Con Bio Exam 2
heterozygosity
H= proportion (%) of genes the average individual is heterozygous or proportion of heterozygous individuals in a population
heterozygosity usually greater at larger population sizes
(Heterozygosity is a measure of genetic variation within a population, representing the presence of different alleles at a gene locus in an organism’s DNA. An individual is heterozygous for a gene if they carry two different alleles (one from each parent) at a particular gene locus. On a population level, heterozygosity refers to the proportion of individuals that are heterozygous at a given gene locus or across the genome, indicating genetic diversity within that population.)
population divergence
population genetic divergence (Dpt) may also play a key role in local fitness and population survival
population may eventually diverge to the point of local adaptation
ecotype ≈ ecophenotype
(Population genetic divergence, often represented as Dpt, is a measure of genetic differentiation between populations of the same species. This metric quantifies how much genetic variation exists between populations relative to the variation within each population. Genetic divergence can be driven by factors such as natural selection, genetic drift, mutations, or barriers to gene flow (e.g., physical separation or environmental differences).
The Dpt value can range from 0 to 1:
Dpt=0 indicates no genetic differentiation between populations, meaning they share similar allele frequencies and are genetically very similar.
Dpt=1 indicates complete genetic divergence, meaning the populations do not share any genetic material at the loci being studied.
High genetic divergence (high Dpt) suggests limited gene flow and that populations are evolving independently, while low genetic divergence (low Dpt) suggests ongoing gene flow or recent common ancestry. Monitoring Dpt is crucial in conservation biology for managing genetic diversity, especially for species with fragmented habitats or isolated populations. )
panther paper (READ)
bb is blind wolf
consequences and benefits of genetic rescue
Benefits:
The paper on the genetic rescue of the Florida panther highlights several significant benefits from introducing genetic material from Texas pumas. These benefits included:
1. Increased Population Size: After the genetic rescue, the Florida panther population size tripled, indicating a successful rebound. The translocation of Texas pumas led to more robust reproduction rates, which helped stabilize and grow the population.
2. Improved Genetic Diversity and Heterozygosity: Genetic diversity and heterozygosity levels doubled after the introduction of new genes. This increase in genetic diversity helped reduce the inbreeding depression effects that had plagued the population.
3. Enhanced Survival Rates and Fitness: Panthers with mixed (admixed) genetics had higher survival rates compared to the purely native Florida panther lineage. This improvement was evident in various fitness traits, such as increased survival of kittens and subadults, lower mortality rates, and reduced prevalence of physical defects linked to inbreeding.
4. Reduction in Inbreeding-Related Defects: The Florida panther population previously exhibited physical and reproductive issues, including cryptorchidism (undescended testes), kinked tails, and heart defects, all associated with inbreeding depression. Following genetic rescue, these issues decreased, and overall health markers improved in the population.
5. Increased Adaptability and Resilience: The admixed panthers demonstrated behaviors associated with higher fitness, such as improved escape responses during captures, suggesting enhanced physical resilience. Increased genetic diversity also likely improved adaptability to environmental challenges.
These benefits reflect the positive outcomes of genetic rescue in counteracting the negative effects of inbreeding, enhancing overall population health, and contributing to a more sustainable population structure for the Florida panther【22†source】.
Consequences:
Yes, the paper "Genetic Restoration of the Florida Panther" does discuss some potential consequences and challenges following the genetic rescue. Here are a few key points:
1. Ongoing Threats Despite Genetic Rescue: While genetic diversity and overall fitness improved, the Florida panther population continues to face threats like habitat loss and limited territory, which can lead to increased density-dependent issues, such as intraspecific aggression. These ongoing challenges mean that genetic rescue alone may not secure the panther's long-term survival without additional conservation efforts to expand and protect habitats.
2. Potential Habitat Saturation: As the population grew, there was concern about reaching a saturation point in available habitat, which could result in overcrowding and increased competition for resources. Habitat saturation can lead to heightened mortality rates and potentially hinder the benefits gained from genetic diversity.
