Endocrinology
Cell-Cell Communication Revisited
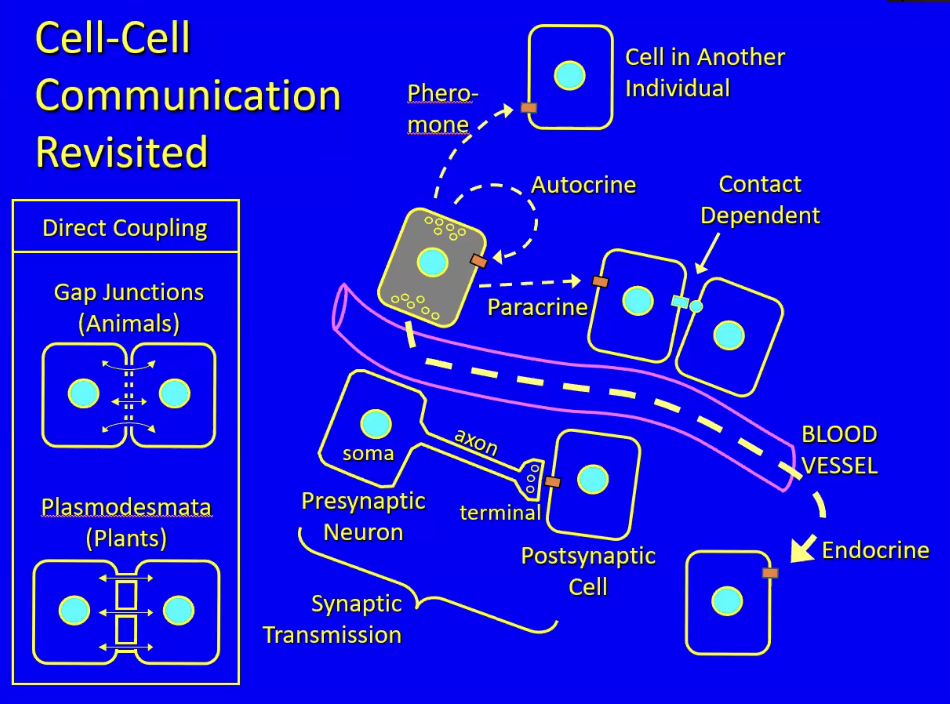
Cells secrete chemical signals (hormones) that are stored vesicles (for those that are hydrophilic) and released via exocytosis. A few things can happen if a hormone is specific or nonspecific
Hormone diffuses into capillary that carries the signal to the rest of the body through the vascular system = classic endocrine signal
Hormone is released into interstitial fluid and stimulates nearby cells with receptors for the hormone = paracrine signaling
When there is a mutual and specific binding between to membranes to convey hormones = Contact dependent signaling
When a nerve cell (with axon and terminal) conveys neurotransmitter to a postsynaptic cell = synaptic transmission (type of paracrine in nervous sys)
When the hormone being secreted is secreted into the external environment and has target cells in other organisms of the same species = pheromone signaling (humans do not have this)
When the cytoplasm of adjacent cells are connected to allow diffusion of hormones = direct coupling (e.g. gap junctions → animals; plasmodesmata → plants)
Hormones of the Posterior Pituitary
The hypothalamus leads down to the posterior pituitary via a stalk
The posterior pituitary is considered nerve tissue while the anterior pituitary is not (anterior pituitary is connected to mouth)
The nerve cells in the hypothalamus control important things
The pituitary gland has two parts:
Posterior pituitary (neurohypophysis)
The hypothalamus has two types of nerve cells that send axons/processes down to the stalk of the posterior pituitary
One type of cell secretes ADH/AVP and the other releases oxytocin. Capillaries pick these hormones up and carry them to their target cells
E.g. Oxytocin → stimulates the muscles of the uterus and the release of preformed milk
