Karbohidratet ne Ushqim (Ligjerata 1).docx

BSc Shkencat e Ushqimit dhe Teknologji
KURSI: KIMIA DHE BIOKIMIA E USHQIMIT
KAPITULLI: KARBOHIDRATET NË USHQIM (LIGJERATA 1)
QËLLIMI I LIGJERATËS:
Qëllimi i kësaj ligjërate është njohja me ndërtimin e karbohidrateve, vetitë fiziko-kimike të tyre, rolin dhe funksionin, metabolizmin, llojet kryesore dhe rëndësinë e tyre ne produkte te ndryshme ushqimore, si dhe punën praktike (laboratorike) që ndërlidhet me karbohidratet në ushqim.
KARBOHIDRATET
Karbohidratet jane komponime të ndërtuara prej C, H dhe O.Termi karbohidrat tregon se komponimet e këtij grupi mund të paraqiten si hidrate të karbonit me formulë të përgjithshme Cn (H2O) n. Për shkak të marrëdhënieve ndërmjet numrit të atomeve të hidrogjenit dhe numrit të atomeve te oksigjenit qe janë si te uji (2:1), shkencëtari rus K.Shmid në vitin 1844 i quajti këto komponime si karbohidrate, sepse në çdo atom karboni vjen një molekulë ujë.
Karbohidratet krijohen në bimët e gjelbra prej komponimeve të thjeshta siç janë CO2 dhe H2O duke shfrytëzuar direkt energjine e rrezeve të diellit. Kjo biosintezë quhet fotosinteza.
Fotosinteza është proces i cili paraqitet tek disa lloje të baktereve, algave dhe në gjethe të bimëve, e shumë rrallë edhe në rrënjët e bimëve të larta.
Fotosinteza si proces mund të ndahet në tri faza:
kapja e energjise se rrezeve te diellit;
përdorimi i energjisë për të formuar ATP dhe energjinë e reduktuar në formë të NADPH;
përdorimi i ATP dhe NADPH për ta fuqizuar sintezën e molekulave organike prej CO2 në ajër (fiksimi i karbonit).
Në bimë procesi i fotosintezës ndodh brenda kloroplasteve, organele qelizore këto të gjetura në disa qeliza. Kloroplastet sigurojnë energji dhe karbon të reduktuar të nevojshëm për rritjen dhe zhvillimin e bimëve, ndërsa bima siguron kloroplastin me CO2, ujë, azot, molekula organike dhe minerale të nevojshme për biogjenezën e kloroplastit. Shumica e kloroplasteve janë të vendosura në qeliza të specializuara të gjetheve, të cilat shpesh përmbajnë 50 ose më shumë kloroplaste për qelizë. Uji kalon lirshëm përmes mbështjellësve të membranave, ashtu si edhe molekulat e tjera neutrale të vogla si CO2 dhe O2.
Brenda kloroplastit është një sistem i komplikuar i membranave, i njohur si membrana fotosintetike (ose membrana tilakoide), që përmban shumicën e proteinave që kërkohen për reaksionet e dritës. Proteinat e nevojshme për fiksimin dhe reduktimin e CO2 ndodhen jashtë membranës fotosintetike, në fazën ujore rrethuese. Fotoliza e ujit është e drejtuar kryesisht nga drita e dukshme (gjatesia valore nga 400 në 700 nm) që absorbohet nga molekulat e pigmentit (kryesisht klorofili a dhe b dhe karotenoidet).
Dy reaksionet e para kryhen nën prezencen e dritës dhe zakonisht njihen si reaksione të dritës (ang. light dependent reactions).
Ashtu siç nënkuptohet edhe nga emri, reaksionet që varen nga drita kërkojnë rrezet e diellit. Në reaksionet që varen nga drita, energjia nga rrezet e diellit përthithet nga klorofili dhe shndërrohet në energji kimike të ruajtur, në formën e molekulës së bartësit të elektroneve NADPH dhe molekulës së bartësit të energjinë ATP (adenozin-trifosfatit). Reaksionet që varen nga drita zhvillohen në membranat tilakoide në grana (pirgun e tilakoideve), brenda kloroplastit.
Procesi që shndërron energjinë e dritës në energji kimike zhvillohet në një kompleks multi-proteinik të quajtur fotosistem. Dy lloje të fotosistemeve janë të fiksuara në membranën tilakoide: fotosistemi II (PSII) dhe fotosistemi I (PSI). Do fotosisteme luajnë një rol kryesor në kapjen e energjisë nga rrezet e diellit nga elektronet e eksituara. Këto elektrone me energji transportohen nga molekula "transportuese të energjisë", të cilat fuqizojnë reaksionet e pavarura nga drita.
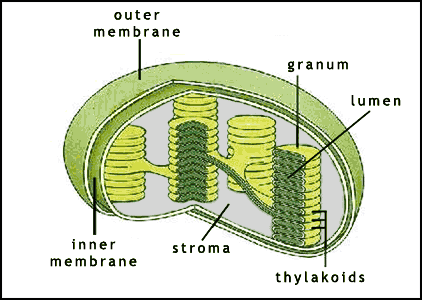
Fig.1. Struktura e kloroplastit
Reaksionet e pavarura nga drita (light independent reactions): reaksione kimike gjatë fotosintezës që shndërrojnë dioksidin e karbonit dhe komponimet e tjera në glukozë, të cilat zhvillohen në stromën e kloroplastit. Rubisco: (ribulozë - 1.5. bisfosfat-karboksilazë/oksigjenazë) një enzimë bimore e cila katalizon fiksimin e dioksidit të karbonit atmosferik gjatë fotosintezës duke katalizuar reaksionin midis dioksidi të karbonit dhe RuBP (ribulozë-1.5-difosfatit).
Krijimi i molekulave organike prej CO2 atmosferik, njihet edhe si Cikli i Kalvinit.
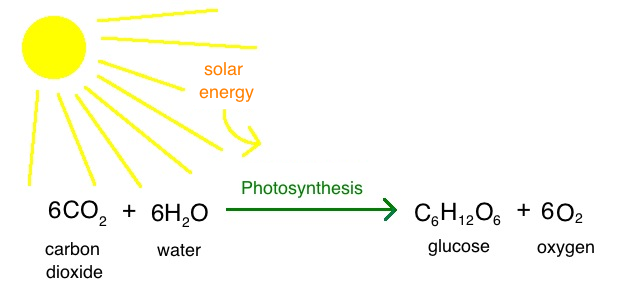
Fig.2. Reaksioni i përgjithshëm i procesit të fotosintezës
Roli i karbohidrateve është i shumëfishtë;
shërbejne si burim kryesor i energjisë për qelizën dhe organizmin në tërësi,
shërbejnë si material rezerv (amidoni dhe glikogjeni),
shërbejnë si material skeletor (celuloza, kitina etj),
shërbejnë si material për krijimin e komponimeve të ndryshme kimike, etj.
Karbohidratet klasifikohen sipas produkteve të hidrolizës në katër grupe kryesore si më poshtë:
I-Monosaharidet: përmbajnë një njësi monosaharide.
II-Disaharidet: përmbajnë 2 njësi monosaharide për molekulë.
III-Oligosaharidet: përmbajnë nga 3 deri në 10 njësi monosaharide për molekulë.
IV-Polisaharidet: përmbajnë më shumë se 10 njësi monosaharide për molekulë.
MONOSAHARIDET
Monosaharidet janë sheqerna të thjeshta të cilat me hidrolizë nuk mund të zbërthehen në sheqerna më të thjeshta. Për nga natyra kimike janë aldo ose keto derivate të alkooleve polihidroksile. Klasifikimi dhe nomenklatura e monosaharideve bazohet kryesisht në tri karakteristika kryesore:
Numri i atomeve të karbonit që ndodhen në molekulën e monosaharideve, dhe se këto molekula emërtohen me shkronja greke duke shtuar prapashtesën “ozë” P.sh, molekulat e monosaharideve me pesë atome të karbonit njihen si pentoza.
Monosaharidet në bazë të grupit të tyre funksional aldehid ose keton ndahen në dy grupe të mëdha: aldoza dhe ketoza.
Klasifikimi i tyre bazohet gjithashtu edhe në konfiguracionin molekular, pra vendosjen në hapësirë (stereopozicioni) të grupeve atomike asimetrike të karbonit.
Monosaharidi më i thjeshtë është trioza, aldehid ose keton 3 karbonik. Gliceraldehidi është një aldotriozë; dihidroksiacetoni është një ketotriozë. Zgjatja e njëpasnjëshme e zinxhirit të triozave jep tetroza, pentozat, heksozat, heptozat dhe oktozat. Në serinë aldo kemi thënë që këto quhen aldotrioza, aldotetroza, aldopentoza, aldoheksoza, etj., në keto-seri, ato janë ketotrioza, ketotetroza, ketopentoza, ketoheksoza, etj.
Brenda një serie të caktuar me të njëjtin numër të atomeve të karbonit, këto molekula ndryshojnë vetëm në rregullimin e atomit të hidrogjenit dhe grupit hidroksil rreth atomeve të karbonit. Krahasoni, për shembull, glukozën dhe manozën - konfigurimi i grupit të hidrogjenit dhe hidroksilit për secilin prej karbonëve është i njëjtë përveç karbonit numër dy. Në glukozë, grupi hidroksil është në të djathtë; në manozë, është majtas. Dy sheqerna që ndryshojnë vetëm në konfigurimin rreth një atomi karboni referohen si epimerë.
Konfigurimi dhe konformimi janë dy terma të përdorur shpesh që ngatërrohen lehtësisht. Konfigurimi është rregullimi në hapësirë i atomeve ose grupeve të atomeve të një molekule që mund të ndryshohet vetëm duke prishur dhe duke bërë lidhje. Konformimi është rregullimi në hapësirë i atomeve ose grupeve të atomeve të një molekule që mund të lindin nga rrotullimi rreth një lidhjeje të vetme dhe që janë të afta të ekzistojnë të fundme.
Monosaharidet e zakonshëm janë përbërje të bardha kristalore, që treten lirshëm në ujë, por të patretshme në tretës polar të tillë si benzeni dhe heksani. Shumica e tyre kanë një shije të ëmbël dhe janë nga më të bollshmet e sheqernave të thjeshta.
Të gjithë monosaharidet, përveç dihidroacetonit, përmbajnë të paktën një atom asimetrik karboni, i cili, në kuptimin më të thjeshtë, është ai për të cilin janë bashkuar katër përbërës të ndryshëm. Molekulat që posedojnë një qendër asimetrike mund të ekzistojnë në forma stereoizomerike.
Stereoizomerët kanë të njëjtën strukturë por ndryshojnë në rregullimin hapësinor të grupeve të ndryshme zëvendësuese. Stereoizomerët përfshijnë forma cis dhe trans dhe epimerë.
Grupi hidroksil dhe atomi i hidrogjenit në pozicionet e kundërta të karbonit asimetrik; këto dy përbërës janë me pamje të pasqyrës ndaj njëra-tjetrës. Format e imazhit pasqyrues të së njëjtës përbërje quhen enantiomerë.
Në natyrë, D-monosaharidet janë shumë më të bollshme se L-monosaharidet. Shumica e qelizave të gjitarëve kërkojnë sheqer D sepse nuk janë në gjendje të metabolizojnë formën L.
Ashtu si me aldehidet, reaksion shtesë në karbohidrate (reaksionet e formimit të gjysmëacetaleve, etj) prodhojnë një atom karboni asimetrik; referohet si karboni anomerik. Tani kemi dy monosaharide që ndryshojnë vetëm në konfigurimin e tyre në lidhje me karbonin anomerik; quhen anomere dhe referohen si forma α dhe β. Në formën α, grupi hidroksil është nën planin e unazës në të cilën është bashkangjitur. Në forma β, është sipër.
