4.2: Plant reproduction
Flower structure
Flowering plants are known as Angiosperms, and the flowers are their reproductive structures.
Most angiosperms are diploid, with meiosis occuring in the reproductive tissues and producing haploid spores which contain gametes.
Male spores and pollen grains, and female spores are the embryo sac.
Most species are hermaphrodite - containing both male and female parts.
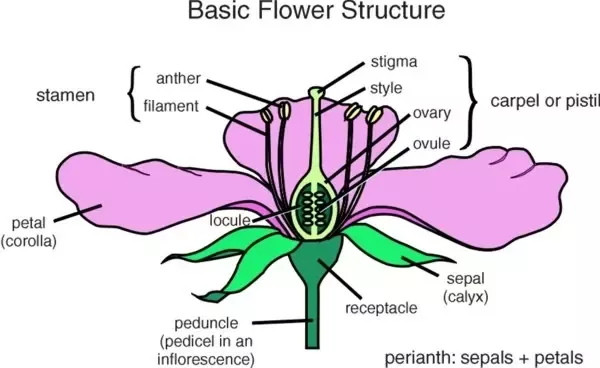
A flower is made up of many structures:
At the base is the receptacle, which helps support the flower.
The outermost ring of structures is the calyx, which is made up of individual sepals.
They are normally green, and protect the flower in bud. They are modified leaves.
In lilies, however, sepals are coloured.
Inside the calyx is the corolla, a ring of petals.
These can range from absent to large, and from pale green to brightly coloured. They are modified leaves.
Nectaries, if present, will be at the base of this. They release scented nectars to attract pollinators.
Inside the petals are the male parts of the flower, known as the stamen.
They consistent of a filament, which supports an anther. This produces the pollen grains.
The filament contains vascular tissue, which transports sucrose, mineral ions and water to developing pollen grains.
The anther usually contains four pollen sacs arranged in two pairs, side by side. When mature, the sacs dehisce - open up and release pollen.
In the centre of the flower is the carpel, which are the female parts of the flower.
A carpel is a closed structure in which one or more ovules develop.
The lower part of the carpel which surrounds the ovules is the ovary.
At it’s tip is the long style, which ends in the receptive surface, the stigma.
Types of cross-pollination
There are two types of cross-pollination, either done by:
Pollinators, such as bees, which are attracted to the large colourful flowers, the scent and their nectar (which is mainly sucrose).
They use long tongues to reach the sugary nectar at the base of the flower.
This causes the bee to rub against the anther, brushing the sticky and sculptured pollen against the bee.
When it enters another flower, the pollen rubs against the ripe stigma, causing cross-pollination.
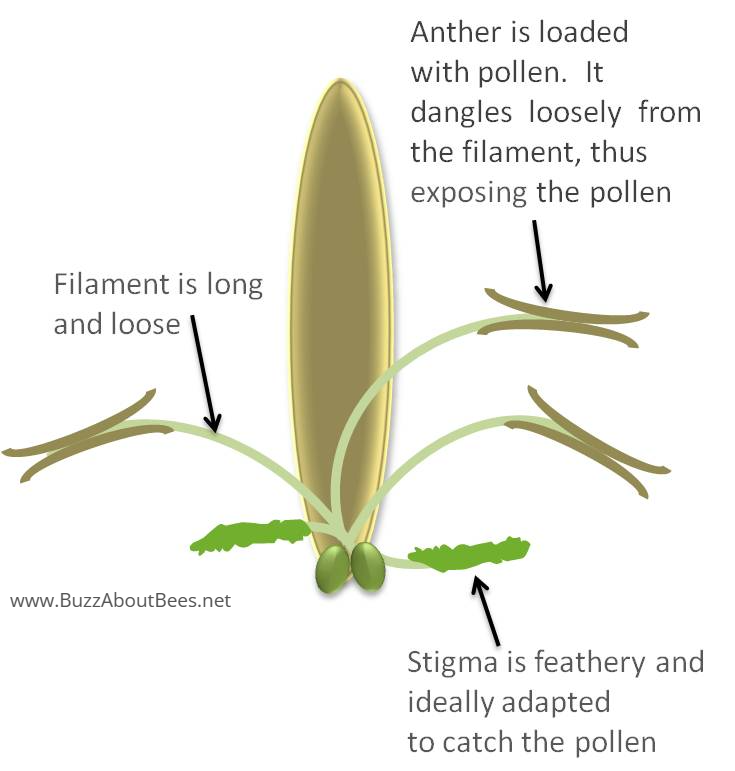
Wind pollinated plants do not have nectar, a scent or bright coloured flowers, relying instead on nature.
