Chapter 7: T Lymphocyte Development
Primary Lymphoid Tissue = Thymus
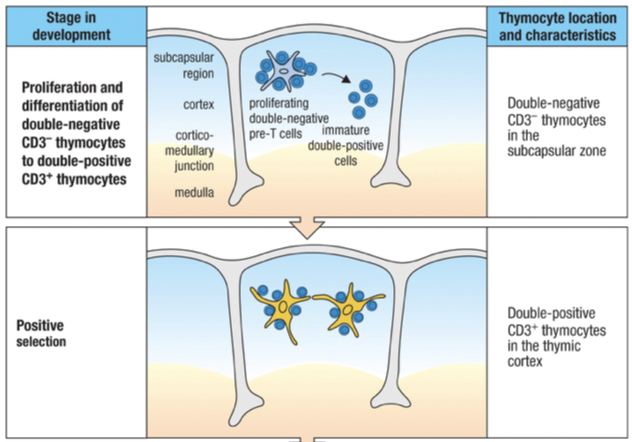
Major function of the thymus function is to ensure that mature T cells that leave thymus are restricted to the particular MHC class expressed by an individual person (self -MHC)
Two selection processes:
Positive selection leads to the death of immature T cells having receptors that do not interact with any self-MHC class I and II
Negative selection induces the death of those immature T cells that are autoreactive (receptors bind too strongly to a self-MHC molecule)
Mature T cell leaving the thymus to circulate in the secondary lymphoid organs is:
Rendered tolerant of self-antigens
Responsive to foreign antigens
Ready to fight infection
Development of T cells in Thymus
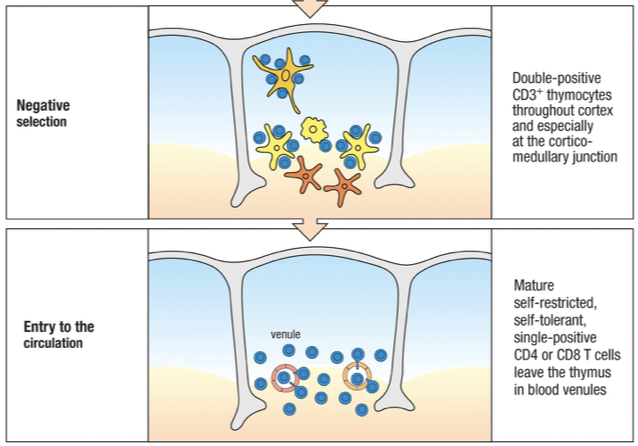
T cells originate from bone marrow as stem cells
Emigrate to mature in the thymus
Named thymus-dependent lymphocytes T cells
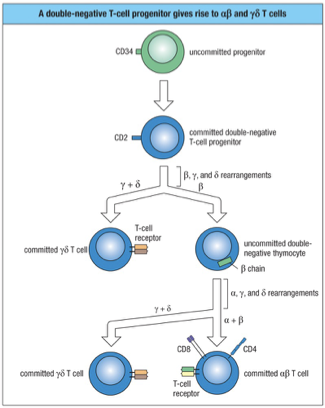
2 lineages develop in parallel from common precursors
Majority are : cells
Minority are : cells
T cells also express other cell-surface proteins related to their eventual functions
Examples are CD4 (for CD4 cells) and CD8 (for CD8 cells) glycoproteins
Mature T cells then leave the thymus in the blood and enter the secondary lymphoid organs to get activated, such as the spleen or lymph nodes.
In the absence of activation mature T cells recirculate between the blood, the secondary lymphoid tissues, the lymph and the GALT.
