Innate Immunity
Slide 3: 3 phases of immune response to pathogen: recognition, reaction and resolution. Two branches to the immune system: innate and acquired/specific. Specific immune response = cell mediated and humoral responses.
Slide 4: Innate immunity is the early phase of host response always found in everybody. Doesn’t change with repeated exposure and discriminates between groups rather than specific pathogens (very primordial system). Adaptive immunity discriminates between individual pathogens, shows ‘memory’ and appears abruptly in evolution in cartilaginous fish.
Slide 5: Anatomical barrier = skin, mucosal barrier etc. Initial infection response when the pathogen enters the body relies on pathogen recognition receptors binding pathogen associated molecular patterns (PAMPs). Early induced innate response happens 4 to 96 hours after initial infection.
Adaptive response uses antigen-specific receptors produced by gene segment rearrangement. Unlike innate receptors these are encoded in multiple regions.
6: The innate immune response consists of soluble mediators and complement eg:
Mucins: may prevent microbes adhering to epithelium
Lysozyme: attacks peptidoglycan - discovered by Alexander Fleming
Defensins: disrupt bacterial and fungal membranes via pore formation
Cathelicidins: disrupt many microbial membranes
Histatins: active against pathogenic fungi
RegIII family: C type lectins which target peptidoglycan
(Martin Luther Decided to Come down Hard on Racism)
Complement is activated by mannose binding lectin, ficolin or C-reactive protein. MBL recognises sugars, ficolin recognises oligosaccharides containing acetylated sugars and CRP recognises phosphorylcholine.
These molecules are secreted by many different cell types. Typically requires proteolysis for activation and often work by disrupting the membrane eg by inserting pores
7: Innate receptors typically expressed non-clonally (expressed on all cells of certain type). NK cells are an exception as they express various combinations of receptors from different families - NK cells are all different. Innate receptors trigger an immediate response and like adaptive receptors are able to discriminate between very closely related molecules., however not as successfully.
Innate PRRs are a defined set of receptors found naturally in the genome. They recognise parts of pathogens
8:
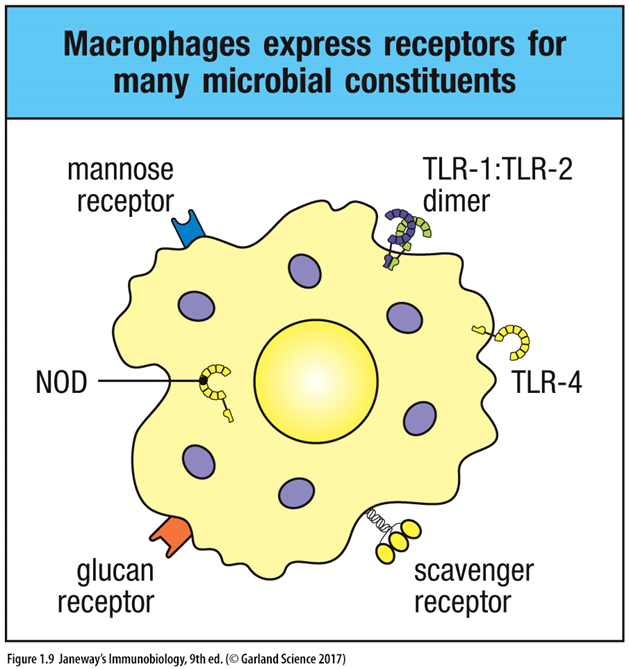
PRR types:
C-type lectin family – recognise beta-1,3-linked glucans (commonly found in fungal cell walls). Mannose receptors recognise pathogenic mannose structures but are also involved in clearing host proteins. Mannose structure binds receptor and is internalised for destruction.
Scavenger receptors – heterogenous, recognise anionic polymers and acetylated LDLs among other structures. Bind pathogens and host products, internalising them. Receptor that mediates atherosclerosis by making macrophages into fatty cells.
