
23 Study Guide-Genetics
A STUDY OF CLASSICAL (MENDELIAN) GENETICS
and
INTRODUCTION TO MOLECULAR GENETCIS
STUDY NOTES – Chapter 23
Genetics is the study of how characteristics are transmitted from parents to offspring.
Gregor Mendel is known as the father of genetics.
Mendel’s Garden peas
Seven characteristics were observed
MOLECULAR GENETICS STUDY NOTES
Just what is a gene, anyway? What is it made of, and how does it do what it does?
This was a question for the biochemists. And they had a good start on the answer already, because the Mendelian geneticists had already answered the question of where genes were in the cell. They were in the nucleus. More specifically, they were in the chromosomes, a fact that was established before the end of the first decade of the century.
So genes were in chromosomes. And everyone knew what chromosomes were made of. They were about 50% histones (a class of protein) and 50% deoxyribonucleic acid (DNA). So if genes were in chromosomes, then they must either be made of protein or of DNA. But which?
Whatever genes were made of, it had to be a substance that could carry information--that's what "instructions" are, information. So the question became, "Which of these molecules is a better candidate for an information-bearing molecule?" It may surprise you to discover that the smart money was on protein, not DNA, as the answer.
Both protein and DNA are made out of long chains of smaller molecules. Proteins are constructed out of a set of twenty different amino acids. A typical protein contains perhaps 150 amino acids, selected from that "library" of twenty amino acids, and strung together in a specific order. The biochemists of the later part of the first half of the twentieth century knew this. Protein... twenty different amino acids.
DNA is made out of smaller molecules called nucleotides. Like the amino acids in proteins, these nucleotides are strung together in long chains, though this was something that was not yet known about DNA structure at the time this puzzle was under consideration. But here's the catch... Where proteins are made from a selection of twenty different amino acids, DNA is made from only four different nucleotides.
A series of extremely clever experiments in the 1940's demonstrated without possibility of doubt that the part of a chromosome that carries genetic information is not protein, it's DNA.
Rosalind Franklin developed her interest in science at school, and by age 15 had decided to become a chemist. She had to overcome the opposition of her father, who did not want her to attend college or become a scientist; he preferred that she go into social work. She earned her Ph.D. in chemistry in 1945 at Cambridge.
Rosalind Franklin joined the scientists at the Medical Research Unit, King's College, when John Randall recruited her to work on the structure of DNA. DNA (deoxyribonucleic acid) was originally discovered in 1898 by Johann Miescher, and it was known that it was a key to genetics. But it was not until the middle of the 20th century when scientific methods had developed to where the actual structure of the molecule could be discovered, and Rosalind Franklin's work was key to that methodology.
Rosalind Franklin worked on the DNA molecule from 1951 until 1953. Using x-ray crystallography she took photographs of the B version of the molecule. A co-worker with whom Franklin did not have a good working relationship, Maurice H. F. Wilkins, Wilkins showed Franklin's photographs of DNA to James Watson, without permission of Franklin. Watson and his research partner, Francis Crick, were working independently on the structure of DNA, and Watson realized that these photographs were the scientific evidence they needed to prove that the DNA molecule was a double-stranded helix.
While Watson, in his account of the discovery of the structure of DNA, largely dismissed Franklin's role in the discovery, Crick later admitted that Franklin had been "only two steps away" from the solution, herself.
Randall had decided that the lab would not work with DNA, and so by the time her paper was published, she had moved on to Birkbeck College and the study of the structure of the tobacco mosaic virus, and she showed the helix structure of the virus' RNA. She worked at Birkbeck for John Desmond Bernal and with Aaron Klug, whose 1982 Nobel Prize was based in part on his work with Franklin.
This discovery set off a flurry of interest in DNA. Laboratories all over the world forged ahead in the study of the structure of this molecule that everyone had considered to be essentially unimportant. A sort of unofficial race among scientists around the world was eventually won by a pair of dark horses in England, James Watson and Frances Crick. Their discovery of the structure of DNA, published in 1953, set off an explosion in an entirely new kind of genetics: molecular genetics.