Thymus
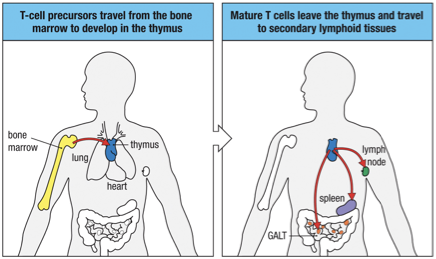
Thymus is found in the upper anterior thorax above the heart
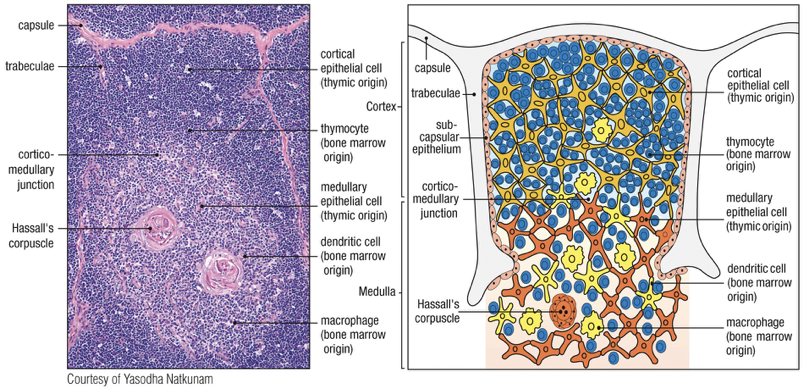
Immature T cells - called thymocytes - are embedded in epithelial cell network called the thymic stroma
Thymus is primary lymphoid organ:
Involved in the development of T cells
Not involved in lymphocyte recirculation via lymph
Blood is the only route through which T cells enter and exit
Embryonic development of thymus
Epithelial cells of cortex outer
Epithelial cells of medulla inner
Rudimentary thymus called thymic anlage is colonized by cells from bone marrow
Progenitor cells thymocytes & dendritic cells populate medulla
Bone marrow derived macrophages also populate medulla (also macrophages scattered throughout the cortex of thymus)
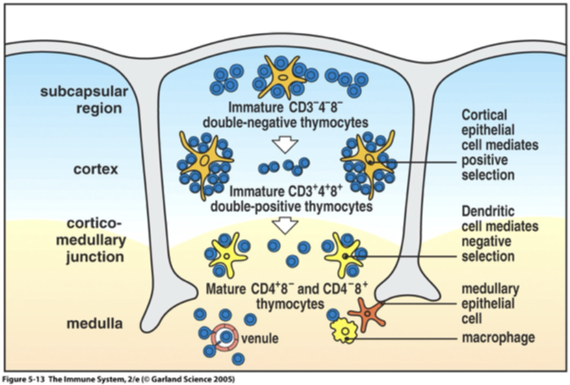
Thymocyte mature progressively move from top to bottom; outer subcapsular region to the inner cortex and the medulla
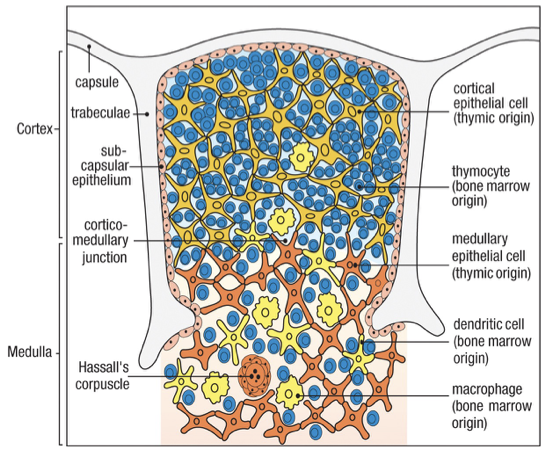
Two regions
Cortex: Immature thymocytes, Cortical epithelial cells, Macrophages
Medulla: Mature thymocytes, Medullary epithelial cells, Dendritic cells, Macrophages
Staining of cortex is dark vs lighter stained medulla
Hassall’s corpuscles
Characteristic feature of the medulla
Macrophages in both cortex and medulla remove the many thymocytes that fail to mature properly
Sites of cell destruction
DiGeorge Syndrome
Characterized by a deletion on chromosome 22, leading to potential congenital absence of the thymus (complete DiGeorge syndrome).
This condition results in disrupted T cell maturation and manifests as severe combined immunodeficiency (SCID-like state).
Symptoms may include cardiac defects, palatal problems, and immunodeficiencies due to reduced or absent T cell populations.