3. Increased Disease Risk: The paper highlights concerns over diseases that might impact the panthers as their population density increases, particularly feline leukemia virus (FeLV) and feline immunodeficiency virus (FIV). Although FeLV infections were reduced through a vaccination program, higher population densities could make disease transmission more likely, which poses a risk to the health and stability of the population.
These consequences illustrate that while genetic rescue can effectively improve genetic diversity and reduce inbreeding depression, it must be integrated with other conservation strategies, such as habitat protection, to ensure a sustainable and resilient population【22†source】.
The article you provided, "Genetic Restoration of the Florida Panther," discusses the genetic rescue effort initiated for the Florida panther in 1995. This effort aimed to counteract the harmful effects of inbreeding by introducing genetic material from eight female Texas pumas. Before the intervention, the Florida panther population was facing severe inbreeding depression, with issues like poor sperm quality, cryptorchidism (undescended testicles), heart defects, and other health problems due to low genetic diversity.
The introduction of Texas pumas was a success, as it increased genetic diversity and heterozygosity, which doubled over time. This genetic diversity boosted population health, leading to improved survival rates and a significant increase in population size. However, ongoing challenges like habitat loss and potential saturation of available resources still pose threats to the long-term viability of the Florida panther population【22†source】.
Owl paper (READ)
abstract and conclusion (hybridization)
impact on rare and endangered species
The Haig et al. (2004) paper focuses on the genetic identification of hybrids between Northern Spotted Owls and Barred Owls, detailing both the legal and conservation implications of hybridization between these two species. Here’s a breakdown of key points and consequences related to genetic rescue:
1. Hybridization as a Conservation Concern: Hybridization with the invasive Barred Owl could pose a threat to the genetic integrity and survival of the Northern Spotted Owl. Barred Owls, which have expanded westward, now occupy overlapping territories with Spotted Owls, leading to interbreeding that could reduce the genetic distinctiveness of the threatened Northern Spotted Owl.
2. Genetic Markers for Conservation: The study developed mitochondrial DNA and AFLP (amplified fragment-length polymorphism) markers to distinguish hybrids from pure-bred individuals. These markers allow for more accurate identification of hybrids, which is critical for conservation efforts aimed at preserving the genetic identity of Northern Spotted Owls.
3. Legal Implications and Protection Challenges: The U.S. Endangered Species Act (ESA) does not provide clear protection for hybrids, complicating conservation and legal efforts for the Northern Spotted Owl. Hybrid identification is essential because Spotted Owls receive protections under the ESA, while hybrids may not, leading to ambiguity in enforcement against their harm or “take.”
4. Potential for Genetic Threats: Continued hybridization and backcrossing (mating of hybrids with either parent species) could lead to outbreeding depression, which occurs when offspring from genetically distant populations (or species) have lower fitness. This could further compromise the Northern Spotted Owl population if hybrid offspring face survival or reproduction issues.
Overall, the paper highlights the importance of genetic research in addressing complex hybridization issues in conservation, particularly where legal protections for hybrids remain unclear.
