Chapter 13
A long time ago, scientists knew chromosomes were primarily made of protein and DNA, but did not know what actually made up genes
DNA composed of four nucleotides while proteins contained 20 distinct amino acids, suggesting proteins had greater capacity for storing information. A series of experiments in the 1920s to 1950s determined DNA is the genetic material
Frederick Griffith (1928): Studied Streptococcus Pneumoniae, a pathogenic bacterium causing pneumonia.
Two strains of Streptococcus
S strain is virulent
R strain is nonvirulent
Griffith infected mice with both strains, hoping to understand the difference between the strains
His hypothesis: Material in dead bacteria cells can genetically transform living bacterial cells → found to be true
Transforming principle: Griffith showed that a nonvirulent strain of bacteria could become virulent when exposed to a killed virulent strain, suggesting the existence of a transferable genetic factor later confirmed to be DNA
He did not know the mechanism for the movement of genetic information, but the modern interpretation is that genetic material was physically transferred between the cells
Transformation: the transfer of virulence from the dead S strain cells into the live R strain cells (Griffith)
Hershey & Chase (1952): Investigated genetic material using bacteriophages- viruses that infect bacteria
Bacteriophages are composed of only DNA and protein
Wanted to determine which of these molecules is the genetic material that is injected into the bacteria
Hershey and Chase showed that DNA enters bacteria during infection. This confirms DNA genetic material.
Used radioactive phosphorus (32P) to track and label bacteriophage DNA and radioactive sulfur (35S) to label bacteriophage protein. They observed that only the bacteriophage DNA (32P) entered the bacteria and was used to produce more bacteriophage, demonstrating that the injected DNA was responsible for directing the production of new viruses within the bacteria.
Conclusion: DNA is the genetic material
Rosalind Franklin: performed x-ray diffraction studies to identify the 3D structure of DNA (captured an important photo)
She discovered that DNA is helical
Franklin aided the research of Watson and Crick
Watson and Crick (1953): deduced the structure of DNA using evidence from Chargaff, Franklin, and others
Did not perform a single experiment themselves related to DNA
Key insight into their molecules was that each DNA molecule was made of two intertwined chains of nucleotides, which is a double helix structure
DNA consists of two strands of nucleotide monomers linked together
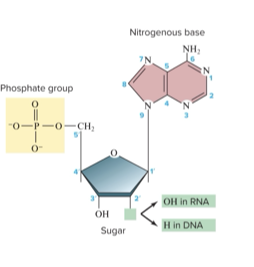
DNA Composition: a nucleic acid made up of nucleotides:
5-carbon sugar called deoxyribose
Phosphate group (PO4) attached to the 5' carbon of the sugar
Nitrogenous base (adenine, thymine, cytosine, or guanine)
Free hydroxyl group (-OH) attached to the 3' carbon of the sugar
DNA Structure:
The two DNA strands of nucleotides pair up and twist around each other to form a spiral-shaped double helix
Sugars and phosphates form the outside backbone
Bases form the internal “rungs”
DNA shape is semiconservative
Each parent strand is a template. New molecules have one old and one new strand- conserves half of the parental DNA in each new copy
Antiparallel strands: direction is determined by sugar-phosphate bonds (two strands run in opposite directions to each other, meaning one strand is oriented 5' to 3' while the other is oriented 3' to 5')
Phosphate groups connect the 3’C of one sugar with the 5’C of the next
One strand has a free 5’ phosphate group- the 5’ end
The other chain has a free 3’ hydroxyl group – the 3’ end
The double helix:
The two DNA strands are arranged as a double helix
Forms two grooves, the larger major groove, and the smallest minor groove
Major groove: wider and more accessible to proteins, with larger spaces between the backbones.
Minor groove: narrower and has less space for protein access
Strands connected via hydrogen bonds between bases on opposite strands
Chargaff’s rule: equal ratios of complementary base pairs contribute to the stability and structure of the DNA double helix
There is always an equal proportion of two-ringed purines (A and G) and single-ringed pyrimidines (C and T)
Amount of adenine = amount of thymine
Amount of cytosine = amount of guanine
The ratio of A-T and G-C varies by species
DNA Replication:
Hydrogen bonds that hold base pairs together are broken and the helicase enzyme unwinds the DNA helix
The DNA polymerase enzyme reads the DNA and adds complementary nucleotides using the rule of base pairings (A to T and C to G). It can only move from 5’ to 3’ direction
DNA is unwound and replication proceeds in both directions, forming two replication forks
Replication fork: Y-shaped region at the end of the strand where DNA unwinds
Replication bubble: an unwound and open region of a DNA helix where DNA replication occurs (2 forks facing each other)
At the replication fork, DNA opens up like a zipper in one direction
The leading strand grows at the 3’ end as the fork opens
In the lagging strand, the exposed 3’ end gets farther from the fork, and an unreplicated gap forms. Synthesis of the lagging strand occurs in small, discontinuous stretches called Okazaki fragments