The Synapse
The Synaptic Cleft
The Synapse Diagram
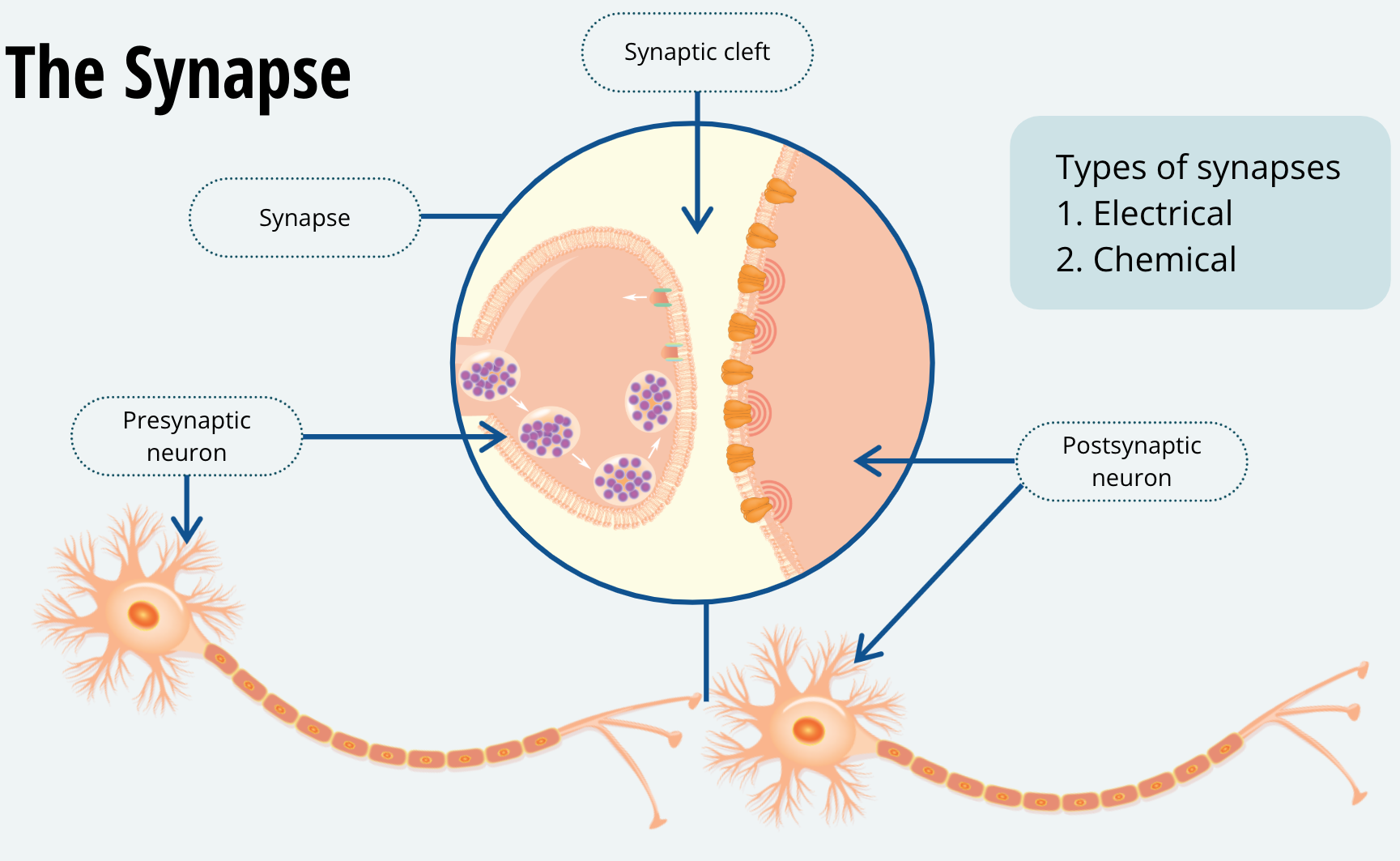
Now that you have an idea of how a neuron fires, it’s time to learn how each neuron conveys information to the subsequent neurons it communicates with. The junction between the terminal button of one neuron and the dendrite (or soma) of another neuron is called a synapse. There are two types of synapses, electrical and chemical. Electrical synapses are quite rare, and we won’t be covering them in this course. The chemical synapse is by far the most common form of communication between neurons. When describing the synapse, we call the neuron that is sending the message the presynaptic neuron and the neuron that is receiving the message, the postsynaptic neuron. Within the synapse, there is a tiny gap between the axon terminal of the presynaptic neuron and the membrane of the postsynaptic neuron. This gap is called the synaptic cleft.
The Presynaptic Terminal
Let’s focus on the presynaptic terminal for a moment. The presynaptic terminal contains vesicles. These are membrane-enclosed containers that hold the neurotransmitter. The membrane of the vesicles is the same phospholipid bilayer that comprises the cell membrane.
Calcium Ions Are Important for Vesicular Docking
When a neuron fires, the action potential travels along the axon to the terminal. Once it arrives at the terminal, the depolarization of the membrane opens voltage-gated calcium channels. These are protein channels that are normally closed but will open when the membrane develops a positive charge. Much like the voltage-gated potassium channels. When they are open, voltage-gated calcium channels selectively allow calcium ions, abbreviated as Ca2+ to cross the membrane. We haven’t discussed calcium ions yet but note that calcium ions will have diffusional forces driving them into the axon terminal.
Calcium and Vesicular Docking
Calcium ions act as a signal to the axon terminal and cause the vesicles to move toward the synapse and fuse with the presynaptic membrane. Exactly how calcium mobilizes the vesicles to the synapse is a question that neuroscientists are still trying to figure out. Once the vesicles fuse with the membrane, they release their contents into the synaptic cleft. Watch this animation to see the sequence of events I just described.
Chemical Synapse Animation
Neurons and Their Neurotransmitters
Each neuron contains a specific neurotransmitter that is released by the vesicles at the synapse. We used to believe that each neuron only contained one type of neurotransmitter. For example, if a neuron contained the neurotransmitter serotonin, we would call it a serotonin neuron. We have since discovered that this isn’t quite true, some neurons can contain more that one type of neurotransmitter. However, the convention of calling a neuron by the major type of neurotransmitter it contains still holds true.
Common Neurotransmitters
There are many types of neurotransmitters, hundreds have been discovered so far, but there are six that are very well known and play the biggest role in communication between neurons. Therefore, it is useful that you become familiar with them:
Acetylcholine
Seretonin
Dopamine
Norepinephrine
Glutamate
Gamma-aminobutyric acid (GABA)
Receptors
Once the neurotransmitter is released into the synaptic cleft, it will bind with proteins on the postsynaptic membrane called receptors. By binding to these receptors, the neurotransmitter creates a conformational change in the receptor. That is the neurotransmitter causes the receptor to change its shape.
Excitatory Neurons
Some neurotransmitters, such as glutamate, will cause receptors to open channels that are permeable to sodium ions. As a result, the postsynaptic membrane will become slightly depolarized, and the postsynaptic neuron will be more likely to fire. Thus, neurons that release the neurotransmitter glutamate are called excitatory neurons.
Inhibitory Neurons
On the other hand, the neurotransmitter GABA, which is an acronym for gammaaminobutyric acid, will bind to receptors that open a chloride ion channel. Recall from our earlier lesson that there is a higher concentration of negatively charged chloride ions on the outside of the neuron. Thus, diffusional forces will drive these ions inside the membrane. As chloride ions flow in, the membrane potential will become more negative. When the membrane is more negative than its resting membrane potential, we say it is hyperpolarized. When the membrane is hyperpolarized, it is more difficult for the axon hillock to reach threshold and more difficult for the neuron to fire. Thus, neurons that release the neurotransmitter GABA are said to be inhibitory neurons.
Modulatory Neurons
On a final note, not all neurotransmitters excite or inhibit the postsynaptic membrane. Some act on different types of receptors that can change the way a neuron fires.