The race to figure out the structure of DNA ended when a young American biochemist named James Watson and a British physicist named Francis Crick published a short paper in 1953. The structure they proposed was both simple and elegant, and complex enough to show how this deceptively monotonous molecule was capable of carrying so much information.
Watson, Crick, and Wilkins were awarded the Nobel Prize in physiology and medicine in 1962, four years after Franklin died. The Nobel Prize rules limit the number of persons for any award to three, and also limit the award to those who are still alive, so Franklin was not eligible for the Nobel. Nevertheless, many have thought that she deserved explicit mention in the award, and that her key role in confirming the structure of DNA was overlooked because of her early death and the attitudes of the scientists of the time towards women scientists.
Watson's book recounting his role in the discovery of DNA displays his dismissive attitude towards "Rosy." Crick's description of Franklin's role was less negative than Watson's, and Wilkins mentioned Franklin when he accepted the Nobel. Anne Sayre wrote a biography of Rosalind Franklin, responding to the lack of credit given to her and the descriptions of Franklin by Watson and others. The wife of another scientist at the laboratory, herself a friend of Franklin, Sayre describes the clash of personalities and the sexism that had faced Franklin in her work. A. Klug used Franklin's notebooks to show how close she had come to independently discovering the structure of DNA.
In 2004, Finch University of Health Sciences/The Chicago Medical School changed its name to the Rosalind Franklin University of Medicine and Science, to honor Franklin's role in science and medicine.
The study of DNA and its functions can be very complex and technical — it is an extremely large molecule, and its functions are the intricate and complicated functions of large molecule chemistry. But a lot of the important things we understand about DNA aren't complicated. They are elegant and beautiful in their simplicity.
DNA is information bearing molecule — a language molecule. It is constructed out of very long chains of four different small composite molecules, and the order of these four molecules determines the message coded into the molecule, just as the order of letters in a word determines the meaning of the word. If this weren't impressive enough, each of these extremely long chains fits precisely against a partner chain in such a way that each of them precisely complements the order of the four components its partner. Because of this aspect of DNA, each molecule can split its two chains apart and build a new matching strand for each — thus duplicating the original molecule. Every time a cell's nucleus goes through mitosis or meiosis, the DNA in the chromosomes of that nucleus must first complete this copying process to produce the two copies of each chromosome necessary for the daughter nuclei. ![]()
Because of the three-dimensional shape of the structure they discovered, DNA has come to be described as a "double helix." A helix is a spiral shape, like a slinky or the hand rail of a spiral staircase. These two strands wrap around each other, creating that double helical shape which has become so well known.
This shape is more than just a geometric form. The double stranded nature of DNA turns out to be a vital functional part of its structure--it couldn't do what it does if it weren't double stranded.
To understand this importance, it's necessary to take a bit closer look at the structures out of which a DNA molecule is made: nucleotides.
Nucleotides are made out of three small molecules bonded together: a sugar, a phosphate group and a nitrogenous base. In DNA, all of the nucleotides contain the sugar deoxyribose (thus the name, "deoxyribonucleic acid"). The nucleotides in RNA contain a slightly different sugar called ribose" (thus "ribonucleic acid).
The nucleotides that make up a DNA molecule come in four types. All of these deoxyribonucleotides are identical in their sugar and phosphate portions. But there are four different kinds of nitrogenous bases, called adenine, guanine, cytosine and thymine. (RNA's ribonucleotides also come in adenine, guanine, and cytosine versions. RNA doesn't have any thymine in it; instead, it has a very similar base called uracil.) It is very common among biologists and chemists to simply refer to these four deoxyribonucleotides as A, G, C and T.
DNA isn't simply a single string of nucleotides--its two strands, side by side and twisted around each other. It turns out that the two strands are held together by interactions between the bases on one strand and the bases on the other--they pair up, holding the two sides together. And the rules about pairing are very strict--Adenine will pair only with Thymine; Guanine will pair only with Cytosine. So Adenine and Thymine are complements of each other, and Guanine and Cytosine are complements of each other. (To be complementary means to "fit together.") This base pairing is strictly maintained in all DNA. So once the base sequence of one side is set, the sequence of the other side is automatic.
