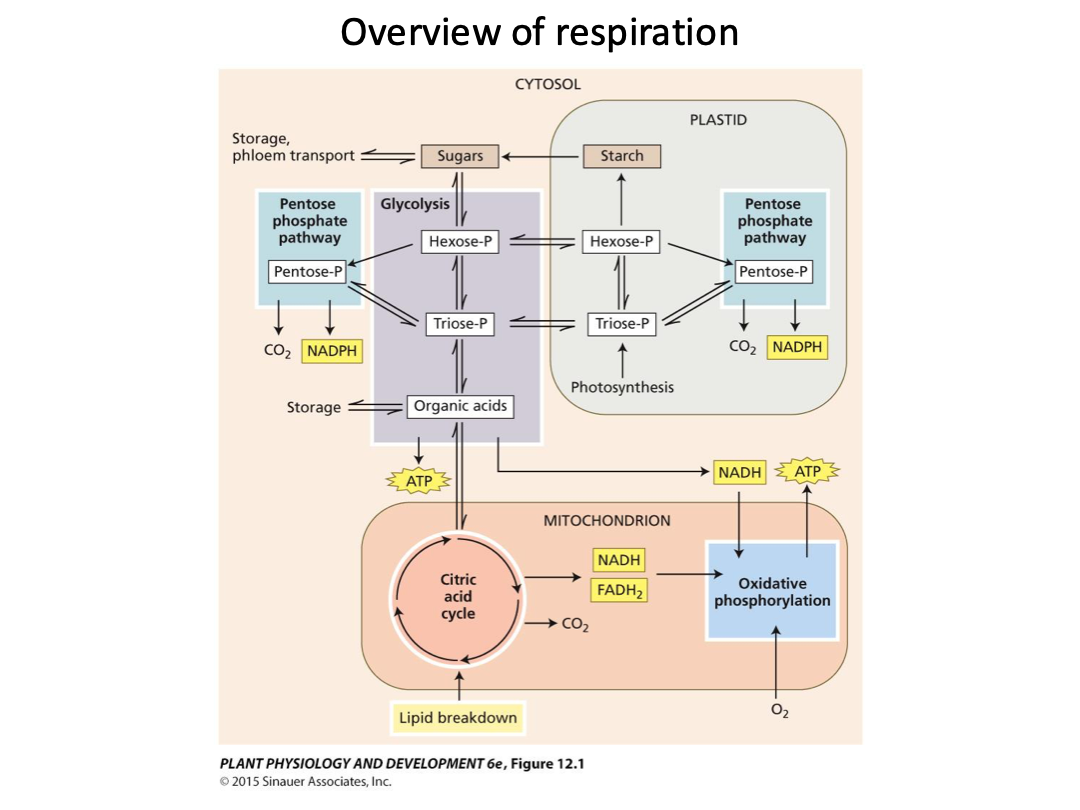
Enters into the mitochondria, does citric acid cycle and oxidative phosphorylation. ETC mainly occurs here.
Glycolysis occurs. NADH and NADPH are similar and they play a role, but they’re in different regions of the cell. Different organelles.
Respiration: glycolysis, oxidative pentose phosphate pathway, citric acid cycle, and oxidative phosphorylation (electron transport chain and ATP synthesis).
Substrates for respiration are generated by other cellular processes and enter the respiratory pathways.
Glycolysis and the oxidative pentose phosphate pathways in the cytosol and plastids convert sugars into organic acids such as pyruvate, generating NADH and ATP.
Organic acids are oxidized in the mitochondrial citric acid cycle, and the NADH and FADH2 produced provide the energy for ATP synthesis by ETC and ATP synthase in oxidative phosphorylation.
Glycolysis
“sugar” + “splitting”
Carbohydrates are converted into hexose phosphates, each of which is then split into two triose phosphates. In a subsequent energy-conserving phase, each triose phosphate is oxidized and rearranged to yield one molecule of pyruvate, an organic acid.
Besides preparing the substrate for oxidation in the citric acid cycle, glycolysis yields some ATP and NADH.
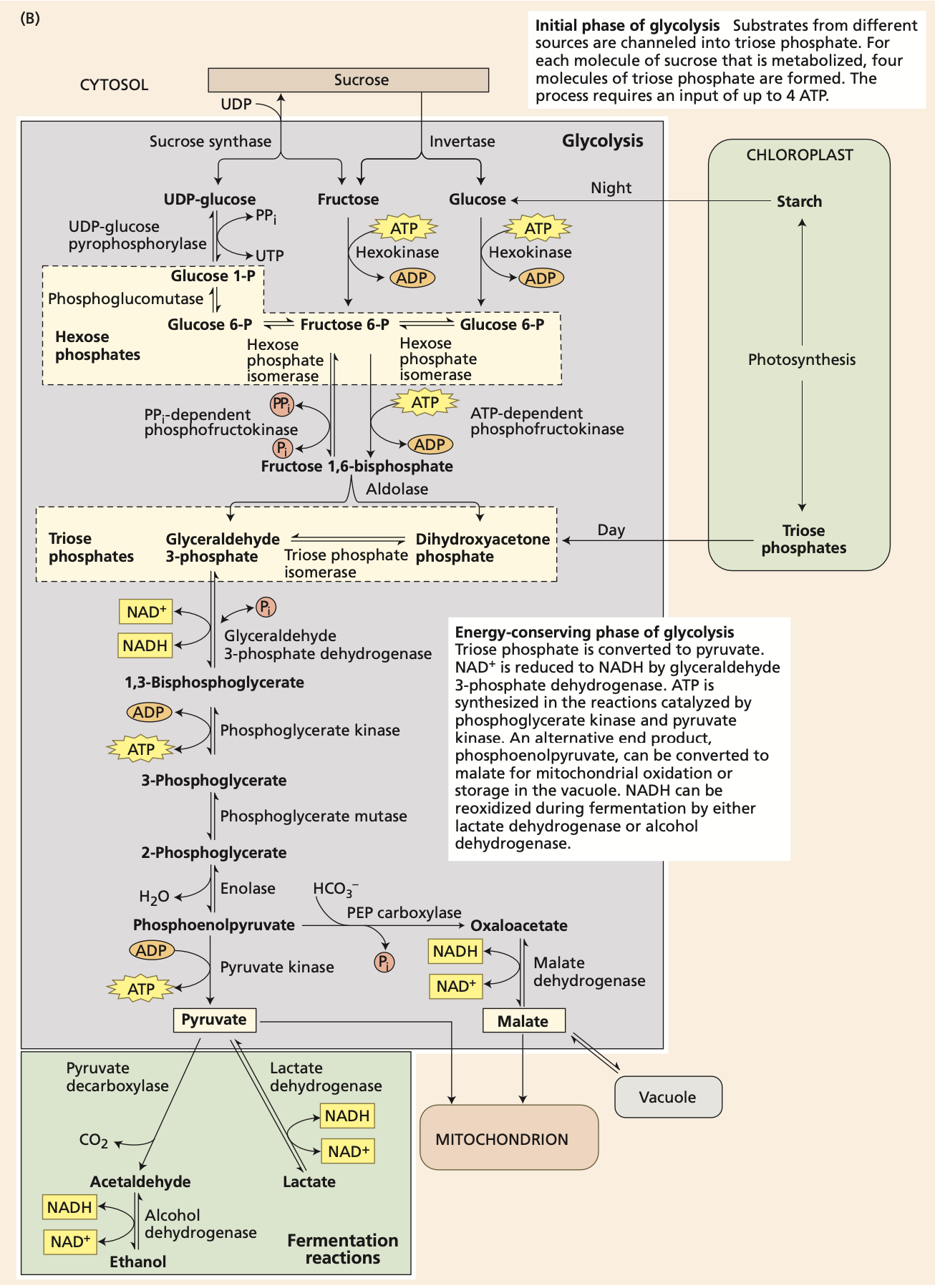
The part at the bottom in green is fermentation. In the absence of oxygen, fermentation regenerates the NAD+ needed for glycolysis. With ethanol or lactate as the final product, the efficiency of fermentation is about 4%.