Neurotransmitters
Six Well-Known Neurotransmitters
Dopamine
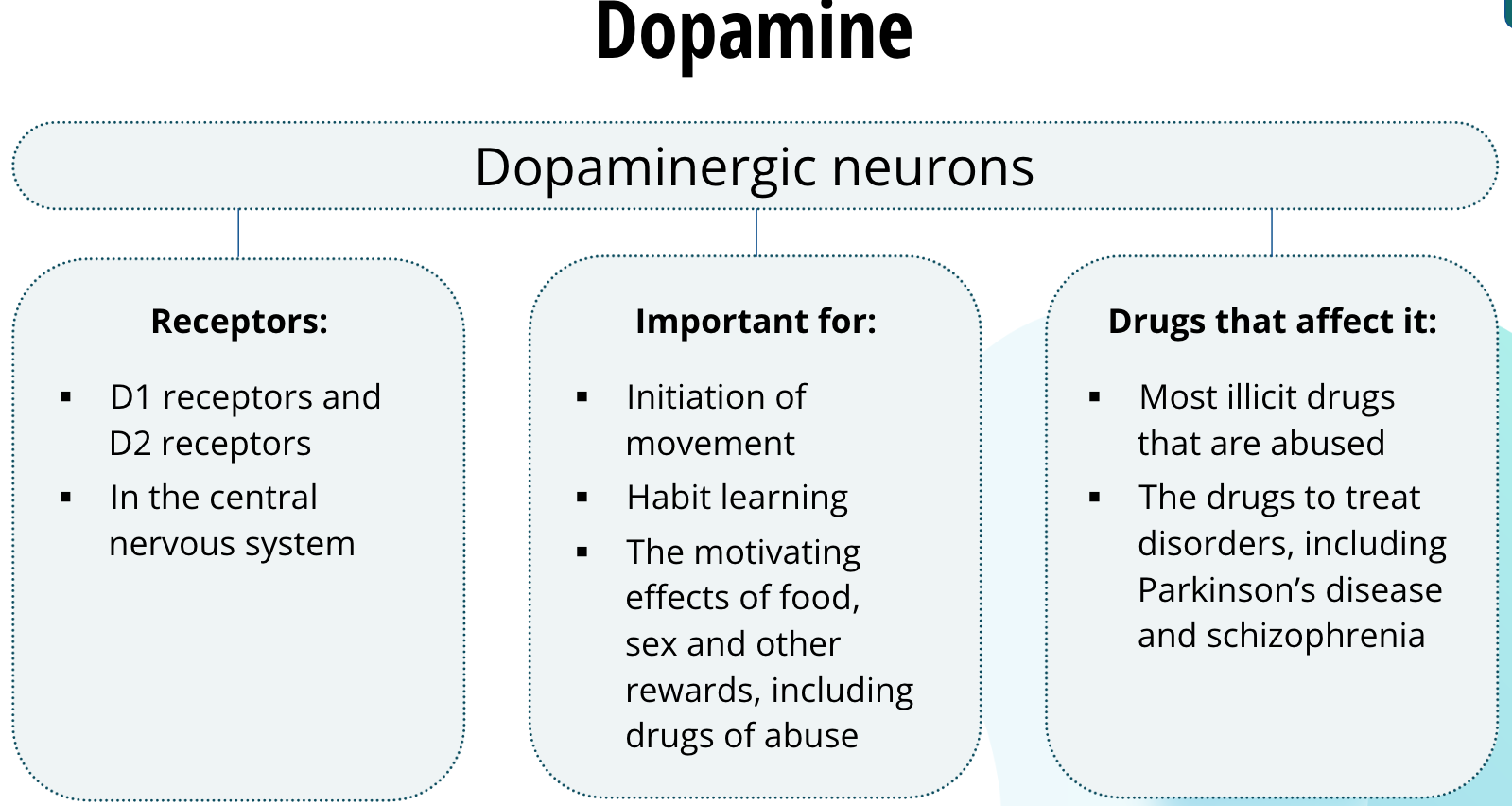
Neurons that release dopamine are called dopaminergic neurons. Dopamine binds to two families of receptors called the D1 receptors and D2 receptors. These receptors are found largely in the central nervous system. Dopaminergic neurons are thought to be important for the initiation of movement, habit learning, and the motivating effects of food, sex, and other rewards, including drugs of abuse. Most illicit drugs that are abused all have in common that they increase dopamine neurotransmission in the brain. Dysregulation of dopamine is thought to underlie several disorders including Parkinson’s disease and schizophrenia. Drugs used to treat these disorders affect dopamine neurotransmission.
Norepinephrine
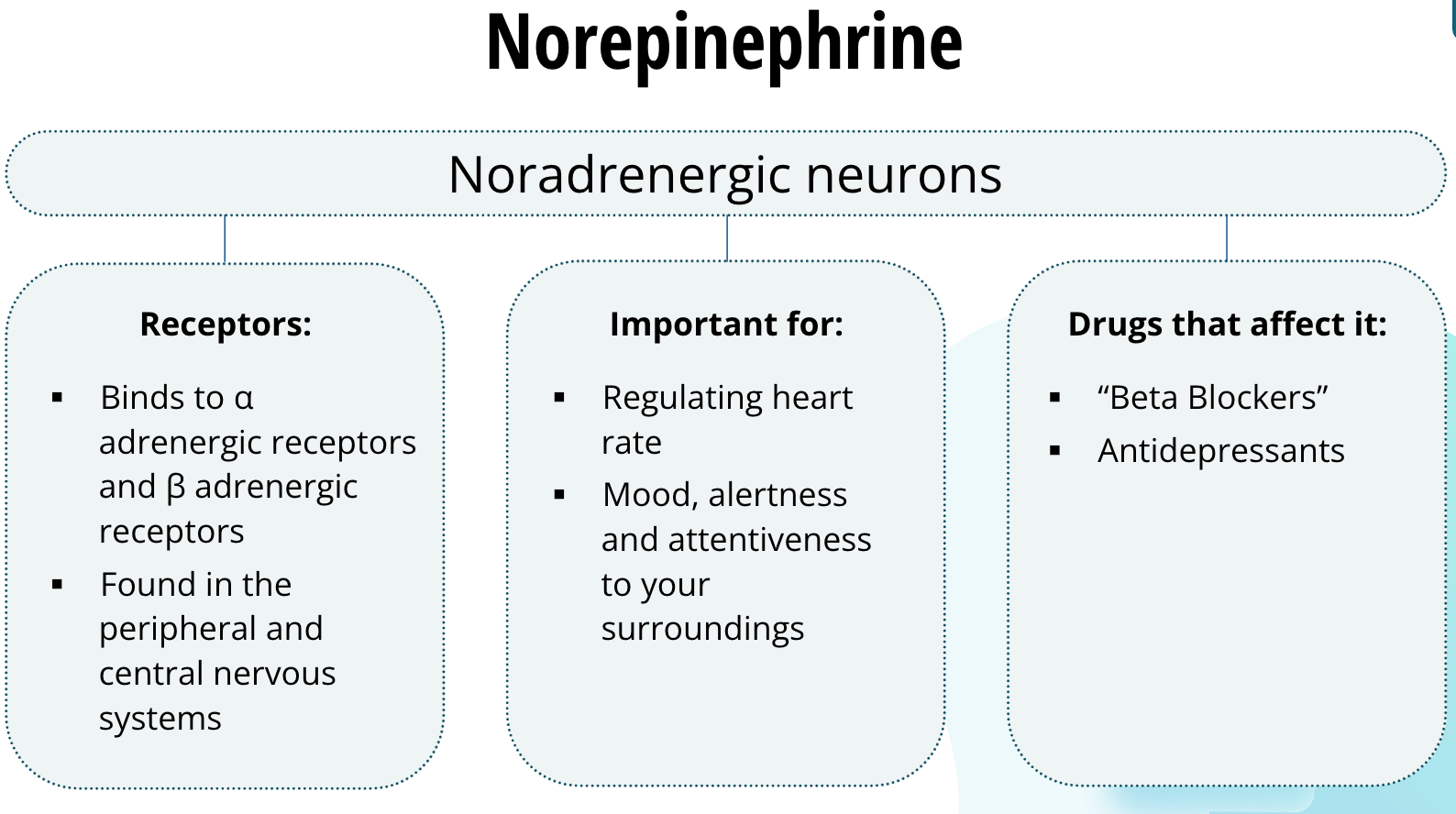
Neurons that release norepinerphrine, which is also called noradrenaline, are called noradrenergic neurons. The precise reason why we have two names for this neurotransmitter is not known — it is just to make the study ofneuroscience that much more complicated, I suppose. Norepinephrine binds to two families of receptors called α adrenergic receptors and β adrenergic receptors. They are found in the peripheral and central nervous systems. In the periphery, they are important for regulating heart rate. In the central nervous system, noradrenergic neurons are thought to be important for mood, alertness, and attentiveness to your surroundings. Patients who have recovered from a heart attack are often prescribed drugs called “Beta Blockers.” These drugs prevent norepinephrine from binding to the β adrenergic receptors and prevent the heart from beating too fast. Some classes of antidepressants also act on the noradrenergic system in the central nervous system.
Glutamate
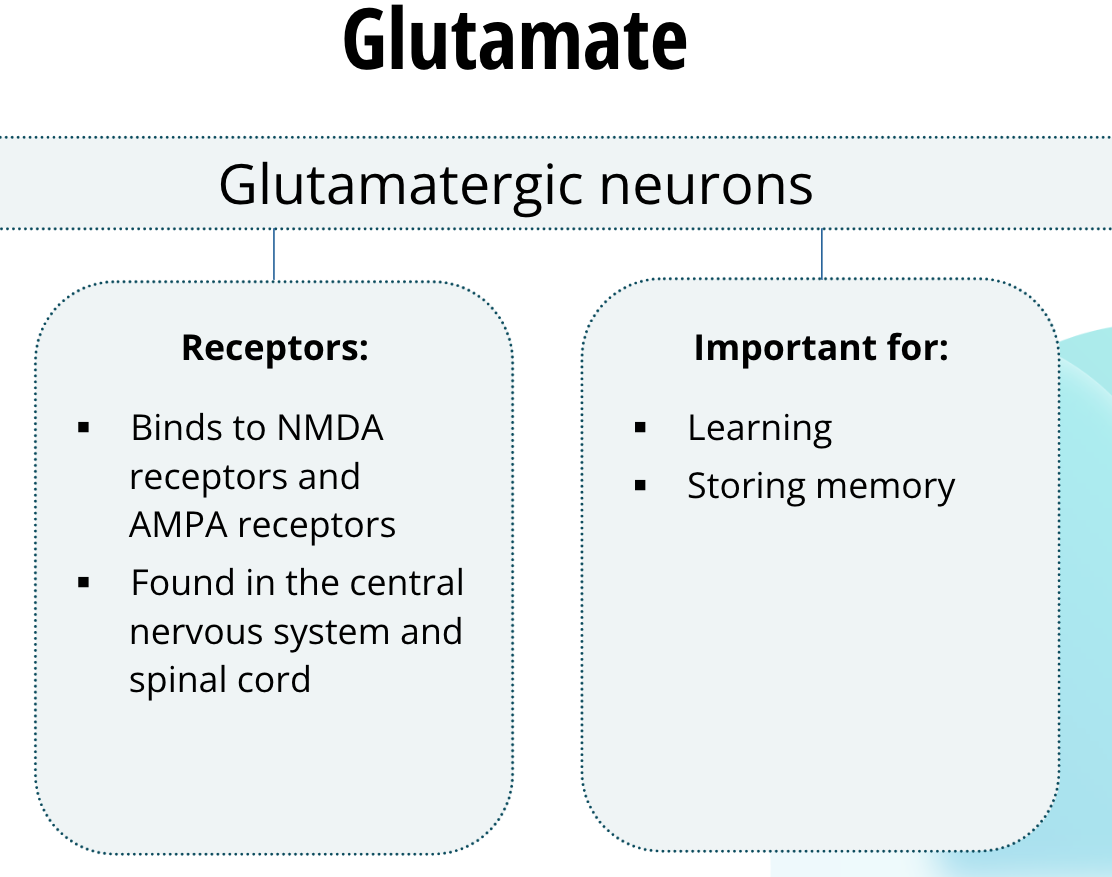
Neurons that release glutamate are called glutamatergic neurons. Glutamate binds to several families of receptors. Two important families are NMDA receptors and AMPA receptors. They are found abundantly throughout the central nervous system and glutamate is the principle excitatory neurotransmitter in the brain, as well as in the spinal cord. These receptors are vitally important for learning and memory. They are involved in neuroplasticity, which is a process by which pathways in the brain, especially at the synapse, are able to change or reorganize. Neuroplasticity is thought to be one way the brain learns and stores memories.