Immunological Aging Factors
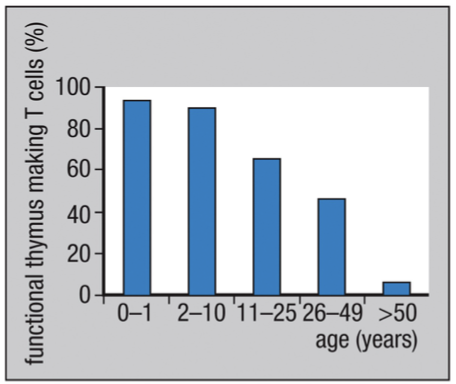
Thymus fully developed before birth; is most active in the young; atrophies with age
Progressively shrinks, fat gradually claiming areas once packed with thymocytes = involution of the thymus
Reduced production of new T cells due to aging does not noticeably impair T cell immunity
Once established, the repertoire of mature peripheral T cells is long lived and/or self-renewing
Differs from the mature B-cell = shorted lived cells that are continually being replenished from the bone marrow
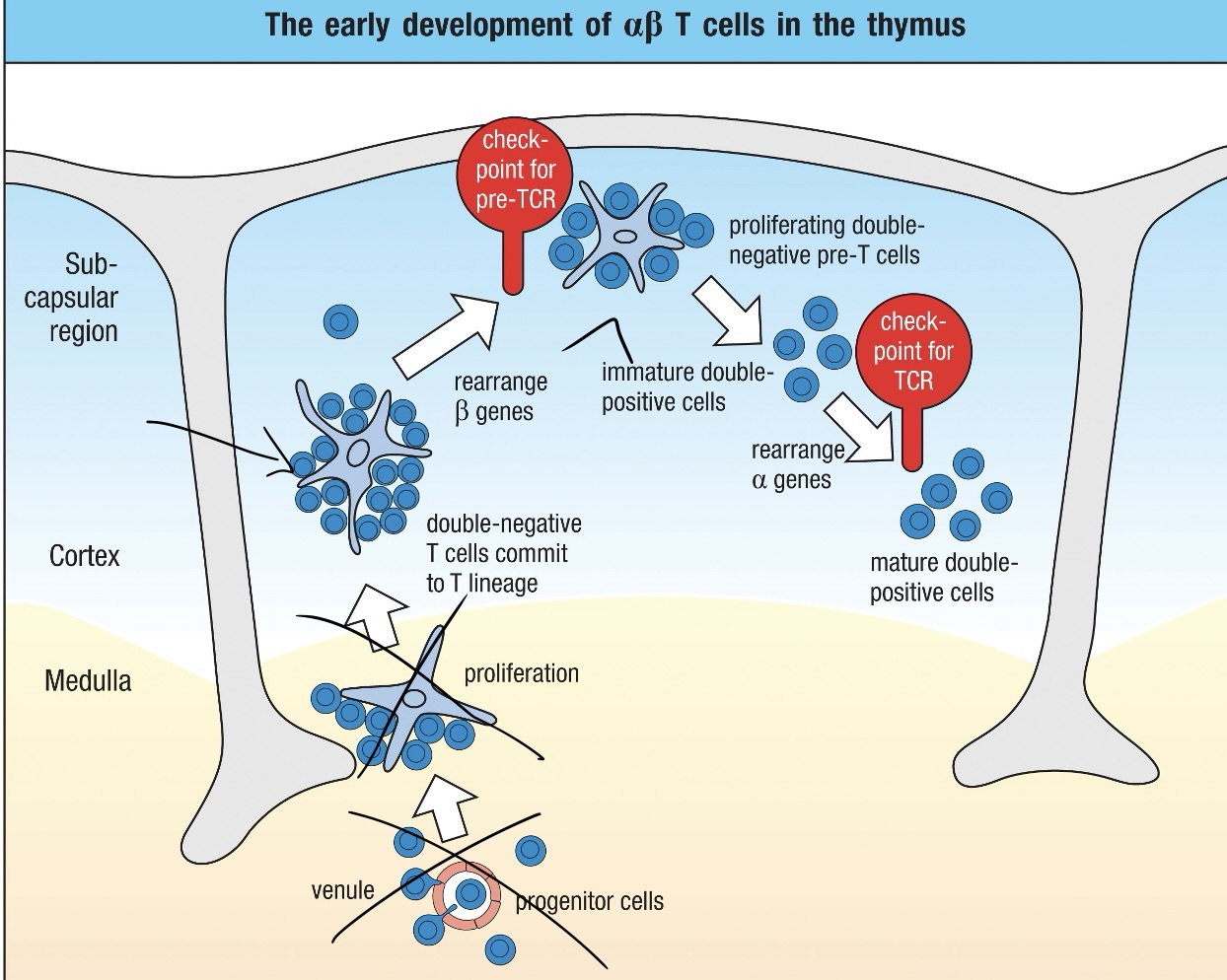
T Cell Maturation Process
Entry as Stem Cells: Progenitor cells enter the thymus.
