Oct 15th - Small molecule Transport & Electrical Properties of Membranes
movement of an atom/molecule across membranes is governed by its concentration and properties
the hydrophobic layer of membranes is a barrier to the movement of some molecules
larger and more polar molecules have more difficult time crossing than smaller & non-polar molecules
for practical purposes, ions can’t cross
an imbalance in the concentration of a solute on either side of the membrane is a driving force for the movement of the solute
transport proteins grant selectivity to lipid bilayers
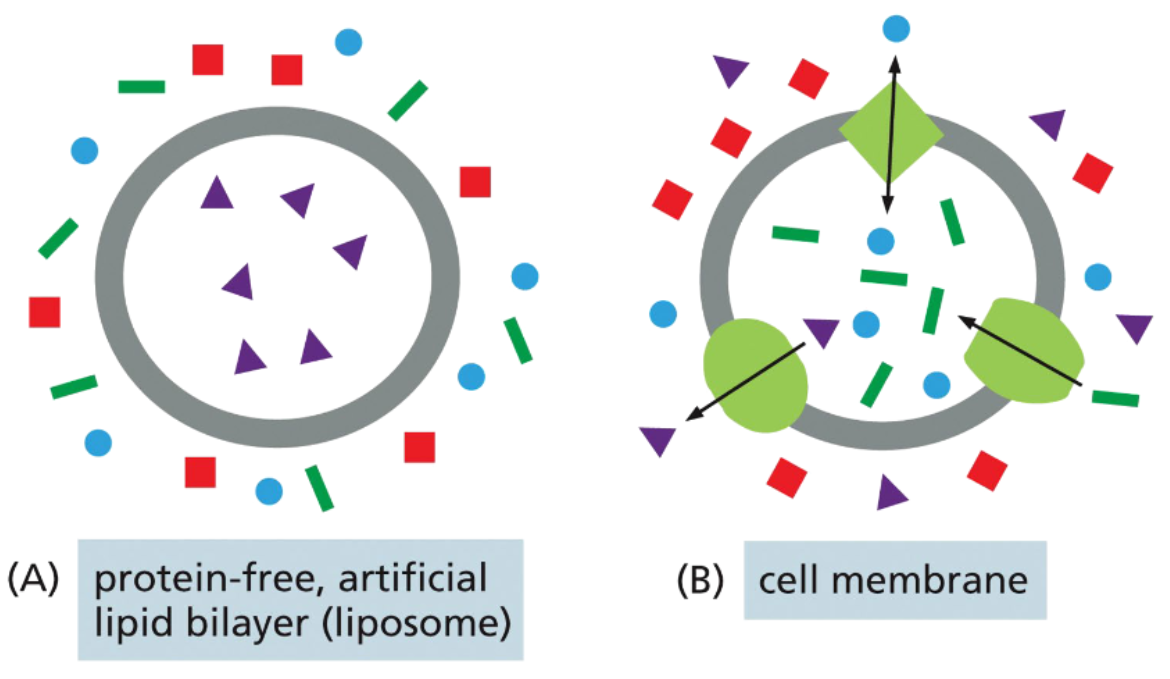
channels and transporters allow charged and polar solutes across membranes
proteins that facilitate movement of solutes across membranes belong to two broad classes
transporters move fixed amounts of solutes at a time via conformational changes; they are slower than channels
some transporter participate in facilitated diffusion & others use energy to move solutes against their gradient (active transport)