Serotonin
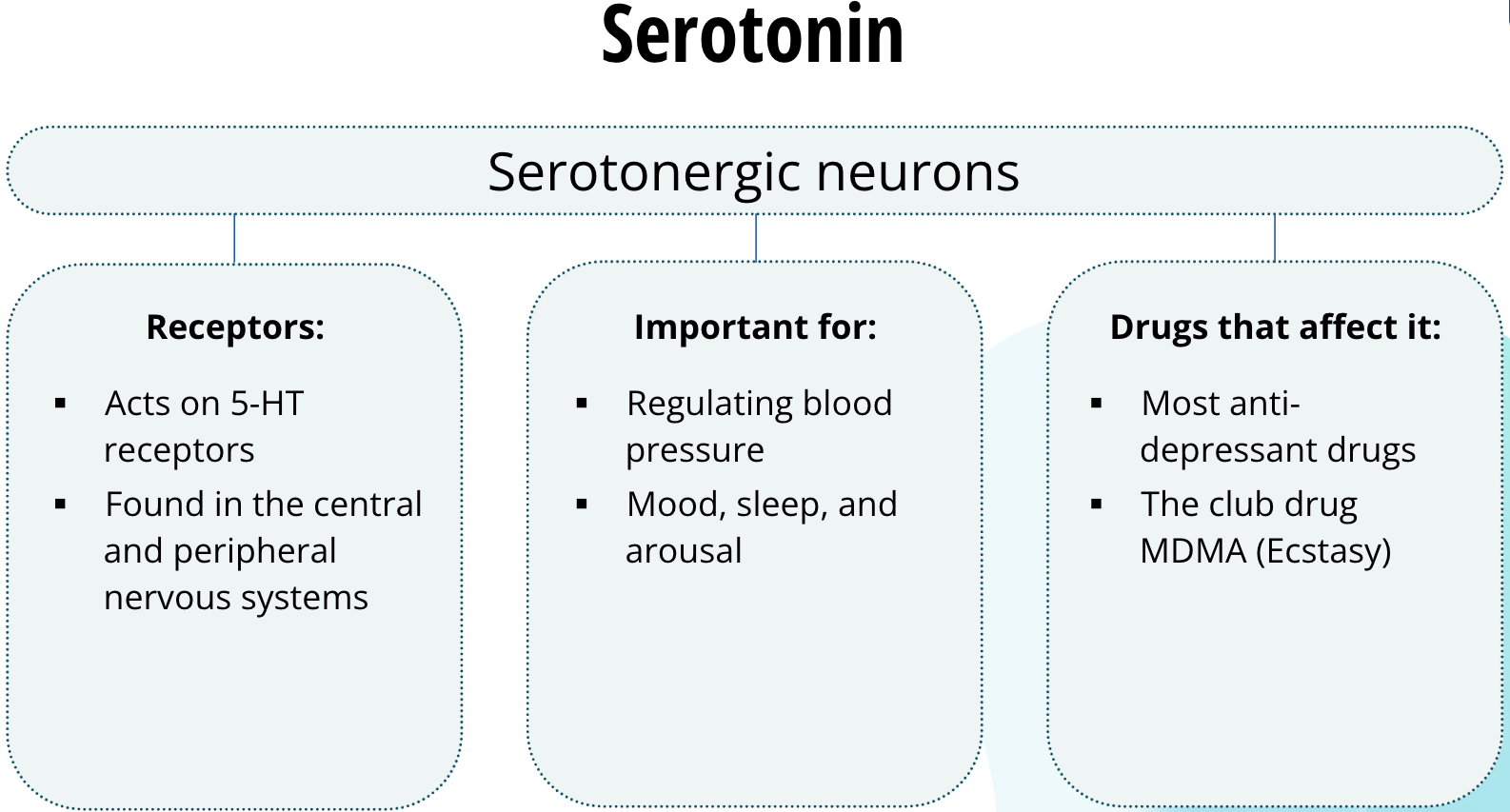
Neurons that release serotonin are called serotonergic neurons. Serotonin acts on a family of receptors called 5-HT receptors. These receptors are found in the central and peripheral nervous systems. In the peripheral nervous system, they are important for regulating blood pressure. In the central nervous system, serotonergic neurons are important for mood, sleep, and arousal. Most anti-depressant drugs act on serotonergic neurons to regulate serotonin neurotransmission, the dysregulation of which is thought to play a role in major depression. The club drug, MDMA, commonly called ecstasy, leads to increased serotonin in the synapse. It is believed that this increase in serotonergic neurotransmission relates to the drug’s mood elevating effects.
Gamma-Aminobutyric Acid (GABA)
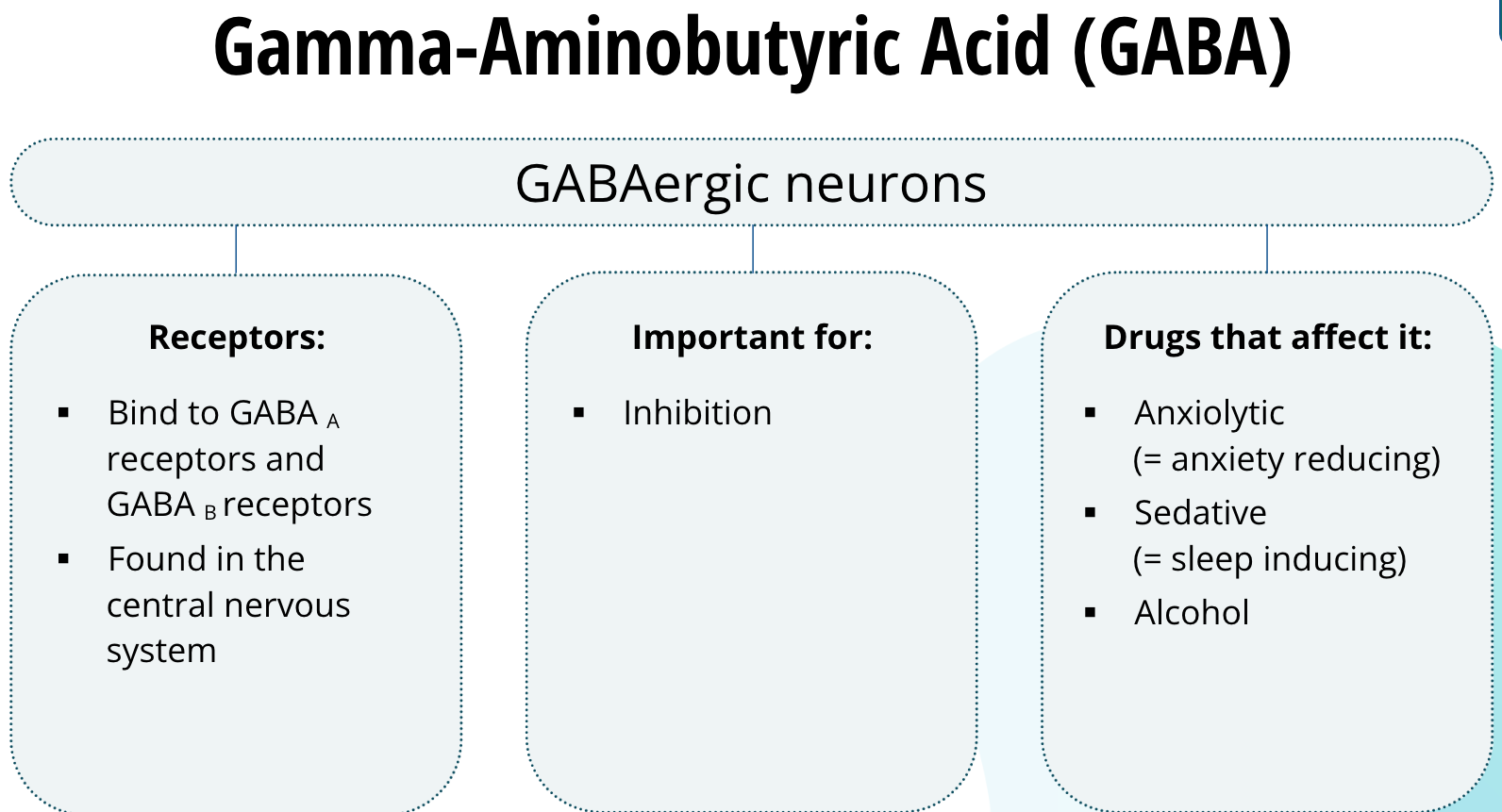
Neurons that release GABA are called GABAergic neurons. GABA binds to two main types of receptors called GABAA receptors and GABAB receptors. They are found abundantly in the central nervous system. GABA is the predominant inhibitory neurotransmitter in the brain and spinal cord. Anxiolytic (meaning anxiety reducing) and sedative (meaning sleep inducing) drugs bind to GABA receptors. Alcohol is also thought to affect the GABA receptors.