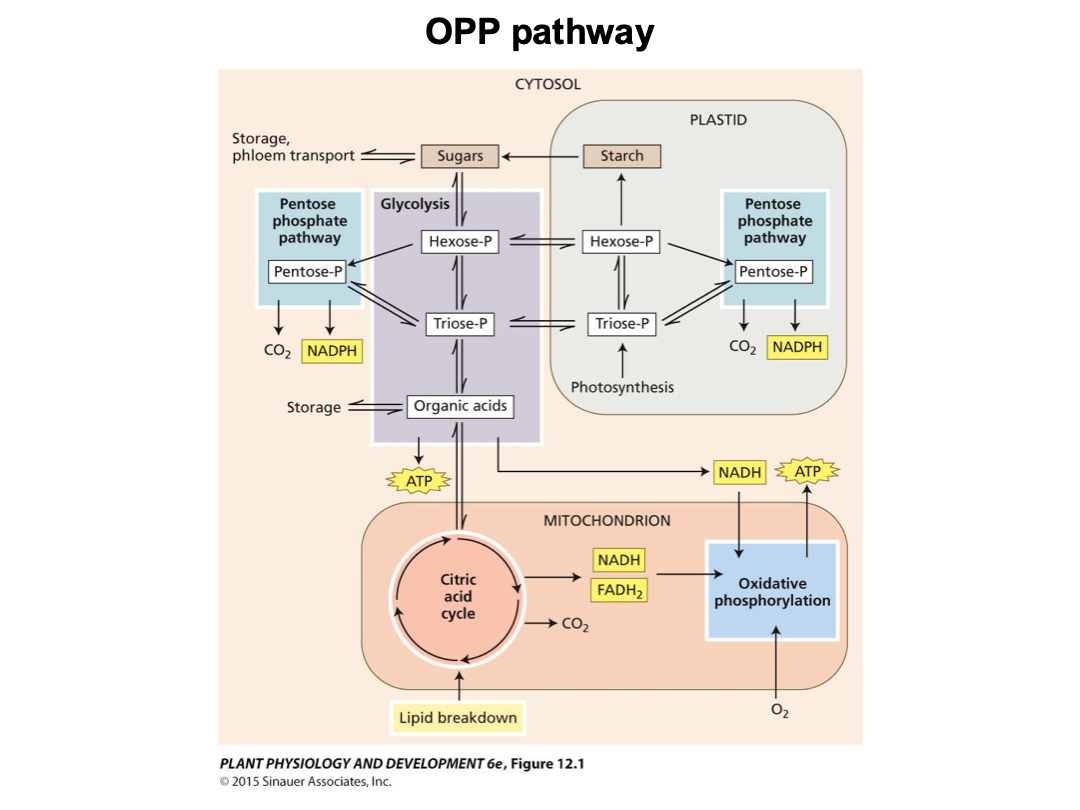
The Citric Acid Cycle
Pyruvate is oxidized to CO2 within the mitochondrial matrix through the citric acid cycle, generating a large number of reducing equivalents in the form of NADH and FADH2 and a small amount of ATP.
One molecule of pyruvate gives rise to three molecules of CO2.
We should know what comes out.
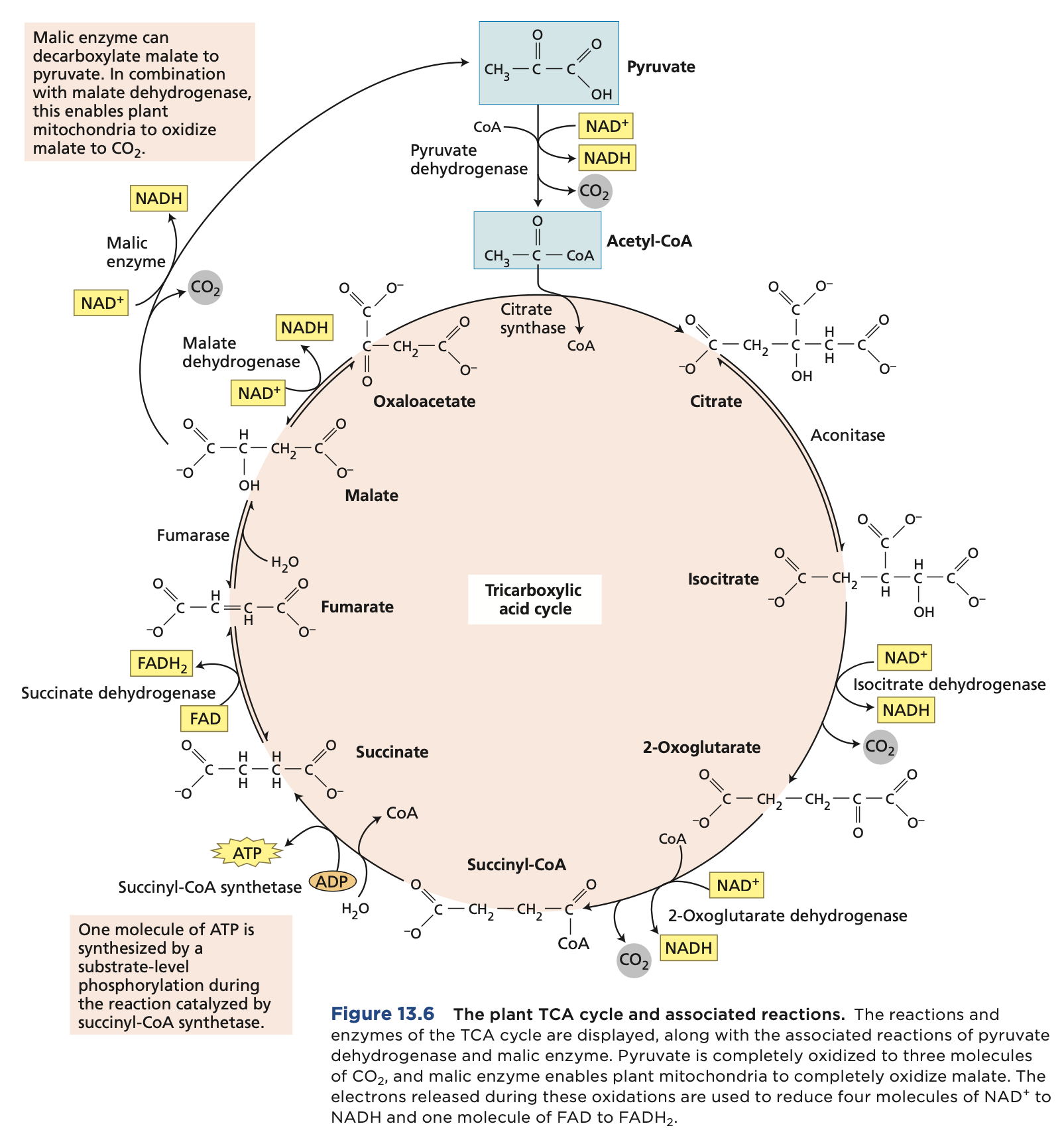
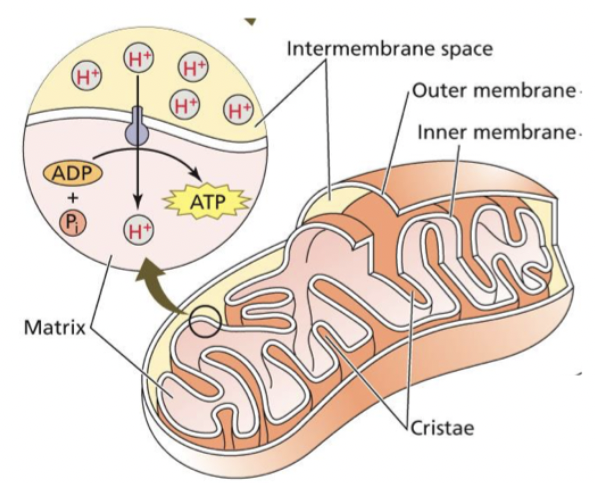
This occurs in the mitochondrial matrix of the plant mitochondria. While the ETC occurs in the inner membrane of the plant mitochondria.