The uterus has a middle, muscular layer, and during pregnancy this layer grows thicker, during childbirth the contraction of the muscles that expel the baby are stimulated by oxytocin
Oxytocin is also present in males and acts on the brain’s reward center in both genders to stimulate sexual and social functions, as well as bonding (e.g. skin to skin)
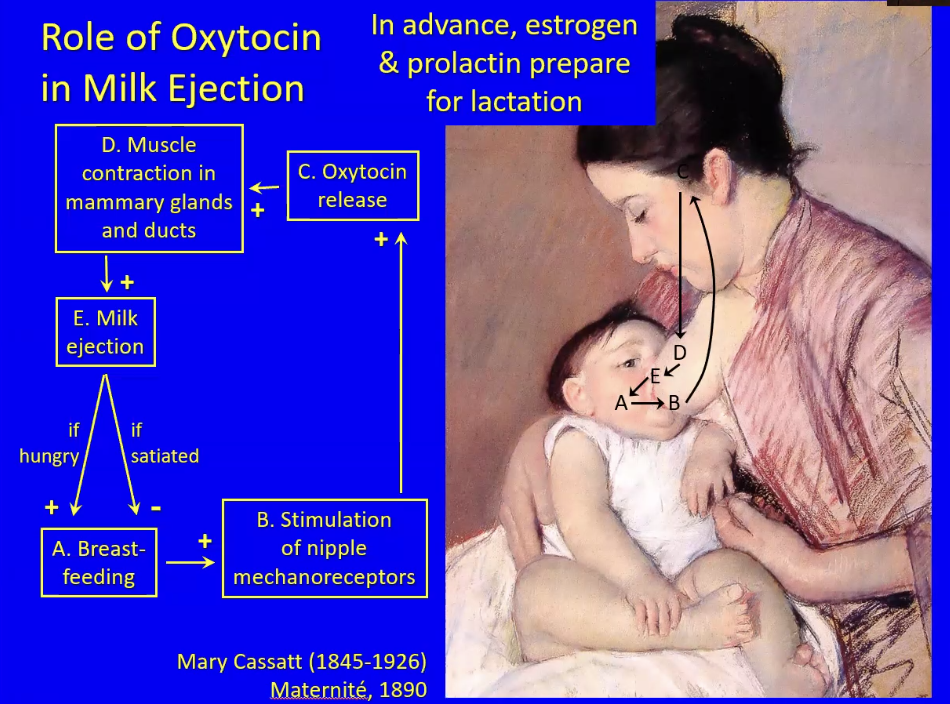
A hungry baby stimulates breastfeeding and in the mother this stimulates mechanoreceptors (touch-sensitive nerves) that connect directly to the hypothalamus. This stimulates the release of oxytocin, and thus the release of milk from the mammary glands via contraction of the smooth muscles around the breast glands/ducts
If the baby is still hungry a positive feedback loop occurs. If the baby is satiated, that interrupts the positive feedback and becomes negative feedback where breastfeeding stops
Estrogen and prolactin cause the growth of the mammary gland tissue NOT the release of milk
Hormones of the Anterior Pituitary
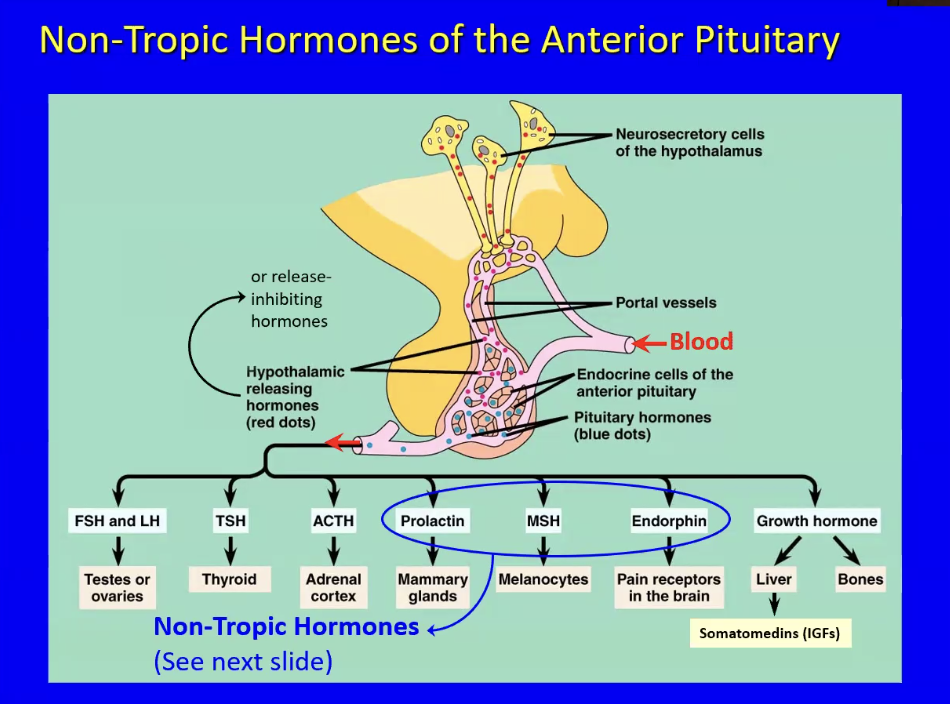
Anterior pituitary (adenohypophysis) releases
Has 7 endocrine cells types that release 8 hormones
Nerve cells in the hypothalamus release hypothalamic releasing hormone which control the release of hormones from the anterior pituitary by binding to specific receptors on anterior pituitary cells
This is possible through secretion of hypothalamic releasing hormones onto a hypothalamic capillary bed, which travel down a portal vein, go down into the anterior pituitary, diffuse out of the capillary, are taken up by the anterior pituitary cell type, and trigger a hormonal response. (AKA production or inhibition)
The blood draining out of the hypothalamus goes through another set of capillaries → this is known as a portal system (which in this case does the process described above)