review inbreeding coefficients (4 parts)
describe pedigree
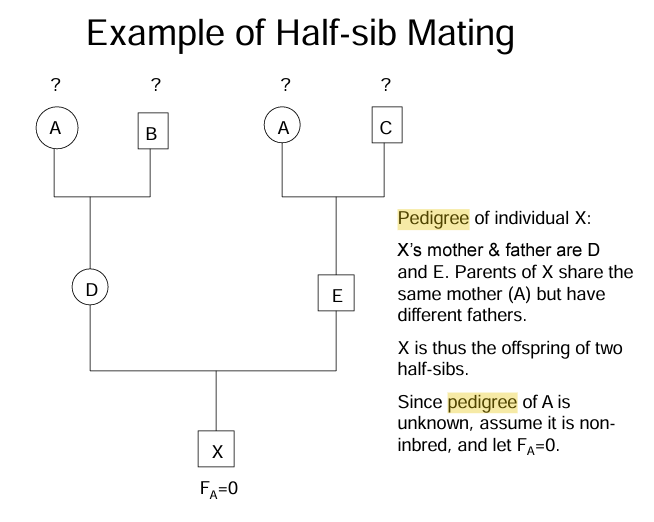
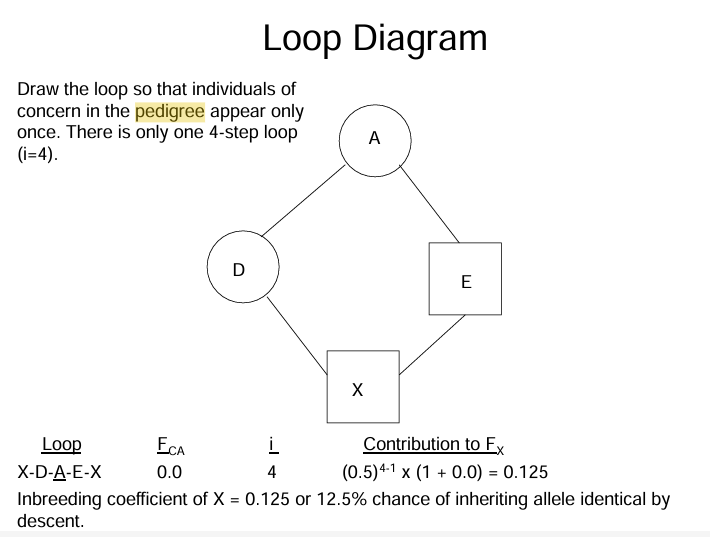
do loop diagram
calculate inbreed coefficient
what does the coefficient mean
Calculation explanation:
Inbreeding coefficient ranges from 0-100% (Fx=0-1) represents the probability of alleles being identical by descent
identical by descent=inherited from the same individual
two of the basic approaches to calculate the inbreeding coefficient is the path analysis technique and the additive relationship method
Path Analysis Technique:
calculates inbreeding coefficient (Fx) X represents individual of interest
draw the pedigree so that any common ancestors appear only one.
a common ancestor is any individual related to both parents of the individual of concern (X)
if there are no common ancestors, then Fx=0 and X is noninbred
If there is no pedigree information on the common ancestors it’s assumed to be noninbred. If the common ancestor is inbred, the its inbreeding coefficient Fca, must be calculated before calculating Fx.
Determine loops in the pedigree. a loop is a path that runs from X, through one parent, to the common ancestor, through the other parent, and back to X without going through any individual more than once
Determine the number of steps in each loop
calculate the contribution of each loop to Fx as:
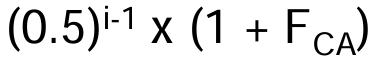
where Fca= inbreeding coefficient of the common ancestor, and i= number of steps in each loop
Sum the contributions of each loop. the summation of all contributions will be the Fx.
island types
Temporary (MS River sandbars)
Continental (MS Gulf islands)
Oceanic (Caribbean)
Island Colonization factors
Island size
Distance to mainland and/or source of colonists
latitude
topography
disturbance regime (i.e. Caribbean hurricane alley)
Dispersal to oceanic islands occurs mostly by sweepstake route. Organisms that colonize these islands may be able to survive in the open ocean for some time (flotsam islands), but not live in it
Ocean islands exhibit high rates of endemism but lower overall species richness.
Plant and animal species best suited for colonized through the air. Small flying forms can ride wind currents, many insects can colonize islands this way
Island Colonization Process
Developing adaptations to island environments can be difficult. Why?
Organisms arrive with adaptations to mainland environments, including morphological and behavioral adaptations (e.g., foraging, mating).
Amount of genetic variation in a colonizing population a fraction of the overall species variability (‘founder effect’).