Oxidative Phosphorylation
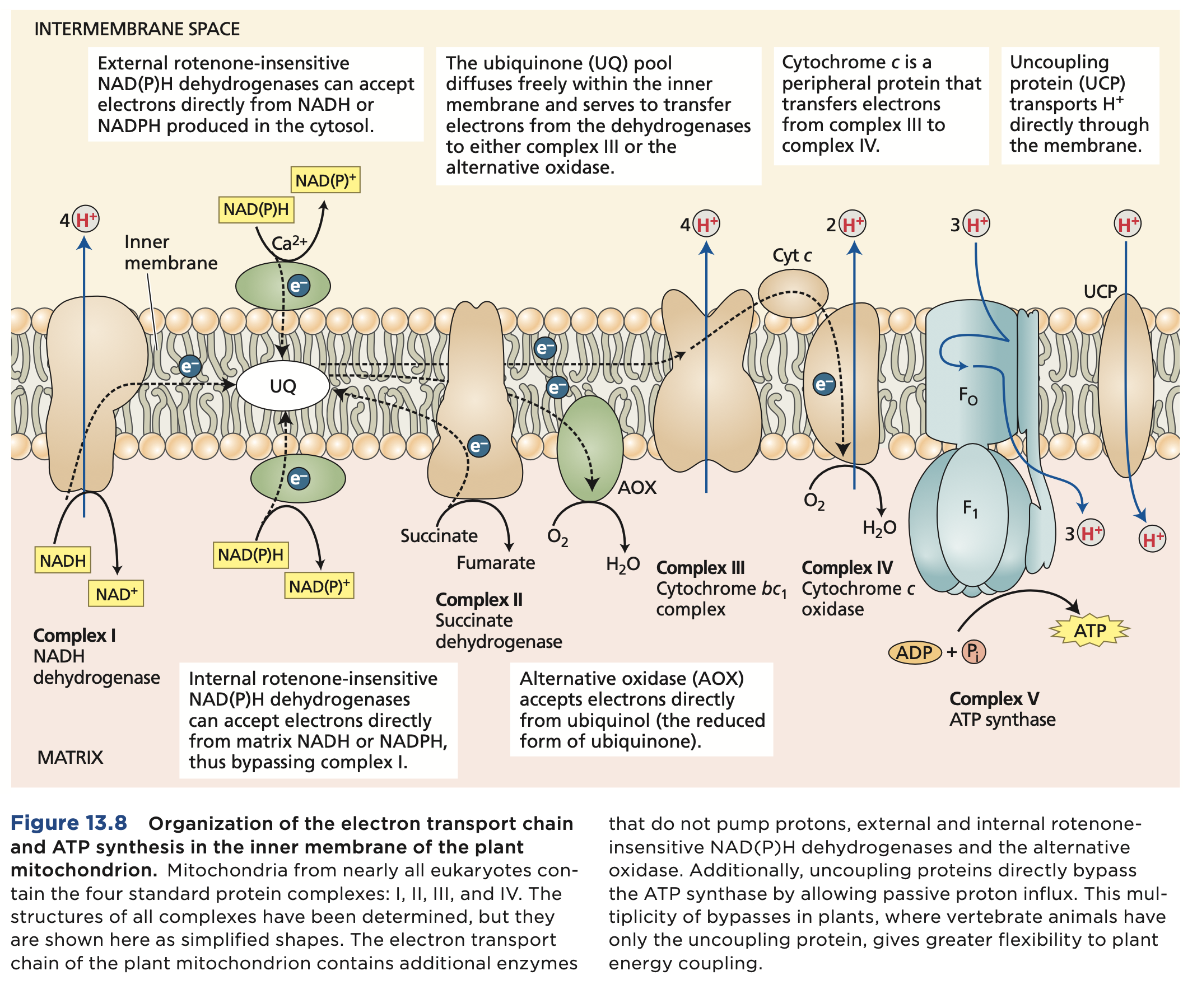
Electron transport from NADH and FADH2 to oxygen is coupled by enzyme complexes to proton transport across the inner mitochondrial membrane. This generates an electrochemical proton gradient used for powering synthesis and export of ATP. The ETC catalyzes a lot number of electrons from NADH to O2.
The individual electron transport proteins are organized into four transmembrane multiprotein complexes (identified by roman numbers I to IV), all of which are localized in the inner mitochondrial membrane. 3 engage in proton pumping, besides complex 2. ATP synthesis in mitochondria.
These are all precursors to biosynthetic pathways.
Plant Growth and Cell Walls
Cell walls have a strong influence on both the direction of cell expansion and the rate of cell expansion. They are often defining features in distinguishing the structure and function of different plant cell types. These are all critical aspects of plant growth and development.