Acetylcholine
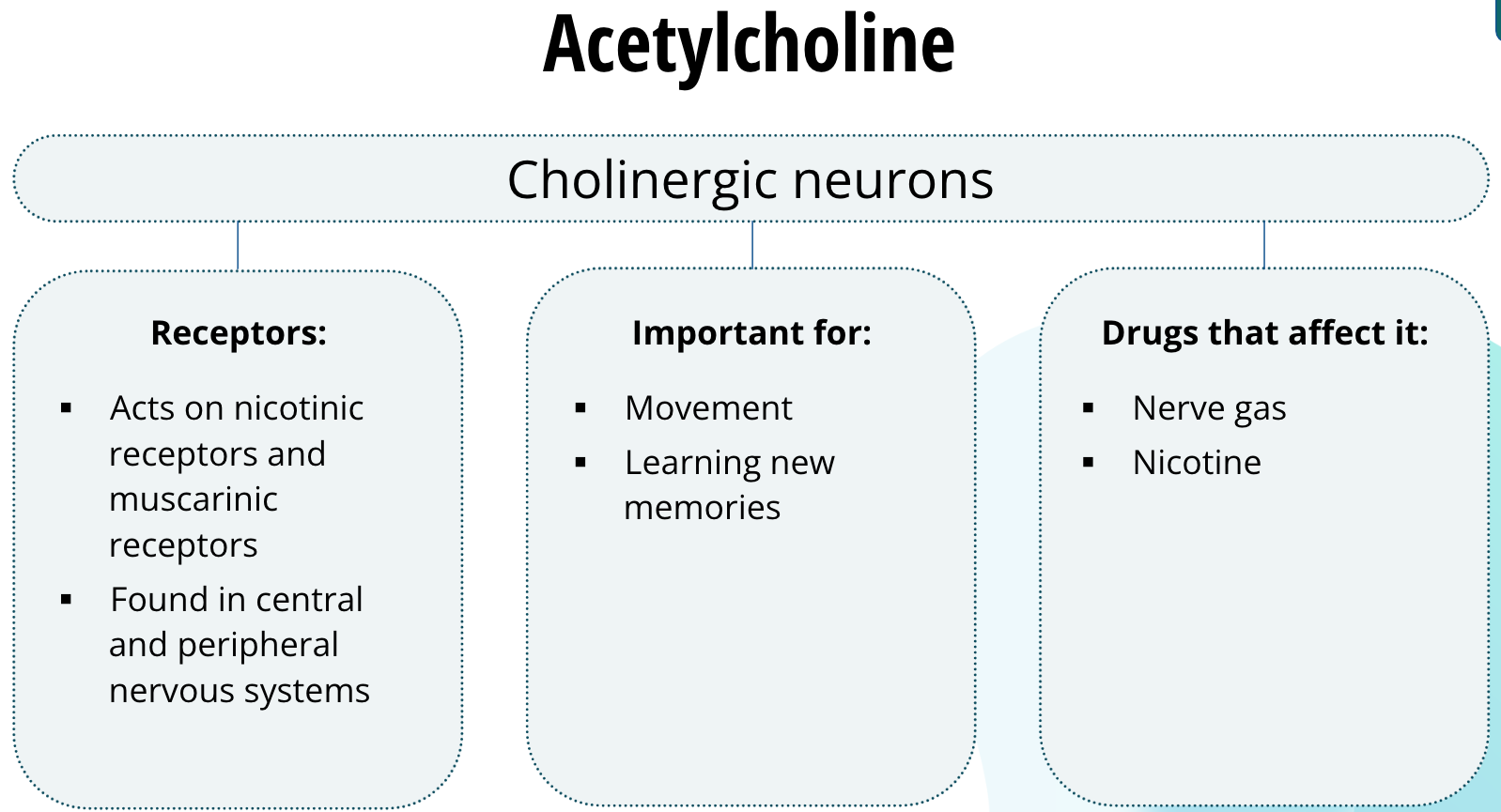
Neurons that release acetylcholine are called cholinergic neurons. Acetylcholine acts on two families of receptors: nicotinic receptors and muscarinic receptors. These receptors are found both in the central and peripheral nervous systems. In the peripheral nervous system, acetylcholine is the neurotransmitter responsible for signalling the muscles to contract. Thus, cholinergic neurons are important for movement. In the central nervous system, cholinergic neurons in the hippocampus are important for learning new memories. Drugs that act at cholinergic synapses in the peripheral nervous system include nerve gases. Nerve gas can block cholinergic neurotransmission to the muscles of the diaphragm and shut down the ability to breathe, causing the victim to suffocate. Another drug that acts on the cholinergic system is nicotine, yes, the same drug found in tobacco. Nicotine binds to nicotinic receptors and enhances cholinergic neurotransmission. It has been shown that nicotine can improve memory function, but that does not mean you should pick up smoking any time soon.
Receptors
Receptor Overview
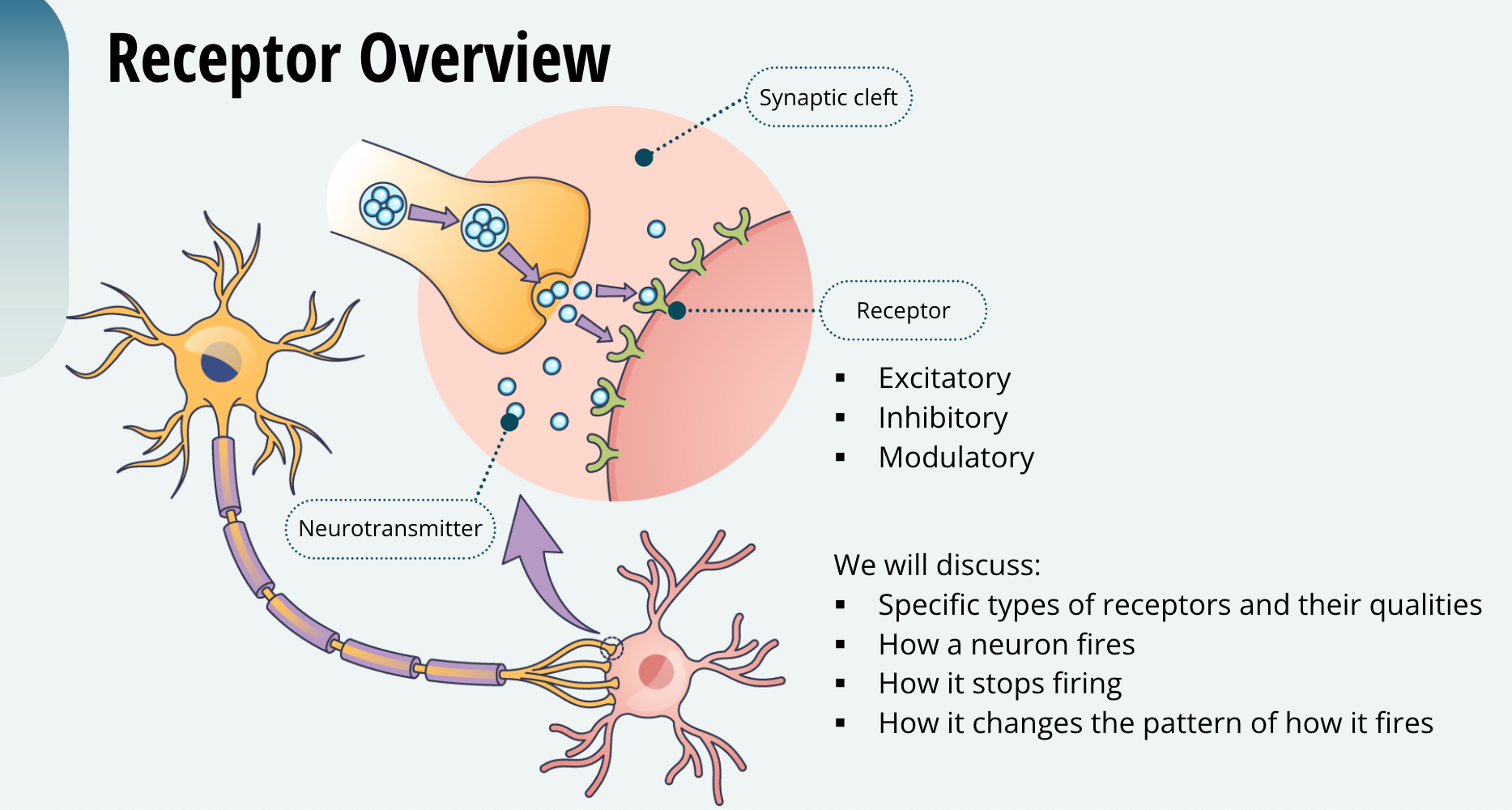
As you learned previously, once the neurotransmitter is released into the synaptic cleft, it will typically bind to a receptor. You also learned that receptors can be excitatory, inhibitory, or modulatory. Next, we will discuss specific types of receptors and the qualities they possess to make a neuron fire, make it stop firing, or change the pattern of how it fires. As you have probably figured out by now, the timing of neuronal firing is how the brain produces behaviour and cognition. So, receptors are very important.
