Chapter 11 - Membrane Structure and Function
Membranes allow for the creation of separate environments, and compartmentalization. They consist of 2 molecules vertically, creating a lipid bilayer and semi-permeable barriers.
Eukaryotic cells have membranes within the main membrane, allowing for environments with different conditions, like varying pHs. Cytosol is 7.2, while mitochondrion are 8. Lysosomes are 5. Lumen are oxidizing, cytosol is reducing.
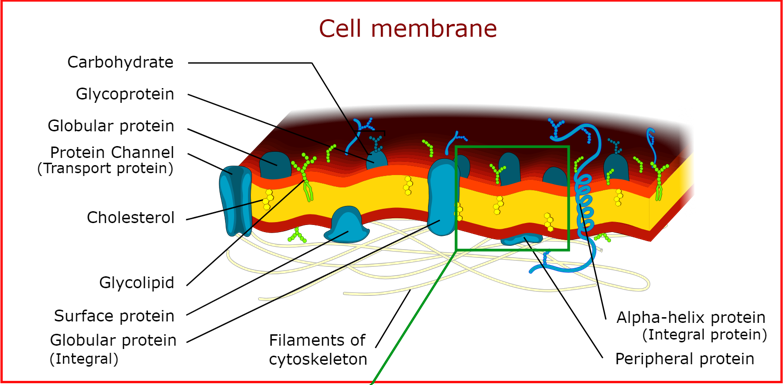
Membranes are mosaics, with an outer and inner facing side. Embedded, there are molecules such as cholesterol, peripheral proteins which are only exterior, channel proteins which travers the membrane to allow certain molecules in/out, and sugar side chains which may be attached to certain proteins.
A given membrane will be, by weight, 45% lipid, 5% protein, and 5-10% carbohydrate, or functionally half protein half lipid.
The membrane is made of phospholipids, with hydrophobic inner-facing tails and hydrophilic outer-facing heads. This makes it amphiphilic, with both portions though not necessarily equally hydrophobic and hydrophilic.
Being semipermeable, non-polar small molecules (like O2, CO2, N2, steroids, and hormones) are allowed in and out on their own via diffusion, and small polar but uncharged molecules (like H2O, ethanol, and glycerol) sometimes make it through by diffusion, but not as easily. There are sometimes channels for these, like aquaporins. Larger uncharged polar molecules (like amino acids, glucose, and nucleosides) usually don’t make it through on their own and require channels, and ions never make it through on their own without channels and transporters.
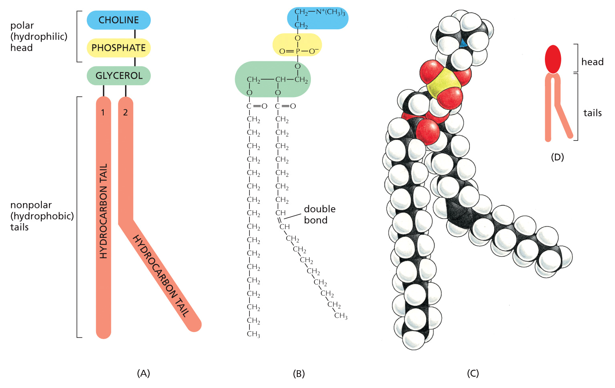
The main phospholipid in animal cell membranes is phosphoglycerate, which have varying head structures, phosphate, and glycerol as head components. Their tails are hydrocarbons, being mostly straight but forming kinks where there are double bonds. In this example it is phosphatidyl choline (spelling?)
Lipid bilayers are facilitated by hydrophobic forces. These exist between polar and non-polar objects.
In vivo membranes are formed via the budding of an existing membrane. Forming a sphere is more energetically favorable.
If there are many phospholipids in a given area, when exposed to water they will spontaneously form a lipid bilayer
Membranes are described under the “Fluid Mosaic Model”, as they have the same dynamic movement of molecules in the plane of the membrane as a fluid does. Individual molecules will rotate and flex their tails.
If temperature rises, so does membrane fluidity and permeability. This happens in reverse too. Drops in temperatures lowers fluidity and permeability. These aren’t good for cell survivability. The amount of double bonds can be changed (more means more permeability). Long tails decrease fluidity, and short tails increase it.
One of the membrane’s main roles is the maintain fluidity and permeability in response to environmental changes, like rising temperatures. More cis double bonds in the tails will lead to higher fluidity. Shorter tails does the same, increasing fluidity. Cholesterol increased the permeability of the barrier.
A saturated fatty acid will have all single binds between hydrocarbons in the tail, whereas an unsaturated fatty acid will have at least one double bond.
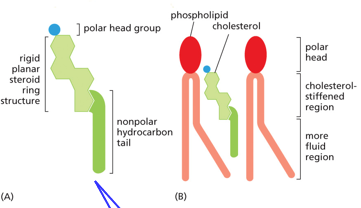
Cholesterol is an essential membrane component, existing primarily in eukaryotic membranes. It has a rigid steroid ring structure, and the polar head group facilitates orientation within the lipid bilayer. More cholesterol means less fluidity (more VDW interactions). The headgroup of cholesterol prevents FA headgroups from clustering, it fits between phospholipids. The only membrane that doesn’t contain this is the inner-mitochondrial membrane
HDL is “good” cholesterol, and LDL is “bad” cholesterol. HDL gets transported to the liver for removal, while LDL builds up on artery walls. These are high/low density lipoproteins, and have nothing to do with the cholesterol themselves but more so the form that carries them that is made of lipid and protein.