Sclerenchyma → phloem → cortex → xylem
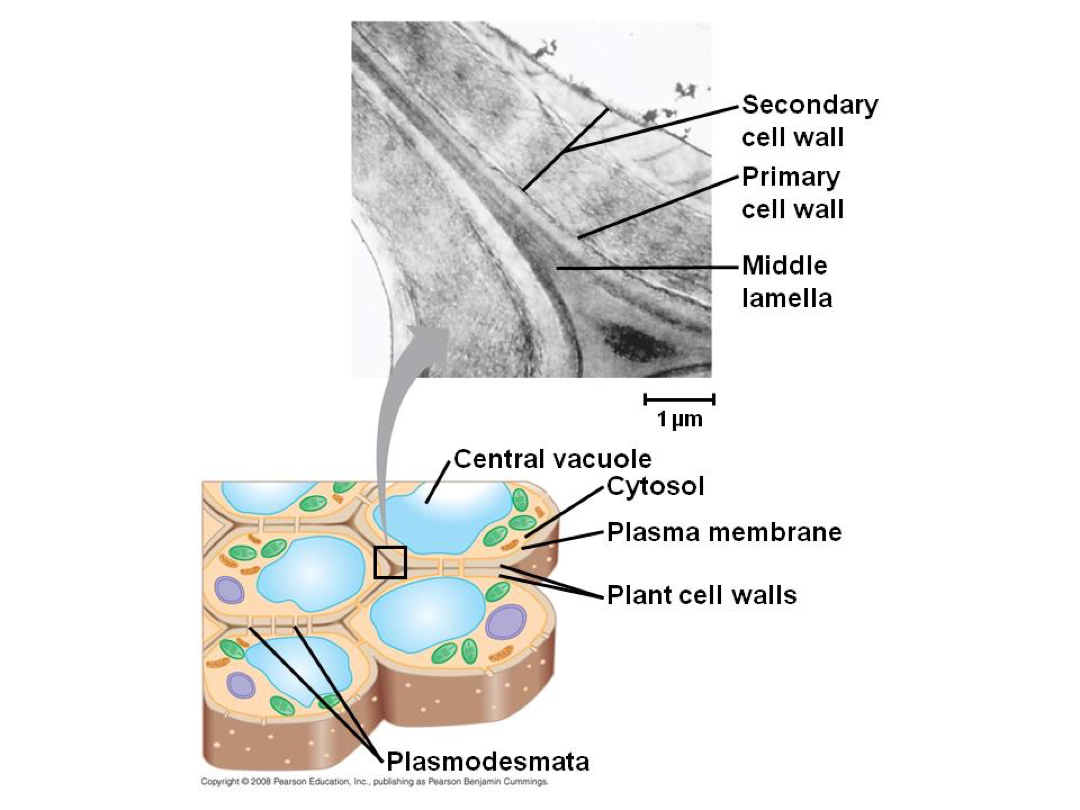
Middle lamella - during cell division (formed). Rich in pectin.
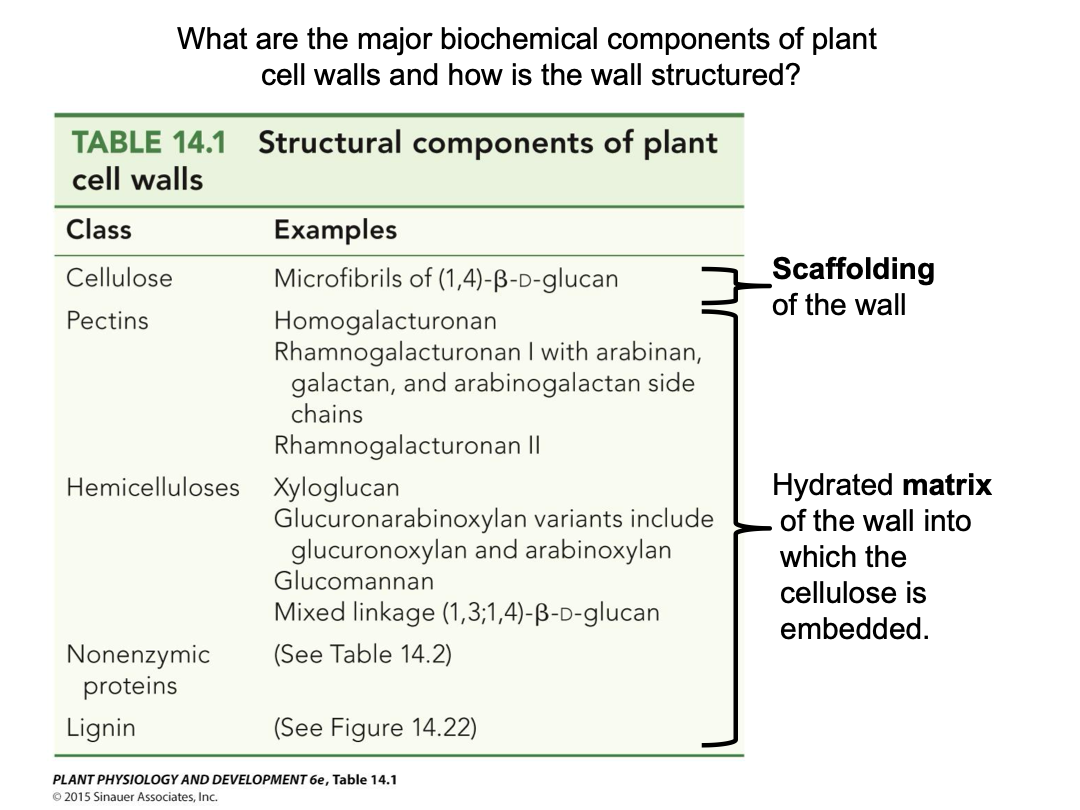
Primary cell wall - after cell division and during cell growth it is created on either side of the middle lamella. A more flexible cell wall. 25% cellulose, 35% hemicellulose, 35% pectin, 2-5% protein. Ratios vary greatly, as do specifics.
Secondary cell wall - after cell growth, it is deposited inside primary cell wall. More rigid due to components. Rigidity is why it comes in after cell growth - too rigid for cell to grow. Typically more cellulose and less pectin. Can contain lignin. Great diversity in specific chemical composition of plant cell walls. Chemical diversity leads to physical diversity.