Random non-adaptive changes to the gene pool of a founder population may lead to its extinction before it can become established.
Generalist species with good dispersal abilities usually make the best island colonizers.
Islands harbor few large predators due to geographic isolation.
Adaptive radiation- evolution of ecologically diverse species from an ancestral colonizer to island and/or island group
species arising from radiation fill different available ecological niches
Ecological release- in the absence of competition and/or predation, island species expand into a greater variety of habitats compared to closely related mainland species
Niche width of species expands due to relaxed competition
Density Compensation- compared with conspecific mainland populations, species on oceanic islands exhibit relatively greater densities
Coexisiting species more dissimilar in ecologically relevant traits than would be expected by chance
Many examples of islands with either widely radiated groups or highly unusual vertebrate forms
“Adaptive space” provided by isolation may promote appearance of dwarfism and gigantism
large animals undergo dwarfism while small animals undergo gigantism
Island Biogeography Example
Suppose an island in M-W equilibrium supports 75 species out of a source pool of 100 species. Maximum extinction rate (E) is 10 extinctions/year. Calculate:
1. Calculate maximum immigration rate (I) at equilibrium.
2. If (I) were doubled:
a. what is the new number of species at equilibrium?
b. what is the turnover rate?
Ŝ = IP/I+E ; solving for I; Ŝ(I+E) = IP ŜE = IP – IŜ
I = ŜE/P-S
Replacing values into formula yields
I = (75)(10)/(100-75)
I = 30 species/year
If immigration rate is doubled, I = 60, use Ŝ = IP/I+E,
a. Ŝ = (100)(60)/(60+10), Ŝ = 85.7 species at equilibrium
b. Turnover: T(hat) = IE/I+E = (60)(10)/(60+10) = 8.57 species
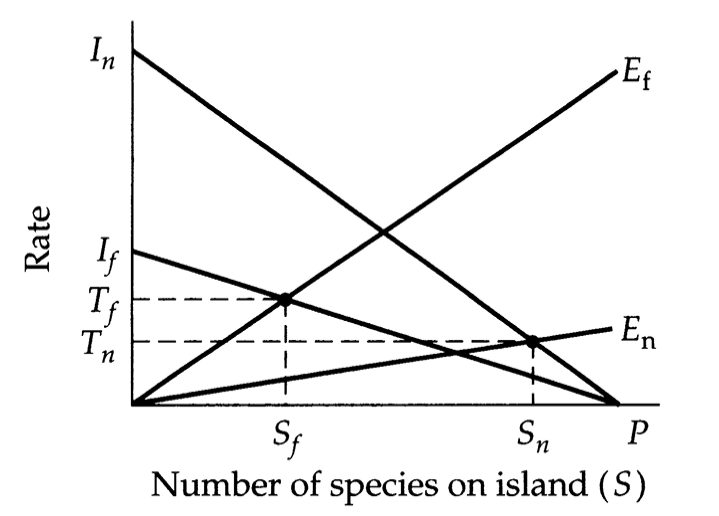
Rescue Effect
Reduction in extinction rate of near islands versus distant ones
M-W model predicts higher turnover on near islands
Rescue effect may increase turnover on more distant islands

As S depends on Immigration and Emigration rates, distance to mainland source may override island size
Target Effect
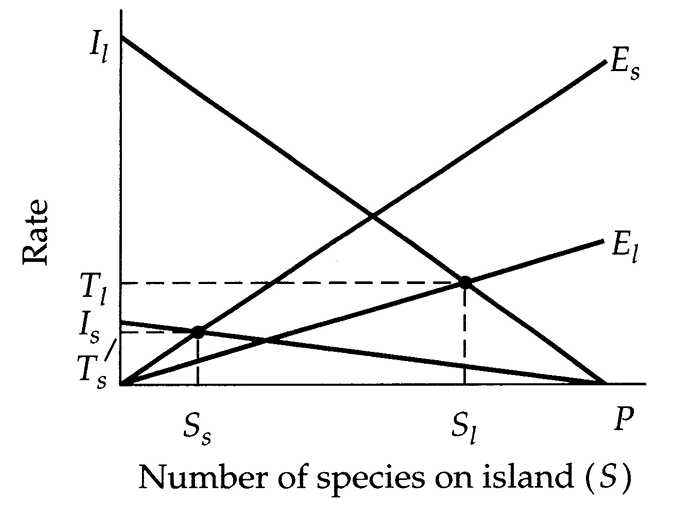
Increase in immigration rate on large islands versus small ones.