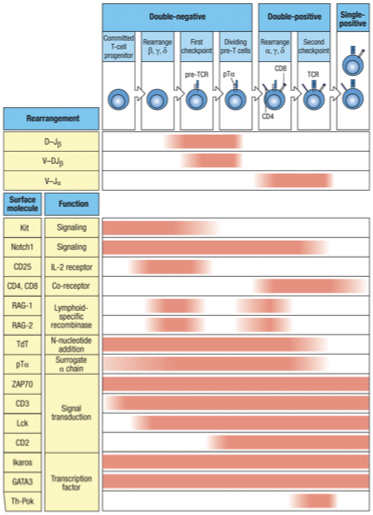
Double Negative Stage: Thymocytes lack both CD4 and CD8 markers at this stage.
Rearrangement: TCR genes undergo rearrangement, with competition between gamma-delta and alpha-beta receptors. The failure of gamma-delta rearrangement leads to Alpha-Beta TCR development.
Double Positive Stage: Thymocytes express both CD4 and CD8 while testing their TCR's ability to bind to MHC molecules. Successful binding promotes survival, while non-binding leads to cell death.
Positive Selection: Thymocytes that can recognize self-MHC molecules receive survival signals.
Single Positive Stage: After selection, thymocytes downregulate either CD4 or CD8, becoming either CD4+ helper T cells or CD8+ cytotoxic T cells.
Mature T Cell Release: Successfully matured T cells exit the thymus and enter circulation, ready to coordinate immune responses.
Thymocyte Progenation
CD3: Part of the T cell receptor complex, essential for T cell activation.
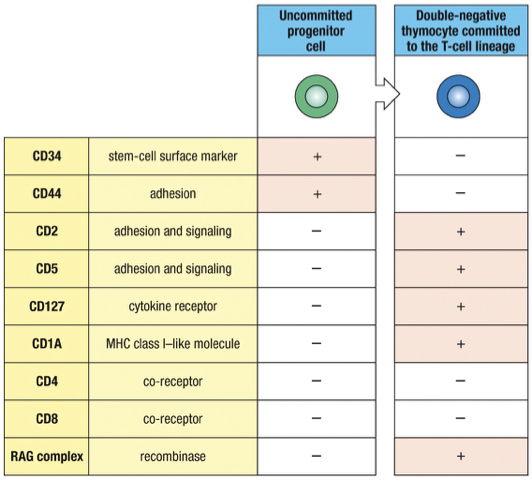
Progenitor Stem cells with TCR genes in germline configuration are marked by:
CD34 as a surface marker
CD44 for adhesion
Approximately one week later, progenitor T cells will express the T-cell specific adhesion molecule CD2 and other surface markers such as CD5 but no TCR complex
No CD4 or CD8 called “double negative” thymocytes
IL-7 receptor on T-cells is essential for binding IL-7 secreted by thymic stromal cells – helps tell the T-cell what to do next in its maturation.
Developed T cells possess specific surface receptors:
CD4 or CD8: Distinguish between helper (CD4+) and cytotoxic (CD8+) T cells.
CD2 for adhesion and signalling
CD5 for adhesion and signalling
RAG complex for RAG1/RAG2
Notch 1 Receptor Process
Notch 1 – at all stages of maturation in the thymus signals are sent through this receptor to drive the T-cell in their differentiation.
Keeps the T cell on the T cell path not allowing the BCR genes to be opened up for rearrangement
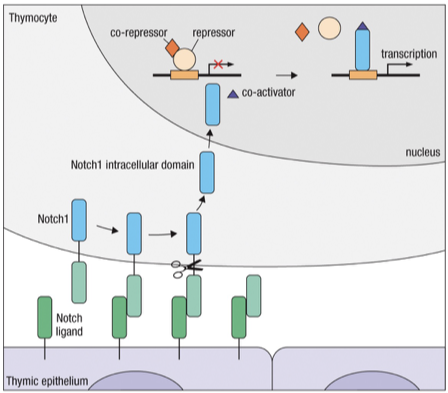
“Bind → Cut →Notch intracellular domain (NICD) → Nucleus → T cell genes ON.”
Lineage Race
-Chain assembles with a surrogate chain = pt (pre-T-cell receptor) which signals the cell to halt rearrangement of , and -chain genes and begin to proliferate
Thymocytes will rearrange their , and -chain genes at about the same time
Productive and -chain gene rearrangement made prior to a productive -chain rearrangement leads to : receptor which signals cell to stop rearrangement of chain
More frequently the chain productively rearranges before the and -chains
These are the signals if chain wins:
Stops additional TCR chain rearrangement (allelic exlucison)
Stimulates proliferation
Stimulates expression of CD4 and CD8 surface receptors
Cell allows for TCR- chain rearrangement (opening DNA)
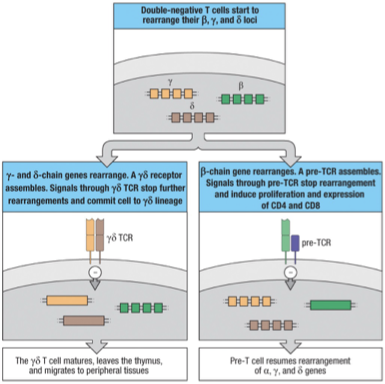
After expression of the pre-TCR, the recombination machinery is reactivated & targeted towards the chain loci (and the and loci)
In a minority of these cells, successful completion of and chain gene rearrangements occurs before the chain gene has rearranged : lineage
In a majority of cells, productive rearrangement of the -chain gene occurs first : lineage
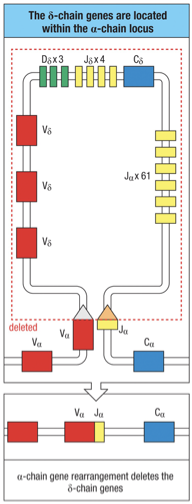
The chain locus is located within the -chain locus…a rearrangement at the -chain locus results in the deletion of the complete chain locus from the chromosome
Therefore, cells committed to one lineage can contain productive rearrangements for the TCR genes of the other lineage (except for -chain).
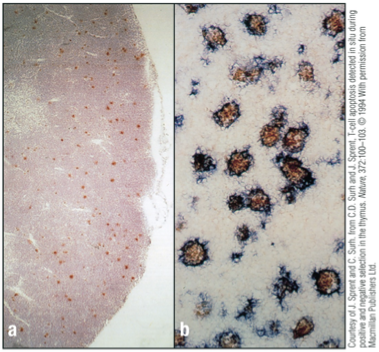
Failure/Apoptosis
Failure to make a productive rearrangement results in death by apoptosis (fate of about 98% of thymocytes)
Macrophages in thymus continually remove dead cells
Cells have been stained for apoptosis with a red dye
Apoptotic cells are scattered throughout the cortex but are rare in the medulla
Higher magnification red for apoptotic cells and blue for macrophages
Apoptotic cells are visible within macrophages
Initial TCR Production
: Receptor most abundant type found on T cells.