Mother of all portal systems is the hepatic portal vein
For each of the 7 different endocrine cells in the anterior pituitary, there is a different hypothalamic hormone regulating its activity
The 8 hormones can be classified into two groups:
Tropic Hormones: FSH, LH, TSH, ACTH, GH
Affect endocrine organs that secrete other hormones
FSH and LH (Follicle Stimulating Hormone and Luteinizing Hormone)
Without reproduction our species wouldn’t survive so it needs control
These are tropic hormones under the hypothalamic/anterior pituitary axis
GnRH is released from the hypothalamus
GnRH (gonadotropin releasing hormone) signals the anterior pituitary to release FSH and LH. These are both gonadotropins.
In short, GnRH tells the anterior pituitary to release gonadotropins, hence its name
Gonads are ovaries in females and testes in males. They have two functions…
Production of gametes, the haploid cells of reproduction. Ovaries produce oocytes and testes produce sperm
Endocrine functions.
Ovaries produce estrogens (female reproductive hormones) such as a kkestradiol, the most abundant estrogen. There are other estrogens
Testes produce androgens such as testosterone, which are male-characteristic steroid hormones. There are other androgens
These hormones are secreted beginning at puberty
secreted throughout life in men (though there is a peak in the 20s).
In women, ovulation starts at puberty but ends at menopause
ONLY THE GONADS HAVE RECEPTORS FOR THESE HORMONES - FSH and LH
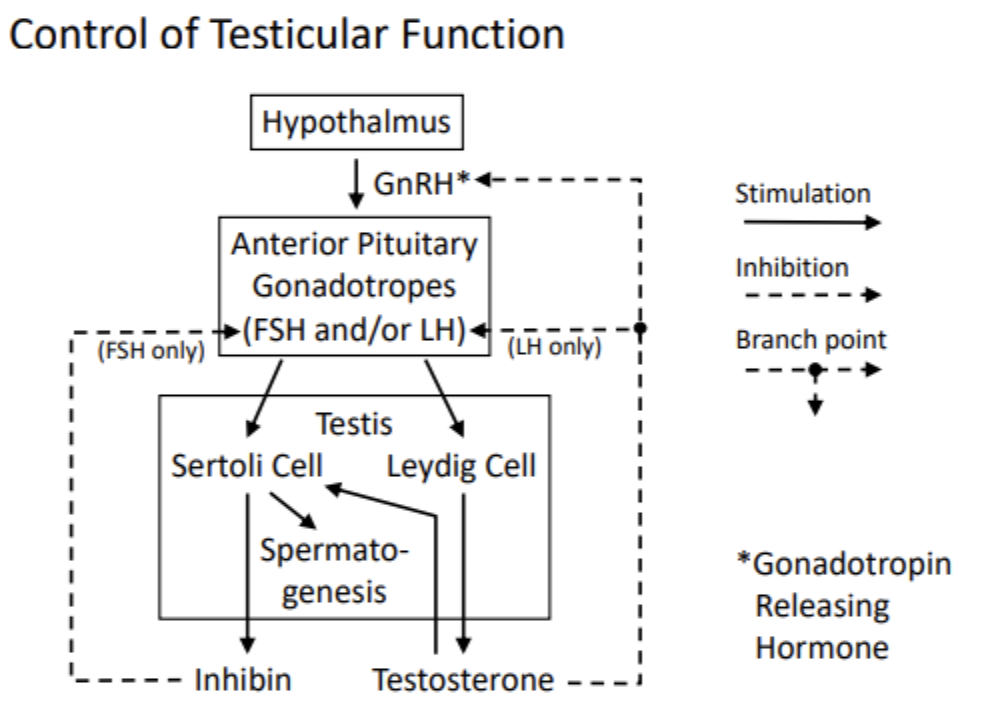
GnRH turns on the system, and is released from the hypothalamus. The reason why we dont make sperm before puberty is because GnRH isnt produced in the first decade of life. At the start of puberty, the hypothalamic neurons make GnRH. This tells the anterior pituitary to release FSH and/or LH.