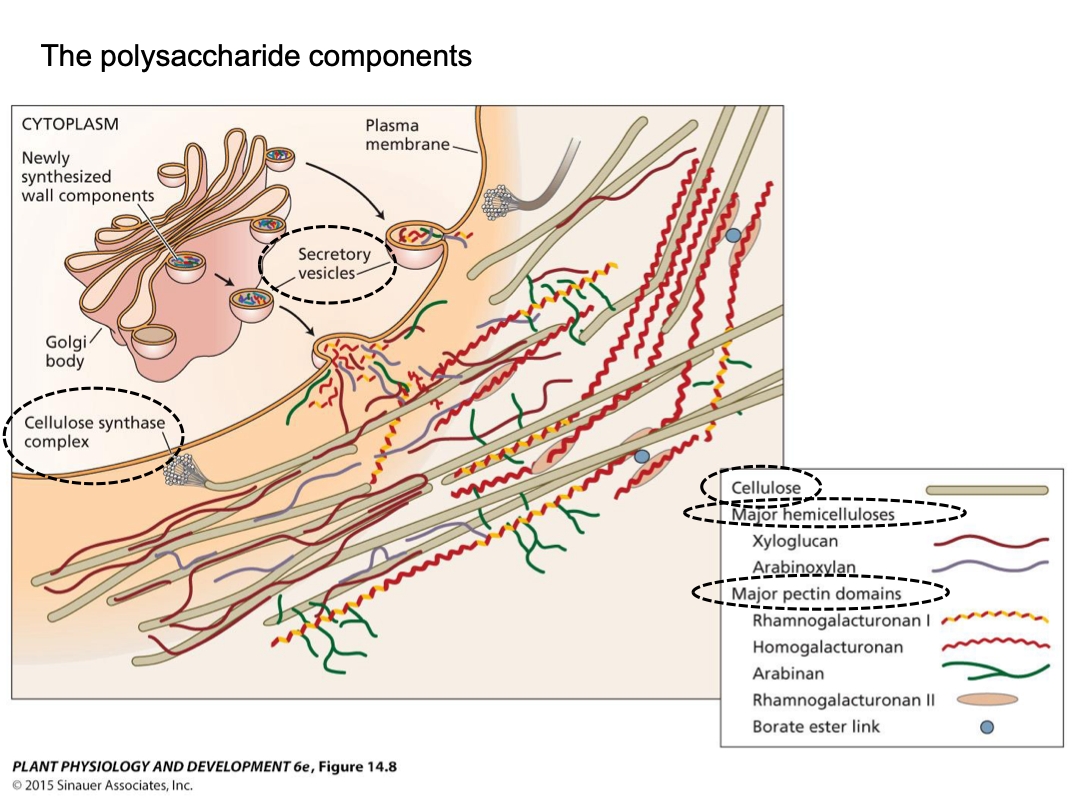
Cellulose microfibrils: very strong physical strength. Glucan chain varies in length, and is not strong. High tensile strength (steel), chemical stability, and resistant to enzymatic attacks. 18-36 glucan chains within, extensive hydrogen bonding within and between glucan chains. Linear chains of 1-4 linked beta-D-glucose individual glucose per chain.
Starch has an alpha(1-4) glycosidic linkage and cellulose has a beta(1-4) glycosidic linkage.
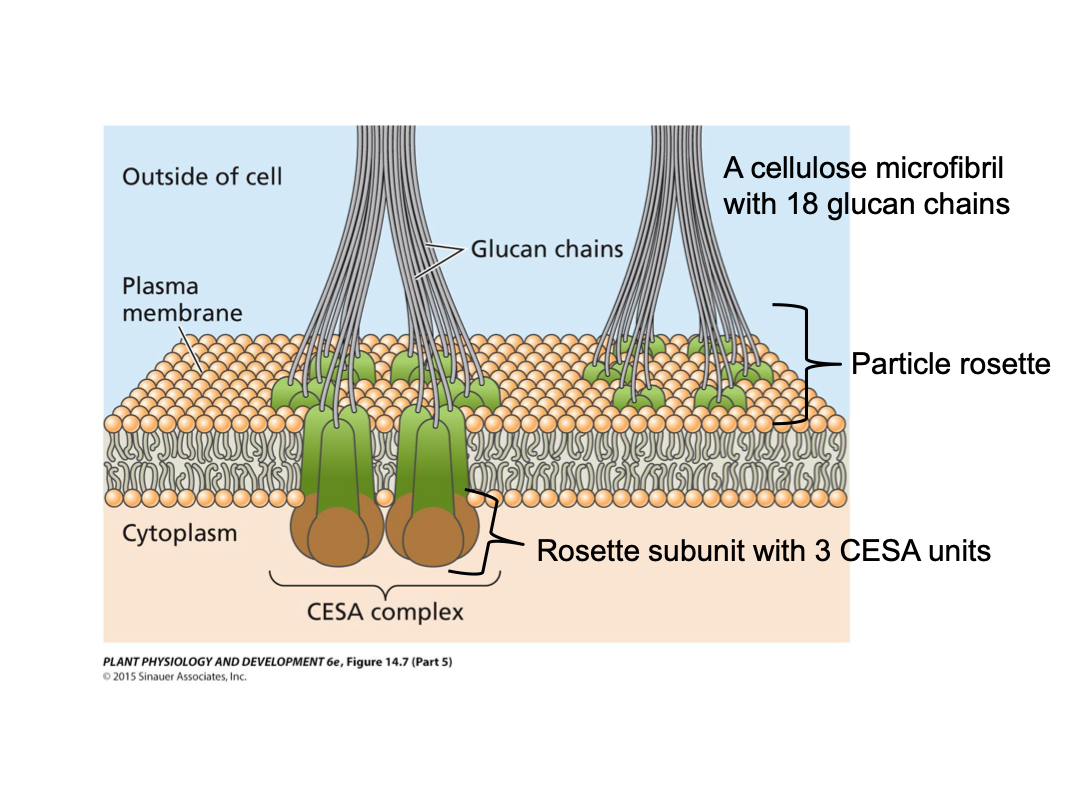
Cellulose microfibrils are synthesized by large protein complexes embedded in the plasma membrane, termed particle rosettes (36 glucan chains, bundled together). CESA (cellulose synthase) synthesizes the individual glucan chains. CESA units together make up a rosette subunit (3 to 6 units). Six rosette subunits together make up the particle rosette. If a rosette has 3 subunits it has 18 glucan chains total. It is embedded in the plasma membrane. As soon as CESA synthesizes, it leaves the cell.
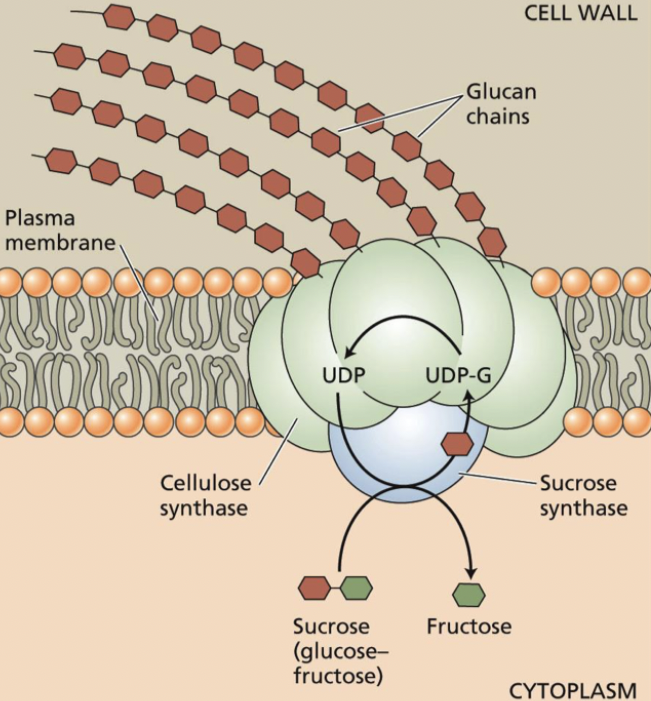
CESA may be complexed with sucrose synthase → cleaves intracellular sucrose to UDP-glucose and fructose → UDP-glucose is then used for synthesis of the glucan chain, which is being released into the cell wall space.
When is it bundled together, it catalyzes the reverse reaction (produces sucrose). Substrate to be synthesized. Easily incorporated into a chain.
There is a close association between the cytoskeleton and microfibril deposition in the wall. Plasma membrane has texture (cytoskeleton, microtubules). Microfibrils have a direction to that of microtubules. Embedded in the plasma membrane.