Their anthers hang outside so the wind can blow any their small, smooth and light pollen.
Feathery stigmas hang outside the flowers and provide a large surface area for catching pollen grains.
They normally group together in an inflorescence - a cluster of flowers on a branch.
Pollination
This is the transfer of pollen grains from the anther to the mature stigma of a plant of the same species.
This brings the pollen grains into contact with the female part of the flower, which can result in fertilisation.
Self-pollination
This is when the pollen from a flower’s anthers is transferred to the mature stigma of the same flower, or another flower on the same plant.
This is also known as inbreeding.
It means that genetic variation can only occur via independent assortment, crossing over and mutations.
This means they have lower genetic variation, and therefore are more susceptible to natural selection as they are less adaptable.
This also increases the chances of two harmful recessive alleles from being continued at fertilisation.
However, it is successful at maintaining successful genomes which are well-suited to stable environments.
It also requires less energy.
Cross pollination
This is when pollen from a flower’s anthers are transferred to the mature stigma on another flower on another plant of the same species.
Occurs in most Angiosperms.
This is also known as outbreeding.
It produces high amounts of genetic variation, as there are two different haploid sets as well as crossing over, independent assortment and mutations.
This allows a species a higher chance of survival in a changing environment, as some individuals are likely to have a suitable set of alleles.
The chance of harmful allele combinations is reduced.
It requires more energy.
Ensuring cross-pollination
There is a variety of methods to avoid self-pollination:
Dichogamy, which is when the stamen and stigma ripen at different times. There are two types:
Protandry - Stamens ripen first, such as in daisies.
Protogyny - Stigma ripens first, is rarer, such as in bluebells.
The anther is below the stigma to avoid pollen falling on to it, such as in pin-eyed primroses.
Genetic incompatibility, where pollen cannot germinate on the stigma of the flower which produced it, such as in red clover.
Separate male and female flowers on the same plant, such as maize.
Separate male and female plants, such as holly.
Gamete development
Male gametes
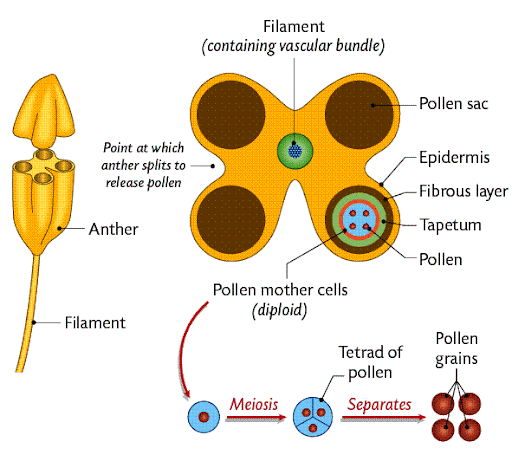
Anther structure
The anther is made up of many structures:
The outermost layer is known as the epidermis.
In the centre is the vascular strand, which contains xylem and phloem tissue.
Around each of the four pollen sacs is multiple layers:
The outermost layer is the fibrous layer.
Then is the tapetum.
This provides nutrients and regulatory molecules to the developing pollen grains.
It also has a significant role in the formation of the pollen cell wall, which is chemical resistant and tough.
It resists desiccation, so the pollen grains can transfer from one flower to another without drying out.
UV light can also not pass through this cell wall, protecting the DNA inside the pollen from mutations. This is especially useful as pollen can be carried at high altitudes.
In order to release pollen when it is mature, dehiscence occurs:
The outer layers of the anther dries out, causing tension in the lateral grooves.
This tension then pulls the walls of the anther apart, and the edges of the pollen sacs curl away. This is dehiscence.
An opening called the stomium exposes pollen grains, and they are then carried away by pollinators or the wind.
Pollen grains
This occurs in the pollen sacs of the anther:
The diploid pollen mother cell undergo meiosis, forming a tetrad.
This tetrad is made up of four haploid cells, which become four pollen grains.
These cells then undergo mitosis to produce two nuclei, one generative and one tube nucleus.
The generative nucleus then produces two male nuclei by mitosis.
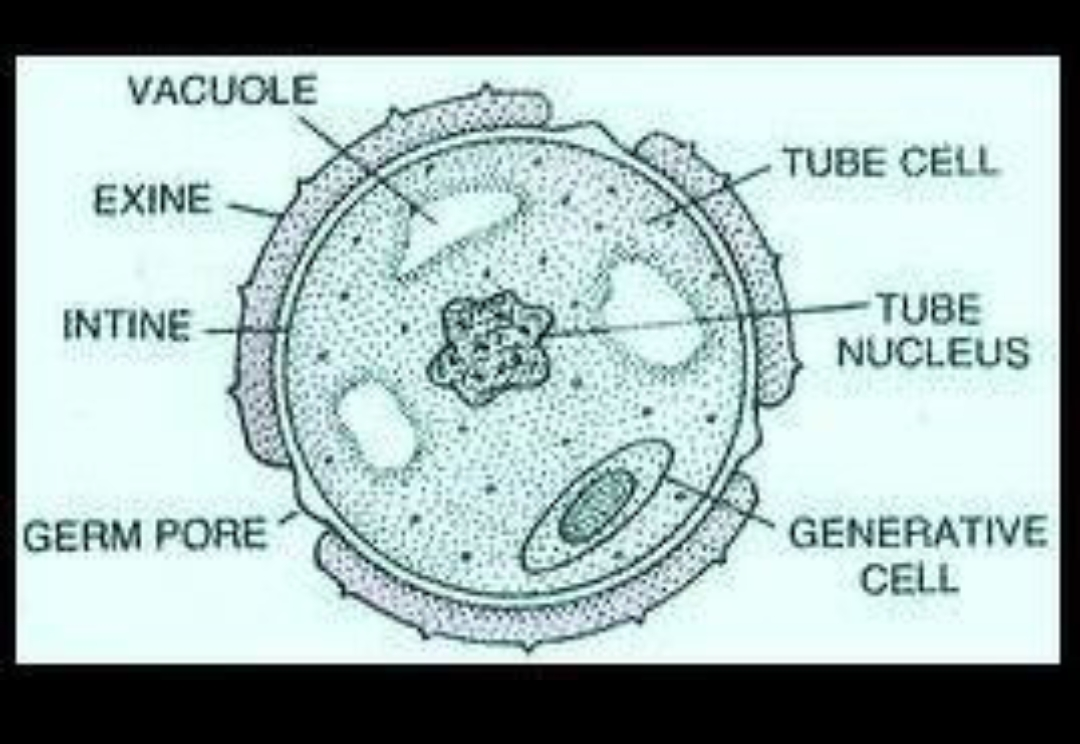
A pollen grain also has many structures:
It has a special cell wall made up of three elements:
The outermost layer, the exine. This is a waterproof and tough layer.
The innermost layer, the intine. This is a thinner layer.
The pits, which are holes in the exine but not the intine. They are used for gas exchange.
It has the two nuclei; the generative and tube.
The generative splits via mitosis before double fertilisation occurs.
Female gametes
Ovule structure
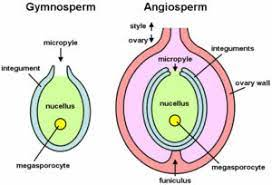
There are many structures surrounding the ovule:
The ovary wall surrounds the ovary.
Attached to this wall by the funicle is the ovule. There can be multiple ovules in one ovary.
The ovule is made up of:
2 cell layers known as integuments surround the embryo sac.
They have a gap known as the micropyle.
The innermost layer is the nucellus, a cell layer which provides nutrients.
The centre is the embryo sac.
Embryo sacs
This occurs in the ovule:
A megaspore mother cell, surrounded by the nucellus, undergoes meiosis and makes four megaspores cells.
Three of these disintegrate.
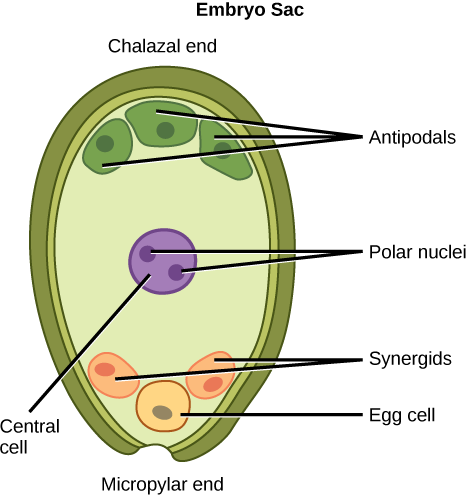
The remaining cell undergoes 3 rounds of mitosis - from 1 to 2 to 4 to 8. This produces 8 haploid nuclei. They have different names:
2 are polar nuclei, which fuse into 1 polar nuclei, making it a diploid cell. These are in the centre of the embryo sac.
2 synergids. They are located at the bottom of the embryo sac.
1 oosphere, which is the female gamete. It is positioned above and between the synergids.
3 antipodals. They move to the opposite side of the synergids.
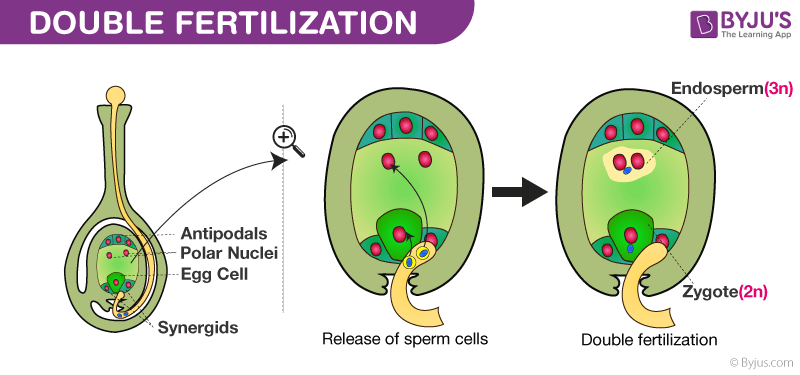
Fertilisation
Pollen tube formation
Pollen tube formation occurs in 6 steps:
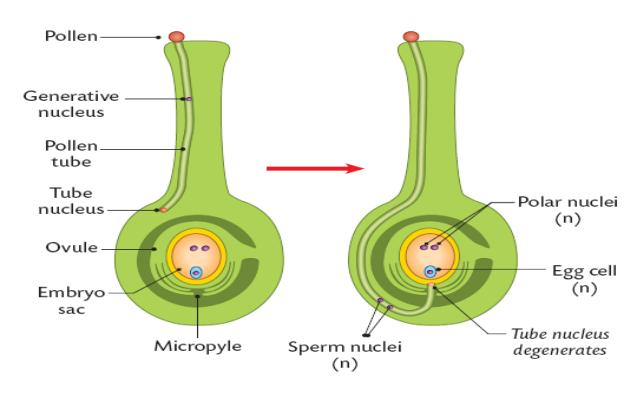
The pollen lands on the stigma, and germinates in the sucrose. If they are chemically compatible, the tube nucleus begins to make a pollen tube:
The nucleus grows the pollen tube out of one of the pits, and leads the pollen tube.
The pollen nucleus codes for hydrolases, such as proteases and cellulases to digest through the stigma and the style. Products of digestion are used by the pollen tube.
It moves up a gradient chemoattractants such as GABA released by the ovule.
It grows through the micropyle and passes into the embryo sac.
The pollen tube nucleus then disintegrates, as it has completed it’s function of controlling pollen tube growth.
Double fertilisation
The pollen tube tip opens, releasing the two male gametes, the generative nuclei, into the embryo sac.
One generative nucleus fuses with the oosphere, the female gamete. This forms a diploid zygote.
The other generative nucleus fuses with the diploid polar nucleus to form the triploid endosperm nucleus.
It then divides rapidly by mitosis, creating endosperm tissue, which takes over from the nucellus in providing nutrition for the embryo.
 Fruit and seed development
Fruit and seed development
The seed develops from the fertilised ovule, and contains an embryonic plant and a food store:
The zygote:
Divides by mitosis into an embryo. This consists of a plumule (the developing shoot), a radicle (the developing root) and one or two cotyledons (seed leaves).
The endosperm:
Develops into a food store for the developing embryo.
The ovule:
The integuments dry out, harden and become waterproof with deposits of lignin.
This is known as the seed coat or testa.
The micropyle remains as a hole in the seed.
The ovule; comprising the embryo, endosperm and testa becomes the seed.
The ovary:
The funicle of the ovule becomes the funcile of the seed.
Where it is attached becomes known as the hilum, which is a mark or scar which remains from the funicle.
The ovary itself becomes the fruit:
It can be sweet, juicy and pigmented like cherries, or dry and hard like almonds.
Fruit and seed structure
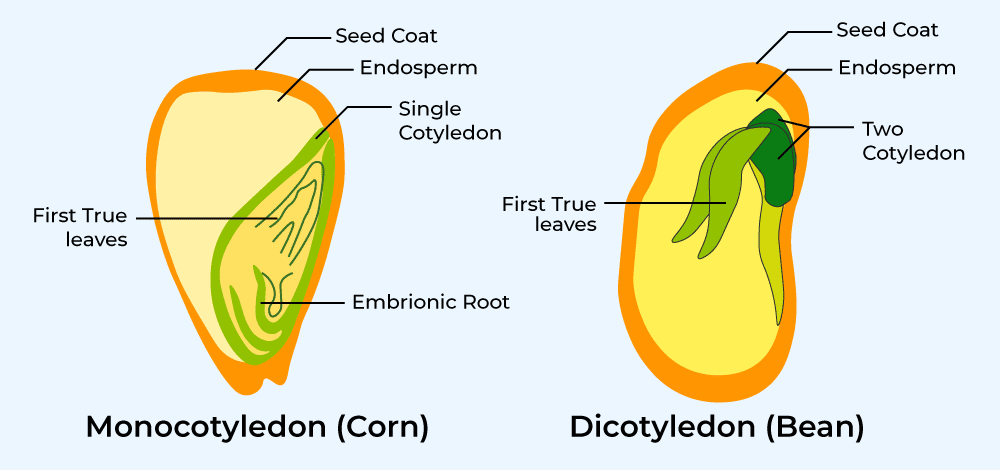
Dicotyledons
Dicotyledons have two seed leaves, or cotyledons, with the embryo lying between them.
The endosperm is absorbed into these cotyledons, so therefore it is a non-endospermic seed.
An example is the broad bean, which has several ovules.
After fertilisation, the ovary elongates into a pod, which is the fruit. The broad beans of the seeds.
Other traits include:
A network of leaf veins.
Sepals, petals and stamens are in multiples of 4 or 5.
Vascular bundles in a ring around stems.
Vascular bundles in the centre of roots.
Monocotyledon
Monocotyledons have one seed leaf, or cotyledon.
The endosperm remains as the food store, so it is therefore endospermic.
The cotyledon remains small and does not develop further.
Seeds then become dormant, with their water content falling below 10% to reduce their metabolic rate.
They can remain like this for long time periods and do not germinate until conditions are suitable.
An example is maize plants, which are similar to other cereal grains and grasses.
The testa fuses with the ovary wall, making it a one seeded fruit.
Other traits include:
Leaf veins are parallel.
Sepals, petals and stamens are in multiples of 3.
Vascular bundles scattered in stems.
Vascular bundles are scattered in roots.
Seed dispersal
The methods by which seeds move away from the parent plant.
If they germinated too close, the parent plant would outcompete it’s seedling, and it would likely die.
These have been developed via natural selection, and there is 6 types:
Wind:
Fruits can either have pores, sails, or parachute-like structures to allow wind dispersal.
Seeds are often shaken out when the stem is blown by the wind, and the wind allows them to travel great distances.
An example is dandelions.

Transport:
Birds, mammals, reptiles and fish eat seeds, which are dispersed via their faces.
Some species can only germinate after scarification has occurred, a process where an animal’s digestive system weakens the testa via acid and enzymes.
Tasty fruits have evolved to attract this type of transport, such as cherries.

Rolling:
When the fruit breaks open to release the seed, which falls and rolls across the ground, away from the parent tree.
An example is conkers.

Bursting:
When legume pods dry, they split and the seeds scatter.
In some species, the pods rotate as they burst open, sending the seeds in many different directions.