TCR -chain locus = variable (V), diversity (D) and joining (J) gene segments and is rearranged first (similar to heavy chain in Ig’s)
TCR -chain locus has no D segments and is rearranged after the -chain (similar to light chain in Ig’s)
Testing chain with a Surrogate -chain (preT)
Still double negative (no CD4/CD8) stage
Production of a functional -chain gene -chain translation and assembly with a surrogate -chain (preT ), CD3 proteins and chain to form a pre-T cell receptor transported to the cell surface
Role of pre –T-cell receptor is analogous to the pre-B-cell receptor in B-cell development
Triggers the thymocyte to proliferate and halt -chain gene rearrangement
Ensures only one type of T-cell receptor -chain is expressed by the T cell
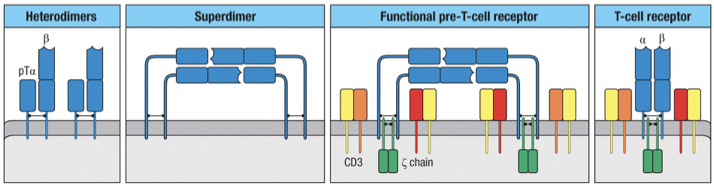
Two β-chain–pTα heterodimers can associate to form a pre-TCR superdimer, which enables ligand-independent signaling during early thymocyte development.
The superdimer cannot signal by itself, so it associates with CD3 signaling molecules on each heterodimer:
CD3γε
CD3δε
and two ζ chains
Success Signals:
Close chain (allelic exlusion)
Somatic recombination of chain
Proliferation
Double negative to double positive development (CD4 + CD8 co-receptor presence)
chain will be rearranged (somatic recombination) Checkpoint Positive selection (double positive to single positive)
Chain rearrangement attempts
Unproductive Rearrangement At One -Chain Locus Can Lead To Rearrangement Of The -chain on the homologous chromosome
Rearrangement At the Same Locus Can Also Occur
Potential to “try out” up to 4 gene rearrangements per cell
80% of T cells make successful rearrangement of the -chain gene.
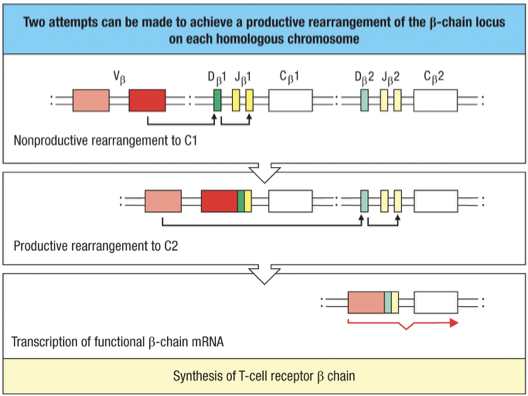
Maximum attempts = 4 (2 per chromosome x 2 chromosome)
Minimum = 2 (1 per chromosome x 2 chromosome); if V to D/J is completed
Testing chain
During cell proliferation initiated by signaling through the pre-TCR, expression of the RAG-1 and RAG-2 genes is repressed (allelic exclusion of the beta chain)
No rearrangement of the -chain genes occurs until the double-positive cells stop dividing
Ensures each cell with a productive -chain gene rearrangement produces many daughter cells that have the potential to express a different -chain gene
The TCR -chain can undergo several successive gene rearrangements
Presence of many V and over 60 J gene segments allows many successive rearrangements
Almost every developing T cell will make a productive -chain rearrangement
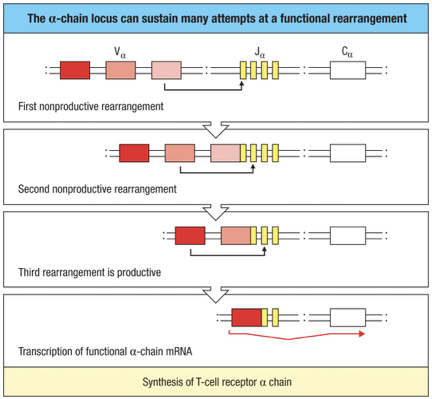
Checking for full-length alpha chains is crucial:
Concern: Possible premature stop codons can disrupt functional TCR formation.
Key Mechanism: Apoptosis serves as a default pathway when unsuccessful.
Additionally the entire chain is located within chain locus; this is where the second race between : and : occurs.