M-W model predicts higher turnover on small Islands.
Target effect may increase turnover on large islands.
Metapopulations
Levins (1969) first described the term
“A collection of subpopulations linked by immigration and extinction process”
Metapopulations are defined by a balance between extinction rates in occupied patches and the colonization rate of empty patches.
Extinction rate related to local patch conditions and stochastic nature of small populations
Colonization rate depends on dispersal ability and location of suitable patches in the landscape
Subpopulations in each patch fluctuate in size, rate dependent on subpopulation size
local extinctions prevented by rescue effect of neighboring patches.
Metapopulations cont’d
“A population of populations”
Spatial scale allows dispersal among patches.
movements constrained if patches separated by expanses of unsuitable habitat
under metapopulation concept, habitat patches are of similar quality
Rate of movement frequent enough to avoid genetic differentiation but rare enough to generate patch-specific population dynamics
birth/death rates become patch-specific
However, not ALL patchily-distributed species are metapopulations
Metapopulation models describe structure and dynamics
Key distinction lies in being able to identify:
individual subpopulations (not as easy as you may think)
habitat patches available (hard to define)
links critical to metapopulation maintenance
Three key processes for metapopulations
Extinction:
constant (risk) multiplied times number of occupied patches
Colonization
dependent on number of occupied (sources) and empty (targets) patches
Turnover
Extinction of local populations and establishment of new local populations in empty habitat patches by migrants from existing local populations
focus on populations not species (contrast to island biogeography)
key to determine metapopulation “persistence”
Metapopulation Types (name 1 and describe)
General metapopulation “Levins metapopulation”
Large network of similar-size small patches
local patch dynamics occur at faster time scale than metapopulation dynamcis
sometimes used to describe system where all local populations have a high risk of extinction
Mainland-island metapopulation “Boorman-Levitt metapopulation”
set of patches (“islands”) within dispersal distance from large patch (“mainland”) population that never goes extinct
Metapopulation persists only if ‘mainland’ patch persists
Source-sink metapopulation
subpopulations with negative growth rates (in absence of dispersal) at low population numbers
source patches generate dispersers
Non-equilibrium metapopulation
long-term extinction rates exceed colonization
extreme case isolation among subpopulations is so that dispersal (and hence recolonization) is precluded
Example
A metapopulation exists as a set of habitat islands composed of small satellite populations and an adjacent large population that serves as a permanent source of colonists .
Assume large population is only source extinctions of propagules and that on satellite populations are independent of each other
a. If pi = 0.2, and pe = 0.4:
a. calculate the fraction of patches occupied at equilibrium.
b. A development proposal calls for elimination of the source population while retaining the satellite populations as nature reserves. Assuming values for pi and pe remain constant:
predict metapopulation fate once the source population is eliminated
Solutions:
Source of colonists is external and extinctions are independent, metapopulation is described by the island-mainland model
island-mainland f(hat)= pi/pi+pe = 0.2/0.2+0.4 = 0.33.
approximately one habitat ‘island’ in three (33%) will support a population
Once the source (mainland) population is removed, colonization is strictly internal while extinctions remain independent
f(hat) = (1-pe/i) = (1-0.4/0.2) = (1-2) = -1.
as f(hat) <0, populations in all patches will go extinct despite being designated as reserves
Metapopulation persistence depends on retaining source population