Neurotransmitters are Ligands
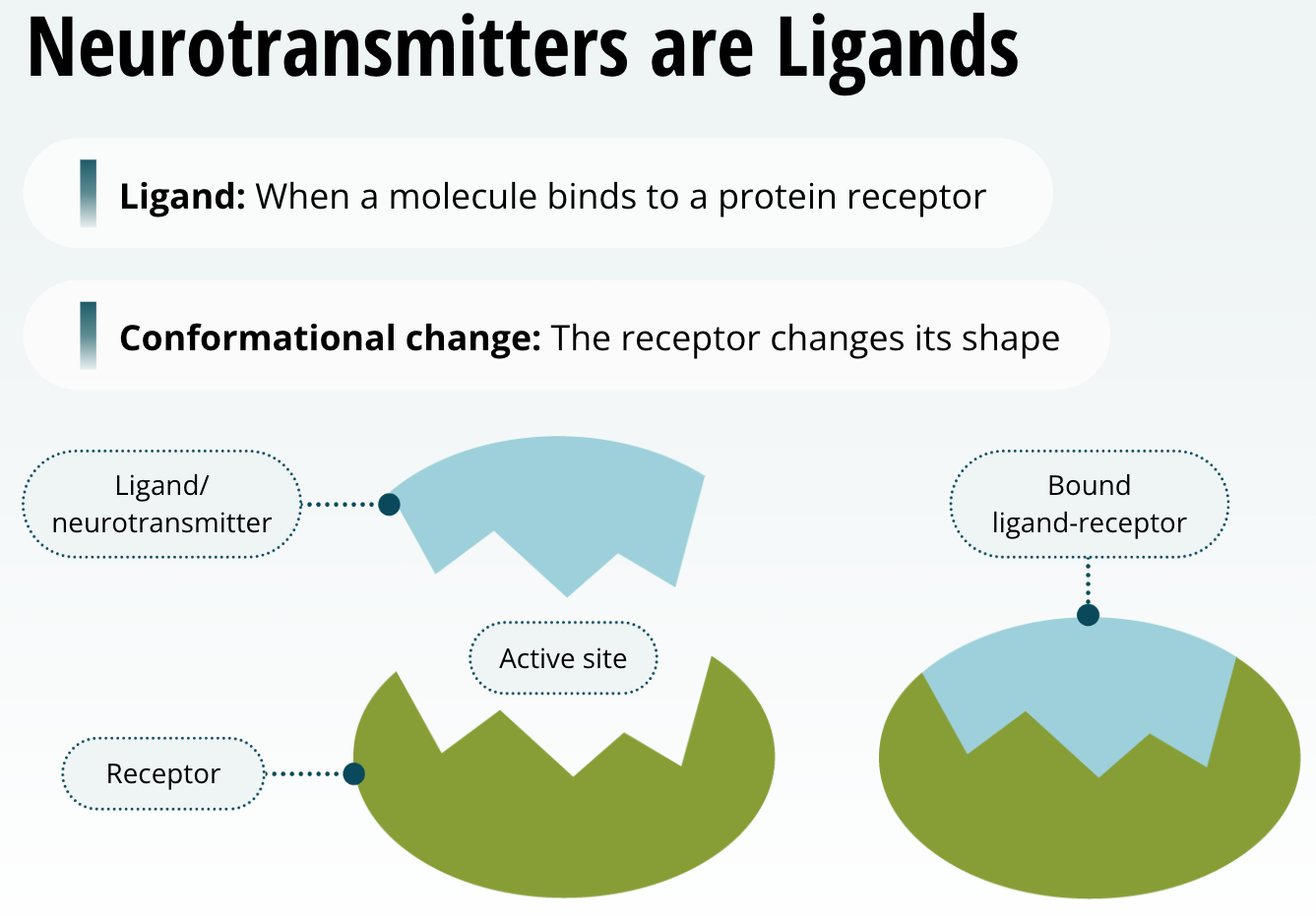
When a molecule binds to a protein receptor, we refer to it as a ligand. When a ligand binds to a receptor, it causes a change in the structure of the protein receptor, or as we say, it causes a conformational change. That means that this causes the receptor to change its shape. So, when a neurotransmitter is released into the synaptic cleft and binds to a receptor, it acts as a ligand. Neurotransmitters are in fact ligands when they bind to their receptors. Just remember for the purposes of this course, neurotransmitter = ligand.
The Relationship between a Ligand and its Receptor
The relationship between a ligand and its receptor is much like a lock and key. Only very specific shapes will bind to, or fit, the protein receptors. In other words, each neurotransmitter has a specific protein that it binds to preferably. For example, dopamine preferably binds to dopamine receptors, and glutamate preferably binds to glutamate receptors.
Receptor Types
Ligand-Gated Ion Channels (LGIC)
G-protein-coupled Receptors (GPCR)
There are many receptor types on and within cells of the body. However, for the purposes of this course we will focus on two types that are very important for neurotransmission, that is, neuronal communication.
The two types of receptors we will be learning about are: 1) Ligand-gated ion channels (abbreviated as LGIC’s) which are also called ionotropic receptors. 2) G-protein-coupled receptors (abbreviated as GPCR’s) which are also referred to as metabotropic receptors. Of these two types, the LGICs are the fastest acting and the GPCRs are slower to cause a change in neuronal firing. As you learn about them, think about why that is the case.
LGIC Receptors
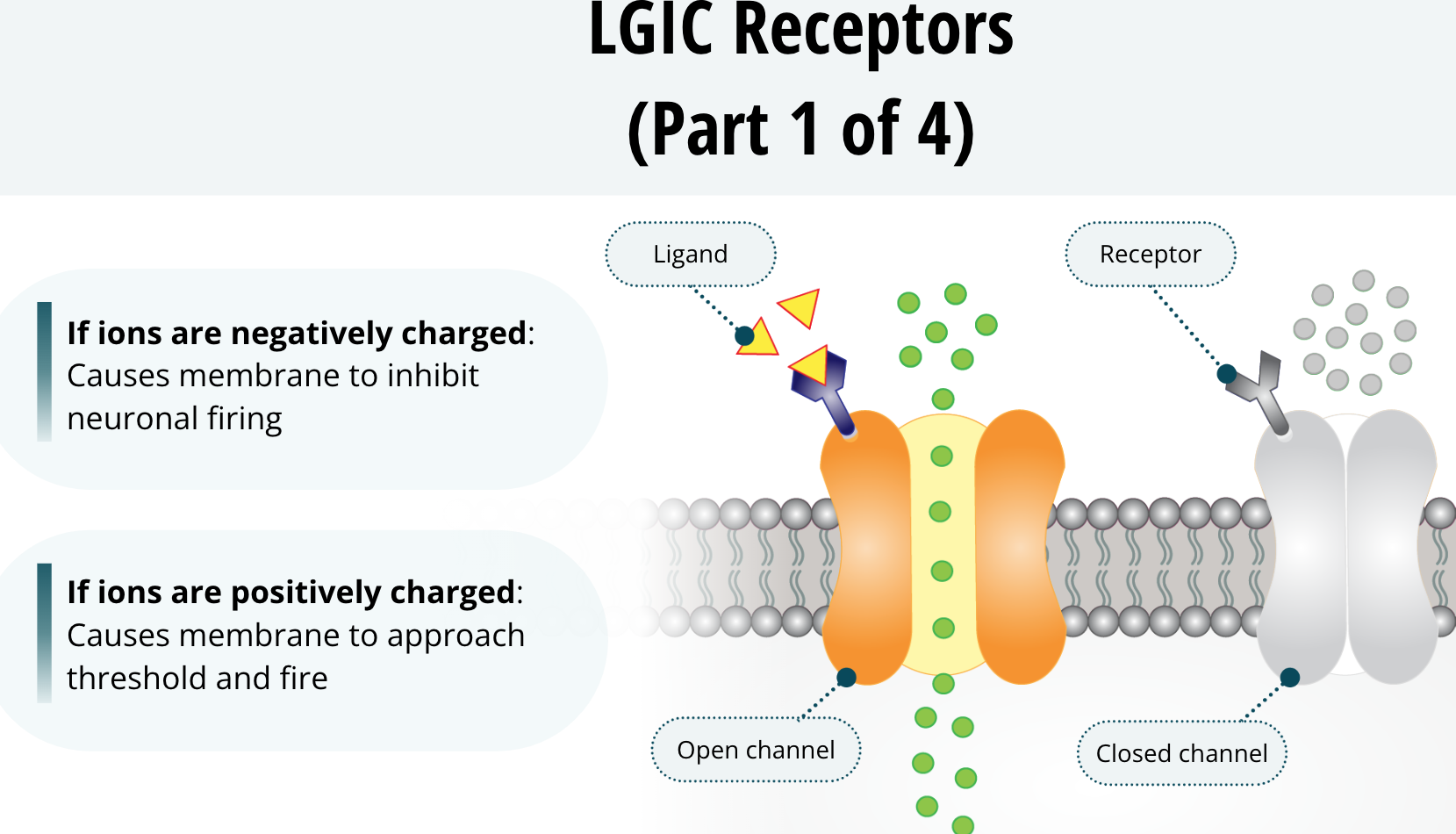
These receptors span the phospholipid bilayer of the neuronal membrane. When they are open, they allow ions to pass though the membrane. Think about how the sodium and potassium channels allow ions to pass through the membrane. These receptors are very similar to those ion channels, except, they can open or close depending on whether a ligand binds to them. Once the ligand does bind, the receptor undergoes a conformational change that opens the channel. This channel will allow select ions to flow across the membrane based upon their electrostatic and diffusional forces. Ultimately, it can cause the membrane to be more negatively charged and inhibit neuronal firing. Conversely, this could cause the membrane to be more positively charged toward threshold and fire.