Përfaqësuesit më të rëndësishëm në kuadër të monosaharideve janë grupi i heksozave (sheqerna të cilat në strukturën e tyre kanë 6 atome te karbonit), siç janë: glukoza, fruktoza, galaktoza, etj.
D-Glukoza (sheqeri i rrushit ose dekstroza) është monosaharidi më i përhapur në natyrë. Glukoza hyn në përbërje të shumë saharideve tjera, siç janë p.sh. saharoza, amidoni, glikogjeni, celuloza, etj. Për nga ndërtimi kimik (fig.2.) glukoza është një aldoheksozë dhe sheqer reduktues, e cila paraqitet me këtë strukturë:
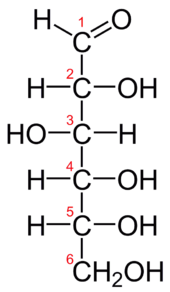
Fig.5. D-Glukoza
Glukoza me reduktim kalon në alkoolin gjashtëhidroksil – sorbitolin (glukitolin) nën ndikimin e enzimit aldehid reduktazë, e cila enzim grupin aldehid të glukozës e metabolizon (konverton) në grup hidroksil.
Me oksidim glukoza kalon në acidin e njohur si acidi glukonik, ndërsa me oksidim pak më të fortë dhe nën ndikimin e HCl ajo kalon në acidin e njohur si acidi glukarik.
Nën ndikimin e acidit të nxehtë hidrojodik nga molekula e glukozës largohen grupet funksionale te O2, me ç’rast formohet heksani (fig.5.).
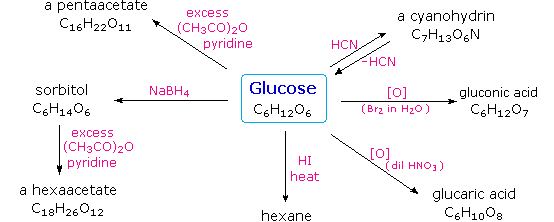
Fig.6. Produktet e ndryshme të glukozës
D-Glukoza është një polialkool dhe një aldehid. Është klasifikuar si një aldozë, një përcaktim për sheqernat që përmbajnë një grup aldehid. Fundi -ozë nënkupton një sheqer; dhe-nënkupton një grup aldehidi. Kur D-glukoza është shkruar në një mënyrë të hapur ose vertikale të zinxhirit të drejtpërdrejtë, i njohur si një strukturë aciklike në kiminë organike, me grupin aldehid (pozicioni 1) në krye dhe grupin primar hidroksil (pozicioni 6) në pjesën e poshtme, shihet se të gjitha grupet sekondare hidroksile janë në atome karboni që kanë katër zëvendësues të ndryshëm të lidhur me to.
Glukoza ka katër atome karboni kirale: C-2, C-3, C-4 dhe C-5. Glukoza e natyrshme ndodh si formë D, në veçanti D-glukoza. Ajo ka një imazh të pasqyrës molekulare të njohur si forma L, veçanërisht L-glukoza.
D-Fruktoza është lloj tjetër i monosaharidit e cila është shumë e përhapur në natyrë dhe në nektarin e luleve. Fruktoza me shumicë ndodhet edhe në mjaltë. Ka shije më të ëmbël sesa glukoza. Fruktoza përfitohet gjatë hidrolizës së saharozës. Në organizmin e njeriut dhe të kafshëve kryesisht fitohet si rezultat i hidrolizës së saharozës ose edhe gjatë metabolizmit të glukozës. D-Fruktoza ka vetëm tre atome karboni kirale, C-3, C-4 dhe C-5. Kështu, ka vetëm 2³ ose 8 D-ketoheksoza. D-Fruktoza është ketoza kryesore tregtare dhe e vetmja e gjetur e lirë në ushqime natyrale, por, si D-glukoza, vetëm në sasi të vogla. Fruktoza për nga ndërtimi kimik është një ketoheksozë ose polihidroksiketon 6 - karbonik (fig.6.) e cila paraqitet me këtë formulë strukturale:
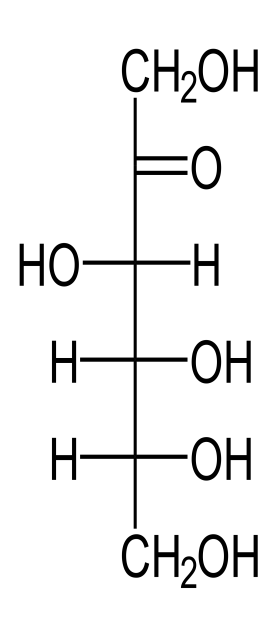
Fig.7. D-Fruktoza
D-Galaktoza (α-D-Galaktopiranoza) është monosaharid (sheqer reduktues) e cila në botën bimore kryesisht ndodhet në përbërjen e polisaharideve të njohura si galaktomananet, pastaj hyjnë në përbërjen edhe të polisaharideve bimore, siç janë hemiceluloza dhe agar-agari, etj. Tek njeriu dhe organizmat tjerë shtazorë galaktoza hynë në ndërtimin e lipideve të përbëra siç janë: cerebrozidet dhe gangliozidet (lipide që hynë në përbërjen e sistemit nervor), pastaj galaktoza është pjesë përbërëse e disaharidit laktozë, etj.
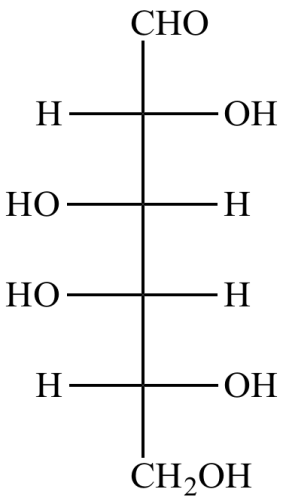
Fig.8. D-Galaktoza
Derivatet e monosaharideve
1-Acidet e sheqerit:
a-Acidet aldonike:
Grupi aldehid i aldozave oksidohet për të formuar acidin aldonik përkatës.
-Glukoza oksidohet për të formuar acidin glukonik
-Galaktoza oksidohet për të formuar acidin galaktonik.
b-Acidet uronike:
Grupi primar i alkoolit i monosaharideve oksidohet për të formuar acidin uronik përkatës.
-Glukoza oksidohet për të formuar acidin glukuronik (GlcUA).
-Galaktoza oksidohet për të formuar acidin galakturonik (GalUA).
c-Acidet aldarike:
Këto janë monosakaride në të cilat si grupi aldehid ashtu edhe ai primar i alkoolit oksidohen për të formuar acidin aldarik përkatës p.sh. glukoza jep acidin glukarik.
d-L-Acidi askorbik (vitaminë C):
Ajo formohet te disa kafshë (jo te njerëzit) nga glukoza. Konsiderohet si një acid sheqeri.
H – C = O
COOH
COOH
O = C
H – C – OH
H – C – OH
H – C – OH
HO – C
O
HO – C – H
HO – C – H
HO – C – H
HO – C
H – C – OH
H – C – OH
H – C – OH
H – C
H – C – OH
H – C – OH
H – C – OH
HO – C – H
COOH
D-Glucuronic
CH2OH
D-Gluconic acid
COOH
D-Glucaric acid
CH2OH
L-Ascorbic acid
Fig.5. Acidet e sheqerit
2-Alkoolet e sheqerit:
Këto janë sheqerna në të cilët grupi karbonil reduktohet në grupin e alkoolit. Sorbitoli është alkooli i glukozës, dulcitoli është alkooli i galaktozës dhe manitoli është alkooli i manozës.
Reduktimi i ketoneve prodhon 2 alkoole epimerike, p.sh. reduktimi i fruktozë jep sorbitolin dhe manitolin.
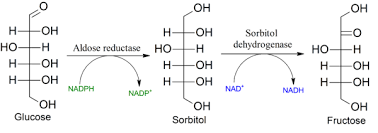
Fig.6. Shembull i alkoolit të sheqerit
3-Deoksi sheqerna:
Këto janë sheqerna në të cilat grupi hidroksil zëvendësohet nga një atom hidrogjeni. Shembujt më të rëndësishëm janë:
2-deoksi ribozë:
Është i pranishëm në strukturën e ADN-së.
L-Fukozë (6-deoksi-L-galaktozë):
Është i pranishëm në disa glikoproteina të membranës qelizore dhe antigjene të grupit të gjakut.
4-Aminosheqernat:
Këto janë sheqerna në të cilat një grup amin (NH2) zëvendëson grupin hidroksil në karbonin e dytë p.sh. glukozamina (GluN), galaktozamina (GlaN) dhe manozamina (ManN)
Aminosheqernat janë përbërës të rëndësishëm të glukozaminoglukaneve (GAG) dhe të disa llojeve të glikolipideve dhe glikoproteinave.
Disa antibiotikë përmbajnë aminosheqerna të cilët janë të rëndësishëm për aktivitetin e tyre.
5-Acidet aminosheqerore:
Këto formohen nga shtimi i acideve në aminosheqerna. Shtimi i acidit piruvik në manozaminë jep acid neuraminik. Derivatet N-acetile të acideve të aminosheqerit quhen acide sialike p.sh. Acidi N-acetil neuraminik (NANA). NANA hyn në strukturën e glikolipideve dhe glikoproteinave.
6-Formimi i esterit:
Grupet hidroksile të monosaharideve mund të formojnë estere me acide
a-Esteret e fosfatit: Për shembull glukoza 1-P dhe glukoza 6-P.
b-Esteret sulfate : Janë të pranishëm në disa lloje të polisaharideve dhe glikolipideve (sulfolipide) p.sh. β-D- galaktozë 3-sulfat.
7-Glikozidet:
Glikozidet janë produkte të kondensimit të karbonit anomerik të sheqerit me:
a) Një sheqer tjetër (glikon): p.sh. formimi i disaharideve dhe polisaharideve. Ose
b) Përbërja jo-karbohidrate (aglikon): të tilla si alkoolet, fenolet ose bazat azotike.
Lidhja glikozidike emërtohet sipas karbonit anomerik me të cilin është ngjitur (α & β) dhe sipas sheqerit mëmë p.sh. lidhje glukozidike, galaktozide ose fruktozidike.
Shembuj të glikozideve:
a) Nukleozidet janë glikozide të formuara nga riboza ose deoksiriboza dhe një bazë azotike që gjendet në acidet nukleike.
b) Glikozidet kardiake: përmbajnë përbërës steroid si aglikon në kombinim me molekulat e sheqerit
DISAHARIDET
Disaharidet janë sheqerna të cilat janë të ndërtuara prej 2 molekula të monosaharideve. Ato gjatë hidrolizës zbërthehen në monosaharide prej të cilave edhe janë të ndërtuara. Në kuadër të oligosaharideve si përfaqësues më të rëndësishëm janë ato të cilat bëjnë pjesë në grupin e disaharideve (saharoza, laktoza, maltoza). Vetëm disa oligosaharide ndodhin në natyrë. Shumica prodhohen nga hidroliza e polisaharideve në njësi më të vogla.
Saharoza (C12H22O12) kryesisht njihet si sheqeri i tavolinës dhe paraqet disaharidin më të rëndësishëm për ushqimin e njeriut. Është disaharid shumë i përhapur ne botën bimore dhe gjendet në: fara, fruta, kërcej, gjethe dhe rrënjë të shumë bimëve. Ky disaharid nuk ndodhet në botën shtazore. Në aspektin kimik saharoza (fig.7) është një glukozid: α-D-Glukopiranozil-(1→2)-β-D-fruktofuranozë dhe paraqitet me këtë strukturë të përgjithshme:
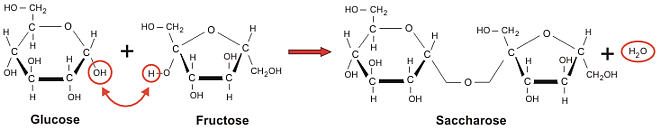
Fig.7. Struktura kimike e disaharidit saharozë (glukoza + fruktoza)
Në kimi lidhja glikozide paraqet një lloj të lidhjes kovalente ndërmjet një molekule të karbohidratit me ndonjë grup tjetër i cili mundet me qenë karbohidrat ose jo.