An example is pea pods.

Water:
Seeds that are developed to be buoyant via air cavities, and float in the water when they fall, and are carried away.
An example is coconut trees.

Carrying:
Hooked seeds that attach to animals coats to be carried away.
An example is burdock.
 Seeds and survival
Seeds and survival
There are many evolutionary developments in seeds:
Dormancy:
Dormant seeds have a low metabolic rate and can survive very cold weather.
The water content is reduced below 10% so seeds can survive very dry conditions.
Testa:
It is chemically resistant, so seeds can survive adverse chemical conditions.
It can physically protect the embryo.
Dispersal:
Seeds can be dispersed great distances to not compete with the parent plant.
It allows for the colonisation of new environments.
Nutrients:
These are provided by cotyledons or the endosperm, and lasts until the embryo can photosynthesise on its own.
Inhibitors:
Allow germination to occur at a suitable time of year. Vernalisation is the process of these inhibitors being broken down in very cold weather, allowing for germination in the spring.
These inhibitors can be in the seeds or the fruits.
Germination
Germination is the process in which a plant grows from a seed.
It lasts until the first photosynthesising leaves are produced, which is when all the endosperm and cotyledon food stores will be used.
It will occur after a period of dormancy, and when environmental factors are favourable.
Requirements
A suitable temperature - This is for enzyme action, and varies from species to species, usually between 5-30 degrees.
Water - Mobilises enzymes for transport via the xylem and phloem, and vacuolates (forms vacuoles) cells, which makes them turgid.
Oxygen - Aerobic respiration is needed for energy, which is needed for metabolism and growth.
Lighting - This differs between species, with some seeds needing light to germinate, some needing darkness, and some unbothered.
Dicotyledon/ broad bean germination
This occurs in 6 steps:
When conditions are suitable, water is imbibed (absorbed into pores) rapidly by the seed through the micropyle.
The water allows enzyme action and tissues to swell.
Food reserves in seeds are insoluble in water, and need to be broken down to be transported to the embryo.
Amylose hydrolyses starch to maltose, and protease hydrolyses proteins to amino acids.
These soluble products are transported to the embryo.
They are also transported to the apical meristems (tissue with unspecialised cells) of the plumule and radicle via the phloem.
Rapid cell division then occurs.
Some sugars are converted to cellulose for cell wall synthesis.
Aerobic respiration releases energy from sugars, and amino acids synthesise proteins.
The swollen tissues then rupture the testa, and the radicle emerges.
It is positively geotropic (moves in response to gravity) and negatively phototropic (moves in response to light), and therefore grows down into the soil.
The plumule then emerges.
This is negatively geotropic and positively phototropic, so it grows upwards and through the soil.
The part of the plumule between the embryo and cotyledons (which remain below ground) elongates rapidly, pushing the plumule upwards.
The plumule is bent over in the shape of a hook as it pushes through, protecting it from damage via soil abrasion.
If planted at the correct depth, the plumule hook will straighten and the leaves will unfurl, beginning to photosynthesise.
By now food stores in the cotyledons will have been depleted.

Monocotyledons/ barley
Barley is often used as an example due to the extensive research done by the alcohol industry into its germination.
Malting as a term refers to the maltose generated when the starch in barley is digested.
This occurs in 6 steps:
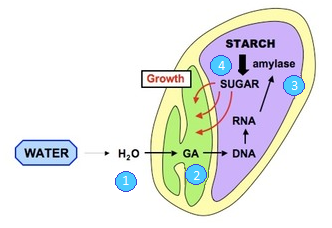
The barley embryo secretes gibberellic acid, a plant growth regulator, which diffuses through the endosperm to the outer aleurone layer.
This is a layer of cells which has a high protein content.
The gibberellic acid switches on genes in the cells of the aleurone layer, starting transcription and translation, producing enzymes such as protease and amylase.
Proteases hydrolyse proteins in the aleurone layer to amino acids, which are used to make amylase.
Amylase diffuses out of the aleurone layer to hydrolyse the starch in the endosperm cells.
Maltose and glucose produced are diffused through the endosperm to the embryo, therefore the plumule and radicle.
These sugars are respired, which fuels biosynthesis and cell division, bringing the seed out of dormancy.