H5 GENETICA
Je kan de geslachtelijke en ongeslachtelijke voortplanting beschrijven en daarmee de genetische variatie bij prokaryoten en eukaryoten verklaren %%(BINAS 86D)%%****:
- ^^Geslachtelijke voortplanting^^: genen worden verdeeld (=^^recombinatie^^).
- Een ^^gen^^ is een bepaald stuk DNA van een chromosoom, codeert voor een bepaalde eigenschap. De ^^locus^^ is de plaats van het gen op het chromosoom.
- Een ^^allel^^ is één van de verschillende varianten van een gen. ^^Homozygoot^^ = beide allelen zijn hetzelfde; ^^heterozygoot^^ = beide allelen zijn verschillend).
- ^^Ongeslachtelijke voortplanting^^: nakomelingen hebben allemaal hetzelfde genotype (=^^kloning^^).
De combinatie van allelen samen op één chromosoom noem je een ^^haplotype^^.
^^Crossing-over:^^ tijdens meiose wisselen ^^homologe chromosomen^^ stukken DNA uit.
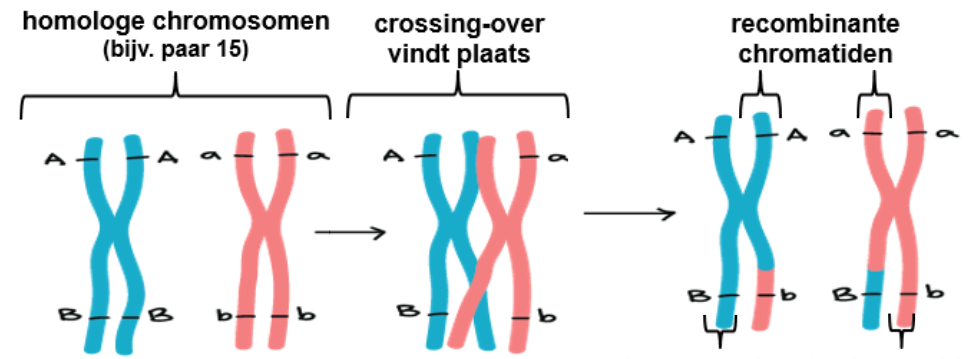
^^Mutaties^^ door bepaalde stoffen/straling leiden tot verandering in het DNA en variatie in genotypen.
Je kan uitleggen dat een fenotype tot stand komt door de combinatie van genotype en de invloed van milieufactoren, en verschillen herkennen met de epigenetische overerving:
Het ^^fenotype^^ zijn de zichtbare kenmerken van de tot expressie gebrachte genen onder invloed van de omgeving. Het ^^genotype^^ is de genetische samenstelling van een individu.
- ^^Epigenetica^^: studie van overerfbare veranderingen in het DNA, die niet in de nucleotidenvolgorde terug zijn te vinden. Het betreft veranderingen in de schakelaars, die bepaalde genen ‘aan’ en ‘uit’ zetten.
Je kan de verschillen tussen autosomen en geslachtschromosomen benoemen en toelichten dat bij de mens de geslachtschromosomen het geslacht bepalen %%(BINAS 70B)%%****:
^^Autosomen:^^ alle 23 gepaarde chromosomen
^^Geslachtschromosomen^^: bij zoogdieren man XY en vrouw XX. Het SRY-gen op het Y chromosoom stimuleert bij een embryo de vorming van mannelijke geslachtshormonen.
- Karyotypes: man 46,XY en vrouw 46,XX
Je kan de frequentie van genotypen en fenotypen van nakomelingen afleiden uit stambomen of kruisingsschema’s bij monohybride en dihybride kruisingen, zowel voor onafhankelijke als gekoppelde overerving, voor autosomale en X-chromosomale genen, multipele allelen en lethale factoren:
Het ^^dominante^^ (t.o.v het ^^recessieve^^) allel is het overheersende allel. Legenda stamboom:
![]()
- ^^Monohybride kruising:^^ dominante allel krijgt een hoofdletter, recessieve een kleine letter.