The testes are composed of a densely packed array of seminiferous tubules. The walls of these tubules are a multilayered epithelium, outside of which lay stem cells that divide, forming another stem cell and a sperm, providing continual production of sperm starting at puberty.
The seminiferous tubules are surrounded by sertoli cells, which provide nourishment to the tubules, and promote spermatogenesis– approximately several hundred sperm per day. Sperm are pushed through the tubule and stored in the epididymis where they sit.
The sertoli cells have receptors for FSH, which trigger the sertoli cells to promote spermatogenesis. Thus, FSH binding to sertoli cells → spermatogenesis
In the spaces between the sertoli cells are leydig cells, which have receptors for LH. When LH binds, testosterone is released from the leydig cell.
Testosterone is produced in high levels, spills out of the testes, binds to carrier proteins, and circulates around the body. This molecule is responsible for male secondary sex characteristics (muscle development, body mass, larynx, pubic hair, etc.)
The highest level of testosterone is in the testes. The leydig cells are located in between the sertoli cells which wrap around the tubules, so the sertoli cells get the highest level of testosterone of any cell in the body
Testosterone, androgens, estrogens are all lipophilic steroid hormones derived from cholesterol
Leydig cells have androgen receptors on the nuclear membrane. The testosterone-receptor complex acts as a transcription factor.
Sertoli cells are highly androgen dependent. The reason why men have more skeletal muscle is male skeletal muscle has more androgen receptors coupled with there being more testosterone in men.
Sertoli cells require testosterone from the leydig cell
Thus, FSH and LH secretion promotes spermatogenesis and release of a protein hormone called inhibin from the sertoli cells
Let’s cover the negative feedback mechanisms
Testosterone binds to the anterior pituitary cells, preventing them from releasing LH but not FSH
Testosterone binds to the hypothalamus and prevents it from releasing GnRH
Thus, testosterone directly inhibits LH and indirectly inhibits FSH and LH
Inhibin is senses by the anterior pituitary, inhibiting FSH release
Although sperm accumulate mutations that increase the risk of birth defects, older men can still have children due to spermatogenesis even in old age
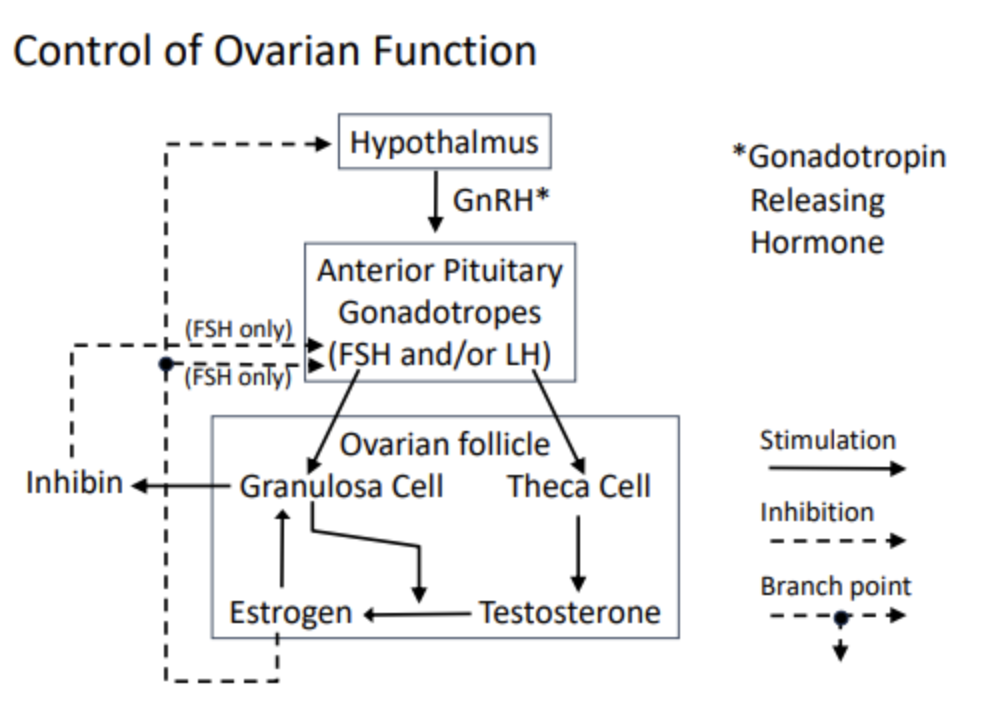
Pretty similar to men
Hypothalamus releases GnRH, which tells the anterior pituitary to release FSH and/or LH.
These hormones target the ovarian follicles, which lie within the ovary
The two cell types in the ovarian follicle are the granulosa and theca cells