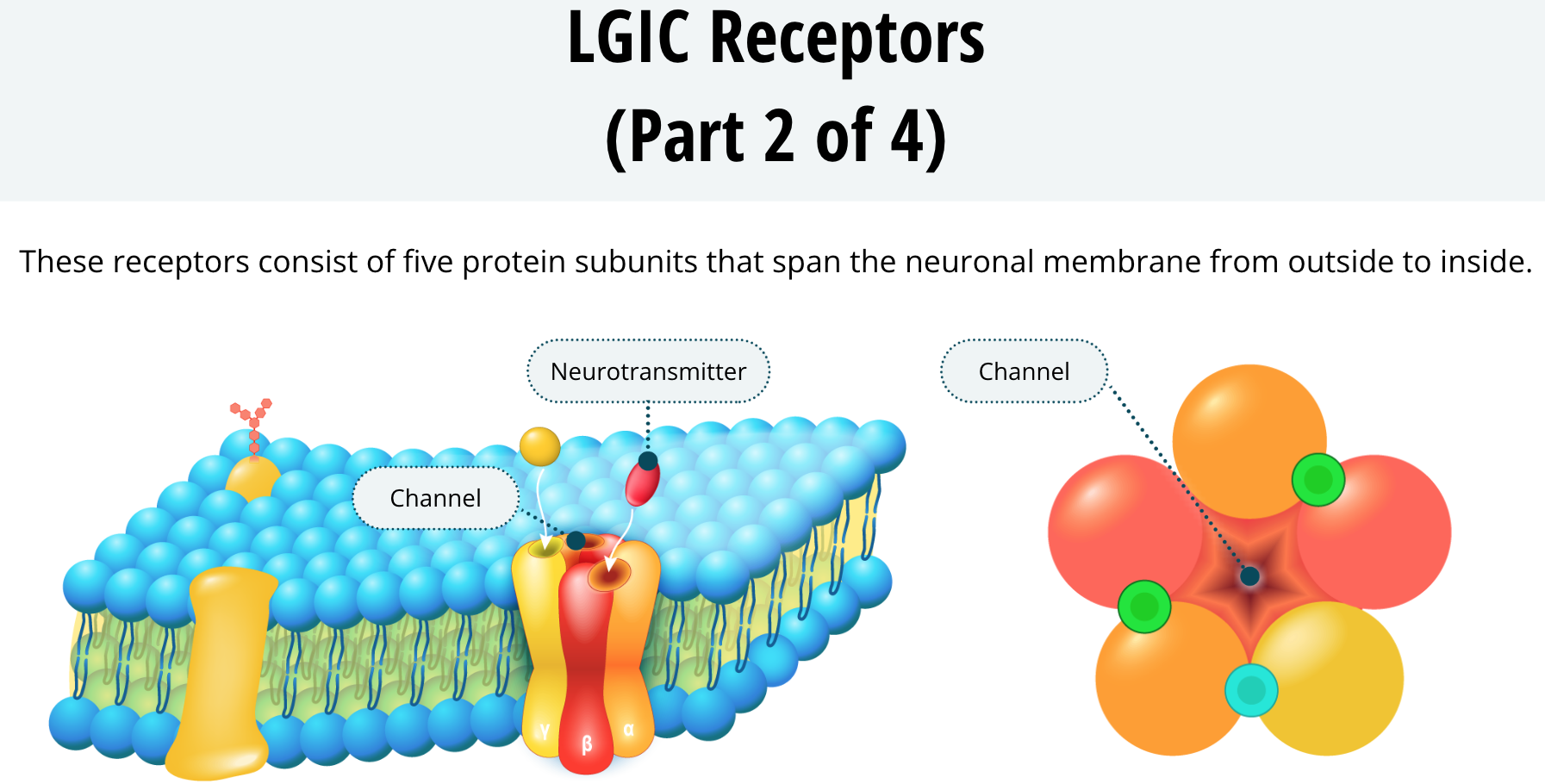
Something all LGIC receptors have in common is that they consist of five protein subunits that span the neuronal membrane from outside to inside. The five subunits form a circle with a channel in the middle.
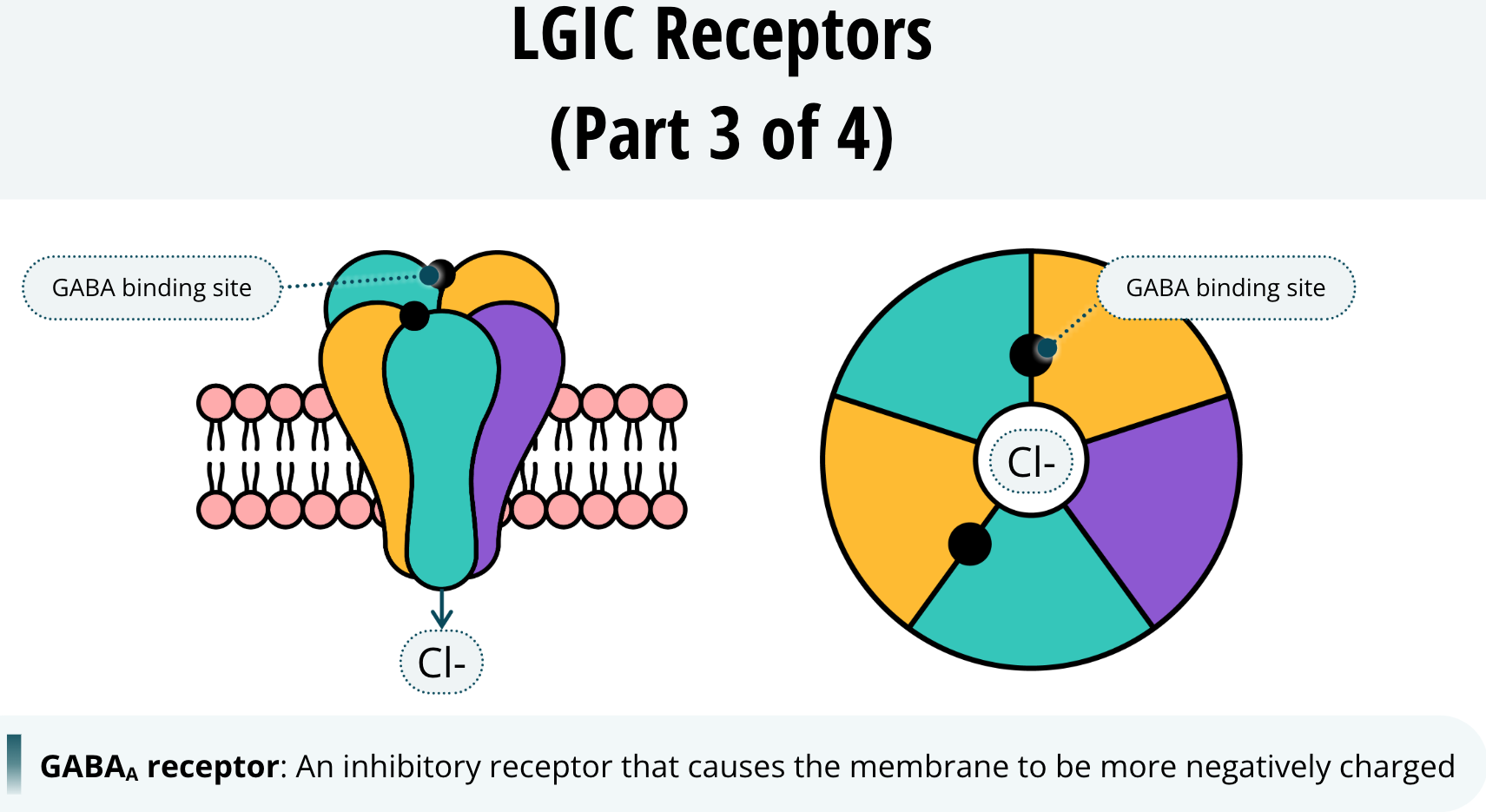
An example of a receptor that, when open, causes the membrane to be more negatively charged is the GABAA receptor. When this receptor is activated, its channel allows chloride ions to cross the membrane into the neuron. As these negatively charged ions enter the neuron, it maintains its negative charge making it more difficult to reach threshold and fire. This is why we say GABAA is an inhibitory receptor. When GABA binds to GABAA receptors, it inhibits the neuron from firing.
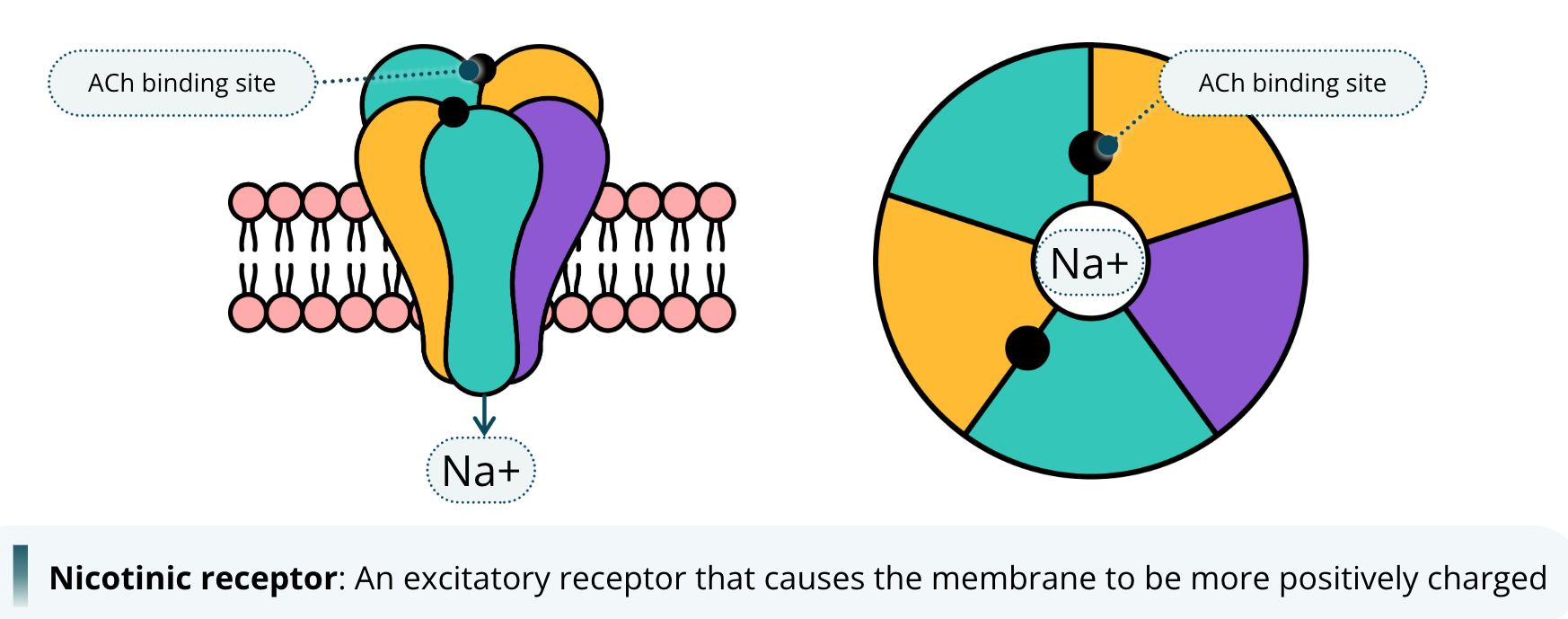
An example of a receptor that causes the membrane to be more positively charged when it is open is the nicotinic receptor. This receptor is bound by the neurotransmitter acetylcholine, abbreviated as ACh. We call it the nicotinic receptor because it is the acetylcholine receptor that also potentially binds to the drug nicotine. Yes, the same nicotine found in tobacco products. When acetylcholine binds to the nicotinic receptor, the channel opens and allows sodium ions to cross the membrane. The influx of sodium ions into the neuron causes the membrane to become more positively charged, and has the potential to depolarize the neuron, causing it to fire. For this reason, we say that nicotinic receptors are excitatory.