A final aspect of the structure of DNA is that the way the two sides’ pair together forces a twist in each of the backbones, creating the famous and beautiful three dimensional shape of a double helix--two helices (coils) wrapped around each other.
Looking at the structure they'd proposed, Watson and Crick immediately realized the potential for self-replication intrinsic to complementary base pairing. Each side of this molecule imposes the order of the other side. So if you separate the two sides ("unzip" the molecule), and assemble new partners out of free (single) nucleotides, the bases of the old chains will select the "correct" partners by complementary base pairing, and you will end up with two new molecules, identical to each other and to the original molecular. Though the specifics of DNA replication are much more complex than this simple model, in fact what happens to the molecule is very much exactly what Watson and Crick expected.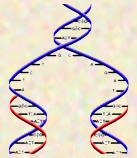
So complementary base pairing explains this vital ability of the genetic material--self-replication. But that's not all DNA has to do.
One more component is a very interesting group of small RNA's called Transfer RNA. The Transfer RNA is the adapters which allow the nucleotide language of DNA and RNA to be converted to the amino acid language of proteins.
The two processes involved in making protein are called transcription and translation.
The word "transcription" means "copy." In the transcription process, a segment of DNA (typically, though not always, a single gene) is used to create a messenger RNA which contains a copy of the information in the gene. Remember that the structure of DNA and the structure of RNA are very similar. This messenger RNA is created by complementary base pairing between ribonucleotides (to make RNA) and one side of the DNA. Uracil behaves just like Thymine in base pairing. If DNA is our cookbook, messenger RNA is a recipe card. DNA contains the entire library of the cell's genetic information; messenger RNA typically contains the information for only one protein. And messenger RNA is easily replaceable--if it gets damaged or degraded, all the cell has to do is transcribe the gene again.
Translation" means "convert from one language to a different language." In the translation process, a ribosome reads the information coded into a messenger RNA & constructs a string of amino acids according to those instructions. The information in the messenger RNA is interpreted by the ribosome in three-base "words" called codons. Each codon is translated into a particular amino acid.
A STUDY OF CLASSICAL (MENDELIAN) GENETICS
and
INTRODUCTION TO MOLECULAR GENETCIS
STUDY NOTES – Chapter 23
Genetics is the study of how characteristics are transmitted from parents to offspring.
Gregor Mendel is known as the father of genetics.
Mendel’s Garden peas
Seven characteristics were observed
MOLECULAR GENETICS STUDY NOTES
Just what is a gene, anyway? What is it made of, and how does it do what it does?
This was a question for the biochemists. And they had a good start on the answer already, because the Mendelian geneticists had already answered the question of where genes were in the cell. They were in the nucleus. More specifically, they were in the chromosomes, a fact that was established before the end of the first decade of the century.
So genes were in chromosomes. And everyone knew what chromosomes were made of. They were about 50% histones (a class of protein) and 50% deoxyribonucleic acid (DNA). So if genes were in chromosomes, then they must either be made of protein or of DNA. But which?
Whatever genes were made of, it had to be a substance that could carry information--that's what "instructions" are, information. So the question became, "Which of these molecules is a better candidate for an information-bearing molecule?" It may surprise you to discover that the smart money was on protein, not DNA, as the answer.
Both protein and DNA are made out of long chains of smaller molecules. Proteins are constructed out of a set of twenty different amino acids. A typical protein contains perhaps 150 amino acids, selected from that "library" of twenty amino acids, and strung together in a specific order. The biochemists of the later part of the first half of the twentieth century knew this. Protein... twenty different amino acids.
DNA is made out of smaller molecules called nucleotides. Like the amino acids in proteins, these nucleotides are strung together in long chains, though this was something that was not yet known about DNA structure at the time this puzzle was under consideration. But here's the catch... Where proteins are made from a selection of twenty different amino acids, DNA is made from only four different nucleotides.
A series of extremely clever experiments in the 1940's demonstrated without possibility of doubt that the part of a chromosome that carries genetic information is not protein, it's DNA.