The granulosa is analogous to the sertoli cell, which surrounds and nourishes the developing gamete. It is stimulated by estrogen. Testosterone can be converted to estrogen in one enzymatic step– this conversion is stimulated/carried out by the granulosa cell. Granulosa cells only have estrogen receptors, not testosterone receptors.
Positive feedback: a loop is formed between estrogen, testosterone, and granulosa cell. The more testosterone → estrogen, the more the granulosa cells trigger more conversion
The granulosa cells, similar to the sertoli cells, secretes inhibin release, which inhibits FSH release from the anterior pituitary
The theca cell is analogous to the leydig cell in the testes. It is stimulated by LH, and releases testosterone.
The steroid hormone that escapes the ovary is estrogen, which travels through the blood on carrier proteins and goes on to inhibit FSH release from the anterior pituitary and GnRH release from the hypothalamus.
Thus, estrogen directly inhibits FSH and indirectly inhibits both FSh and LH
This hormone cycling creates the menstrual cycle/ovarian cycle in women
Ovary & Follicle Development Summary
Basic Structure:
Each follicle = 1 oocyte + granulosa cells.
At birth: baby girls have hundreds of primary follicles per ovary, each with a single granulosa cell layer around the oocyte.
Follicle Maturation:
Daily from birth, some primary follicles begin maturing into secondary follicles:
Secondary = many granulosa layers + outer theca cell layer.
If FSH & LH levels are right (after puberty), secondary follicles continue developing.
Graafian Follicle Formation:
Follicle enlarges, forms a fluid-filled cavity.
Oocyte is suspended by a stalk of granulosa cells.
As fluid builds, pressure and digestive enzymes weaken the ovary's outer layer → ovulation occurs.
Ovulation:
Around day 14 of a 28-day cycle (± a few days).
Follicle ruptures, releasing oocyte + some granulosa cells into abdominal cavity.
Oocyte is swept into oviduct/fallopian tube by cilia.
Fertilization occurs in the oviduct, if sperm is present.
Fertilization Window:
Oocyte survives ~1 day.
Sperm can survive 7–10 days in reproductive tract.
So, intercourse up to 10 days before ovulation can result in pregnancy.
Post-Ovulation:
Remaining follicle cells form the corpus luteum under LH stimulation.
Corpus luteum = temporary endocrine organ, secretes estrogen + progesterone.
Lifespan: ~10 days if no fertilization → then degenerates to scar tissue.
Cycle Details:
Follicle/oocyte development starts daily from birth (~30/day), but only continues past primary stage after puberty.
Most oocytes die off before puberty.
Menopause = depletion of oocytes.
One oocyte typically ovulates per cycle; two = fraternal twins.