channels create open paths through which solutes can rapidly flow
some channels can have open and closed states that are regulated
eg ligand, voltage and mechanically-gated channels
channels can only allow solutes to move according to their gradient (passive transport / facilitated diffusion)
both channels & transporters are selective with respect to which solute they move
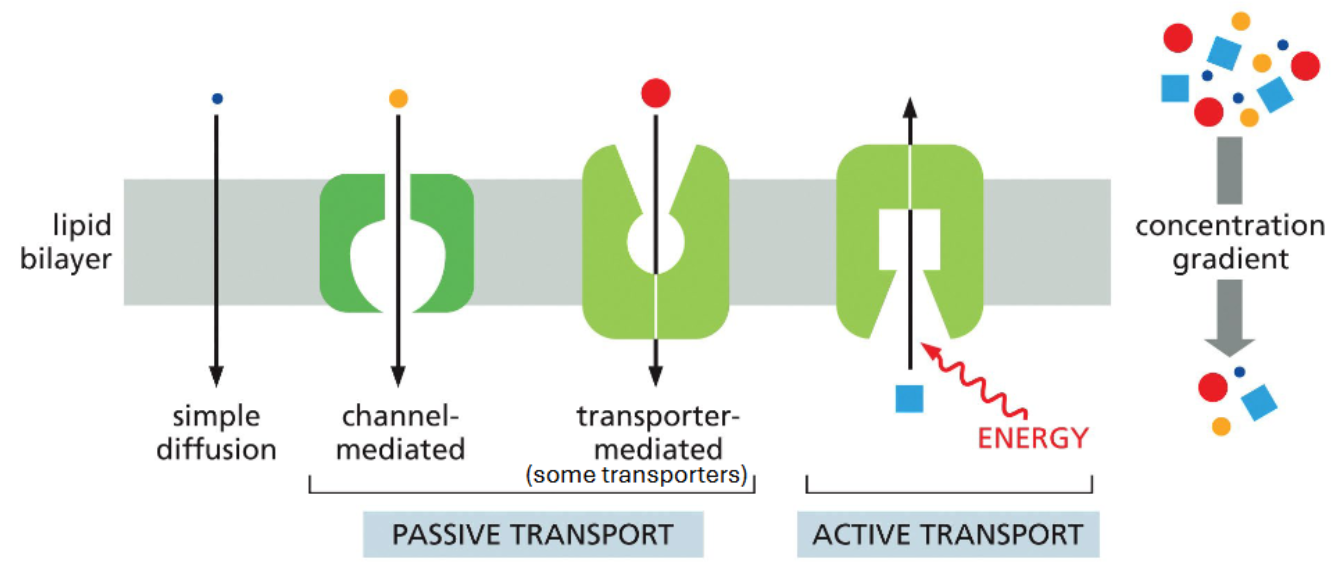
multiple modes of transmembrane movement
simple diffusion and passive transport both involve movement of a solute down its concentration gradient
active transport involves solute movement against its concentration gradient
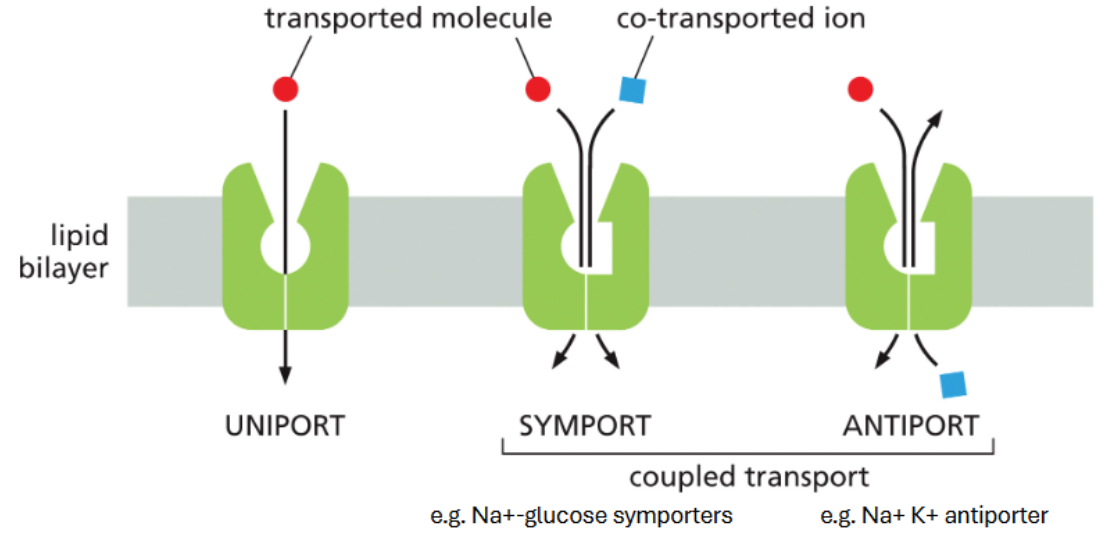
transporters are defined by number & direction of solutes
uniport — transport single molecules across the membrane
symporters — transfer two solutes, same direction
antiporters — transfer two solutes, opposite directions
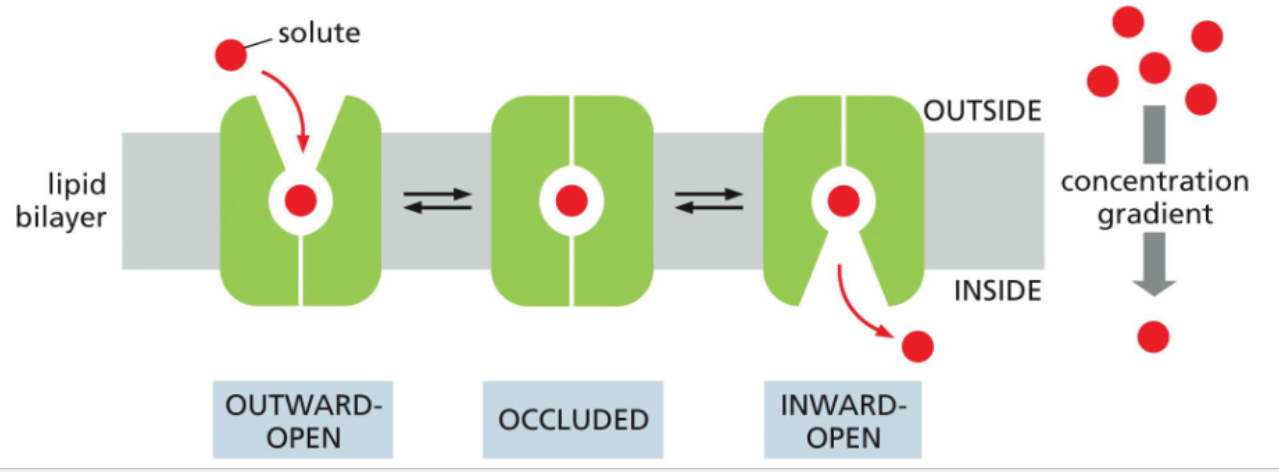
passive transporters alternate between different conformations independent of solute binding