==Voorbeeld==: P-generatie Aa x AA (A is bruin haar, a is blond haar)
| A | A | |
|---|---|---|
| A | AA | AA |
| a | Aa | Aa |
genotype-ratio → AA:Aa = 1:1; fenotype-ratio → blond:bruin = 4:0
- ^^Dihybride kruising^^:
==Voorbeeld== (niet-gekoppeld): P-generatie AaBb x AaBb
| AB | Ab | aB | ab | |
|---|---|---|---|---|
| AB | AABB | AABb | AaBB | |
| Ab | AABb | AAbb | Aabb | |
| aB | AaBB | aaBB | aaBb | |
| ab | Aabb | aaBb | aabb |
Kans op AaBb = 1:4
Je kan ook losse kruisingsschema’s maken per gen en vervolgens de kansen met elkaar vermenigvuldigen.
==Voorbeeld== (gekoppeld → genen liggen op hetzelfde chromosoom, allelen erven in paren over): P-generatie GL//GL x gl//gl betekent F1-generatie GL//gl en F2-generatie …
| GL/ | gl/ | |
|---|---|---|
| GL/ | GL//GL | GL//gl |
| gl/ | GL//gl | gl//gl |
- ^^Intermediaire overerving:^^ beide allelen zijn even ‘sterk’ en bij combinatie van beide allelen ontstaat bij het fenotype een ‘mengvorm’.
==Voorbeeld==: P-generatie BrBr (rode bloem) x BwBw (witte bloem) betekent F1-generatie BrBw (roze bloem) en F2-generatie …
| Br | Bw | |
|---|---|---|
| Br | BrBr | BrBw |
| Bw | BrBw | BwBw |
genotype-ratio → BrBr : BwBw : BrBw = 1:1:2; fenotype-ratio → rood : wit : roze = 1:1:2
- ^^Lethale allelen:^^ een lethaal allel levert in homozygote toestand geen levensvatbaar individu op.
Voorbeeld: P-generatie AAy x AAy (met Ay is lethaal allel).
| A | Ay | |
|---|---|---|
| A | AA | AAy |
| Ay | AAy |
genotype-ratio → AA:AAy = 1:2
- ^^X-chromosomale overerving:^^ gen komt alleen voor op het X-chromosoom.
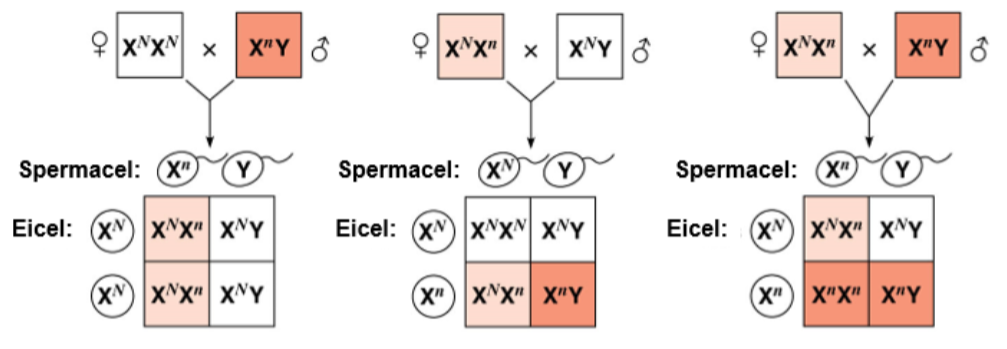
Is een eigenschap X-chromosomaal?
- Een dominante vader krijgt een recessieve dochter, of…
- Een recessieve moeder krijgt een dominante zoon, DAN NIET X-CHROMOSOMAAL.
- ^^Multiple allelen^^: overerving berust op drie of meer allelen.
==Voorbeeld==: overerving van bloedgroepen; Ia en Ib zijn ^^co-dominant^^ en i is recessief. P-generatie: Iai (A) x IaIb (AB)
| Ia | i | |
|---|---|---|
| Ia | IaIa | Iai |
| Ib | IaIb | Ibi |
genotype-ratio → Ibi:IaIb:IaIa:Iai = 1:1:1:1; fenotype-ratio → B:AB:A = 1:1:2
Je kan verklaren dat mitochondriale overerving en epigenetica kunnen leiden tot een ander overervingspatroon dan volgens de wetten van Mendel:
- ^^Mitochondriale overerving^^: kind erft mtDNA via de moeder.
- ^^Wetten van Mendel^^:
- alle individuen uit de eerste generatie vertonen hetzelfde kenmerk als het kenmerk van een van beide ouders.
- de verschillende kenmerken worden onafhankelijk van elkaar overgeërfd, indien ze op verschillende chromosomen liggen.
Je kan standpunten over het ingrijpen in de erfelijkheid van pro- en eukaryote organismen toelichten met ethische en biologische argumenten: