lecture 6: single cell plankton (marine communities)
lab report for the field course
will be put in canvas
plankton diversity
Plankton communities are taxonomically and functionally diverse
definition of plankton →
they drift
cannot actively swim against currents
organisms that can be pluricellular to unicellular
plankton diverity can be found across several lineages form viruses to bacteria/archaea and eukaryotes, both unicellular (protists/micro-eukaryotes) and multicellular
size range:
picoplankton
nanoplankton
microplankton
mesoplankton
macroplankton
megaplankton
today only unicellular eukaryotes
size-based framework
plankton size classes
Plankton classification was ecologically inadequate
The traditional division of plankton is overly simplistic and taxonomically inconsistent:
bacterioplankton
phytoplankton
zooplankton
It blurs important ecological distinctions like mode of nutrition, reproductive rate, and trophic role
so instead of the above classification, it was changed to different terminology:
viroplankton
bacterioplankton
mycoplankton
hytoplankton
protozooplankton
metazooplankton
nekton
then there is the other temrinlogoy for plankton (look at the diagram in teh slide)
femtoplankton
picoplankton (<2 (3) µm)
nanoplankton (…)
microplankton
mesoplankton
macroplankton
megaplankton
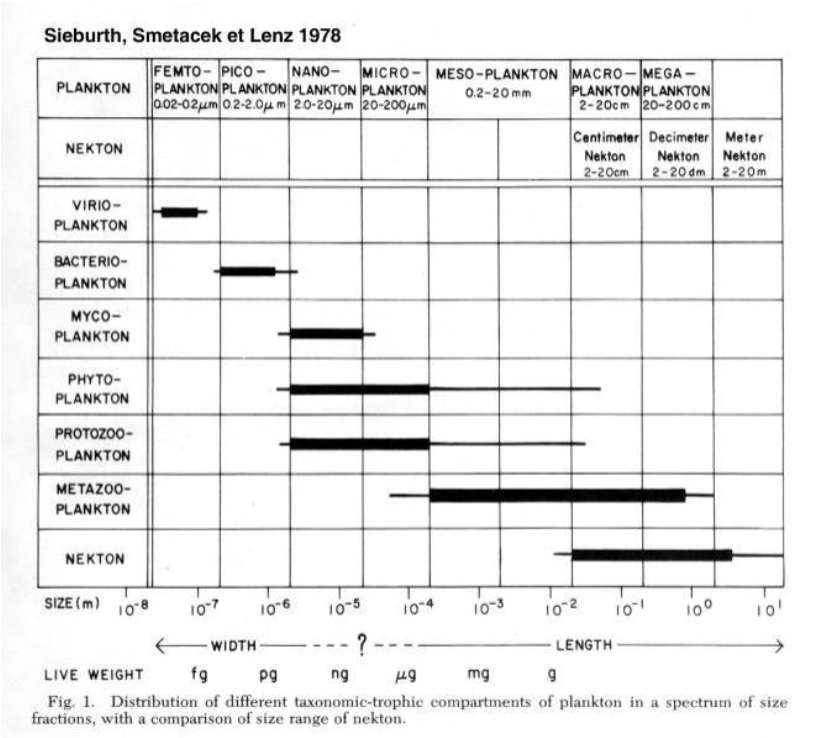
This is because it makes it easy to classify them according to the amount of energy they consume
emphasis on size-based calssification
advocates for organising plankton by size fractions (femto-, pico, nano, micro, meso, macro, mega) to align with:
sampling methods
metabolic scaling
trophic interactions
plankton size range and available optical imaging methods
flow cytometry mostly used for pico and nano plankton (0.2-20µm)
insitu imaging equipment
flow cam
different types of flow cams
insitu imaging equipment
50 - 200 µm
zoo scan
200 - 2000 µm
plankton size range and sampling methods
The difference between using a wp2 net and a phytoplankton net is becasue the amount of water samples is more using a wp2 net than with a phytoplankton net
functional diversity
plankton communities are functionally diverse
phytoplankton →
photosynthetic prokaryotes (cyanobacteria)
microbial eukaryotes
several lineages
zooplankton →
heterotrophic uni and multicellular eukaryotes
protozooplankton and metazooplankton
mixoplankton →
microbial eukaryotes exhibiting a combination of autotrophy and heterotrophy
planktonic protist functional groups based on mechanisms for energy and nutrient acquisition
phago-heterotroph
photo-autotroph
constitutive mixotroph
behaves like a consumer
generalist non constitutive mixotroph
eats anything
but when it eats an algae, it keeps teh chloroplast temporarily and photosynthesizes for that time period
endosymbiotic specialist non-constitutive mixotroph
plastidic specialist non constitutive mixotroph
marine planktonic protists
protists = single celled eukaryotes
protists are more abundant than all animals together
planktonic protists are hte engine of marine ecosystems
eukaryotes have invested in cell forms, apparatus and lifestyle
plasmodium and toxoplasma have evolved from free living algae
apicomplexans harbour a unique organelle called Apicoplast which is a non-photosynthetic chloroplast
ciliates and the algae symbiodinium are genetically closely related
they belong to the alveolata, a very diverse group of protists
they are from a diverse group that evolved from algae
eukaryote phylogeny before the molecular era - a brief history
ernst haeckel: the plankton artist
haeckel - general morphology of the organism
enrst haechel believed that the natural system proposed darwim (1895), should be represented as a geanological tree
haeckel’s book provided major imporvements to teh theory of descent, including:
a large vocabulary of neologisms, some of which became succesful, such as phylogeny, monophyletic, and polyphyletic
the term protists (the first of all or primordial) to distinguish unicellular organisms
haeckel - pedigree of man
metazoa - today use to describe multicellular animals
the use of phylogeny to establish series of ancestral morphological characters
eukaryote phylogeny in the molecular era - 2000s
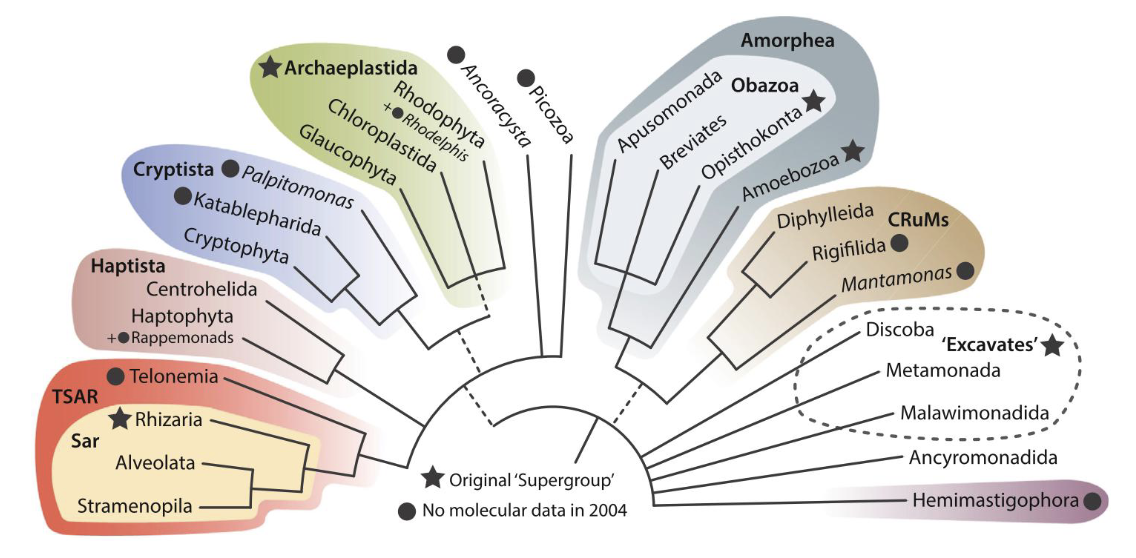
most of the supergroups had at least one morphological synapomorphy (a characteristic present in an ancestral species and shared exclusively by its evolutionary descendants)
the precise number and membership of the supergroups has varied, reflecting the rapid pace with which importnat taxa are being discovered and added to the tree (broad molecular phylogenetic analyses)
orphan taxa - Metamonada and Discoba
Discoba - Euglenoids
discoba consists of four main lineages: Jakobida, Euglenozoa, Heterolobosea, and Tsukubamonadida
Euglenozoa are a group of >1500 described species of single-celled flagellates with diverse modes of nutrition, including phagotrophy and photoautotrophy
upiquitous flagellates
covered with rigid pellicle allowing euglenoid specific motion (metaboly)
notes from video about euglenoids:
mostly found in freshwater
get food through phagocytosis
they dont have chloroplasts, instead they rely of feeding mechanisms like phagocytosis and osmosis
however, the older ones of these ingested green algae, and took the algae’s chloroplast and the rest of the photosynthetic apparatus, evolving the ability to make their own food in addition to consuming nutrients from their environment
the common link between the euglenoids is the striped surface called the pellicle, other than that, there is nothing else.
the most common shape among euglenoids is a sort of elongated almond shape and the end comes to a tapered point
then there is th emonomorphina pyrum, whose bodies are round and have a clear tail coming out from one end
almost all the Phacus species have a flat diamond body, resembling a leaf, and some Phacus helicoides can twist the leaf into a corkscrew structure; they have a pellicle, but are rigid and can’t engage in metaboly (euglenoid movement)
all photosynthetic euglenoids have two or more flagella, even if they are capable of metaboly
the eyespot in eugelnoids works as a shading device, telling the cell where it can find light to drive photosynthesis
the chloroplasts in the euglenoids are different from their ancestral counterparts, particulary in how they store their energy
green algae create a starch that is stored in the chloroplast that is used later for their energetic needs, but Euglenoids produce a different carbohydrate called paramylon that is kept in the cytoplasm of the cell instead of the chloroplast
Symbiontids are anaerobic deep-sea microbial eukaryotes with sulfur or sulfide oxidizing epsilon proteobacteria epibionts
kinetoplastids are ubiquitous flagellates, free-living (bodonids) or parasitic (trypanosomes)
Supergroup Amorphea - Opisthokonts
amorphea is robustly supported in most phylogenomic analyses
there is a great diversity of lifestyles and morphologies among unicellular opisthokonts
there is a larger diversity of basal lineages on the animals side of opisthokonta, including the Icthyosporea
sister-group to Metazoa = Choanoflagellata
supergroup Archaeplastida - Glaucophytes, Phodophyta and Chlorophyta
Primary endosymbiosis
the first one occurred ~1.6 billion years ago and gave rise to glaucophytes, red algae and green algae (the ancestors of all plants)
the second one occurred ~90-140 million years ago, establishing a permanent photosynthetic compartment (the chromatophore) in amoebae in the genus Paulinella
Chloroplastida
Chloroplastida or Viridiplantae includes two evolutionary branches:
Chlorophyta (dominated Proterozoic oceans): form a large and morphologically diverse clade of marine, freshwater and terrestrial green algae
Stretophyta: include a paraphyletic assmeblage of green algae (charophytes) and the land plants
it is one of the main groups of photosynthetic eukaryotes
supergroup Cryptista - Cryptophytes
Cryptophytes, a lineage that has been central to the stufy of secondary origin of chloroplast
chryptophytes plastids (red algal origin) is surrounded by four membranes; presence of a nucleomorph (NM) = residual nucleus of the red algal endosymbiont
they are ubiquitous in marine, brackish, and freshwater habitats
mostly photosynthetic with some mixotrophic species
because they produce PUFA, it’s used as food for copeopds in fish farming
chlorplast contains phycobiliprotein (phycoerythrin)
supergroup Haptista - Centrohelida, Rappemonads and Haptophytes
haptista are characterised by the haptonema, a third appendage used for attachement and food handling
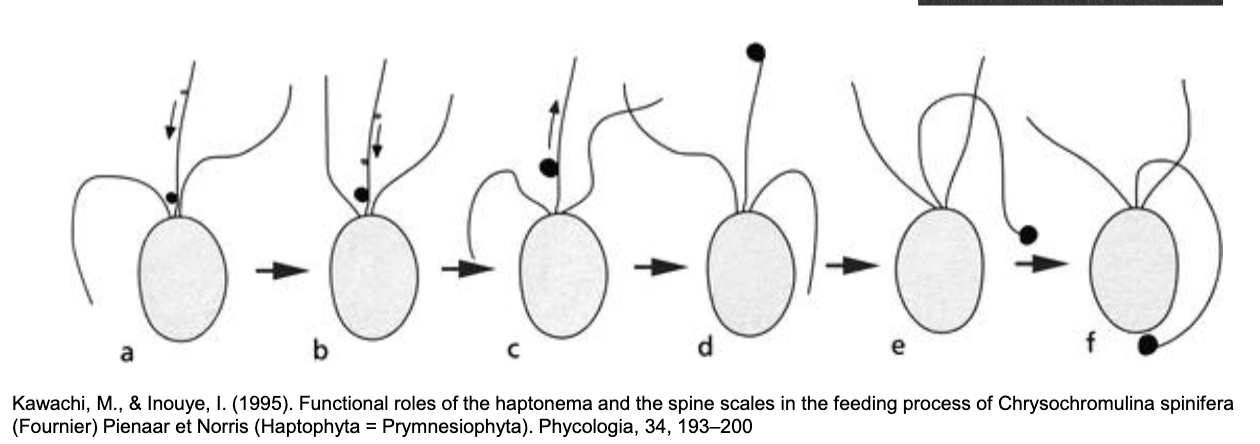
three groups:
Haptophyte
Centrohelida
Rappemonads
predominantly planktonic
some cells are covered of calcified plates, the coccoliths
marine environments worldwide
major component of the marine ecosystem
play a significant role in the global carbonate cycle through photosynthesis and calcification
blooms can be seen from space
the turquise color is a result of the light getting scattered by their calcium carbonate coccoliths (scales)
diversified in the Mesozoic to become a significant component of the phytoplankton (todays evidence, the white cliffs of Dover)
Supergroup TSAR - the secret rulers of our world
TSAR: Telonemids, stramenopiles, alveolates, and rhizaria
it is a gigantic assemblage of eukaryote lineages and possible a rival to Opisthokonta in diversity
it includes several
major groups of microbial algae (e.g. diatoms, dinoflagellates)
large seaweeds (e.g. kelp)
ecologically important free-living heterotrophic microbial eukaryotes (e.g. ciliates, foraminiferans, radiolarians)
many well-studied parasites (e.g. apicomplexans, oomycetes)
Alveolata
molecular data is supported by a morphological synapomorphy: the cortical alveoli under the cell membrane
two main subclades:
ciliophora (ciliates)
Myzozoa (apicomplexa, dinoflagellates, perkinsea)
evidences of seocndary endosymbiosis:
apicomplexa: derived non-photosynthetic plastid called apicomplast
cilliates: multiple genes of apparent algal origin present in the macronuclei
Ciliophora →
more than 8000 described species
nuclear dualism:
micronucleus (2N, “germ line”)
macronucleus (> 2N, “somatic line")
macronucleus polyploid and undergo direct division without mitosis. It controls the Non-reproductive cell functions, such as metabolism
sexual reproduction by conjugation
cilia covering most og the cell, sometimes arranged in dense patches (e.g. cirri)
perfectly adapted to life in soil/sediments, but also present in the plankton (e.g. tintinnids)
all heterotrophic species (kleptoplasty of cryptophytes)
form a lorica with debris
they feed primarily on other protists and bacteria
they are an important part of the fossil record
Apicomplexa →
very succesful group of endoparasites or endocommensals of animals, from corals to humans:
COccidiomorphea: contain well known vertebrate parasites like Plasmodium (malaria) and Toxoplasma which means that these aprasites evolved from free living algae
gregarinomorphea: contain mostly invertebrate parasites
Dinophyta →
atypical nucelus (dinokaryon) with high DNA content and permanently condensed chromosomes
550 genera with > 2000 species (~50% photosynthetic)
athecate (naked) or thecate (cellulose cell)
many cases of plastid loss (and re-aqusition from variuos donors: haptophytes, chlorphytes, cryptophytes by kleptoplastid)
Rhizaria →
still poorly known genetically compared with other major TSAr clades
united by molecular and morphological data (amoeboid “tendency” - filopodia and reticulopodia)
foraminifera - the benthic builders
main group with microfossil record
more than 12 000 species
mostly benthic
several live in symbiosis with teh dinoflagellates Symbiodinium (same found in corals)
radiolaria
the favourite protists of artists
great carbon sinkers
Stramenopila
major clade supported by both moelcular and morphological data (falgella with tripartite tubular hair - mastigonemes)
diatoms and brown algae
heterotrophic protists
including parasites
commensals of plants and animals
lineages known only from environmental DNA
Bacillariophyta →
the jewels of the sea
enclosed in a siliceous frustule
heterokont flagella present during sexual reproduction
250 genera, > 10000 species
~20% of the phytoplankton primary