Rosalind Franklin developed her interest in science at school, and by age 15 had decided to become a chemist. She had to overcome the opposition of her father, who did not want her to attend college or become a scientist; he preferred that she go into social work. She earned her Ph.D. in chemistry in 1945 at Cambridge.
Rosalind Franklin joined the scientists at the Medical Research Unit, King's College, when John Randall recruited her to work on the structure of DNA. DNA (deoxyribonucleic acid) was originally discovered in 1898 by Johann Miescher, and it was known that it was a key to genetics. But it was not until the middle of the 20th century when scientific methods had developed to where the actual structure of the molecule could be discovered, and Rosalind Franklin's work was key to that methodology.
Rosalind Franklin worked on the DNA molecule from 1951 until 1953. Using x-ray crystallography she took photographs of the B version of the molecule. A co-worker with whom Franklin did not have a good working relationship, Maurice H. F. Wilkins, Wilkins showed Franklin's photographs of DNA to James Watson, without permission of Franklin. Watson and his research partner, Francis Crick, were working independently on the structure of DNA, and Watson realized that these photographs were the scientific evidence they needed to prove that the DNA molecule was a double-stranded helix.
While Watson, in his account of the discovery of the structure of DNA, largely dismissed Franklin's role in the discovery, Crick later admitted that Franklin had been "only two steps away" from the solution, herself.
Randall had decided that the lab would not work with DNA, and so by the time her paper was published, she had moved on to Birkbeck College and the study of the structure of the tobacco mosaic virus, and she showed the helix structure of the virus' RNA. She worked at Birkbeck for John Desmond Bernal and with Aaron Klug, whose 1982 Nobel Prize was based in part on his work with Franklin.
This discovery set off a flurry of interest in DNA. Laboratories all over the world forged ahead in the study of the structure of this molecule that everyone had considered to be essentially unimportant. A sort of unofficial race among scientists around the world was eventually won by a pair of dark horses in England, James Watson and Frances Crick. Their discovery of the structure of DNA, published in 1953, set off an explosion in an entirely new kind of genetics: molecular genetics.
The race to figure out the structure of DNA ended when a young American biochemist named James Watson and a British physicist named Francis Crick published a short paper in 1953. The structure they proposed was both simple and elegant, and complex enough to show how this deceptively monotonous molecule was capable of carrying so much information.
Watson, Crick, and Wilkins were awarded the Nobel Prize in physiology and medicine in 1962, four years after Franklin died. The Nobel Prize rules limit the number of persons for any award to three, and also limit the award to those who are still alive, so Franklin was not eligible for the Nobel. Nevertheless, many have thought that she deserved explicit mention in the award, and that her key role in confirming the structure of DNA was overlooked because of her early death and the attitudes of the scientists of the time towards women scientists.
Watson's book recounting his role in the discovery of DNA displays his dismissive attitude towards "Rosy." Crick's description of Franklin's role was less negative than Watson's, and Wilkins mentioned Franklin when he accepted the Nobel. Anne Sayre wrote a biography of Rosalind Franklin, responding to the lack of credit given to her and the descriptions of Franklin by Watson and others. The wife of another scientist at the laboratory, herself a friend of Franklin, Sayre describes the clash of personalities and the sexism that had faced Franklin in her work. A. Klug used Franklin's notebooks to show how close she had come to independently discovering the structure of DNA.
In 2004, Finch University of Health Sciences/The Chicago Medical School changed its name to the Rosalind Franklin University of Medicine and Science, to honor Franklin's role in science and medicine.
The study of DNA and its functions can be very complex and technical — it is an extremely large molecule, and its functions are the intricate and complicated functions of large molecule chemistry. But a lot of the important things we understand about DNA aren't complicated. They are elegant and beautiful in their simplicity.