Disaharidi saharozë nuk aftësi reduktuese dhe si pasojë nuk krijon ozacone. Është substancë kristalore me ngjyrë të bardhë, tretet mirë dhe shpejtë në ujë dhe hidroliza e saj mund të bëhet me acide, baza dhe enzime.
Maltoza (C12H22O11) është disaharid që ndryshe njihet edhe si sheqeri i mjaltit (gjendet ne sasi të mëdha në mjaltë). Në aspektin kimik paraqet një α-D-Glukopiranozil-(1→4)-α-D-glukopiranozë. Pra përbëhet prej dy njësive të glukozës të lidhura me lidhje α-(1,4) glikozidike. Maltoza në organizmin e njeriut fitohet nga hidroliza e amidonit dhe glikogjenit në traktin digjestiv.
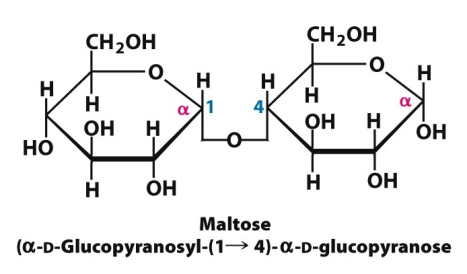
Fig.8. Struktura kimike e disaharidit maltozë (glukoza + glukoza)
Laktoza (C12H22O11) që ndryshe njihet si sheqeri i qumështit (përbën 2-8% të qumështit) është disaharid shumë i rëndësishëm i cili gjendet në qumështin e gjitarëve dhe është produkt i gjëndrave qumështore. Në aspektin kimik paraqet O-β-D-Galaktopiranozil- (1→4) – α-D-glukopiranozë. Është substancë kristalore me shije të dobët të ëmbël. Ka aftësi reduktuese dhe formon ozacone (laktozaconet). Me hidrolizë disaharidi laktozë jep një molekulë D-glukozë dhe nje molekulë D-galaktozë. Qumështi i lopës përmban laktozë 4.1-5.4./100 ndërsa në qumështin e nënës sasia e laktozës sillet prej 6.7-7.8/100.
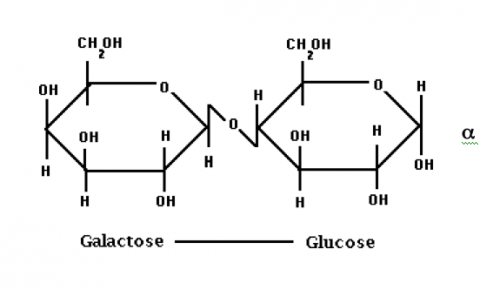
Fig.9. Struktura kimike e disaharidit laktozë (galaktoza + glukoza)
OLIGOSAHARIDET
Këto janë polimere me 3-10 njësi monosaharide. Ato nuk janë një burim i rëndësishëm i karbohidrateve në dietë për njerëzit, pasi shumica e tyre nuk janë të tretshëm. Ato janë të pranishme si përbërës i shumë llojeve të glikolipideve dhe glikoproteinave p.sh. oligosaharidet që janë përbërës të substancës së grupit të gjakut ABO, imunoglobulinave dhe glikolipideve dhe glikoproteinave të membranave qelizore
POLISAHARIDET
Polisaharidet janë karbohidrate të cilat janë të ndërtuara prej më shumë se gjashtë molekulave të monosaharideve. Në ndërtimin e tyre më së shpeshti marrin pjesë monosaharidet: glukoza, fruktoza, ksiloza dhe arabinoza. Polisaharidet në ujë zakonisht nuk treten ose japin tretje viskoze koloidale. Përfaqësues më të rëndësishëm në kuadër të grupit të polisaharideve janë: amidoni dhe celuloza (bota bimore) dhe glikogjeni (bota shtazore).
Amidoni (Amyllum) është karbohidrati më i përhapur në botën bimore dhe atë kryesisht tek bimet e larta. Është polimer kompleks i cili është produkt i procesit të fotosintezës. Përbëhet prej një numri të madh të molekulave të glukozës të lidhura me lidhje glikozide C1-C4 dhe C1-C6. Tek bimët kryesisht shërben si material rezerv ndërsa tek njeriu dhe organizmat tjerë shtazor shërben si substancë ushqyese. Amidoni është i përbërë prej dy polisaharideve:
amilozës, e cila tretet ne ujë, përbëhet prej një numri të madh të mbetjeve të glukozës (250-300) të cilat në mes veti janë të lidhura me lidhje glikozide C1-C4. Me tretje lugoli jep ngjyrë të kaltër kurse molekula e saj nuk është e degëzuar;
amilopektinës, e cila përbëhet poashtu nga molekulat e glukozës të cilat këtu përveç lidhjeve C1-C4 posedojnë edhe lidhjet C1-C6. Me tretje lugoli jep ngjyrë vjollce ose të kuqe dhe molekula e saj është e degëzuar.
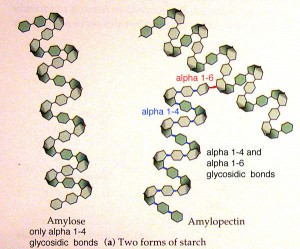
Fig.10. Ndërtimi i polisaharidit amidon (amiloza dhe amilopektina)
Glikogjeni është polisaharid shumë i ngjashëm me amidonin dhe shumë shpesh quhet edhe si amidoni shtazor. Është polimer i cili ndërtohet nga një numër shumë i madh i molekulave të glukozës (24.000-25.000). Shumica e mbetjeve të glukozës tek makromolekula e glikogjenit janë të lidhura me lidhje C1-C4 ndërsa degëzimet me lidhje C1-C6 glikozide. Dy vende (organe) kryesore të deponimit të energjisë në formë të glikogjenit në organizmin tonë janë: mëlçia dhe muskujt. Ekziston përqendrim më i madh i glikogjenit në mëlçi sesa në muskuj, mirëpo për shkak të masës së përgjithshme të indit muskulor, në muskuj deponohet sasi më e madhe e glikogjenit krahasuar me mëlçinë. Në qelizë (citosol) glikogjeni paraqitet në formë të granulave të vogla me diametër prej 10-40 nm.

Fig.11. Granulat e polisaharidit glikogjen në indin muskulor (EM)
Celuloza është polisaharidi më i përhapur në botën bimore. Është polimer i cili përbëhet prej 3000 e më shumë njësive të glukozës. Është substancë bazë e botës bimore dhe komponenta kryesore e murit qelizor. Pambuku përmban 90-95g/100. Gjatë hidrolizës së gjatë me acide minerale celuloza jep glukozën. Për herë të parë është izoluar në vitin 1834 nga shkencëtari francez Anselme Payen. Rezultatet e hulumtimeve shkencore (rentgenografike) kanë vërtetuar se polisaharidi celulozë është e ndërtuar nga vargjet e gjata dhe të drejta (jo të degëzuara). Cdo varg është i ndërtuar prej 40-50 njësi glikozide kurse 40-60 vargje të tilla e ndërtojnë një fije e cila quhet micele. Molekulat e celulozës në micele janë të lidhura me lidhje hidrogjenore. Trakti digjestiv i njeriut dhe i shtazëve nuk mund ta zberthejë celulozën mirëpo mikroorganizmat (bakteret intestinale) të cilat gjenden në traktin digjestiv e zbërthejnë atë nën veprimin e enzimit celulazë. Fibrat e celulozës tek njeriu i ndihmojnë peristaltikës së zorrëve dhe si pasojë edhe tretjen (digjestionin) në përgjithësi.
Njeriut i nevojiten çdo ditë nga 400/600 gr gluçide ose 5-8 gr/kg të peshës trupore.
METABOLIZMI I KARBOHIDRATEVE
Pas resorbimimit shumica e monosaharideve transformohen në glukozë andaj me metabolizëm të karbohidrateve shpesh nënkuptohet edhe metabolizmi i glukozës. Metabolizmi i sheqernave pas fosforilimit të tyre merr kahje në varësi nga nevojat energjetike të organizmit në drejtim të katabolizmit ose anabolizmit.
Metabolizmi i sheqernave ne organizmin e gjitareve përfshinë katabolizmin dhe anabolizmin e karbohidrateve.
Katabolizmi i karbohidrateve
Glikoliza (fig.12): Oksidimi i glukozës ose glikogjenit përmes rrugës së Embden-Meyerhof-Parnasit deri në piruvat ose laktat (acid piruvik ose laktik).
Glikoliza si proces katabolik kryhet në hapësirën ekstra-mitokondriale respektivisht në citoplazmën qelizore. Është reaksion në të cilin glukoza si heksozë në fund të reaksionit zbërthehet deri në dy molekula të piruvatit 3 karbonik me pjesëmarrjen e 10 enzimeve të ndryshme.
Reaksioni (procesi) i glikolizës përbëhet prej dy fazave: fazës I dhe fazës II.
Faza I e glikolizës zakonisht njihet edhe si faza përgatitore në të cilën përfshihen 5 nënfaza:
Fosforilimi i glukozës me anë të ATP nën veprimin e enzimit të njohur si heksokinazë ose glukokinazë me ç’rast fitohet një produkt i ndërmjetëm i njohur si glukozë - 6 – fosfat;
Glukozë-6-fosfati me ndihmën e enzimit fosfohekso – izomerazë metabolizohet në ndërproduktin tjetër i cili quhet fruktozë - 6-fosfat;
Fruktozë-6-fosfati fosforilohet me anë të ATP nën veprimin e enzimit fosfo-fruktokinazë1 me ç’rast fitohet fruktozë - 1.6 - difosfati;
Fruktozë - 1.6 - difosfati nën veprimin e enzimit aldolazë zbërthehet në dy trioza, në glicerinaldehid-3-fosfat (aldozë) dhe dihidroksiaceton- fosfat (ketozë).
Faza II e glikolizës zakonisht njihet edhe si faza e përfitimit ose konservimit të energjisë me ç’rast fitohen 4 molekula të ATP respektivisht 2, sepse dy molekula të ATP kanë shërbyer në fillim për inicimin e procesit të glikolizës. Këtu përfshihen këto nënfaza:
Shndërrimi (izomerizimi) i dihidroksiaceton - fosfat (ketozë) në glicerinaldehid-3-fosfat nën ndikimin e enzimit fosfotriozë-izomerazë;
Nën veprimin e enzimit glicerinaldehid-3-fosfat dehidrogjenazë glicerinaldehid-3-fosfati kalon në acidin 1.3. difosfoglicerinik me ç’rast formohet NADH2;
Acidi 1.3 - difosfoglicerinik nën veprimin e enzimit kinaza e acidit fosfoglicerinik kalon në acid 3-fosfoglicerinik me ç’rast fitohet molekula e ATP;
Acidi 3-fosfoglicerinik nën veprimin e enzimit mutaza e acidit 3-fosfoglicerinik kalon në acid 2-fosfoglicerinik;
Acidi 2-fosfoglicerinik nën veprimin e enzimit enolazë kalon në acidin fosfoenolpiruvik me ç’rast lirohet një molekulë ujë;
Acidi fosfoenolpiruvik nën veprimin e enzimit piruvat kinazë kalon në acidin piruvik me ç’rast lirohet (fitohet) molekula e ATP.