Hemicellulose and pectins are complex polysaccharides that are the flexible hydrated matrix into which cellulose microfibrils are embedded. Wide variety of monosaccharide building blocks. Polysaccharides synthesized in Golgi apparatus. Delivered to cell wall via exocytosis (vesicle).
Hemicellulose: tightly bound polysaccharides of the wall, long chains often with short side branches, bind to surface of microfibrils, form tethers between microfibrils. Commonly xyloglucan.
Xyloglucan is a typical and common hemicellulose. Xylose sticks out of glucan chain (characterization). Xylose and glucose are two basic subunits, very simple structure.
Pectins: highly hydrated gel phase of the wall (more easily extracted), complex polysaccharides, typically containing galacturonic acid. Methyl esters can be removed, exposing carboxyl groups. Can get ionic bridging of the carboxyl groups of adjacent chains by Ca2+. This can generate large pectin networks.
Galacturonic acid: for each subunit, 6th carbon position has a carboxyl group. Almost all the different cell types include different methylated and non-methylated pectin. Every cell type is a mix of the two.
Methylated pectin: hydrogen of COOH is actually a methyl group → COOCH3. Methyl group can be removed by the enzyme by pectin methyl esterase (PME).
If you have 2 different cell types, one with active extension, and one more mature - mature one has more demethylated pectin because it is more rigid. More crosslinking, more rigidity.
Cell wall proteins often have unusual and unique properties. Rich in particular amino acids or highly glycosylated or highly repetitive.
Cell wall protein roles: structural, enzymes for cell wall assembly and modification, cell adhesion, cell signaling roles. Proteins contain things attached to it.
Cell wall biosynthesis: polymer synthesis → polymer deposition → assembly of components → modification of components (to alter wall properties)
Assembly of the cell wall is self assembly (wall polymers aggregate) and enzyme-mediated assembly.
Enzymes:
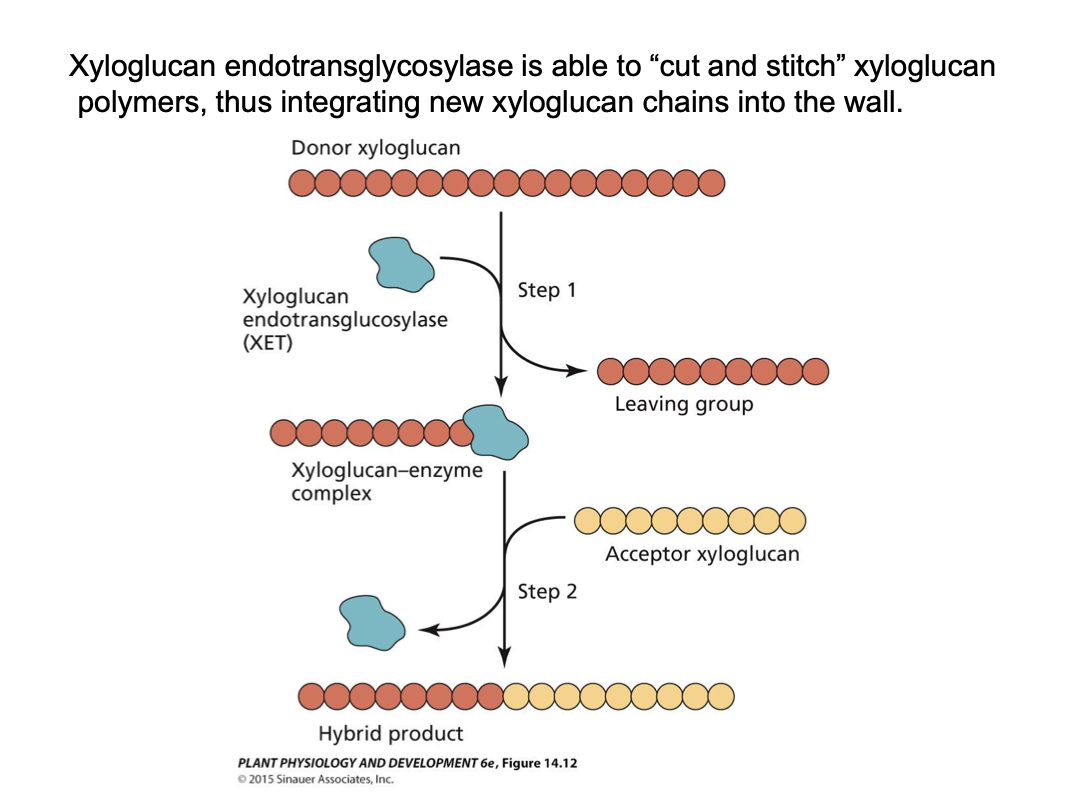
Xyloglucan endotransglycosidase - cut and stitch xyloglucan polymers
Pectin esterases: remove methyl groups in pectin to allow more extensive ionic bridging - more rigid gel.
Peroxidase - cross-link phenolic groups in lignin
Cell wall expansion
Stretching (modification of chemical bonds). Need to break or maintain interactions of bonds, moving them apart.
New cell wall components. Avoid stretching thinner and thinner.
Secondary cell wall contains lignin. Lignin base is an aromatic ring with a hydroxyl group (phenol) that is derived from phenylalanine. These are secreted into the wall, where oxidation by cell wall peroxidases cross-links the phenol groups into a large complex macromolecule. This complex binds tightly to cellulose and displaces water. These factors greatly strengthen the wall, but also prevent any further expansion of the cell wall.
The aromatic ring peroxidase is trying to transfer electrons from the phenol group and then transfering in the electrons. Electron into hydrogen peroxide. Peroxidase moves away from phenol group. Becomes radical. Phenol radical. Lignin is the networking structure.
Cell expansion (growth): plant cells typically expand 10-100 fold in volume (sometimes 10k-fold). Irreversible stretching (yielding) of cell wall. Volume increases don’t thin because of integration of new cell wall polymers into the existing cell wall.
Direction (pattern) of cell expansion
Rate of cell expansion
Patterns:
Tip growth - localized cell expansion due to localized (polar) new wall synthesis. Root hairs and pollen tubes expand by tip growth.
Diffuse growth - expansion distributed over cell surface and due to both wall yielding and new wall synthesis. Expansion can still be directional. Most plant cells expand by diffuse growth.