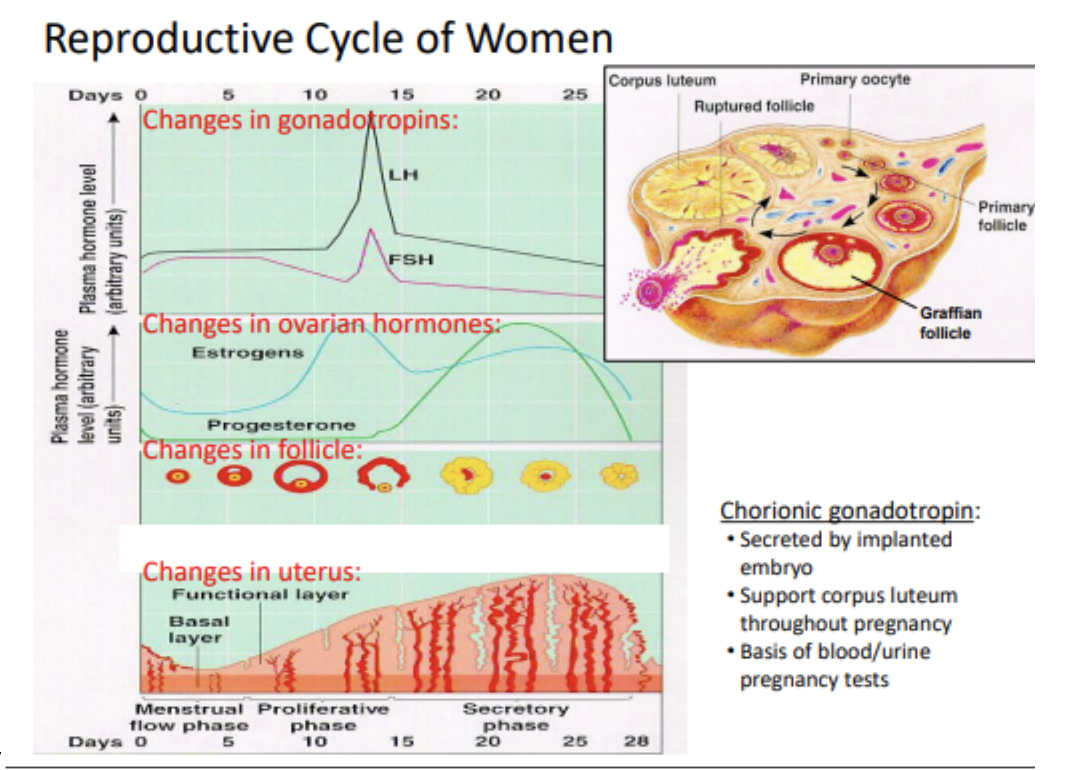
Hormone Changes Summary
Early Cycle (Days 1–6):
FSH stimulates follicle growth (primary → secondary → Graafian).
Granulosa cells produce estrogen using testosterone from theca cells.
Low estrogen levels = negative feedback on FSH and LH → keeps them flat.
Mid Cycle (Days 7–14):
Rising estrogen from growing follicle → increased production (positive feedback).
Around Day 12: Estrogen surge triggers LH surge (+ smaller FSH surge).
Day 14: Ovulation occurs due to LH surge; follicle ruptures.
Post-ovulation: LH/FSH drop again; estrogen falls slightly.
Late Cycle (Days 15–28):
Follicle remnants become corpus luteum, secreting estrogen + progesterone.
Progesterone: maintains endometrium, develops glands for glycogen secretion.
If no fertilization, corpus luteum dies after ~7–10 days → hormone levels fall.
Drop in estrogen/progesterone → prostaglandin not inhibited → spiral arteries constrict → endometrial necrosis and menstruation (bleeding = Day 0).
Endometrium Changes:
Estrogen: stimulates thickening from basal layer starting Day 3–4.
Spiral arteries grow to supply blood.
Progesterone: differentiates glands to support zygote.
Without hormones, prostaglandin causes ischemia → tissue dies → menstruation.
If Fertilization Occurs:
Implantation ~Day 21–24.
Embryo forms chorion, which secretes chorionic gonadotropin (hCG).
hCG mimics LH → saves corpus luteum → continues estrogen/progesterone production.
Supports pregnancy and uterine growth.
hCG is detected in pregnancy tests.