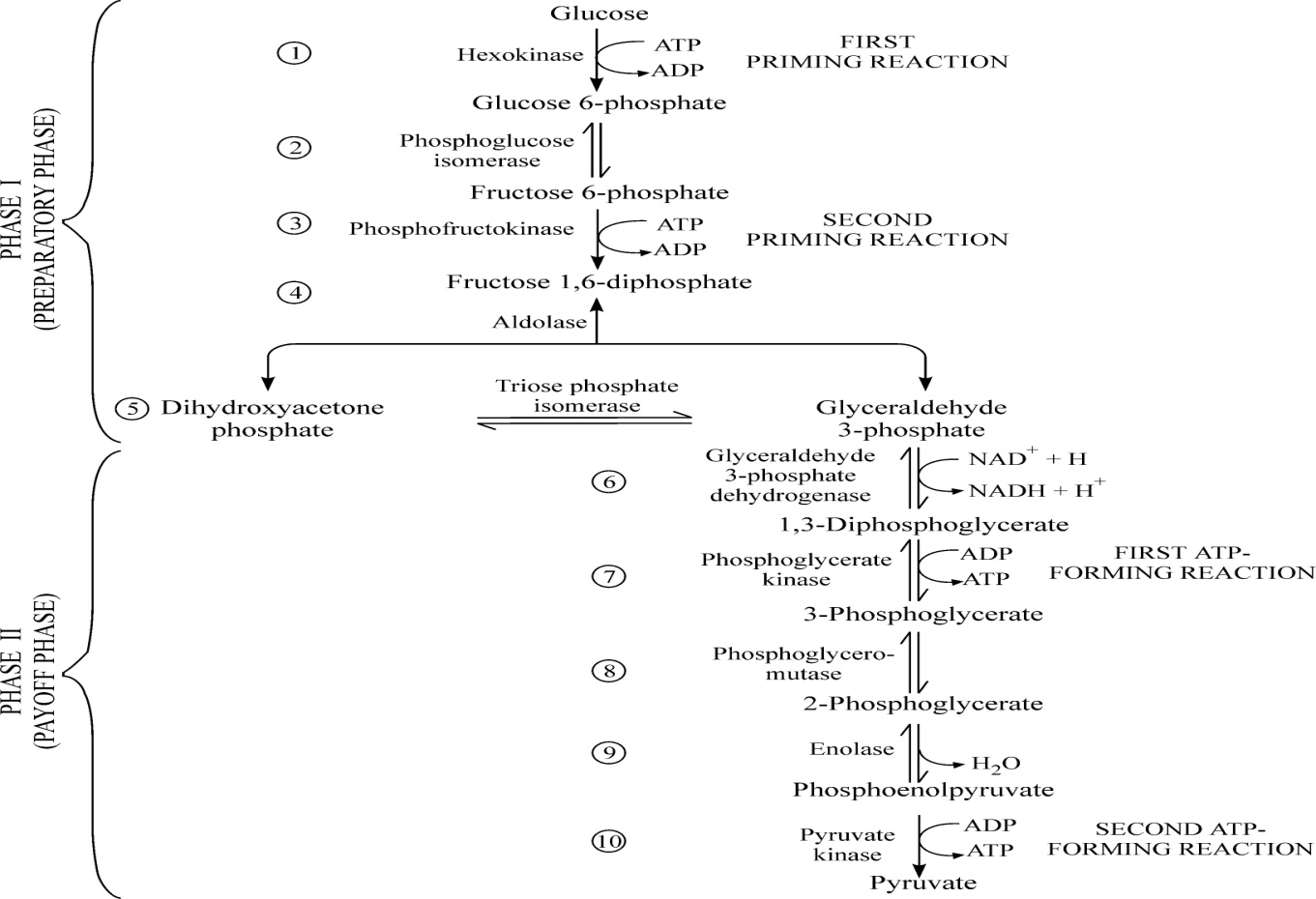
Te glikoliza oksigjeni molekular nuk është akceptor i hidrogjeneve dhe elektroneve, por rolin e akceptorit e luajnë substancat tjera.
Është vërtetuar se kur bëhet kontraktimi i muskujve në kushte anaerobe (në mungesë të oksigjenit) kemi zhdukjen e glikogjenit dhe krijimin e acidit piruvik dhe laktik si produkte kryesore finale. Kur furnizimi i muskulit me oksigjen është i mjaftueshëm atëherë lajmërohet glikogjeni e zhduket piruvati dhe laktati. Pra kur kontraktimet bëhen në kushte aerobe nuk kemi grumbullim të laktatit dhe acidi piruvik oksidohet më tutje në CO2 dhe H2O.
Gjatë glikolizës anaerobe ku si produkt i fundit është acidi laktik, fitohen gjithsej 2 molekula ATP për çdo molekulë të glukozës së zbërthyer, ndërsa gjatë glikolizës aerobe fitohen gjithsej 8 molekula të ATP.
Glikogjenoliza (fig.13): Zbërthimi i glikogjenit. Produkti final i glikogjenolizës në mëlçi dhe veshkë është glukoza ndërsa në muskuj është piruvati (acidi piruvik) dhe laktati (acidi laktik).
Zbërthimi i glikogjenit fillon nën veprimin e enzimit fosforilazë i cili enzim është specifik për fosforilizën e lidhjeve C1-C4 në molekulën e glikogjenit dhe lirimin e glukozë -1- fosfatit ndërsa në shkëputjen e lidhjeve C1-C6 te glikogjeni merr pjesë enzimi amilo - 1.6 -glukozidaza. Mandej glukozo-1-fosfati nën veprimin e enzimit fosfoglukomutazë transformohet në glukozë-6-fosfat, e pastaj rruga e zbërthimit të glukozë-6-fosfatit është e njëjtë me atë të glikolizës ndërsa në mëlçi (ang.liver) dhe veshkë (ang. kidney) nën veprimin e enzimit glukozë-6- fosfatazë lirohet glukoza e lirë.
Dy hormonet të cilat kontrollojnë procesin e glikogjenolizës janë glukagoni dhe adrenalina dhe të dy këto hormone janë me natyrë peptidike.
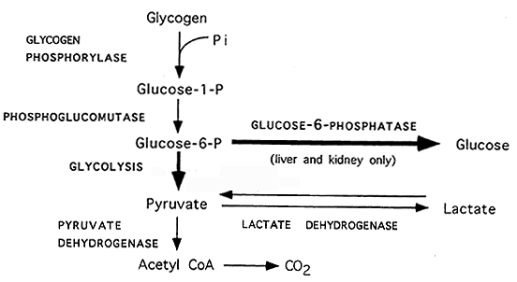
Fig.13. Glikogjenoliza
Cikli i Krebsit (fig.14) - Paraqet rrugën e zakonshme të oksidimit të sheqernave, yndyrnave dhe proteinave. Ky cikël i reaksioneve zhvillohet në prani të O2 dhe si i tillë cikli (procesi) është aerob.
Metabolizmi aerob (Cikli i Krebsit) kryhet në mitokondritë e qelizës dhe zhvillohet në tri etapa kryesore:
Krijimi i acetil-CoA prej acidit piruvik, acideve yndyrore dhe aminoacideve;
Oksidimi i acetil-CoA deri ne CO2 dhe H2O;
Transportimi i hidrogjeneve të liruara në hallka të ndryshme të Ciklit të Krebsit deri tek oksigjeni molekular. Kjo etapë është e përcjellur me fitimin (konservimin) e energjisë në formë të ATP dhe thirret fosforilimi oksidues.
Reaksioni (Cikli i Krebsit) fillon me oksidimin e acidit piruvik i cili poces tek eukariotët ndodh në matriksin mitokondrial me ç’rast fitohet acetil-CoA e cila pastaj futet në të ashtuquajturin reaksion të acideve trikarboksilike. Kalimi i acidit piruvik deri në acetil-CoA mundësohet nga një kompleks enzimatik i njohur si piruvat dehidrogjenazë, i cili përbëhet nga tri enzime të ndërlidhura të përbëra prej 60 subnjësive me ç’rast fitohet NADH2 dhe lirohet CO2.
Pas formimit të acetil-CoA 8 etapat kryesore të Ciklit të Krebsit janë:
Acetil-CoA reagon me acidin oksalacetik me ç’rast formohet acidi citrik dhe lirohet CoA nën veprimin e enzimit citrat-sintetazë;
Kalimi i citratit në izocitrat nën ndikimin e enzimit akonitazë;
Nën ndikimin e enzimit izocitrat-dehidrogjenazë acidi izocitrik oksidohet duke formuar fillimisht acidin oksalo-sukcinik (NADH2) e pastaj nën ndikimin e enzimit dekarboksilazë acidi oksalo-sukcinik kalon në intermedierin e njohur si acidi-α-ketoglutarik me ç’rast lirohet gjatë reaksionit CO2;
Acidit-α-ketoglutarik i bashkohet CoA dhe kalon në një intermedier jo stabil të njohur si sukcinil-CoA me ç’rast edhe këtu lirohet CO2 dhe formohet NADH2;
Sukcinil-CoA sintetaza mundëson kalimin e sukcinil-CoA në acid sukcinik me ç’rast largohet një molekulë e CoA;
Acidi sukcinik kalon në acid fumarik nën ndikimin e enzimit sukcinat – dehidrogjenazë me ç’rast formohet FADH2;
Acidi fumarik nën ndikimin e enzimit fumarazë dhe me bashkimin e një molekule ujë kalon në acid malik;
Acidi malik oksidohet nën veprimin e enzimit malat-dehidrogjenazë dhe kalon përsëri në acid-oksalacetik me ç’rast formohet NADH2.
Gjatë Ciklit të Krebsit lirohen (formohen) gjithsejt 38 molekula të ATP.
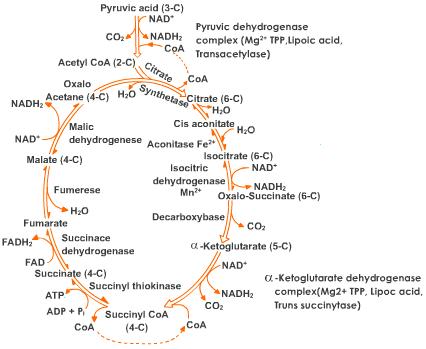
Fig.14. Cikli i Krebsit
Cikli pentozik (fosfopentozik): Është rrugë alternative e Glikolizës dhe Ciklit të Krebsit. Ndryshe njihet edhe si Cikli i Fosfopentozave. Cikli Fosfopentozik përbëhet prej dy fazave:
Faza oksiduese e cila përfshin dy hapa e në njërën prej të cilës fillimisht glukozë-6-fosfati oksidohet deri në lakton (6-fosfoglukonat) e pastaj me largimin e CO2 në hapin e dytë formohet ribulozë-5-fosfati;
Faza jo-oksiduese e cila karakterizohet kryesisht me krijimin e ribozë-5-fosfatit nga ribulozë-5-fosfati, molekulë kjo (ribozë-5-fosfatit) e cila hynë në ndërtimin e acideve nukleike.
Fermentimi alkoolik (fig.15): Rrushi paraqet substratin bazë ku me anë të procesit biokimik të njohur si fermentimi alkoolik, tharmet e gjinisë Sacharomyces metabolizojnë ose konvertojnë sheqerin prezent në pulpën e kokrrës së rrushit (glukoza dhe fruktoza) në alkool etilik dhe CO2, me ç’rast lirohet energjia në formë të nxehtësisë.
Konkretisht, procesi i fermentimit alkoolik përfshinë shndërrimin e një moli të glukozës në dy mole të alkoolit etilik (C2H5OH) dhe dy mole CO2, duke prodhuar dy mole ATP në proces. Acidi piruvik i formuar gjatë procesit të glikolizës nën ndikimin e enzimit piruvat dekarboksilazë shndërrohet në acetaldehid, i cili pastaj nën ndikimin e enzimit alkool dehidrogjenazë shndërrohet në alkool etilik, duke regjeneruar me këtë rast NAD+ (të oksiduar) të nevojshëm për procesin e glikolizës.