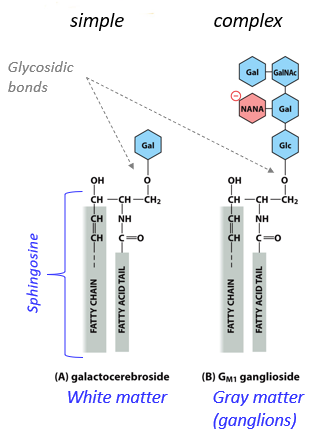
A glycolipid is a lipid with a glycosidic bond, connected to a sugar. They have a sphingosine basis, and exist only on the extracellular side of a membrane. They self-associate with each other due to polarity/charges, and have simple/complex forms. They function to help protect membranes from harsh conditions, influence electric fields and ionic concentrations when charged, aid in cell-cell recognition, and form the cell coat (glycocalyx) with glycoproteins.
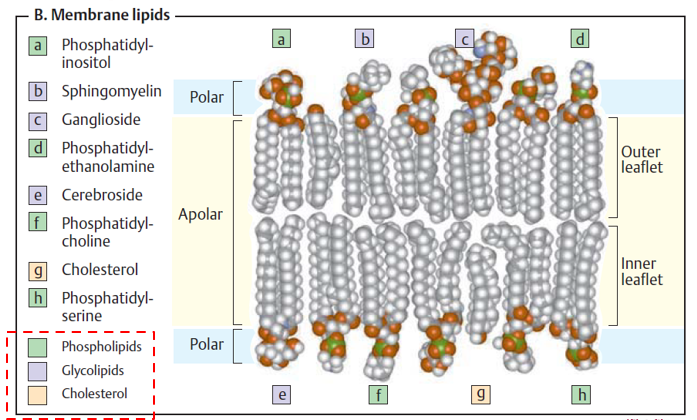
The composition of a given membrane is diverse, and varies based on cell type
The lipid bilayer is asymmetric, meaning the population of each side is distinct between the leaflets. There are different compositions facing extracellularly and the cytosol. Cholesterol is commonly in both.
A lipid raft is a local area that exhibits low fluidity, facilitated by cholesterol. They are organizing centers for the assembly of signaling molecules. Being more orderly and tightly packed than the surround bilayer, they influence membrane fluidity and protein trafficking.
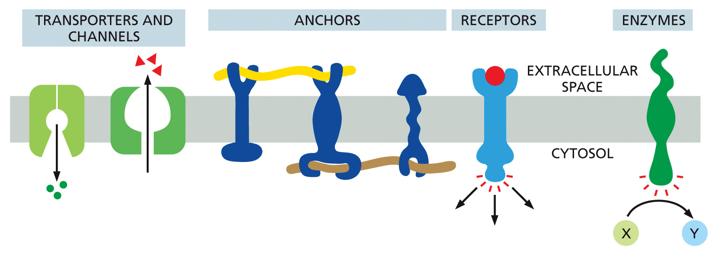
There are many different kinds of proteins within a membrane, namely transport channels, anchors, receptors, and various enzymes to name a few. They are larger than lipids, meaning there are fewer of them despite being around half of the membrane’s mass.
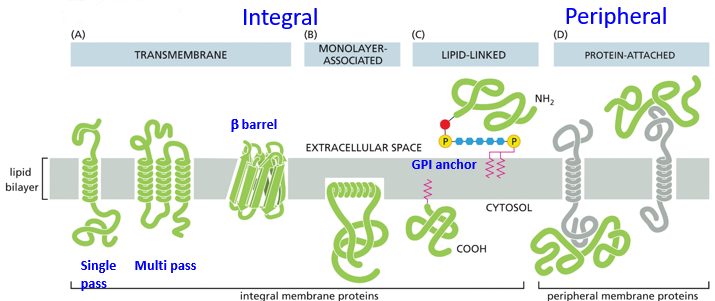
Transmembrane proteins go through both layers while monolayer-associated don’t. They can be lipid-linked as well, existing outside but bound to the membrane. These are further subdivided into single pass, multi pass (for transmembrane). Peripheral proteins may attach to other proteins on the membrane, typically associating with transmembrane proteins. Above, c is an anchor. Transmembrane proteins are often formed by alpha helices
Multi pass proteins may start crossing the membrane with lipid molecules between passes, through once properly folded will be next to each other.
Beta barrels tend to form transmembrane channels, like porin proteins
Proteins with oligosaccharide and disulfide bonds can only exist extracellularly, as cytosol is reducing
A cell coat is a carbohydrate rich cell surface, with glycolipids, glycoproteins, and proteoglycans. This aids cell-cell recognition and adhesion
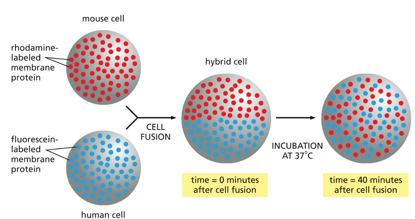
Things like FRAP and mouse-human hybrid cells show that over time, proteins will diffuse around a membrane and intermix.
The movement of proteins can be restricted using molecular fences. This is done with epithelial cells particularly. This can happen through aggregate assembly, interactions with external molecules, interactions with extracellular proteins on other cells, and molecular fences
Everything outside the cell is glycosylated. There are also proteoglycans. Glycoproteins are proteins that carry sugar, and proteoglycans are sugar structures that have adaptive proteins. They are named off the main active structure. Proteoglycans accompany water molecules.