TSH (Thyroid Stimulating Hormone = Thyrotropin)
Release is controlled by TRH (thyrotropin releasing hormone)
Affect the thyroid gland in the neck, which which releases the thyroid hormones T3= Triiodothyronine and T4=Tetraiodothyronine
Synthesized from Tyr in thryroglobin molecules
Mammals secrete mainly T4 but T3 is more potentn
Stimulate: cellular metabolism, protein synthesis, O2 consumption, Lipid breakdown, ATP production, Heat Production
Effects of Thyroid Hormones
Hyperthyroidism
High temp, sweating, irritability
Graves’ disease: autoantibodies stimulate TSH receptors and expothalmos (wide, bulging eyse)
Hypothyroidism
Weight gain, lethargy, intolerance to cold
In infants → congenital iodine deficiency syndrome
Due to autoimmune destruction of thyroid (Hashimoto’s) pr dietary iodine deficiency
Lack of negative feedback can cause goiter
Negative feedback regulation → The production of T3 and T4 inhibits the release of TRH from the hypothalamus and TSH from the anterior pituitary
The cross-section of a thyroid gland shows the thyroid follicle consisting of colloid material and epithelial cells, that secrete thyroglobulin, enzymes, and iodine
Thyroglobulin is rich in tyrosine residues (an amino acid with a six carbon ring and hydroxyl group). On tyrosine’s six carbon ring, there can be iodine atoms added on.
Follicle cells concentrate and secrete iodide ions into the colloid, along with synthetic enzymes, which attach iodine onto the rings of tyrosine to form monoiodinated or diiodinated tyrosines
Other enzymes move the carbon rings from one iodinated tyrosine onto another iodinated tyrosine, cross-linking them to form a dimer-like molecule.
If a diiodinated tyrosine is linked to a monoiodinated tyrosine, then T3 is formed
If a diiodinated tyrosine is linked to another diiodinated tyrosine, then T4 is formed
TSH stimulates the thyroid follicle cells to phagocytose some of the colloid, which contains deactivated+iodinated tyrosine dimers. Once inside the cell, the lysosomes digest the thyroglobulin, cleaving the peptide backbones of the dimer/thyroglobulin, releasing the active hormone.
Once liberated, the lipophilic T3 and T4 diffuses out of the cell and into the capillary, where it binds to specific and nonspecific carrier proteins produced by the liver. The hormones then go around to the body
This is the only place where iodine is used in our body– to make thyroid hormones
ACTH (Adrenalcorticotropic Hormone or Corticotropin
Targets the adrenal glands that sit atop of the kidneys
Consists of two parts: inner medulla and outer cortex
The outer part releases a bunch of steroid hormones (synthesized from cholesterol = corticosteriods → lipophilic hormones
Mineralocorticoids
Aldosterone: sodium ion and water reabsorption, increased blood volume and pressure
Glucocorticoids
Cortisol: gluconeogenisis, muscle catabolism, immunosuppression (incl. anti-inflammatory effects
Cortisol is a principle response to stress
ensures the body has glucose available for metabolic fuel.
Hypoglycemia can kill the brain due to metabolic failure
Hyperglycemia is more of a long term problem
We have overlapping hormone systems in place to ensure hypoglycemia doesn’t happen
Cortisol is the most abundant glucocorticoid in humans, but there are others
Cortisol functions to maintain glycogen stores in the liver. Note that we also store glycogen in the muscle.
Glucagon functions to breakdown liver glycogen. Cortisol is the hormone that makes sure that there is always glycogen in the liver. In short, cortisol makes sure glucagon works
Cortisol also helps people survive in prolonged periods of fasting and food deprivation. The relative abundance of food for us to eat is pretty new– our bodies aren’t really used to having excess nutrition. Thus, our ancestors had periods of food deprivation, and therefore adapted mechanisms to survive– this mechanism is cortisol. Takes about a month for someone to die of starvation, given that they have sufficient water.
Cortisol promotes muscle catabolism– it takes muscle protein in contractile fibers, breaks it down, releases it into the blood, and converts it to liver glycogen. This process doesn’t kill the muscle, it just makes it skinnier, so it is still functional.
Cortisol stimulates gluconeogenesis: the formation of glucose from non-carbohydrates. Deaminating an amino acid creates an intermediate that can feed into carbohydrate pathways. This occurs in the liver– some of the glucose is released into the blood and some is stored.
In short, cortisol tells the body to break down its own muscle for fuel
Cortisol is also an immunosuppressant– slows antibody secretion, T cell activity, cytokine secretion, etc.
Not evolutionarily clear why we developed this. Scientists speculate that when in extreme conditions like starvation, trauma, and other stresses, both physical and mental– there is cortisol release. If you’re in a stressful situation– there is no time to feel sick!
Cortisone is a synthetic derivative of cortisol. It is a potent anti-inflammatory agent.
Taking oral glucocorticoid pills/steroid pills/injections are typically more potent, relieving inflammation but also suppressing the immune system, leaving you susceptible to cancer/infection.
Sex steroids
Androgens in men
Estrogens and progestins in women
Inner part releases epinephrine (flight/fight hormone) and is controlled by sympathetic nerves
Epinephrine is a principle response to stress
The effects of epinephrine are widespread– dilated pupils, blood flows shifted to skeletal muscles/brain/heart at the expense of internal organs/skin/digestive tract blood flow.
Plasma glucose rises very quickly. Liver glycogen is broken down and used as fuel in our brain and skeletal muscle during short term emergencies.
Cortisol is more of a long term (hours to days) hormone and epinephrine is more of a short term (seconds to a few minutes) hormone. Remember that glucagon is also involved, and maintains glucose in between meals at a medium time scale (minutes to hours). These hormone systems all overlap to some degree.
Release is controlled by CRH (corticotropin releasing hormone)
Stimulates the growth of adrenal cortex and secretion of steroid hormones → exert negative feedback: inhibit secretion of ACTH and CRH
GH (Growth Hormone)
Both a tropic and non-tropic hormone (activates liver and stimulates growth of muscle/bone)
Stimulated by: GHRH (Growth Hormone Releasing Hormone) from the hypothalamus.
Inhibited by: GHIH (Growth Hormone Inhibiting Hormone / Somatostatin).
Somatostatin also slows digestion and inhibits growth in the GI tract and brain
Direct Effects: Stimulates growth in muscles, bones, etc.
Indirect Effects: Stimulates liver to release IGFs (Insulin-like Growth Factors).
IGFs promote cell division, protein synthesis, glucose uptake.
GH contributes to lean body mass by promoting mitosis, protein synthesis, and fat breakdown (lipolysis).
Pituitary Dwarfism (GH Deficiency):
Caused by defects in GH/IGF production or their receptors (200+ causes).
Results in short stature and disproportionate body size, but normal brain development.
Lifespan is normal.
Requires functional GHRH, GH, IGFs, and their receptors for normal growth.
Gigantism & Acromegaly (GH Excess):
Caused by GH-secreting tumor (usually benign).
Gigantism: Occurs before bone growth plates close → excessive height.
Acromegaly: Occurs after growth plates close → enlargement of hands, skull, organs.
Leads to pain, organ overgrowth, and shortened lifespan (3–4 decades).
GH secretion in these cases ignores normal negative feedback.
Non-Tropic Hormones: Prolactin, MSH, Endorphin
Prolactin
stimulates the growth of the mammary glands and secretion of milk.
Milk is produced after pregnancy because estrogen inhibits the effect of prolactin on milk production. Thus after birth estrogen levels drop allows for prolactin to have an effect
During pregnancy high levels of estrogens and prolactin make breasts larger by increasing fat deposition. Pigment of nipple increases, and ducts/glands grow
Its release is stimulated by PRH (prolactin releasing hormone and is inhibited by Dopamine (prolactin inhibiting hormone)
In men/women, results in the sexual refractory. (where men achieve a second orgasm)
MSH (Melanocyte Stimulating Hormone)
Disperses pigment (present in all vertebrates) in melanocytes to darken skin
Melanin dispersion in granules is variable not fixed → in animals
In humans, is a satiety signal in brain (inhibits hunger signal)
Endorphin
Naturally occurring opiates
Why do opiates bind to our brain cells? Because there are naturally occuring opiates released from the anterior pituitary called endorphins. It just so happens that morphine has the same binding domain as endorphin. Plants like opium evolved this so that animals would get sick after eating it. Endorphins help us survive in emergency situations where we would otherwise be impaired by pain
Inhibit pain, induce euphoria.