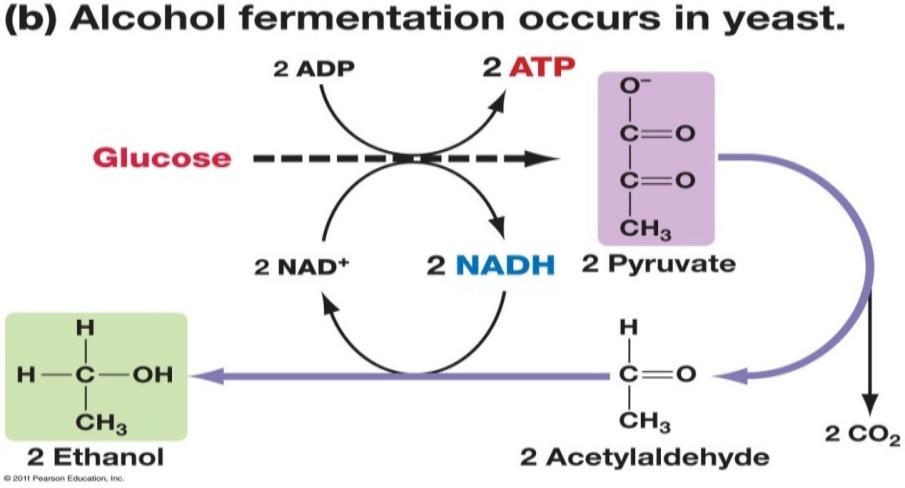
Fig.15. Reaksioni i fermentimit alkoolik tek tharmet (ang.Alcohol fermentation occurs in yeast)
PROCESET ANABOLIKE TE KARBOHIDRATEVE
Glikogjeneza (fig.16): Sinteza e glikogjenit prej glukozës.
Në procesin e glikogjenezës glukoza hyn si në proceset katabolike në formë të fosforiluar si glukozë-6-fosfat.
Glukoza shndërrohet në glukozë-6-fosfat nga veprimi i enzimeve glukokinazë ose heksokinazë me shndërrimin e ATP në ADP;
Glukoza-6-fosfati shndërrohet në glukozë-1-fosfat me veprimin e fosfoglukomutazës, duke kaluar nëpër glukozën e detyrueshme të ndërmjetme të njohur si glukozë -1,6-bisfosfat;
Glukoza-1-fosfat shndërrohet në UDP-glukozë nga veprimi i enzimës pirofosforilazë UDP-glukozë. Formohet pirofosfati, e cila më vonë hidrolizohet nga pirofosfataza në dy molekula fosfat;
Enzima glikogjenina është e nevojshme për të krijuar zinxhirë fillestarë të shkurtër të glikogjenit, të cilat më pas zgjaten dhe degëzohen nga enzimat e tjera të glikogjenezës. Glikogjenina, një homodimer, ka një mbetje tirozine në secilën njësi që shërben si spirancë për fundin reduktues të glikogjenit. Fillimisht, rreth shtatë molekula UDP-glukozë shtohen në secilën mbetje tirozine nga glikogjenina, duke formuar lidhje α (1 → 4);
Sapo të formohet një zinxhir prej shtatë monomerësh të glukozës, glikogjen-sintetaza lidhet me zinxhirin e glikogjenit në rritje dhe shton glukozën UDP në grupin 4-hidroksil të mbetjes glukozile në fundin joreduktues të zinxhirit të glikogjenit, duke formuar më shumë lidhje α (1 4) në proces;
Degët bëhen me anë të enzimës së degëzimit të glikogjenit (e njohur edhe si amilo-α (1: 4) → α (1:6) transglikozilaza), e cila transferon fundin e zinxhirit në një pjesë të mëparshme përmes lidhjes glikozidike α-1:6, duke formuar degë, të cilat më tej rriten duke shtuar më shumë njësi glikozidike α-1: 4.
Ky proces kryhet në mëlçi dhe muskuj.
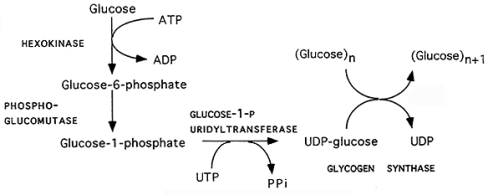
Fig.16. Glikogjeneza (sinteza e glikogjenit prej glukozës)
Glikoneogjeneza & Glikogjenogjeneza: Krijimi i glukozës ose glikogjenit prej substancave te cilat nuk bëjnë pjesë në karbohidrate (aminoacidet glikogjenike, acidi laktik, gliceroli).
Gjatë glikoneogjenezës e cila zakonisht paraqitet kur përqendrimi i sheqerit në gjak është i ulët dhe glikogjeni i mëlçisë është shpenzuar, 7 prej 10 reaksioneve të glikolizës janë reverzibile përjashto këtu reaksionet e katalizuara nga enzimi heksokinazë, PFK-1 (fosfofruktokinazë) dhe piruvat kinazë të cilat janë ireverzibile. Për këtë arsye reaksionet alternative të katalizuara nga enzime të ndryshme përdoren për t’i tejkaluar këto pengesa.
Një numër shumë i madh i substancave organike shërbejnë si substrat për krijimin e sheqernave (acidi laktik, piruvik, acetik, propionik, sukcinik, glicerina, dihidroksiacetoni, alanina, serina, valina, ac.glutaminik, ac. asparaginik, histidina, treonina, arginina).
Ky proces kryhet në mëlçi e jo në muskuj. Acidi laktik i krijuar në muskuj kalon në gjak e mandej në mëlçi përdoret për glukoneogjenezë. Pra acidi laktik mund të metabolizohet vetëm nëse kalon përsëri në acid piruvik, e mandej reaksionet për glukoneogjenezë siç e cekëm më lartë janë të njëjta (pothuajsë në tërësi) me ato të glikolizës, vetëm në kahje të kundërt.
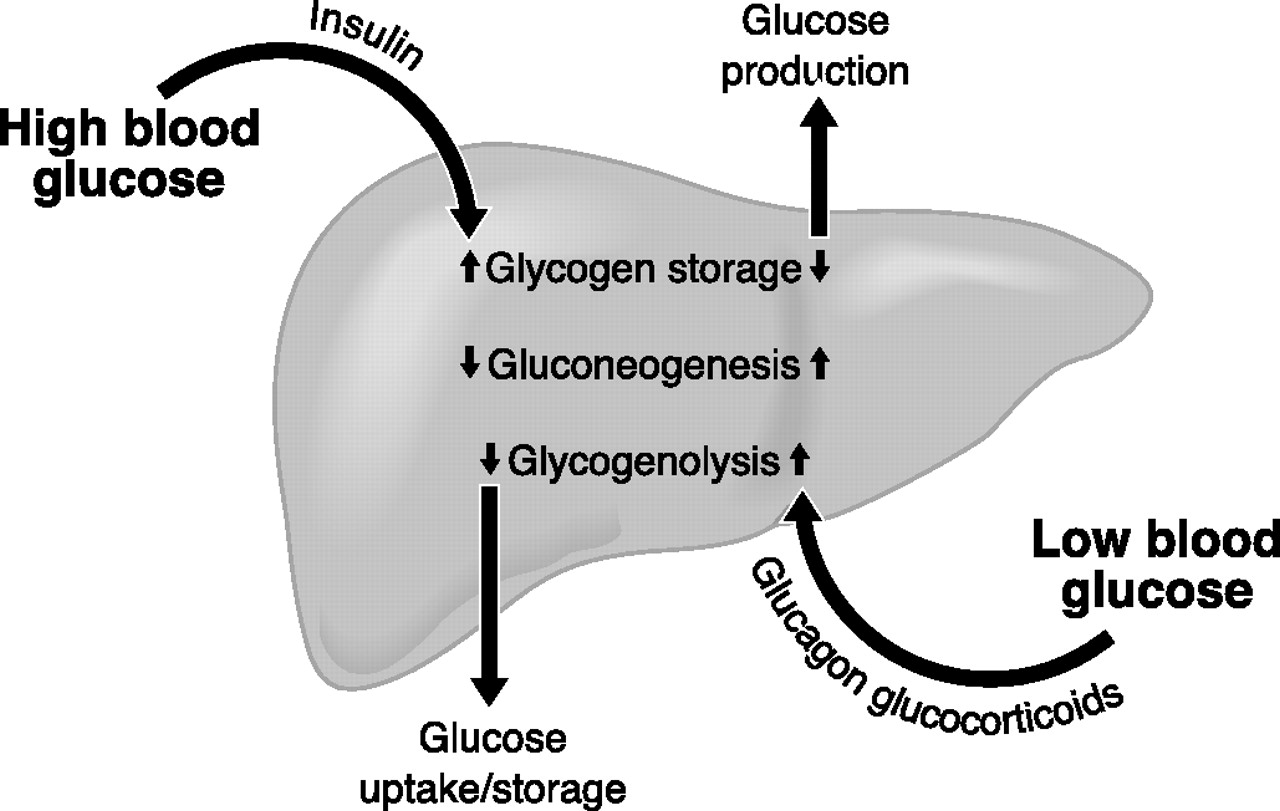
Fig.17. Efekti hiper dhe hipoglikemik i disa hormoneve (insulina, glukagoni, glukokortikosteroidet)
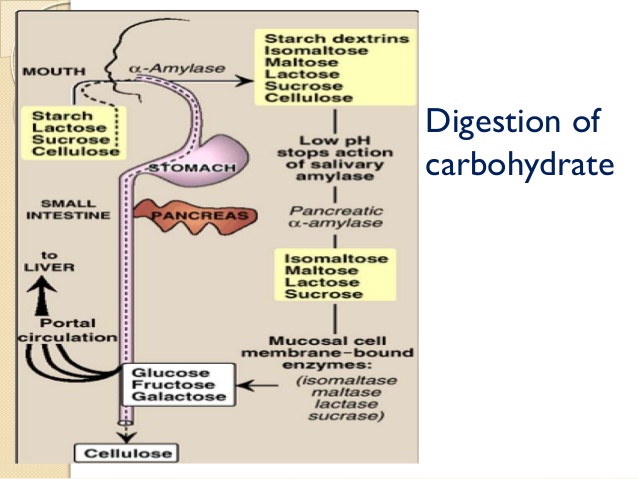
Fig.18. Skema e përgjithshme e metabolizmit (zbërthimit - tretjes) të karbohidrateve në traktin digjestiv
BIOKIMIA E KARBOHIDRATEVE TË USHQIMIT
Siç e cekëm edhe më lartë, karbohidratet mund të jenë komponime organike që përfshijnë prej monosaharideve të thjeshta e deri në struktura shumë komplekse siç janë polisaharidet.
Karbohidratet janë përbërës të rëndësishëm dhe të domosdoshëm të dietës njerëzore. Megjithëse ato kryesisht luajnë funksion energjetik me të cilën edhe karakterizohen, ato gjithashtu kanë edhe rol struktural dhe funksional. Sipas Autoritetit Evropian të Sigurisë Ushqimore, marrja e karbohidrateve duhet të shkojë në mes të 45 dhe 60 % të energjisë tek të rriturit dhe fëmijët më të mëdhenj (rritur) se një vit. Burimet dietike të sheqernave përfshijnë: pemët, frutat, perimet, qumështin dhe produktet e qumështit, si dhe ushqimet që përmbajnë saharozë të shtuar dhe hidrolizat të amidonit.
Polisaharidet në kombinim me proteina, lipide dhe acide nukleike luajnë rol të rëndësishëm në sistemet metabolike të shumë organizmave bimor dhe shtazor.
Karbohidratet kanë rol të shumëfishtë në sistemin (zingjirin) ushqimor, ku ato funksionojnë për të siguruar shijen, strukturën, dhe cilësinë e produkteve të ndryshme ushqimore, si dhe përfitimet ushqyese të konsumatorëve.
Sa i përket strukturës së tyre, dihet se karbohidratet përmbajnë numër të caktuar të grupeve hidroksile (OH) për molekulë, karakteristikë kjo që jep mundësi të mëdha për lidhje hidrogjenore, duke i bërë ato shumë hidrofilike. Kjo karakteristikë i lejon atyre të shërbejnë si mjet për kontrollin e lagështisë në produkte të ndryshme ushqimore.