Complement and Fc receptors – bind complement- and antibody-coated organisms
Toll-like receptors (TLRs) – recognise molecular patterns not found in healthy vertebrate cells
9: The Toll receptor protein was first discovered in fruit flies (lower level organism used for study as it’s easier to spot outcome due to reduced genome). Flies deficient in Toll receptor were more susceptible to infection, especially fungal infection – showed uncontrollable Aspergillus fumigatus growth. Later found in humans and other mammals
10: later found that TLRs had many different subtypes which respond to different types of pathogenic structures eg TLR5 responds to flagellin in bacteria whereas TLR-7 and 8 respond to viral ssRNA
11: TLRs contain extracellular leucine rich repeats whereas IL-1Rs contain an Ig-like domain. Both TLRs and IL-1Rs have an intracellular signalling domain Toll/IL-1 receptor domain which is important for downstream innate immune signalling (internally similar region containing ‘boxes’ 1 to 3). Similar signalling cascades are activated – MAPK and NFkB respectively. Both domains recruit adaptor proteins, mainly MyD88. These receptors share significant structural and functional similarities – both are involved in inflammatory responses, TLRs by recognising PAMPS and IL-1R binds proinflammatory cytokine IL-1.
12: TLRs not all found on cell surface.
Extracellular TLRs eg TLR-2:TLR6 contain 18-25 copies of leucine-rich repeats. Intracellular TLRs found in the endosomial compartment recognise parts of microbes eg DNA or RNA. DNA only accessible after microbe broken down and RNA not usually found in healthy mammalian endosomes. Diacyl lipopeptides recognised by heterodimeric TLR6:TLR-2 receptors are derived from lipoteichoic acid of Gram +ve cell walls and lipoproteins from Gram -ve surfaces.
13: Activating a TLR with a certain type of pathogen activates a response. Once the receptor is activated it interacts with adaptor proteins (bind something to something else). Some pathways share adaptor proteins but not always, leading to a different outcome eg cytokine production or type I IFN production.
14: Dimerisation important in many aspects of biology as it helps provide a maximal response eg bringing signalling components together. Sometimes described as producing a highly active stage of receptor. TLR-1 and 2 found on the cell surface form heterodimers bringing their cytoplasmic TIR domains together, initiating signalling. Dimerisation is caused by each TLR binding the same lipopeptide.
TLR-4 is associated with accessory protein MD2 which recognises LPS. 5 of LPS’s acyl chains bind a pocket in MD2 and 1 remains free. The few chain binds the outer convex surface of another TLR-4 inducing a homodimer. LPS bound to the second molecule stabilises the dimer which leads to signal transduction. MD2 helps to stabilise the dimer so it doesn’t ‘fall apart’
15: TLR4
· LPS binding -> dimerisation.
· Signal transduction downstream is divided into MyD88 dependent and TRIF dependent pathways:
- MyD88 dependent -> proinflammatory cytokine production eg TNF-alpha, IL-6 and IL-1beta.
- TRIF dependent -> type I interferon (alpha and beta) production and late phase NF-kB activation.
· Adaptor recruitment:
- MyD88 and TIRAP (TIRAP binds MyD88) -> series of intracellular reactions -> Nf-kB activation. NF-kB moves into nucleus -> cytokine production
- OR TRIF and TRAM -> event cascade -> T1 interferon production and (slower) activation of NF-kB
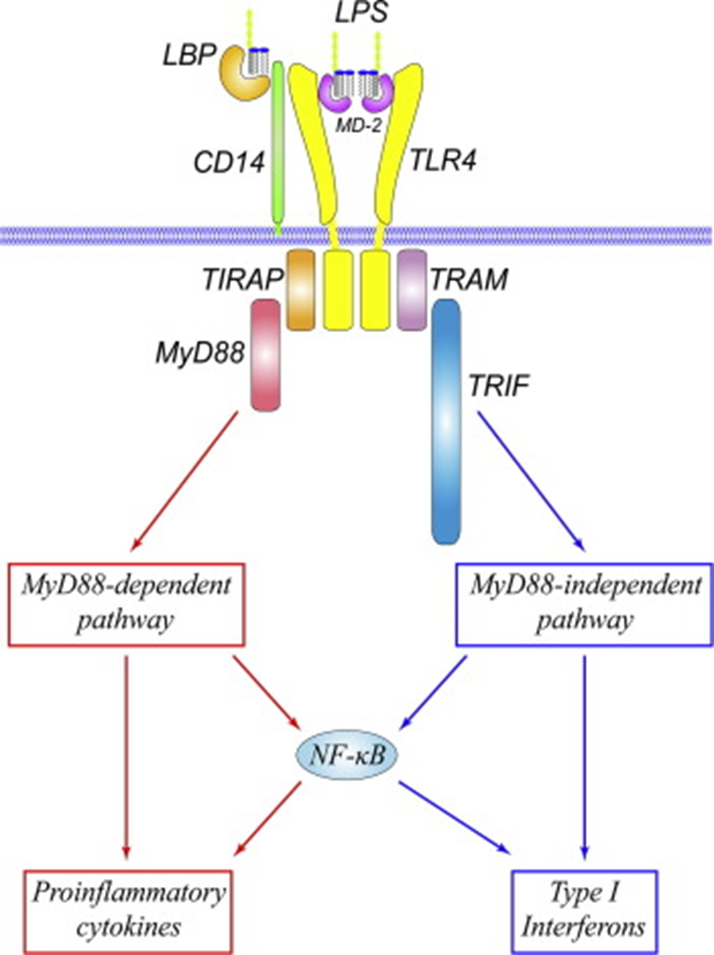
16: Because of TLR discovery we now know how the innate immune response is initiated. As a result of TLR4 signalling:
- NF-kB activation -> proinflammatory cytokines TNF-alpha, IL-6 and IL-1beta
- IRF3 activation -> type I interferon production
- MAPK pathways (ERK, JNK and p38) activated -> inflammation, cell survival and apoptosis
- AM defence through phagocytosis promotion and recruitment of immune cells to infection site
17: NOD-like receptors (NLRs) are cytoplasmic (found inside the cell) receptor proteins of which 22 have been found in humans. They sense bacterial invasion and cellular damage within the cell. They have a central nucleotide-binding oligomerisation domain (NOD) as well as other domains which detect microbial products and cellular damage.
Some NLRs activate NFkB (transcription factor which turns on inflammatory mediator production- read into it) for the same responses as TLRs whereas others activate a distinct pathway inducing cell death and inflammatory cytokine production. High number of genetic mutations associated with chronic inflammatory/autoimmune conditions associated with NLRs.
18: NLRs contain amino terminal caspase recruitment domains (CARD). CARD structurally related to MyD88 TIR death domain. It can dimerise with CARDs on other proteins to induce signalling. NOD1 senses gamma-glutamyl diaminopimelic acid (iE-DAP) which is produced peptidoglycan breakdown from Gram -ve eg Salmonella and some Gram +ve eg Listeria. NOD2 recognises muramyl dipeptide (MDP) found in most peptidoglycan.
Signalling proteins contain protein-interaction domains which mediate assembly of larger complexes. Domains direct or suggest function of the protein eg what it activates.
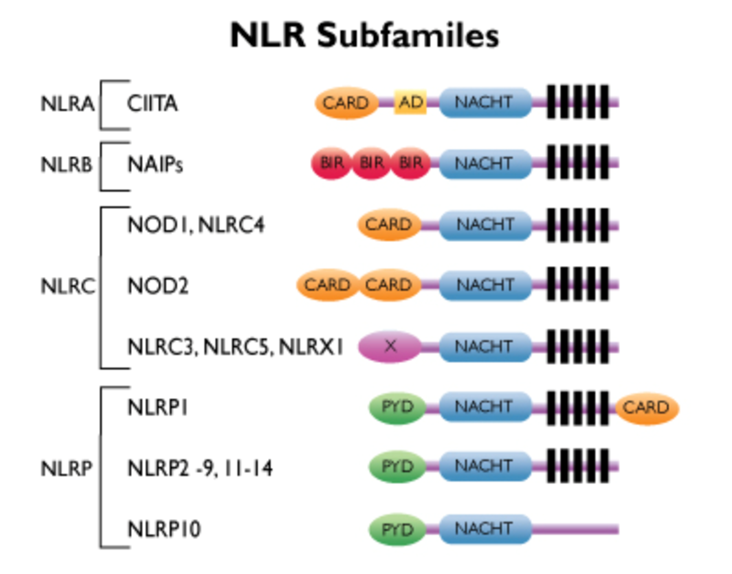
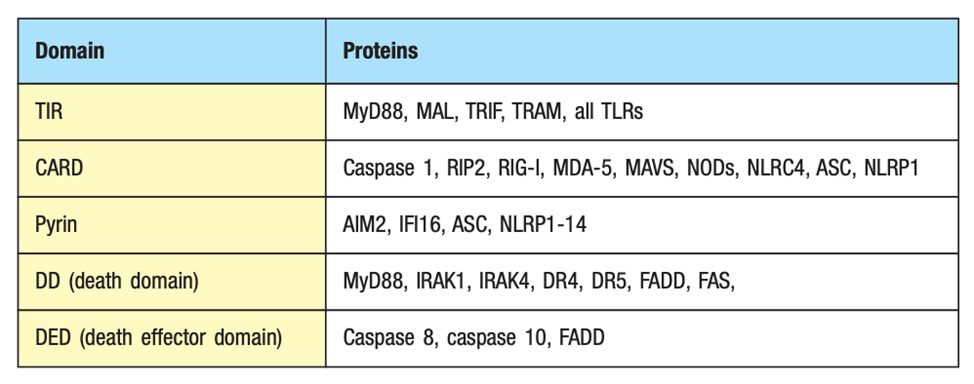
19: Pyrin domains are structurally related to CARD and TIR domains important to forming inflammasome. They interact with other pyrins. Humans have 14 NLRs which contain a pyrin domain, the two most well-known being NLRP1 and NLRP3. These receptors contribute to a complex called the inflammasome which drives proinflammatory cytokine production and induces cell death. They detect many PAMPs and DAMPs (damage associated - damaged host cells). Activating the inflammasome is needed for pathogen host defence however it’s been recently shown to contribute to pathogenesis of IBD, rheumatoid arthritic and atherosclerosis. Different receptor families have different inflammasomes which they induce. Cryo-EM can be used to observe inflammasome formation and structure, a diagram of which is shown below.
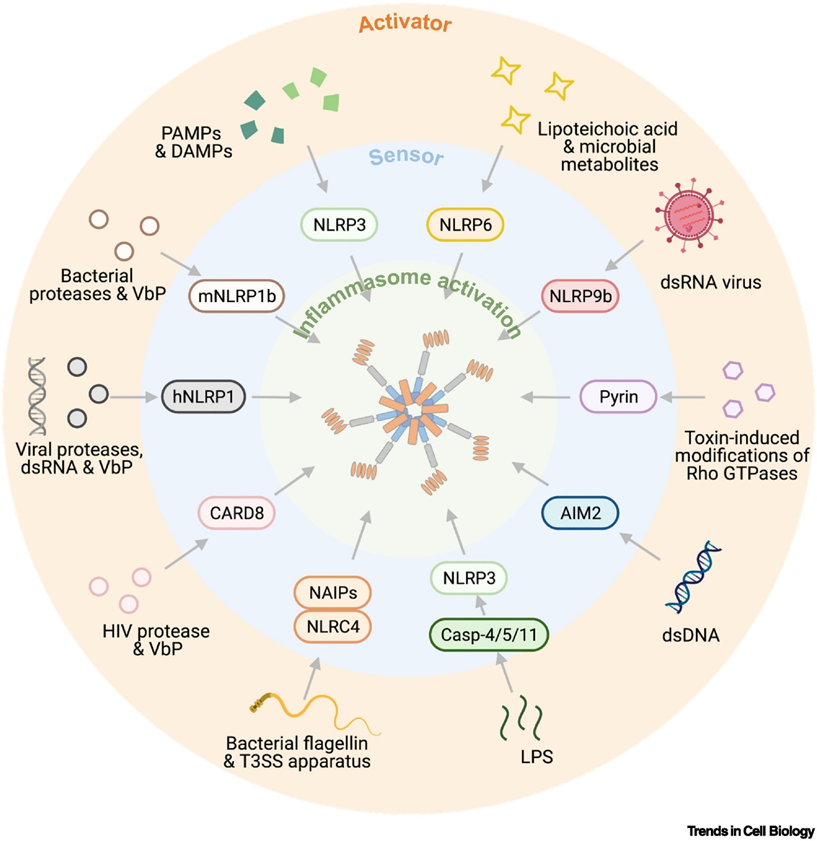
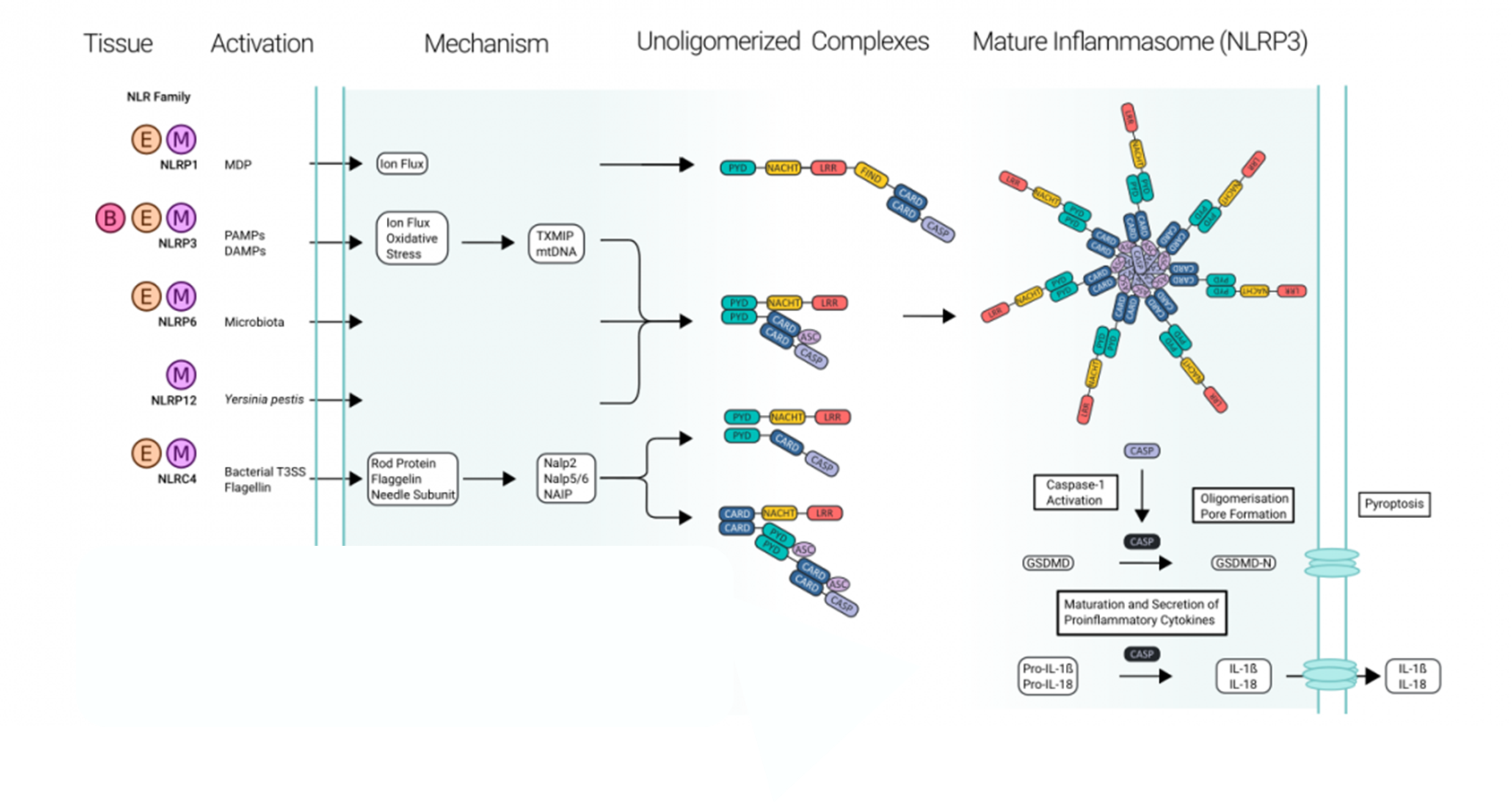
20: NLRP3 forms an inflammasome complex which contains adaptor protein ASC (apoptosis associated speck-like protein containing a CARD). ASC contributes to recruiting and activating caspase-1 which processes inactive cytokines eg IL-18 into their active forms. These cytokines trigger inflammation and recruit other immune cells. Overactivation of the NLRP3 pathway can cause excessive inflammation and contribute to autoimmune/chronic inflammation conditions.
Caspase = enzyme associated with apoptosis (cleave), kinases phosphorylate and ubiquitinases cleave ubiquitin, phosphorylases dephosphorylates.
Procaspase = active form
21: diagram
1 cleavage event caspase is involved in is GSDMD-N → GSDMD cleavage which causes holes to form in cell membrane, a form of apoptosis called pyroptosis. Also takes inactive version of IL-1beta and cleaves it to form active version
22: regulation of the immune response is important to prevent overreactive immune response which could cause chronic inflammation, autoimmune disease and inflammatory disorders eg gout and atherosclerosis. Targeting the pathways mentioned in this lecture could enable us to treat autoimmune disorders etc and allow us to design vaccines.
Extra reading REQUIRED:
Toll/IL-1 Receptor (TIR) domain
TLR signalling
TLR4
activation of MAPK pathways (ERK, JNK, p38) - don’t need to learn every step but must understand process and result of pathway
NOD-like receptors (NLRs)
pyrin-containing NOD-like receptors (NLRP) inc NLRP3
Robin’s study: Knockout of TLR2 has no effect on LPS and Leishmania activating MAPK pathway whereas TLR4 knockout leads to no activation of MAPK pathway in response to LPS.
Studies could use knockout mice, CRISPR..