passive transporters continuously alternate between different conformations that position the solute binding site on either side of the membrane
this conformation is independent of solute binding so a solute moved in one direction can be moved in the opposite direction just as easily
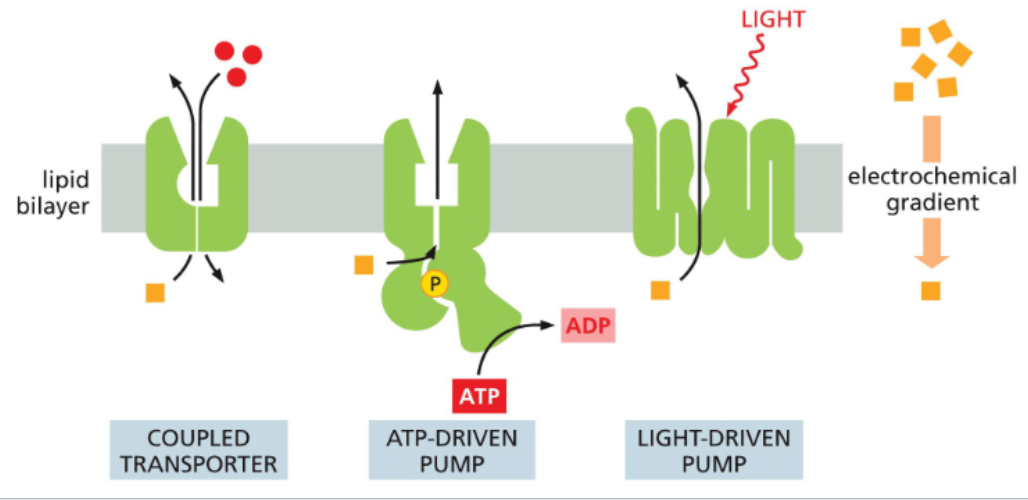
active transporter use energy to move solutes against their electrochemical gradient
movement of a solute against its electrochemical gradient is energetically unfavorable, so additional energy must be applied
coupled transporters use the movement of one solute with its gradient to power movement of another against its gradient
ATP-driven pumps use the energy of ATP hydrolysis to power solute-power moving conformational changes
light-driven pumps use photon-powered conformational changes to move solutes
Redox driven pumps use the energy of electron transfers to move solutes
ion movement is governed by both gradient and electrical charge
unequal distribution of ions results in a charge difference (voltage) across the plasma membrane
opposite charges on either side of the membrane collect at the membrane; the cytosol and extracellular fluid are electrically neutral — the voltage exists only at the membrane
this membrane potential influences the movement of ions
ions have both a chemical and an electrical gradient
no membrane potential only the concentration of ions matters
with a typical membrane potential, inward movement of cations is assisted by the voltage; inward movement of anions is resisted by the voltage
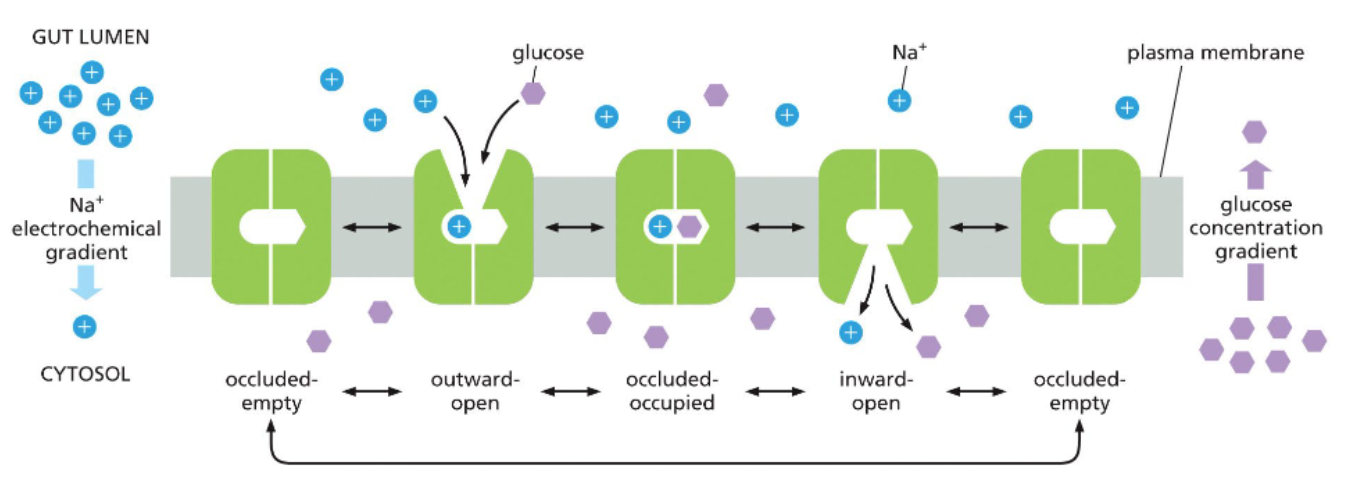
Na+ glucose symporter facilitate glucose uptake by cells of the intestines
passive glucose transporters would not allow glucose to accumulate in the absorptive epithelial cells
use Na+ electrochemical gradient to transport glucose
to maintain a high conc of glucose in the cells, glucose uptake is couple to inflow of sodium, which follows its conc gradient
the transporters won’t face the cytosol until both sodium and glucose are bound
the Na+ gradient is maintained by an ATP-powered sodium pump
secondary active transport involves using ATP to establish a gradient that is used to move another solute against its gradient
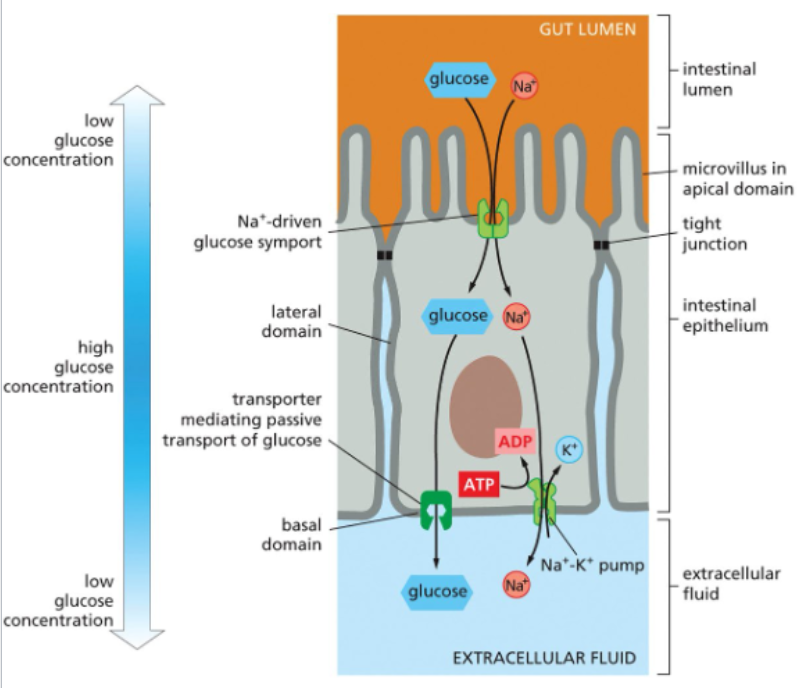
different membrane domain-restricted glucose transporters in the gut
the Na+ glucose symporter brings glucose from the intestinal lumen into the absorptive cell
tight junctions limit transporter diffusion and keep them on the apical side of the cell
glucose is also low in the extracellular fluid so passive glucose transporters on the basolateral surface are sufficient to move glucose out of the cells
Na is also moved out the cell by a Na+ K+ pump, which prevents Na accumulation within the cell
an almost identical mechanism moves amino acids from the gut to the extracellular fluid
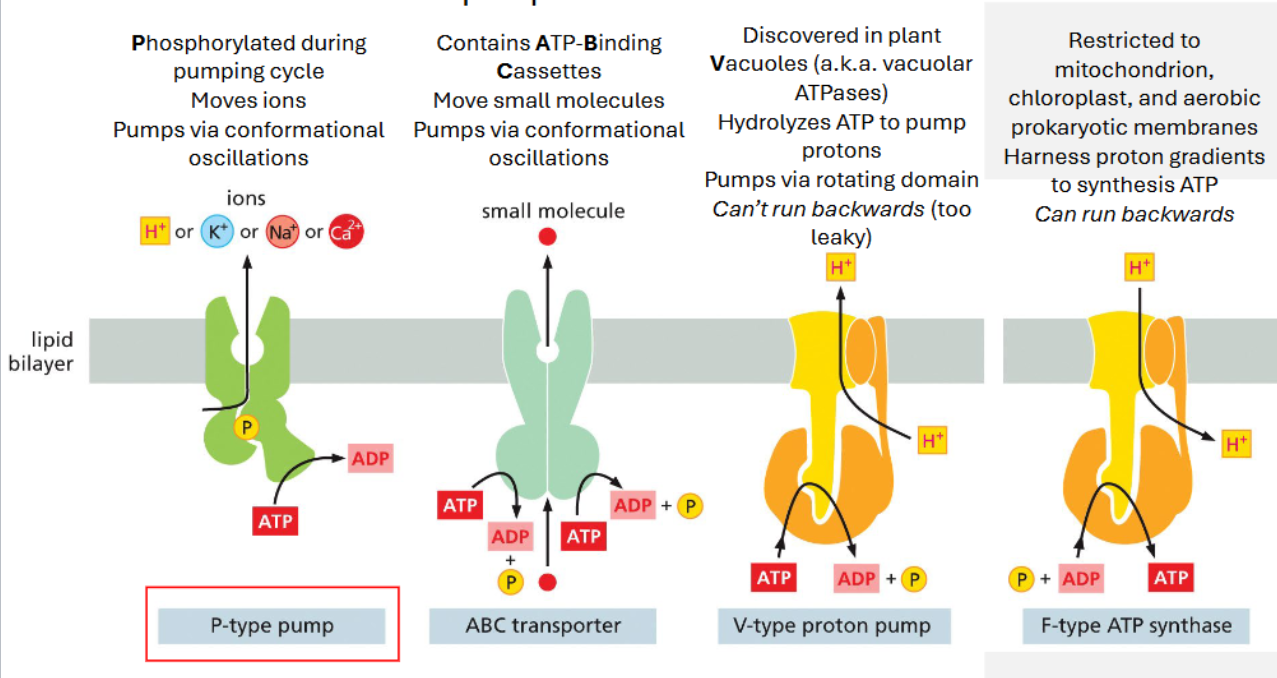
ATP-driven pumps take one of three forms
P-type pump: phosphorylated during pumping cycle | moves ions | pumps via conformational oscillations
ABC transporter: ATP-Binding Cassette | move small molecules | pumps via conformational oscillations
V-type proton pump: discovered in plant vacuoles | hydrolyzes ATP to pump protons | pumps via rotating domain | cannot run backwards (too leaky)
F-type ATP synthase: restricted to mitochondrion, chloroplast, and aerobic prokaryotic membranes | harness proton gradients to synthesize ATP | can run backwards
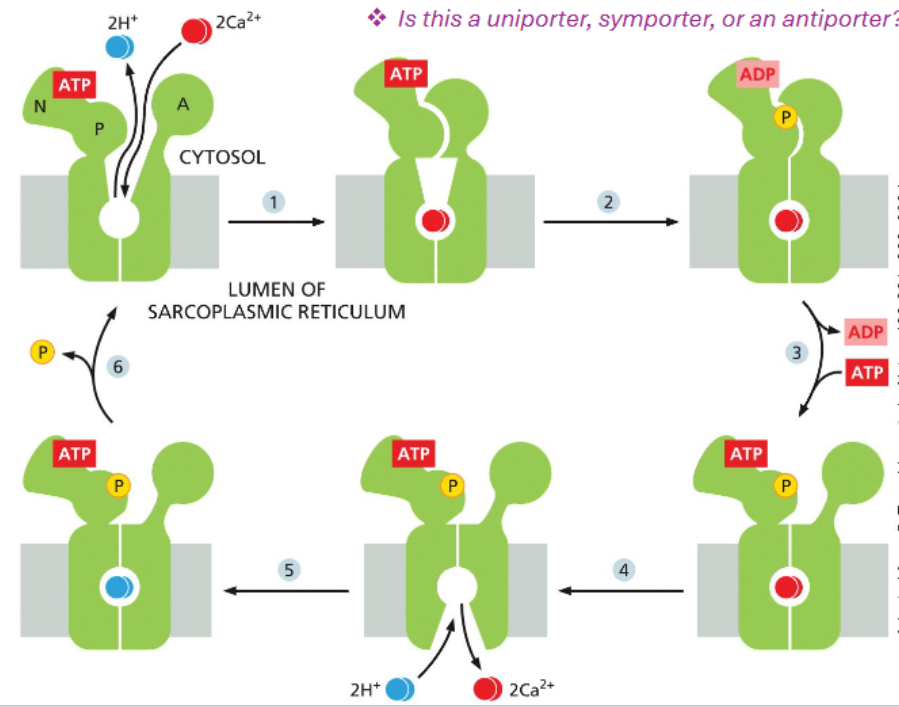
SERCA: the sarcoplasmic reticulum Ca2+ transporter (P-type pump)
SR is a Ca2+ rich modified ER that is found in muscle cells
muscle contraction is governed by rapid increases in cytosolic calcium conc due to opening of Ca2+ channels on the SR membrane
contraction is terminated by movement of Ca2+ back into the SR by ATP-powered pumps
SERCA is a family
different family members can be found in different cell types
conformational cycling of SERCA
SERCA moves two protons into the cytosol in exchange for moving 2 Ca2+ ions into the SR/ER lumen
1. In the ATP-bound, unphosphorylated state, SERCA is open to the cytosol, where it can bind calcium.
2. Calcium binding promotes ATP hydrolysis and phosphorylation of a regulatory aspartic acid that converts SERCA to an occluded state.
3. ADP is exchanged for ATP, but the protein
remains phosphorylated.
4. The ATP-bound phosphorylated SERCA
opens to the SR/ER lumen where calcium is
released and protons bound.
5. Bound protons shift SERCA conformation
and promotes removal of the phosphate.
6. Phosphate release opens SERCA to the
cytosol.
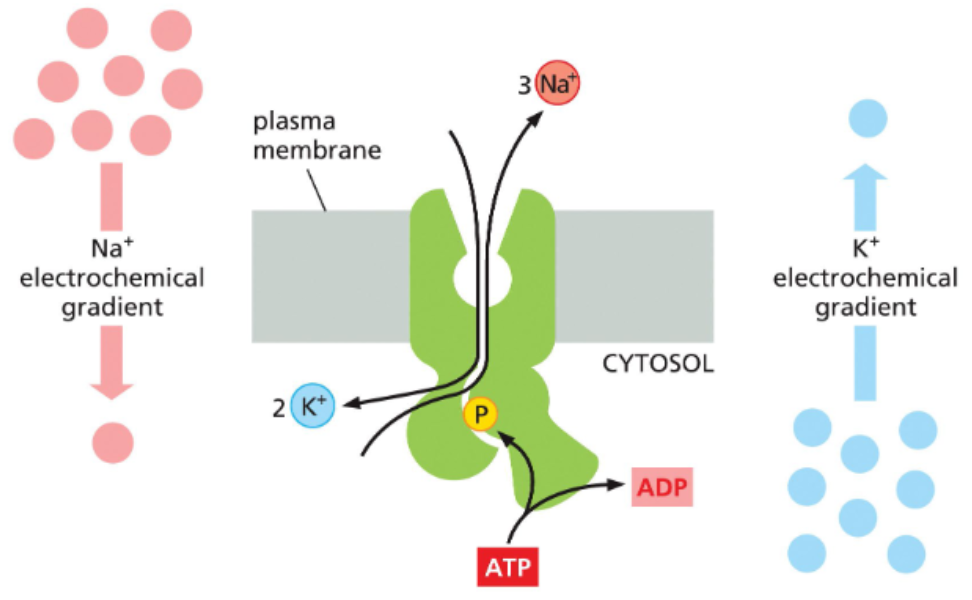
Na+ K+ pumps use ATP to maintain gradients of both ions
P-pump
Na+ K+ pumps hydrolyze ATP to move both Na+ and K+ against their electrochemical gradients
cells with these pumps maintain reciprocal gradients of Na (higher outside) and K (higher inside)
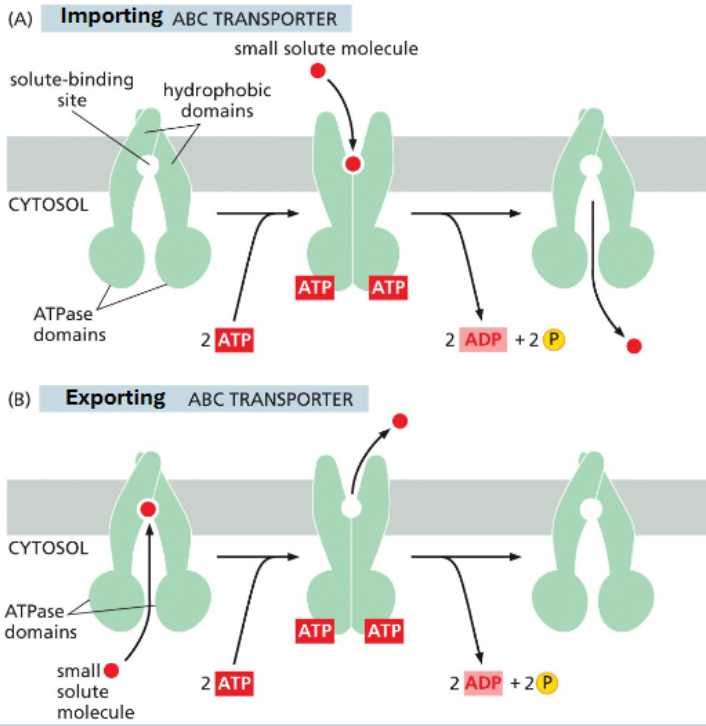
ABC transporters
ABC transporters are the most numerous and diverse
bind ATP but aren’t phosphorylated
importers & exporters vary based on whether solute binding precedes or follows ATP binding
most eukaryotes have only exporters; prokaryotes have both
alternate between openings regulated by ATP binding
ATP bound — faces cytosol
ATP unbound — face periplasmic space
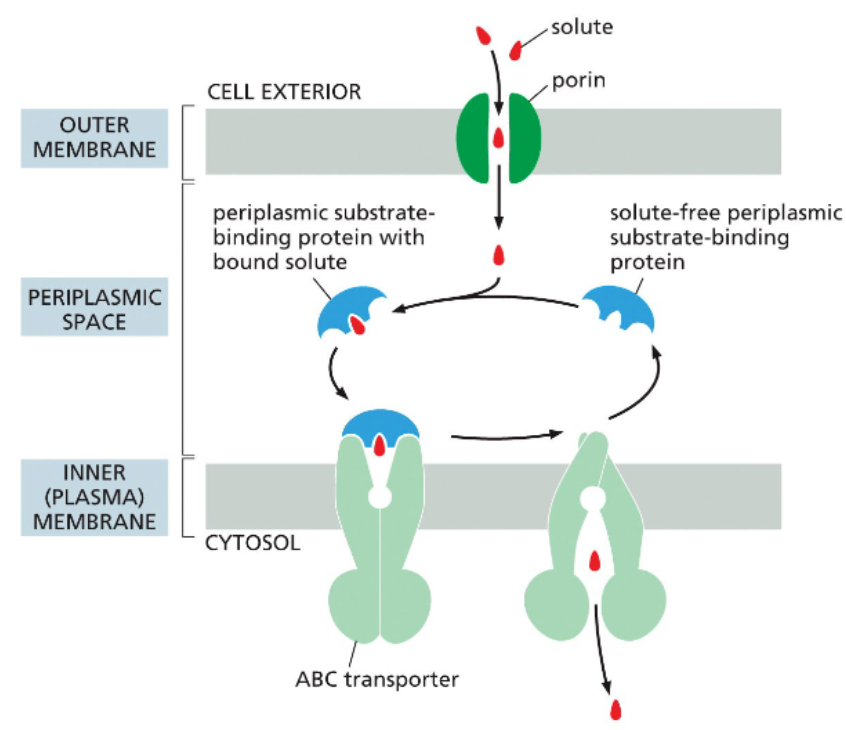
gram-negative bacteria have a double membrane with a periplasmic space
the space between (periplasmic space) contains a porous network of peptidoglycan
gram-positive bacteria have a single membrane but a thicker peptidoglycan layer that stains dark purple with crystal violet
gram-negative bacteria use auxiliary transport in conjunction with ABC transporters
Movement of materials into gram-negative bacteria requires cooperation between non-selective porins on the outer membrane, ABC transporters on the inner membrane, and a substrate binding protein that moves between them
The spacing of peptidoglycan in the periplasmic space is wide enough to allow free diffusion
ABC transporters can contribute to drug resistance by pumping toxins out of the target cell
multi-drug resistance (MDR) protein is upregulated in many cancer cells and contributes to resistance to chemotherapy