GPCR Receptors
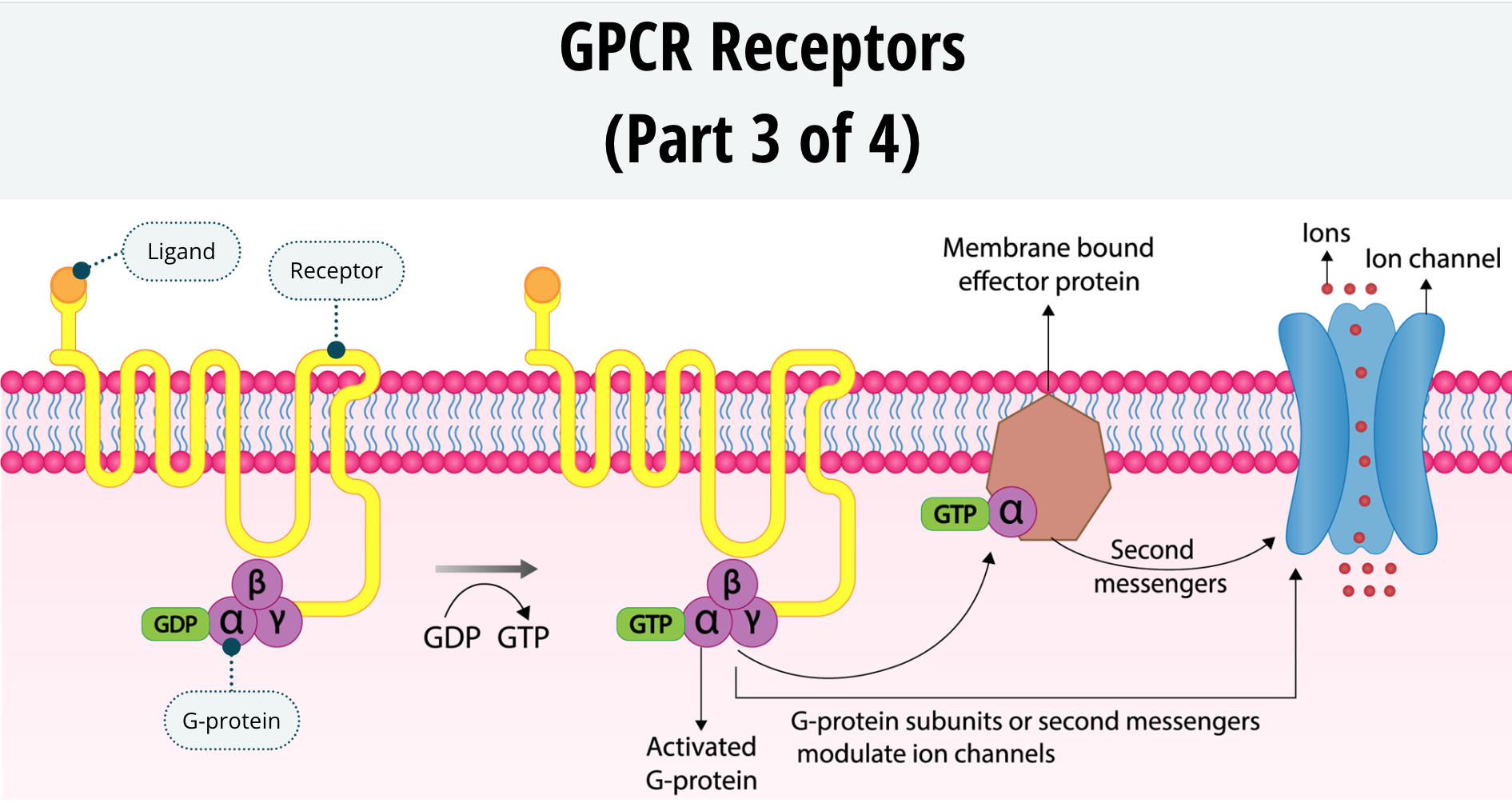
The next category of receptors are the G-protein-coupled receptors, or GPCRs. These receptors have seven transmembrane domains. That is just another way of saying that the protein receptor has seven sections that cross the neuronal membrane back and forth. These receptors have an N terminus on the outside of the neuron and the C terminus on the inside. The ligand binds to the N terminus and causes a conformational change in the receptor resulting in the C terminus interacting with a G-protein.
G-proteins are so called because they bind to two types of nucleotides starting with the letter G, called guanosine diphosphate, or GDP, and guanosine triphosphate, or GTP. The G-protein itself is composed of three proteins labelled alpha, beta, and gamma. The G-protein sits just inside the neuronal membrane and, when quiescent, is bound to GDP.
Once the GPCR is activated by a ligand, it will interact with the G-protein. Activation of the G-protein will cause it to decouple from the GDP and take on a GTP. Once the GTP is bound to the G-protein, the protein itself splits into two sections. The alpha subunit decouples and acts as a signalling mechanism within the neuron. Additionally, the beta and gamma subunits stay coupled to each other but also act as a signalling mechanism. The alpha subunit and the beta/gamma subunits of the G-protein can then cause longterm cellular changes within the neuron by either activating second messengers or modulating protein channels. You will learn more about second messengers later.
After a while, the GTP will lose a phosphate group and revert back to GDP. Once this happens the alpha and beta/gamma subunits will re-couple, the G-protein will once again become quiescent as the system resets until the next time the GPCR is activated by a ligand.
Differences Between LGICs and GPCRs
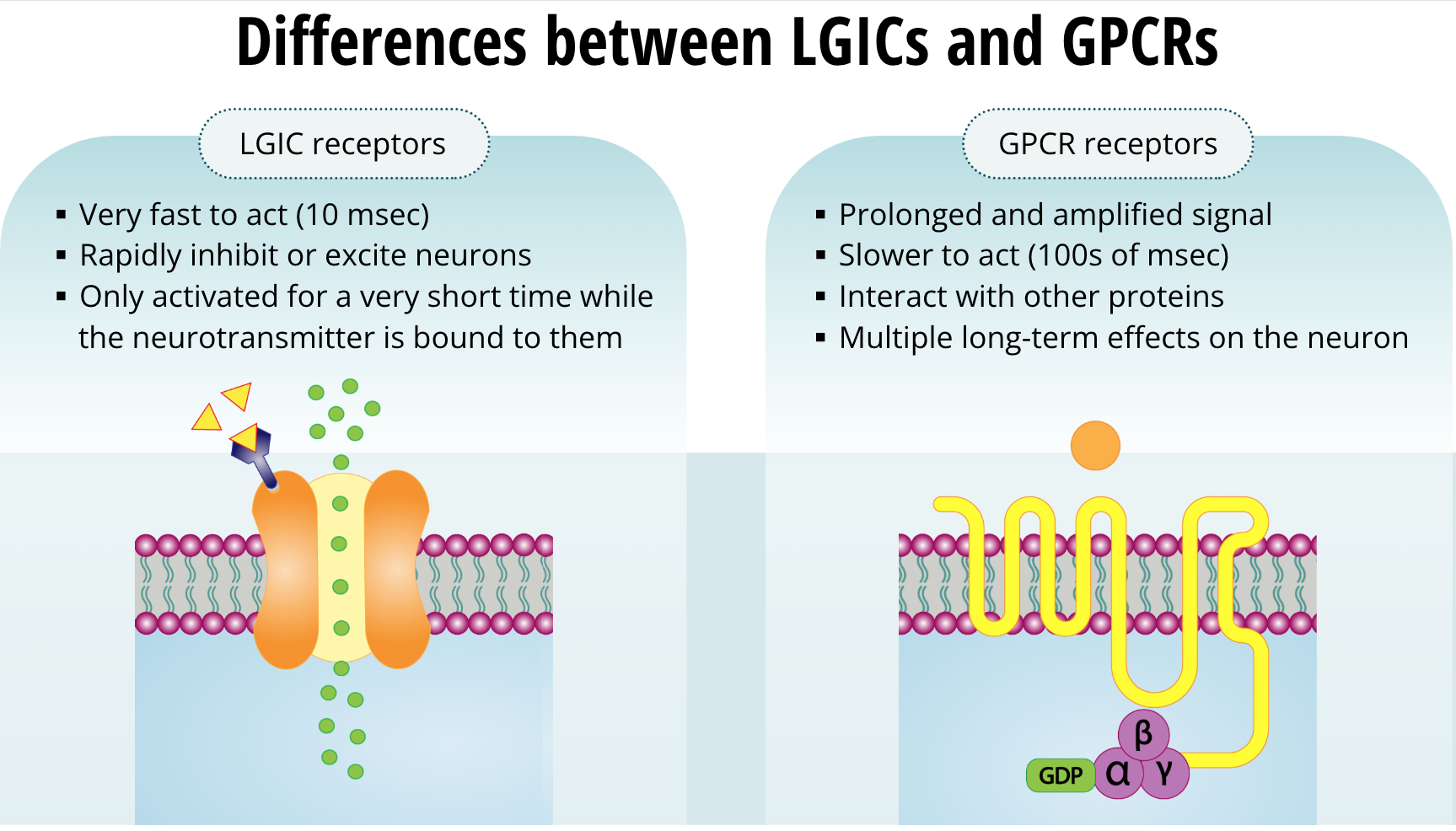
It appears that mother nature developed these two types of receptors for different functions. The LGIC receptors are very fast to act. They can act as fast as a 10 msec. They can rapidly inhibit or excite neurons. However, the drawback is that they are only activated for a very short time while the neurotransmitter is bound to them. Once the neurotransmitter decouples, the receptor actions on the neuron are finished. On the other hand, with GPCR receptors, the signal is both prolonged and amplified. The actions of these receptors are slower (at about 100s of msec) because they must interact with other proteins. Once they do, they have a bigger impact on the neuron. With these types of receptors, the actions last long after the neurotransmitter is no longer bound. Because these receptors interact with other proteins, and those interact with yet other proteins, the signal lasts a much longer time and the effects of one molecule of the neurotransmitter can have multiple long-term effects on the neuron. We will learn about those next.