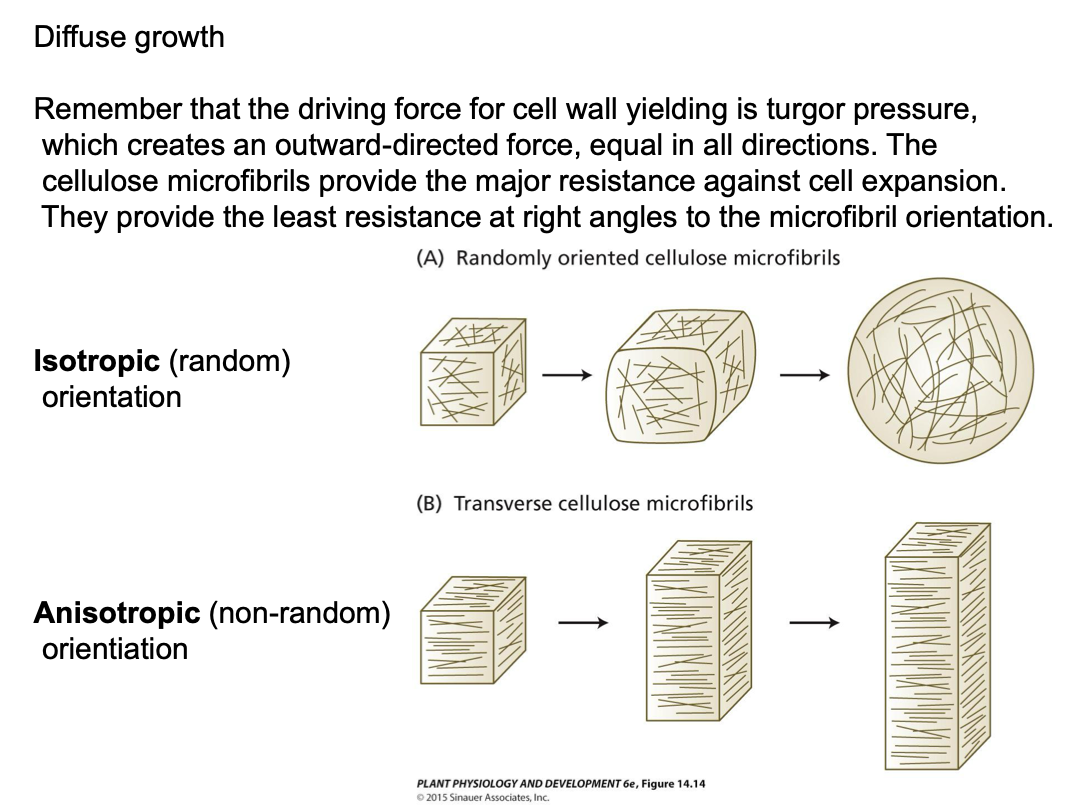
Understand how direction is confirmed. Direction for the extension.
The orientation of microtubules in the cytosol mirrors that of newly synthesized cellulose microfibrils in the cell wall.
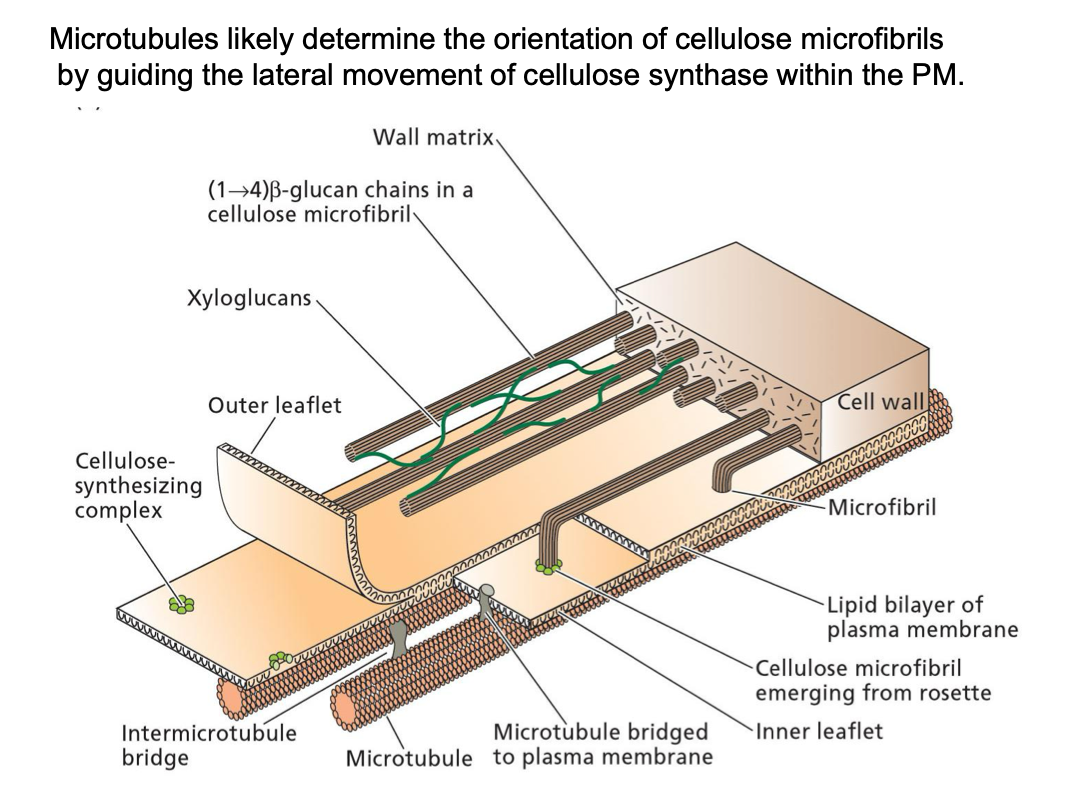
The orientation of microtubule deposition is disrupted by drugs that depolymerize microtubules (oryzalin, colchicine). An immunofluorescent technique shows that microtubules are clearly visible in root cortex and oriented at right angles to the direction of cell expansion. Microtubules are not visible after oryzalin treatment. Oryzalin treatment disrupts normal cell elongation - abnormal root morphogenesis (bulbous, not elongated)
Each new cell wall layer is stretched and thinned during expansion. Synthesizing new cell wall layers is important because the microfibrils in older layers become passively reoriented parallel to the direction of cell expansion. The newest layers have the greatest influence on the direction of cell expansion, since the older layers have become thin, providing less resistance to expansion.
Plants grown under continuous water stress require more turgor pressure.
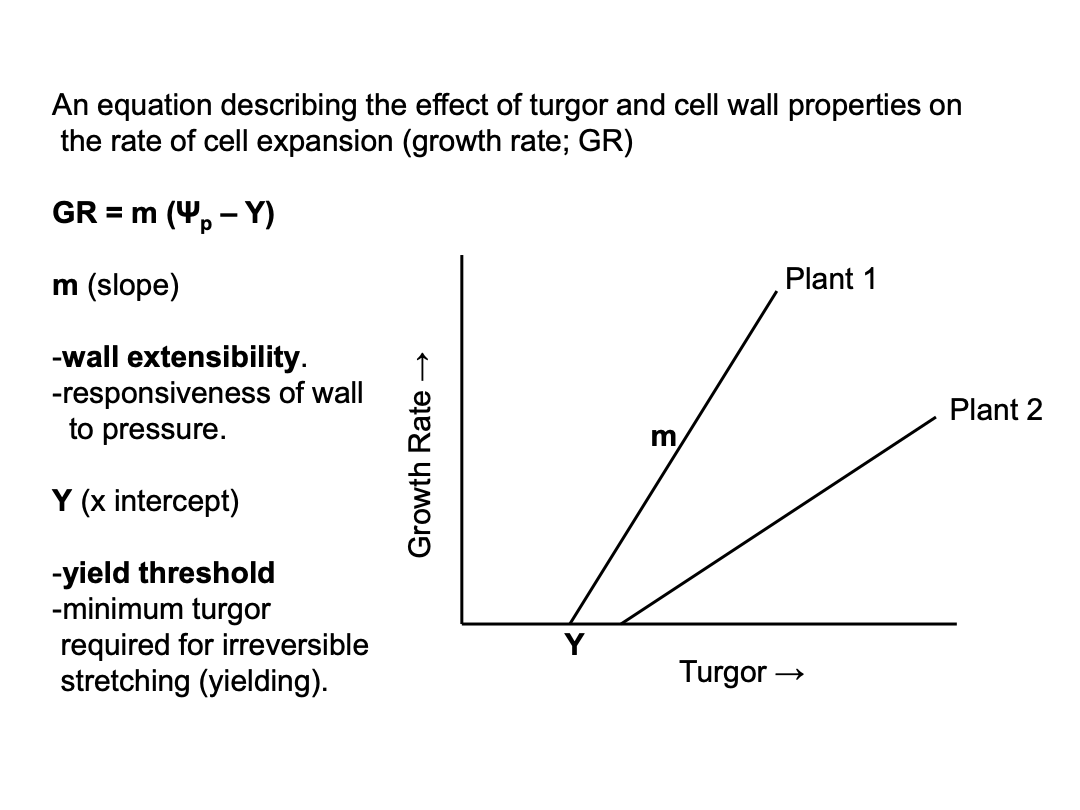
m and Y depend on the structure and biochemical properties of the wall and either or both can change in response to short-term signals (eg. the hormone auxin) or long-term conditions (eg. water deficit). Plants in long-term water deficit will grow slower than well-watered plants, even at the same turgor pressure, based on differences in cell wall structures. about 0.3 turgor pressure to drive water stress (Plant 2 is at 0.3, plant 1 is at 0.25).