Aftësia (afiniteti) e substancave për ta lidhur ujin njihet si lagështithithja (ang.humectancy), njëra ndër karakteristikat kryesore të karbohidrateve në ushqime.
Përveç prezencës së grupeve hidroksile (OH) aftësia për ta mbajtur lagështinë varet edhe prej karakteristikave të përgjithshme strukturore të karbohidrateve të caktuara, p.sh. fruktoza lidh më shumë ujë krahasuar me glukozën.
MONOSAHARIDET E USHQIMIT
Monosaharidet dihet se grupohen në dy familje (grupe): aldo-sheqernat dhe keto-sheqernat.
Sheqernat më të zakonshëm në kuadër të monosaharideve janë heksozat dhe pentozat. Prej pentozave më të zakonshme në ushqim janë D-ksiloza dhe L-arabinoza ndërsa prej heksozave më të rëndësishme janë D-glukoza, D-galaktoza, D-manoza dhe D-fruktoza. Prej këtyrë D-glukoza, D-galaktoza dhe D-manoza janë aldo-sheqerna.
Prej monosaharideve të gjetura tek bimët, vetëm glukoza dhe fruktoza janë gjetur të pakombinuara.
Glukoza gjendet në shumicën e frutave dhe në shumë perime. Është monomer i polimereve komplekse në kafshë (glikogjeni) dhe në bimë (amidoni). Shumica e glukozës është e pranishme në formë të një polimeri të patretshëm që njihet si celulozë.
Në aspektin komercial glukoza mund të prodhohet me degradimin e amidonit me anë të acideve të forta. Zakonisht përdoret acidi klorhidrik (HCl) i cili në kombinim me amidonin e përzier mirë dhe në temperaturë adekuate mundëson zbërthimin deri në glukozë. Pastaj acidi neutralizohet dhe produkti përfitohet me anë të filtrimit ose centrifugimit, në fund ai lahet (pastrohet) dhe thahet. Amidoni i lëngët mund të përdoret për të prodhuar shurup me vlera ekuivalente të dekstrozës prej 50 - 98.
Ekuivalenti i dekstrozës është një masë e sasisë së sheqernave reduktuese të pranishme në një produkt sheqeri të shprehur në përqindje në një bazë të thatë në krahasim me dekstrozën. Një maltodekstrinë me një DE prej 10 do të kishte 10% të fuqisë reduktuese të dekstrozës e cila ka një DE prej 100. Marrim edhe një shembull me disaharidin maltozë, një disaharid i bërë nga dy molekula glukozë (dekstrozë), e cila ka një DE prej 52, kjo pas korrigjimit për humbjen e ujit në peshë molekulare kur dy molekulat e glukozës janë të kombinuara. Glukoza (dekstrozë) ka një masë molekulare prej 180, ndërsa uji ka një masë molekulare prej 18. Për secilën 2 monomere të glukozës lidhëse, hiqet një molekulë uji. DE mund të llogaritet si 100 * (180 / masë molekulare (polimeri i glukozës). Në këtë shembull, DE llogaritet si 100 * (180 / (180 * 2 - 18 * 1) = 52. Këto zakonisht prodhohen duke përdorur enzimin e njohur si ekzoamilazë (glukan 1, 4 – α – glukozidazë), e njohur ndyshe edhe si amiloglukozidazë. Ky enzim mund ta shndërroj 95% - 98% të amidonit të lëngët në glukozë.
Glukoza mund të shndërrohet në fruktozë duka kaluar shurupi i glukozës nëpër kolonën e cila përmban enzimin glukozë – izomerazë. Ky hap shndërron afërsisht 40% të glukozës në fruktozë. Fruktoza koncentrohet duke kaluar përzierja nëpërmjet kolonës kromatografike. Glukoza dhe fruktoza kryesisht përdoren si shurupe në përbërësit ushqimor. Ato kanë aftësi të formimit të mbështjellësve dhe aftësi adhesive, dhe se përdoren si veshje (mbështjellës) të arrave të pjekura, karamelave, mëlmesave, etj. Shurupet e misrit me përqindje të lartë të fruktozës përdoren si ëmbëlsues dhe përdoren pothuasje çdo herë kur nevojitet procesi i ëmbëlsimit.
Fruktoza është sheqeri që karakterizohet me fuqinë e shijes më të ëmbël. Ajo absorbohet pasivisht, më ngadalë sesa glukoza. Ajo është e bollshme në fruta. Ekzistojnë polimerë të fruktozës, natyrale (inulina) të ekstraktuara në mënyrë industriale kryesisht nga rrënjët e bimës të njohur si çikorja (Cichorium intybus) dhe sintetike (fruktooligosaharidet), por këto komponime nuk kontribuojnë në mënyrë të konsiderueshme në ëmbëlsinë e ushqimeve dhe treten dobët, kështu që ato sillen edhe si fibra të tretshme.
Galaktoza përbën pjesë të glikolipideve dhe glikoproteinave të membranave qelizore, sidomos neuroneve. Sintetizohet nga gjëndrat e qumështit në gjitarë, për të prodhuar disaharidin laktozë, prandaj shumica e dietogalaktozës vjen nga konsumimi i laktozës në qumësht. Absorbohet nga zorrët së bashku me glukozën dhe me transportues të njëjtë.
OLIGOSAHARIDET E USHQIMIT
Oligosaharidet ashtu siç e theksuam edhe më lartë, përbëhen prej një numri të caktuar të monosaharideve (2-10).
Oligosaharidet e tretshme paraqesin burim të shpejtë të energjisë për organizmin e njeriut. Oligosaharidet jo të tretshme zakonisht kanë karakteristika prebiotike.
Prebiotikët janë përbërës ushqimor (substanca) të cilat në mënyrë selektive rrisin popullatën e baktereve të dëshiruara (Bifidobacterium dhe Lactobacillus) në zorrën e madhe.
Oligosaharidet si prebiotikë (p.sh. trans-galaktooligosaharidet) mund poashtu të përdoren për të stabilizuar probiotikët në sistemin ushqimor.
Probiotikët në ushqim përmbajnë kultura të mikroorganizmave të cilat përmisojnë balancën intestinale mikrobiale të konsumatorit. Janë baktere dhe tharme të promovuara si të shëndetshme për organizmin tonë. Ato zakonisht shtohen në jogurt ose merren si shtesa ushqimore (ang.food supplements) dhe zakonisht përshkruhen si baktere ose mikroorganizma “të mira” ose “miqësore”.

Fig.19. Ushqimet e pasura me prebiotikë
Karbohidrati më i rëndësishëm në kuadër të oligosaharideve padyshim që është disaharidi saharozë, e cila përbëhet prej njësive të α-D- glukozës dhe β-D-fruktozës të cilat janë të lidhura (1→2) nëpërmjet skajeve të tyre reduktuese. Saharoza formohet në citoplazmën e qelizave, në indet fotosintetikisht aktive dhe translokohet nëpërmjet floemës në vendet aktive metabolike. Saharoza hidrolizohet në zorrë nga veprimi i kompleksit të enzimës saharazë-izomaltazë. Është sheqer jo-reduktues i cili është shumë i tretshëm në ujë dhe kristalizohet lehtë. Gjithashtu përdoret për të ëmbëlsuar ushqimet, për të përmisuar kryesisht shijen e hidhur të shumë prej tyre, si dhe për t'i ruajtur ato duke rritur presionin osmotik i cili pamundëson rritjen e shumë mikroorganizmave të dëmshëm. Tek bimët siç janë kallami i sheqerit (Saccharum spp) dhe panxharsheqeri (Beta vulgaris) saharoza shërben si karbohidrat rezervë.
Në aspektin komercial saharoza përfitohet nga kallami i sheqerit dhe panxharsheqeri. Ekstraktimi nga panxharsheqeri fillon me pastrimin dhe prerjen e rrënjëve në rripa. Rripat pastaj barten në tanqe të mëdha ku sheqeri i papërpunuar ekstraktohet në ujë të nxehtë. Saharoza nga kallami i sheqerit fillon (ekstraktohet) me ndarjen dhe bluarjen e rrënjëve. Lëngu i shtrydhur del jashtë me anë të kalimit të rrënjëve të tokës nëpërmjet serisë së rrotullimeve. Pastaj lëngu i papërpunuar i sheqerit pastrohet duke përdorur përzierjen e gëlqeres dhe CO2 me qëllim përfitimin e CaCO3 (karbonati i kalciumit). Pastaj partikulat jo-sheqerore kapen për karbonatin e kalciumit dhe precipitojnë. Lëngu filtrohet dhe zihet nën vakum, i cili largon ujin dhe lejon shurupin e turbullt (të dendur). Ky material filtrohet dhe centrifugohet dhe pastaj pastrohet me ujë të nxehtë dhe në fund thahet.

Fig.20 Panxharsheqeri (Beta vulgaris)
Saharoza në ushqim mund të reagojë me përbërës të ndryshëm duke rritur në këtë mënyrë kohën e mbajtjes së aromave dhe shijes së produkteve të ndryshme ushqimore. Karakteristikat antioksidante janë përdorur me qëllim ndaljen e keqësimit të shijes së produkteve të ndryshme ushqimore të konservuara. Po ashtu përdoret edhe me qëllim pengimin e humbjes së lagështisë nga produktet e pjekura (pjekurinat). Saharoza mund të sigurojë ngjyrën e verdhë-kafe në produktet ushqimore nëpërmjet degradimit termal, degradimit alkalin dhe produkteve të reaksionit Mailard.
Maltoza dihet se është një sheqer reduktues i përbërë prej dy molekulave të glukozës të lidhura nga një lidhje glikozidike, e pranishme në disa fruta në të cilat përbën deri 15% të sheqernave totale. Është 50% më e ëmbël nëse krahasohet me saharozën. Ajo hidrolizohet në zorrë me anë të veprimit të enzimit maltazë. Maltoza është pjesë e maltodekstrinave dhe shurupeve të glukozës dhe përbën një përbërës të përdorur në shumë ushqime si burim i energjisë, duke u prodhuar industrialisht përmes hidrolizës së orizit ose edhe amidonit të misrit.
Laktoza dihet se është sheqeri i qumështit dhe formohet nga bashkimi i dy molekulave të galaktozës dhe glukozës të lidhura me lidhje glikozidike. Ajo hidrolizohet nga veprimi i enzimit laktazë, një enzimë aktiviteti i së cilës zvogëlohet (reduktohet) në çdo 2 ose 3 vjet në shumicën e njerëzve. Ajo nxirret, në formë të pastër, nga qumështi i lopës, karakterizohet me tretshmëri të ulët dhe ëmbëlsia e saj është vetëm 40% krahasuar me atë të saharozës. Tek foshnjat dhe fëmijët e vegjël laktoza nuk siguron vetëm energji, por gjithashtu ndihmon zhvillimin e mikrobiotës së zorrëve (bifidobaktereve dhe baktereve të tjera laktike), rritë biodiversitetin e kalciumit dhe elementeve të tjera minerale si dhe siguron galaktozën që është direkt i përdorshëm për zhvillimin e sistemit nervor.
Familja e Oligosaharideve Rafinozë (RFO) janë të përbëra nga saharoza dhe numër i ndryshëm i njësive të α-D-galaktozideve.
Rafinoza dhe stakioza janë dy sheqernat e zakonshëm të RFO të gjendura tek bimët. Rafinoza (trisaharid) përbëhet prej glukozës, fruktozës, dhe galaktozës. Stakioza (tetrasaharid) është e ngjashme me rafinozën mirëpo përmban njësinë (grupin) shtesë α-D-galaktozile. Rafinoza dhe stakioza gjenden në formë natyrale tek legumet (leguminozat). Ato mund të degradohen gjatë procesimit të ushqimit sepse janë karbohidrate termolabile. Melasa (ang.molasse) e panxharsheqerit përmban afërsisht 18% rafinozë.