DNA is information bearing molecule — a language molecule. It is constructed out of very long chains of four different small composite molecules, and the order of these four molecules determines the message coded into the molecule, just as the order of letters in a word determines the meaning of the word. If this weren't impressive enough, each of these extremely long chains fits precisely against a partner chain in such a way that each of them precisely complements the order of the four components its partner. Because of this aspect of DNA, each molecule can split its two chains apart and build a new matching strand for each — thus duplicating the original molecule. Every time a cell's nucleus goes through mitosis or meiosis, the DNA in the chromosomes of that nucleus must first complete this copying process to produce the two copies of each chromosome necessary for the daughter nuclei. ![]()
Because of the three-dimensional shape of the structure they discovered, DNA has come to be described as a "double helix." A helix is a spiral shape, like a slinky or the hand rail of a spiral staircase. These two strands wrap around each other, creating that double helical shape which has become so well known.
This shape is more than just a geometric form. The double stranded nature of DNA turns out to be a vital functional part of its structure--it couldn't do what it does if it weren't double stranded.
To understand this importance, it's necessary to take a bit closer look at the structures out of which a DNA molecule is made: nucleotides.
Nucleotides are made out of three small molecules bonded together: a sugar, a phosphate group and a nitrogenous base. In DNA, all of the nucleotides contain the sugar deoxyribose (thus the name, "deoxyribonucleic acid"). The nucleotides in RNA contain a slightly different sugar called ribose" (thus "ribonucleic acid).
The nucleotides that make up a DNA molecule come in four types. All of these deoxyribonucleotides are identical in their sugar and phosphate portions. But there are four different kinds of nitrogenous bases, called adenine, guanine, cytosine and thymine. (RNA's ribonucleotides also come in adenine, guanine, and cytosine versions. RNA doesn't have any thymine in it; instead, it has a very similar base called uracil.) It is very common among biologists and chemists to simply refer to these four deoxyribonucleotides as A, G, C and T.
DNA isn't simply a single string of nucleotides--its two strands, side by side and twisted around each other. It turns out that the two strands are held together by interactions between the bases on one strand and the bases on the other--they pair up, holding the two sides together. And the rules about pairing are very strict--Adenine will pair only with Thymine; Guanine will pair only with Cytosine. So Adenine and Thymine are complements of each other, and Guanine and Cytosine are complements of each other. (To be complementary means to "fit together.") This base pairing is strictly maintained in all DNA. So once the base sequence of one side is set, the sequence of the other side is automatic.
A final aspect of the structure of DNA is that the way the two sides’ pair together forces a twist in each of the backbones, creating the famous and beautiful three dimensional shape of a double helix--two helices (coils) wrapped around each other.
Looking at the structure they'd proposed, Watson and Crick immediately realized the potential for self-replication intrinsic to complementary base pairing. Each side of this molecule imposes the order of the other side. So if you separate the two sides ("unzip" the molecule), and assemble new partners out of free (single) nucleotides, the bases of the old chains will select the "correct" partners by complementary base pairing, and you will end up with two new molecules, identical to each other and to the original molecular. Though the specifics of DNA replication are much more complex than this simple model, in fact what happens to the molecule is very much exactly what Watson and Crick expected.
So complementary base pairing explains this vital ability of the genetic material--self-replication. But that's not all DNA has to do.
One more component is a very interesting group of small RNA's called Transfer RNA. The Transfer RNA is the adapters which allow the nucleotide language of DNA and RNA to be converted to the amino acid language of proteins.
The two processes involved in making protein are called transcription and translation.
The word "transcription" means "copy." In the transcription process, a segment of DNA (typically, though not always, a single gene) is used to create a messenger RNA which contains a copy of the information in the gene. Remember that the structure of DNA and the structure of RNA are very similar. This messenger RNA is created by complementary base pairing between ribonucleotides (to make RNA) and one side of the DNA. Uracil behaves just like Thymine in base pairing. If DNA is our cookbook, messenger RNA is a recipe card. DNA contains the entire library of the cell's genetic information; messenger RNA typically contains the information for only one protein. And messenger RNA is easily replaceable--if it gets damaged or degraded, all the cell has to do is transcribe the gene again.
Translation" means "convert from one language to a different language." In the translation process, a ribosome reads the information coded into a messenger RNA & constructs a string of amino acids according to those instructions. The information in the messenger RNA is interpreted by the ribosome in three-base "words" called codons. Each codon is translated into a particular amino acid.