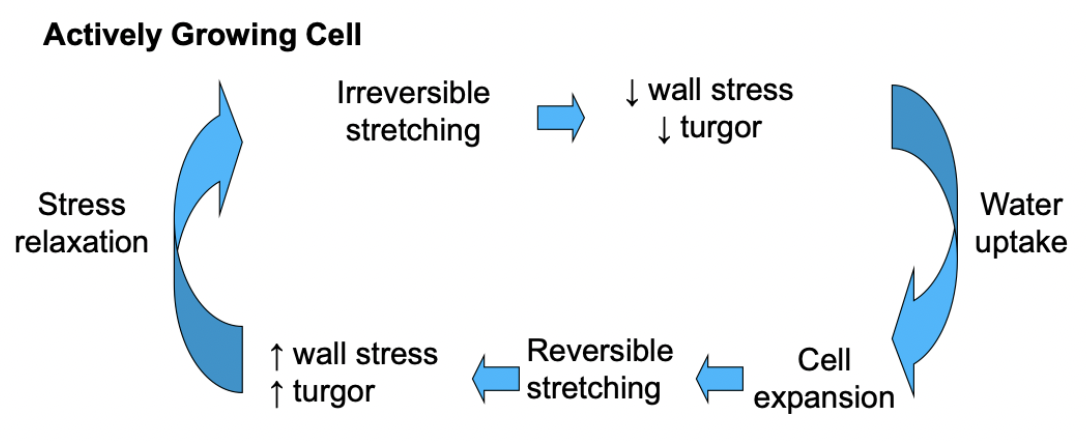
Cell walls of actively growing plant cells also exhibit shorter-term biochemical changes in their cell walls that allow for irreversible stretching (yielding) of the wall in response to turgor. Stress relaxation.
Water uptake triggers cell extension. Increased water stress = increased pressure, a relax in pressure will lead to irreversible stretching and then more water stress. Cell wall stress decreases volume. Keep taking up water, driving cell expansion.
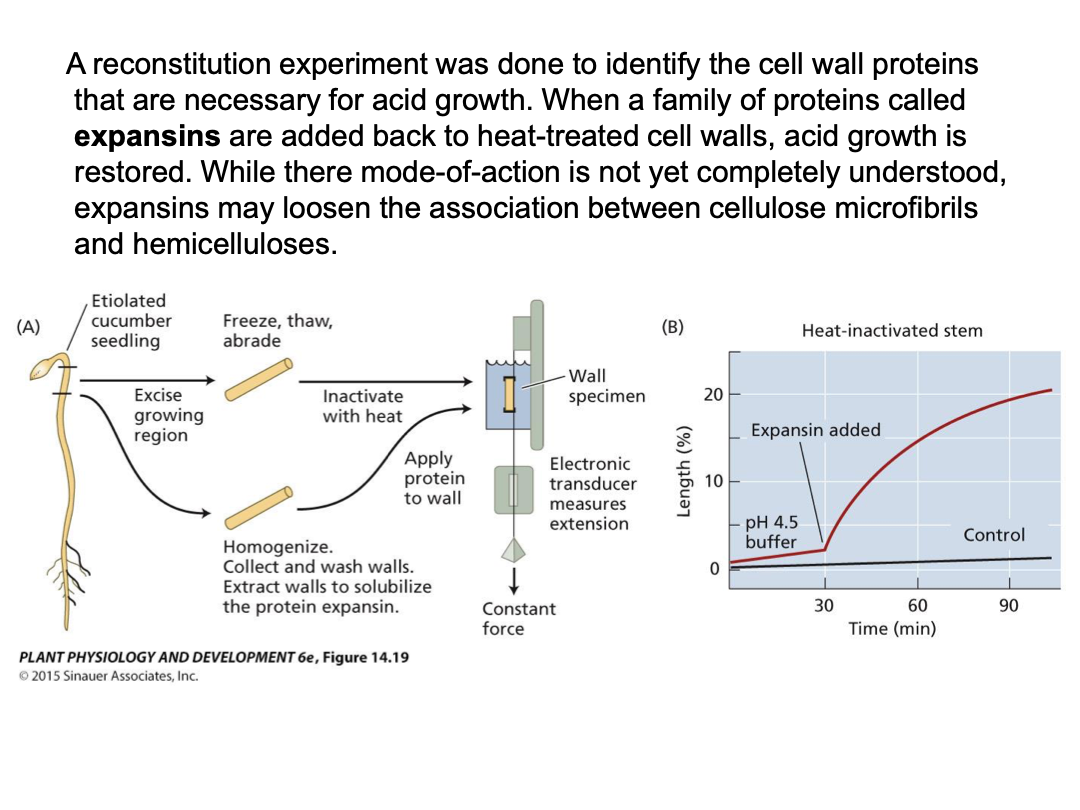
Studying stress relaxation: stem section → isolate cell walls → use extensometer for tension (turgor pressure) → measure wall extension (creep) under different conditions
wall creep enhanced at lower pH
Acid growth response is prevalent in young expanding cells, but not mature ones
Pre-treated with heat or protease denaturation - acid growth response not active
At some stage, the cell walls of older cells exhibit changes in their structure and no longer exhibit stress relaxation. Their walls no longer yield to turgor pressure and they stop growing. Mature cells don’t have stress relaxation. No further water uptake, it is at equilibrium
Changes in mature cell walls that lead to cessation of cell expansion:
Reduction in stress relaxation processes
Increased cross-linking of wall components
New matrix polysaccharides that associate more strongly with each other and/or the cellulose microfibrils.
Enzymes, pectin esterase makes cell walls more rigid. Expose carboxyl groups in pectin.
Lignification.