Second Messengers
Second Messengers
You have learned about two types of receptors, the LGICs and the GPCRs. You also learned that GPCRs interact with other proteins to affect cellular responses. One way to think about neurotransmission is that the neurotransmitter, or ligand, itself is a first messenger, and the intracellular proteins that are affected by GPCRs are second messengers. Second messengers are small molecules and ions that relay signals received by the cell-surface receptors to effector proteins inside the cell. They broadcast the first message relayed by the GPCR. Second messengers are typically present at low concentrations in resting cells and can be rapidly produced or released when cells are stimulated.
Types of Second Messengers
Next, we will explore four types second messengers: cyclic adenosine monophosphate (cAMP), inositol triphosphate (IP3), diacylglycerol (DAG), and calcium ions.
Cyclic Adenosine Monophosphate (cAMP)
cAMP is an acronym for cyclic adenosine monophosphate. This molecule is hydrophilic (i.e. water loving) and an important second messenger. When certain g-proteins are activated, they can change the activity of an enzyme called adenylate cyclase (which is also called adenylyl cyclase, either name is acceptable). This enzyme catalyzes adenosine triphosphate (ATP) into cyclic AMP.
Adenosine Triphosphate (ATP)
ATP (or adenosine triphosphate) is a nucleotide that is important for providing energy for intracellular processes. It is essentially a form of cellular energy, and it is synthesized in the mitochondria.
cAMP Synthesis
Aside from its role in providing cellular energy, when ATP is converted to cAMP it becomes a second messenger. Many molecules of ATP are converted to cAMP when the GPCR is activated, thus this is a good example of signal amplification. Once the cAMP is synthesized, it goes on to activate protein kinases and specifically a kinase called protein kinase A (PKA).
Protein Kinases
Protein kinases are proteins that activate other proteins by adding a phosphate group to them. A phosphate group is characterized by a phosphorus atom bound to four oxygen atoms (denoted as PO4). When a kinase adds a phosphate group to another protein it is called phosphorylation. Phosphorylation is an important way to modify protein function within the cell. Some examples of how phosphorylation alter protein function are: opening or closing an ion channel, altering the activity of an enzyme, or even changing the stability of certain proteins. It is important to note that these events are reversible. That is, like other animal cells, neurons contain protein phosphatases. This enzyme has the opposite effect of protein kinases, insofar that it travels within the cell to remove phosphate groups from proteins - or dephosphorylate them. Thus, phosphorylation of proteins via the activation of kinases by cAMP is an important but temporary event.
cAMP Signaling
A final note on cAMP signalling. When a GPCR is activated, if levels of cAMP increase within the cell, we call that receptor a Gs GPCR. The S stands for stimulatory as in the GPCR stimulates adenylate cyclase to synthesize more cAMP. In some cases, the GPCR causes levels of cAMP to drop. In these cases, the receptor is called a Gi GPCR. The i in this case stands for inhibitory, meaning it inhibits adenylate cyclase. When scientists were first investigating GPCRs, they first discovered their effects on cAMP, either increasing or decreasing it. Hence, the terms Gs and Gi GPCRs. Eventually, they discovered other GPCRs that did not have any effect on cAMP, suggesting that there are other second messengers. For those types of GPCRs they simply call them G0 indicating that they have no effect on cAMP.
Inositol Triphosphate (IP) and Diacylglycerol (DAG)
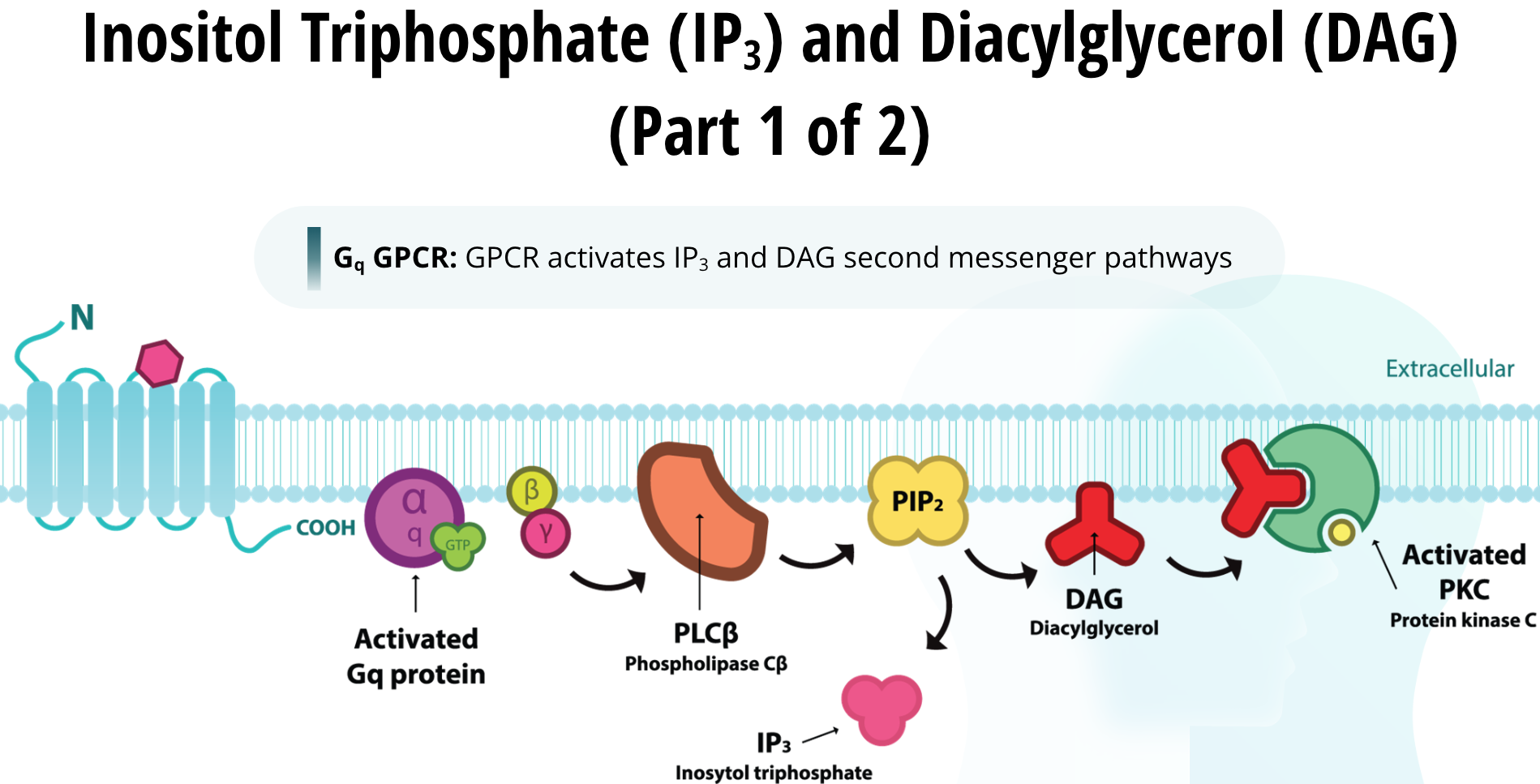
When a GPCR activates the inositol triphosphate (IP3) and diacylglycerol (DAG) second messenger pathways, it is referred to as a Gq GPCR. When a Gq GPCR is activated by its neurotransmitter the beta/gamma subunits of the G-protein activate phospholipase C (PLC). PLC is a class of membrane-linked enzymes that chop up phospholipids. One particular phospholipid is PIP2 (which is an acronym for Phosphatidylinositol 4,5- bisphosphate). It is positioned within the cell membrane. Once PLC is activated by the G-protein, it cleaves PIP2 into two separate molecules called DAG and IP3. DAG remains lipophilic and stays within the fatty cell membrane. DAG is capable of activating a specific protein kinase called protein kinase C (PKC), which in turn produces its cellular effects by phosphorylating other proteins.
The other half of PIP2 is called IP3 and it is hydrophilic. Thus, it leaves the membrane to enter the cytosol. IP3 relocates to the endoplasmic reticulum. There it binds to a receptor called the IP3 mediated calcium channel. The endoplasmic reticulum is a network of membrane enclosed tubules within the cell. It contains a high concentration of calcium ions compared to the cytosol. When calcium ion channels are open, calcium ions will flow out from the endoplasmic reticulum into the cytosol due to diffusional forces.
Calcium Ions (Ca2+)
The fourth second messenger is the calcium ion itself, abbreviated as Ca2+. These ions bind to another molecule called calmodulin. Once calcium binds to its substrate calmodulin (CaM) the complex is able to activate other proteins within the cell. Once such protein is CaMKII, calcium calmodulin kinase two. This protein kinase, like all others, phosphorylates proteins within the cell and has been found to be particularly important for memory formation in the brain. Because intracellular levels of calcium ions are quite low, these ions will flow into the cell if a calcium channel is opened on the cell membrane or on the endoplasmic reticulum.