Melasa paraqet produktin viskoz që rezulton nga rafinimi (pastrimi) i panxharsheqerit dhe kallamit të sheqerit në sheqer. Përveç sheqerit i cili dominon në këtë pëzierje (shurup) ekzistojnë edhe sasi në gjurmë të vitaminave dhe shumë lëndë minerale të rëndësishme të cilat sigurojnë shëndet të mirë për organizmin tonë. Melasa zakonisht përdoret si ëmbëlsues dhe aromatizues i ushqimeve të ndryshme. Ajo (melasa) gjithashtu paraqet komponentën kryesore të sheqerit komercial me ngjyrë kafe.
Oligosaharidet (karbohidratet) tjera me rëndësi janë edhe fruktanet (polimer të fruktozës). Fruktanët janë karbohidrate natyrale të pranishme në shumë ushqime, nga drithërat dhe bishtajoret deri te disa fruta dhe perime. Zakonisht këto oligosaharide përbëhen prej 25 mbetjeve të monosaharideve ose edhe më pak. Ekzistojnë pesë klasë të fruktaneve: inulina, levani, levani i përzier, polimerët e levanit dhe polimerët e inulinës. Bimët të cilët prodhojnë fruktane rriten kryesisht në zona ku thatësirat dhe acari janë zakonisht prezente, përjashtim i dukshëm është orizi, i cili nuk është në gjendje të sintetizojë fruktanet. Fruktanët me gjatësi 2-9 njësi quhen oligofruktozë ose fruktooligosaharide dhe ato me >10 njësi si inulina.
Ushqimet që janë të pasura me fruktane mund të përfshijnë produkte gruri - të tilla si buka dhe pastat - qepë, hudhra, elbi, lakra, brokoli, fëstëku, angjinarja, rrënjët e çikores dhe asparagu.
Prodhimi komercial i fruktaneve zakonisht bëhet me anë të përpunimit të bimëve, siç janë: çikorja (chicory) dhe argjinarja e Jerusalemit (Jerusalem artichoke). Inulina (grumbull heterogjen i polimerëve të fruktozës, lidhja β-2,1 ) për qëllime komerciale përfitohet (ekstraktohet) me anë të hidrolizës enzimatike (enzimi endo-inulinazë) e shoqëruar me tharje, ndërsa fruktooligosaharidet përfitohen nga saharoza dukë përdorur fruktoziltransferazën.
Fruktanet, inulina dhe frukto-oligosaharidet kanë status të përbërësve ushqimor në shumë shtete dhe klasifikohen si karbohidrate (substanca) në përgjithësi të sigurta (GRAS) në SH.B.A.
Fruktanet kanë përdorim të gjerë në produkte ushqimore, siç janë pjekurinat, drithërat e mëngjesit, pijet, dhe produktet e qumështit. Fruktanet e vogla (fruktooligosaharidet) kanë shije të ëmbël. Po ashtu fruktanet kanë afinitet të reduktimit të aktivitetit ujor dhe në këtë mënyrë përmisojnë stabilitetin mikrobiologjik.
POLISAHARIDET E USHQIMIT
Polisaharidet janë karbohidrate të cilat përbëhen prej më shumë se 10 (20) njësive të monosaharideve. Zakonisht ato ndahen si të qenët amidon dhe jo-amidon. Polisaharidet e amidonit përfaqësojnë burim të energjisë tek organizmi i njeriut ndërsa polisaharidet jo-amidonike zakonisht janë të patretshme dhe si të tilla janë të rëndësishme në mbrojtjen e organizmit tonë nga sëmundjet e ndryshme intestinale.
Polisaharidet mund të jenë të formuara nga një lloj i njëjtë monosaharidit (homoglukanet) dhe lloje të ndyshme të monosaharideve (heteroglukanet).
Lidhjet hidrogjenore benda molekulës së polisaharideve kanë rol të rëndësishëm në tretjen e polisaharideve në ujë.
Gjatë hidratimit, lidhja hidrogjenore ndryshon prej grupeve hidroksile të glukaneve (polisaharideve) në lidhjet hidrogjenore me ujë, ndërsa e kundërta ndodh gjatë procesit të dehidratimit.
Shumica e polisaharideve rrisin viskozitetin e tretësirës. Karakteristikat e tretësirave të polisaharideve varen nga struktura, pesha molekulare dhe koncentrimi.
Funksioni primar i amidonit tek bimët është se ai shërben si depo e energjisë dhe si burim i karbonit për biosintezën e re të makromolekulave. Amidoni përkohësisht mund të akumulohet në qelizat e kloroplasteve të cilat gjenden në indet fotosintetike. Shumica e amidonit haset tek organet deponuese të tij, siç janë endosperma e farave dhe rrënjët, si dhe tuberët e llojeve të ndryshme të bimëve.
Izolimi i amidonit nga drithërat e ndryshme përfshin qulljen (futjen në ujë), bluarjen e trashë, qërimin, bluarjen e imët, shoshitjen, ndarjen (separimin) centrifugale dhe tharjen. Izolimi i amidonit nga tuberët/rrënjët përfshinë pastrimin dhe qërimin e tuberëve. Amidoni dhe derivatet janë pothuajse të kudogjendur në sistemin e ushqimit. Ai (amidoni) ndikon në karakteristikat shqisore dhe karakteristikat stukturore (ang. textural) të ushqimit. Fuqia e saj ëmbëlsuese varet nga shkalla e hidrolizës. Vetëm dekstrinat që kanë një fuqi reduktuese në rritje (një shkallë e dekstrozës ekuivalente me 25-45) kontribuojnë deri në një farë mase për ëmbëlsinë e ushqimeve. Ekzistojnë tri lloje të amidonit në dietë, ato që hidrolizohen (treten) shpejt dhe të cilat mund t’i gjejmë në ushqimet e gatuara jo kohë të gjatë dhe në ushqimet që janë të pasura me amidon, ato që treten dobët të cilat gjenden në pasta dhe drithëra, dhe në fund amidoni rezistent. Amidoni Rezistent (RS) është lloj amidoni i cili që nuk treten dhe përthithen në stomak ose zorrët e vogla dhe si i tillë kalon në zorrën e trashë. Amidoni rezistent nuk çliron glukozë brenda zorrës së hollë, por përkundrazi arrin në zorrën e trashë ku konsumohet ose fermentohet nga bakteret e zorrës së trashë (mikrobiota e zorrëve). Fermentimi i amidonit rezistent prodhon acide yndyrore me zinxhir të shkurtër, duke përfshirë acetatin, propionatin dhe butiratin dhe rritjen e masës së qelizave bakteriale. Acidet yndyrore me zinxhir të shkurtër prodhohen në zorrën e trashë ku thithen me shpejtësi nga zorra e trashë, më pas metabolizohen në qelizat epiteliale të zorrës së trashë, mëlçi ose inde të tjera. Amidoni Rezistent (RS) kategorizohet në pesë lloje:
RS1 – Amidon rezistent i patretur fizikisht, siç është ajo që gjendet në fara ose bishtajore dhe drithëra të papërpunuara. Ky amidon është i lidhur brenda mureve qelizore fibroze të ushqimeve të lartpërmendura.
RS2 – Amidon rezistent i paarritshëm për enzimat për shkak të konformimit të amidonit, si tek bananet jeshile, patatet e papërpunuara dhe amidoni i misrit me amilozë të lartë.
RS3 – Amidoni rezistent që formohet kur ushqimet që përmbajnë amidon (p.sh. orizi, patatet) gatuhen dhe ftohen, si p.sh. makaronat. Ndodh për shkak të retrogradimit, që i referohet proceseve kolektive të amidonit të tretur duke u bërë më pak e tretshme pasi nxehet dhe tretet në ujë dhe më pas ftohet.
RS4 – Amidoni që është modifikuar kimikisht për t'i rezistuar tretjes.
RS5 – Amidoni që gjenden në formë komplekse me lipide
Amidoni mund të përdoret si agjent për trashje të marmelatave dhe ëmbëlsirave tjera, si dhe stabilizues koloidal për salcën e sallatave. Ai mund të përdoret edhe si agjent mbulues, lustrues dhe xhel-formues në industrinë e çamçakëzave.
Polisaharidet me natyrë jo-amidonike kanë tri lloje kryesore të funksioneve: mund të shërbejnë si karbohidrate rezerve (p.sh. galaktomananet tek leguminozat endospermike), shërbejnë si material ndërtues (p.sh. polisaharidet e celulozës dhe hemicelulozës, siç janë araboksilani dhe pektina të cilat marrin pjesë në ndërtimin e murit qelizor) dhe mund të kenë edhe rol mbrojtës, p.sh.goma arabike (gum Arabic) eksudatet e të cilës prodhohen dhe mbrojnë bimën si rezultat i dëmtimeve të ndryshme.
Glikogjeni ekziston vetëm në botën e kafshëve (muskuj dhe mëlçi) dhe si edhe amidoni, nuk kontribuon në ëmbëlsinë e ushqimeve.
Prej polisaharideve jo-amidonike komerciale më i rëndësishmi padyshim është polisaharidi celulozë.
Celuloza dhe hemiceluloza formojnë pjesë të murit qelizor të të gjitha qelizave të perimeve. Ato dihet se janë polisaharide të formuara nga lidhjet e glukozës në pozicionin 1-4. Molekulat e celulozës mund të bashkohen dhe të formojnë grup (deng) të fibrave kristalore të cilat janë shumë të patretshme dhe të palëshueshme në ujë. Këto formacione (grupime) janë komponenta kryesore e murit qelizor tek bimët. Celulozën brenda qelizës e sintetizon enzima (kompleksi enzimatik) celulozë-sintetaza, e cila është e lokalizuar në membranën plazmatike. Në mënyrë komerciale si burim i celulozës shërbejnë pulpat e drurit dhe fijet e pambukut. Fijet e pambukut janë fibra të cilat mbesin në farat e pambukut pasi janë larguar fibrat e gjatë. Fibrat e pambukut përmbajnë 98% celulozë ndërsa druri përmban 40%-50% celulozë, 30% hemicelulozë dhe 20% ligninë.
Celuloza përdoret për qëllime të ndyshme në industrinë ushqimore (procesin e filtrimit të lëngjeve, si zëvendësues të lipideve në ushqimet me kalori të ulët, etj). Ajo (celuloza) nevojitet të modifikohet fizikisht ose kimikisht për t’u përdorur pastaj në sistemin ushqimor.
Lidhja e përzier β-glukane (ang.mixed linkage β-glucanes), zakonisht e njohur si β-glukani, është komponenta kryesore e murit qelizor të drithërave të zakonshme (elbi, tërshëra, gruri, thekra, etj). Zbërthimi hidrolitik i β-glukaneve bëhet me anë të enzimeve të njohura si glukanaza. Aftësia e mirë për të formuar viskozitet i bën këto karbohidrate shumë të përdorshme (agjent për trashje) në industrinë ushqimore (akulloret, ëmbëlsirat dhe salcat për sallata). Poashtu mund të përdoret edhe në industrinë e pijeve, dhe ka potencial të përdoret si zëvendësues i lipideve në mish dhe sistemin e produkteve të qumështit).
Karbohidrate tjera me rëndësi në produktet ushqimore janë edhe galaktomananet dhe glukomananet.
Si polisaharid kompleks i murit qelizor të bimëve e që vlen të përmendet është edhe pektina. Pektinat janë pjesë e lamelës së mesme të qelizave të bimëve. Ata janë polisaharide të acidit galakturonik në pozicionin 1-4 me grupet karboksile të cilat shpesh janë të metoksiluara. Ato janë të bollshme, sidomos në fruta. Në mënyrë komerciale pektina ekstraktohet prej bërësisë së frutave agrume dhe bërësisë së mollave. Në industrinë ushqimore pektina përdoret si agjent xhelatinizues, si agjent i trashjes dhe stabilizimit të produkteve ushqimore, siç janë marmelata, reçeli, produktet xhelatinoze dhe produktet tjera ushqimore. Ajo (pektina) mund të përdoret edhe si stabilizues i shpërndarjes së proteinave në produktet acidifikuese të qumështit, siç është jogurti dhe pijet tjera me bazë qumështi. Në industrinë ushqimore në rastë të caktuara nevojitet zbërthimi hidrolitik i pektinës, e kjo mundësohet me anë të enzimës së njohur si pektinazë.
Me rëndësi në industrinë ushqimore dhe atë të pijeve paraqesin edhe grupi i karbohidrateve i njohur si eksudatet rrëshinore (ang. exudates gums), siç janë: gum arabic, gum ghatti, gum karaya, etj, të cilat sintetizohen nga bimët si substanca mbrojtëse. Këto rrëshira (eksudate) gjejnë aplikim të madh në industrinë ushqimore, p.sh. gum arabic përdoret si stabilizues dhe emulsifikator në industrinë e pijeve, stabilizues i emulsionit vaj-ujë, etj.
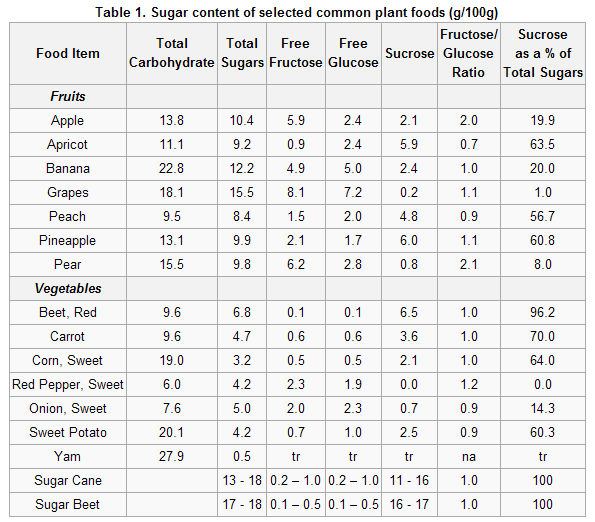
Tab.1. Përbërja e sheqernave në ushqimet e rëndomta bimore (g/100g)
Fibrat dietike janë kryesisht karbohidrate komplekse të patretura në murin e qelizave të bimës (celuloza, hemiceluloza dhe pektina), një shumëllojshmëri e “gomave” (rrëshirë, rezinë, etj), mukozave dhe polisaharideve të algave. Për fibra, zorra e njeriut nuk zotëron enzima të nevojshme për të “thyer” lidhjet ndërmjet njësive të sheqerit. Fibra e patretur udhëton nëpër zorrën tonë dhe duke bërë kështu siguron përfitime shëndetësore. Fibra gjithashtu inkurajon rritjen e baktereve të shëndetshme në zorrën tonë të madhe.
Përfitimet vijnë nga dy lloje të ndryshme të fibrave bimore që klasifikohen në bazë të asaj nëse treten ose jo në ujë.
Fibrat e tretshme janë të “tretshme" në ujë. Kur përzihet me ujë ato formojnë një masë xhelatinoze dhe mufaten. Fibrat e tretshme kanë shumë përfitime, duke përfshirë moderimin e niveleve të glukozës në gjak dhe uljen e kolesterolit. Emrat shkencorë për fibrat e tretshëm përfshijnë: pektinat, mukozat, disa hemiceluloza, β-glukanet, etj. Burime të mira të fibrave të tretshme përfshijnë: tërshërën dhe bollgurin, bishtajoret (bizelja, fasuli, etj), elbi, frutat dhe perimet (sidomos portokalli, molla dhe karrota).
Fibra e patretshme nuk absorbohen ose shpërndahet në ujë. Ato kalojnë përmes sistemit tonë të tretjes në afërsi të formës së tyre origjinale. Fibrat e patretura ofrojnë shumë përfitime për shëndetin e zorrëve, duke përfshirë zvogëlimin e rrezikut dhe shfaqjen e hemorroideve dhe konstipacionin (kapsllëkun), pra ndihmojnë në mënyrë direkte në funksionimin e peristaltikës (lëvizjen) së zorrëve. Emrat shkencorë për fibrat e patretura përfshijnë: celulozën, ligninën dhe gjithashtu disa hemiceluloza të tjera. Shumica e fibrave të patretura vijnë nga shtresat e krundeve të kokrrave të drithërave. Është e rëndësishme që të konsumohen të dy llojet e fibrave për përfitime maksimale shëndetësore.
Lignina është një komponent josheqeror i fibrave dietike i cili gjendet në muret e qelizave bimore. Ligninët janë veçanërisht të rëndësishëm në formimin e mureve të qelizave, veçanërisht në dru, sepse ato japin ngurtësi dhe nuk kalben lehtë.
ASPEKTE SHËNDETËSORE TË KARBOHIDRATEVE USHQIMORE
Glukoza dihet tanimë se paraqet burimin kryesor të energjisë në organizmin e njeriut. Glukoza, galaktoza, manoza, fruktoza, ksiloza, N-acetilglukozamina, N-acetilgalaktozamina dhe acidi N-acetilneuraminik janë identifikuar si substance esenciale për sintezën e glikoproteinave tek njerëzit, të cilat (glikoproteinat) dihet se janë tepër të rëndësishëm për ndërtimin e vendeve receptore të qelizës dhe vendeve njohëse në qelizë.
Krahas glukozës edhe saharoza siguron burim të shpejtë të energjisë për organizmin e njeriut, edhe pse mbikonsumi manifestohet me problem shëndetësore, siç janë: obeziteti, kariesi, etj.
Rafinoza dhe stakioza munden të promovojnë rritjen e popullatës së Bifidobacteria në intestinën e madhe, dhe si rrjedhojë kanë potencial si përbërës prebiotik në produktet e ndryshme të ushqimit.
Sikurse rafinoza dhe stakioza poashtu edhe fruktanet tek njeriu nuk mund të hidrolizohen me anë të enzimeve të traktit digjestiv (zorra e hollë), mirëpo edhe këto shërbejnë si prebiotik për bakteret e dobishme të kolonës - Bifidobacteria, duke ndaluar (supresuar) në të njëjtën kohë zhvillimin e baktereve patogjene. Po ashtu është parë edhe efekti i fruktaneve në rritjen (fuqizimin) e sistemit imunitar, aktiviteti anti-tumoral, etj.
Sa i përket amidonit, dihet se një pjesë e tij zbërthehet (tretet) pas konsumimit me anë të ushqimit në zorrën e hollë, mirëpo një pjesë e tij nuk mund të zbërthehet dhe ky lloj amidoni njihet edhe si amidoni rezistent. Amidoni rezistent paraqet burim të fibrave në sistemin ushqimor.
Në grupin e karbohidrateve të ushqimit të cilat kanë rol të caktuar shëndetësor në organizmin tonë e që vlen të përmendet janë edhe β-glukanet. Ato janë raportuar se reduktojnë nivelin e kolesterolit dhe nivelin e glukozës në serum.
Edhe karbohidrati (polisaharidi) pektinë është parë të ketë efekt pozitiv për shëndetin tonë. Konkretisht është parë se fibrat e pektinës reduktojnë nivelin e kolesterolit të serumit (duke interferuar në absorbimin e kolesterolit), reduktojnë absorbimin e glukozës, kanë efekt të caktuar prebiotik duke mundësuar rritjen dhe zhvillimin e Eubacterium dhe Bifidobacterium, etj.
Efektet Negative të Sheqernave në Shëndet
Ne konsumojmë shumë sheqer të shtuar. Burrat e rritur marrin mesatarisht 24 lugë çaji sheqer të shtuar në ditë, sipas Institutit Kombëtar të Kancerit (SH.B.A.) Kjo është e barabartë me 384 kalori.
Se si sheqeri në fakt ndikon në shëndetin e zemrës nuk është kuptuar plotësisht, por duket se ka disa lidhje indirekte. Për shembull, sasitë e larta të sheqerit mbingarkojnë mëlçinë. “Mëlçia juaj metabolizon sheqerin në të njëjtën mënyrë si alkooli dhe i shndërron karbohidratet e dietës në yndyrë”. Me kalimin e kohës, kjo mund të çojë në një akumulim më të madh të yndyrës, e cila mund të kthehet në sëmundje të mëlçisë yndyrore (steatoza hepatike) një kontribues në diabetin, i cili rrit rrezikun për sëmundje të zemrës.
Konsumimi i tepërt i sheqerit të shtuar mund të rrisë presionin e gjakut dhe të rrisë inflamacionin kronik, që të dyja janë rrugë patologjike drejt sëmundjeve të zemrës. Konsumi i tepërt i sheqerit, veçanërisht në pijet me sheqer, gjithashtu kontribuon në shtimin e peshës duke mashtruar trupin tuaj për të fikur sistemin e tij të kontrollit të oreksit, sepse kaloritë e lëngshme nuk janë aq të kënaqshme sa kaloritë nga ushqimet e ngurta. Kjo është arsyeja pse është më e lehtë për njerëzit që të shtojnë më shumë kalori në dietën e tyre të rregullt kur konsumojnë pije me sheqer.
“Efektet e marrjes së sheqerit të shtuar – presioni i lartë i gjakut, inflamacioni, shtimi në peshë, diabeti dhe sëmundja e mëlçisë yndyrore – janë të gjitha të lidhura me një rrezik në rritje për atakun në zemër dhe goditjen në tru.
Shoqata Amerikane e Zemrës sugjeron që gratë të konsumojnë jo më shumë se 100 kalori (rreth 6 lugë çaji ose 24 gramë) dhe burrat jo më shumë se 150 kalori (rreth 9 lugë çaji ose 36 gram) sheqer të shtuar në ditë.
REAKSIONET E KAFËZIMIT NË KARBOHIDRATE (ANG. BROWNING REACTIONS)
Ekzistojnë në përgjithësi tri reaksione të kafezimit në ushqim: kafezimi oksidativ/enzimatik, karamelizimi dhe reaksioni jo-oksidativ/jo-enzimatik (reaksioni i Mailardit ose kafezimi i Mailardit).
I pari (kafezimi oksidativ/enzimatik) ka të bëjë me proteinat (enzimat) ndërsa karamelizimi dhe reaksioni jo-oksidativ/jo-enzimatik kanë të bëjnë edhe me reaksionet e karbohidrateve.
Karamelizimi përfshin një mori komplekse të reaksioneve të cilat janë rezultat i nxemjes direkte të karbohidrateve, e në veçanti sheqernave. Reaksionet e dehidrimit rezultojnë në formimin e lidhjeve dyfishe së bashku me polimerizimin e strukturave unazore të cilat absorbojnë gjatësi të ndryshme valore, aromatizimin, errësimin dhe ngjyrosjen e këtyre përzierjeve.
Dy role kryesore të karamelizimit në industrinë ushqimore janë: shija e karamelit dhe prodhimi i ngjyrës, proces ky në të cilën saharoza nxehet në tretësirën acidike ose tretësirën e kripërave acidike të amonit për të prodhuar lloje të ndryshme të ushqimit, sheqerkave (ëmbëlsirave) dhe pijeve.
Reaksionet e Mailard-it (ang.Maillard Reactions) janë reaksionet më të rëndësishme që hasen në sistemin ushqimor, dhe të cilat njihen edhe si reaksionet e kafezimit jo-oksidativ/jo-enzimatik. Sheqernat reduktues dhe aminoacidet ose edhe përbërësit tjerë të cilët përmbajnë N reagojnë për të prodhuar N-glikozidet duke zhvendosur ngjyrën nga e kuqe-kafe në ngjyrë kafe intensive, me aromë karameli dhe si rezultat formohen melanoidinet koloidale të patretshme.
Në përgjithësi reaksionet Mailard janë shumë komplekse dhe si të tilla mund të rezultojnë me aromë, shije dhe ngjyrë të dëshiruar ose ndonjëherë edhe të padëshiruar.