Checkpoints
Check-point 1 - Once a beta chain is produced it is tested to make sure it can bind to the surrogate alpha chain (pT ) — to ++
Check-point 2 – Once an alpha chain is made the whole TCR is tested to make sure the TCR can bind to human MHC ++ to +
“Cortex = test MHC (positive), Medulla = test peptide (negative), leave thymus = mature naïve T cell.”
Expression of proteins/surface markers
Notch 1: From beginning of maturation to release into periphery
CD4/CD8: Both turned on (double positive) after checkpoint 1, one will remain with T cell permanently based on if it binds to MHC I or MHC II
RAG1/RAG2: On only during somatic recombination in maturing T cell when chain is rearranged and chain is rearranged, turned OFF in between.
TDT: On during somatic recombination in maturing T cell, turned off when T cell is finished with somatic recombination (checkpoint 2)
pT : On during chain testing only (checkpoint 1)
CD3 complex: Stays on permanantly after checkpoint 1 (testing chain with pT)
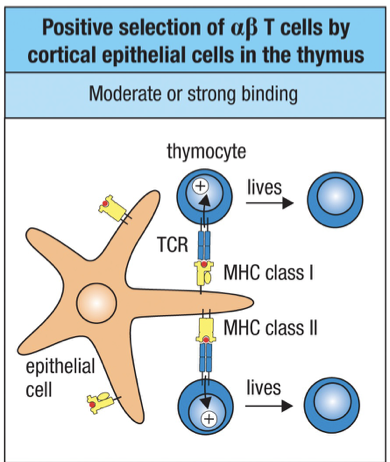
Positive Selection
After checkpoint 2 and the chain is checked Second phase of T cell development involves selection of T cells bearing TCRs that can recognize an individual’s own MHC presenting peptides.
Occurs still in cortex of thymus (moving lower down)
Over time the V gene segments which code for CDR 1 and 2 have evolved to recognize the thousands of different MHC molecules across the human population (HLA’s).
Remember that CDR 1 and 2 recognize the MHC molecule while CDR 3 recognizes the peptide.
The V gene segment chosen may not produce CDR 1 and 2 that recognize the specific MHC of that person.
This selection process involves only : T cells and not : T cells
Positive selection is the process by which this small population of T cells that reacts with the individual’s own MHC molecules is selected.
Only a small population (2%) of double positive thymocytes will be able to bind to a specific individuals MHC’s and the rest die by apoptosis in the cortex.
Positive selection is mediated by cortical epithelial cells bearing complexes of class I and class II self-MHC and self-peptides.
At the point of contact – interactions between the : TCR of thymocytes with self-MHC and self-peptide are tested.
If a peptide:MHC complex is bound by a thymocyte within 3-4 days of expressing a functional TCR, then a positive signal is delivered to the thymocyte.
A thymocyte that does not receive a signal dies by apoptosis and is removed by macrophages.
Self-peptides presented in the MHC molecules of cortical epithelial cells are derived from self-proteins present in the thymus.
The number of different peptides that can be presented by one individual’s MHC molecule is estimated to be about 10,000.
For someone who is heterozygous for the six major HLA genes about 180,000 self-peptides (18 HLA x 10000 per HLA) could be presented by 18 different MHC class I and II molecules.
For someone who is homozygous for the six major HLA genes about 60,000 self-peptides (6 HLA x 10000 per HLA) could be presented by 6 different MHC class I and II molecules
Strong Binding = live
Weak Binding = apoptosis
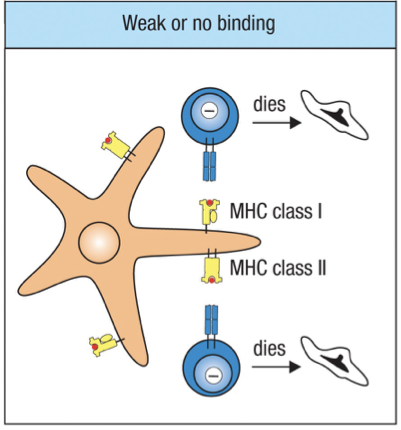
Double Positive to Single Positive
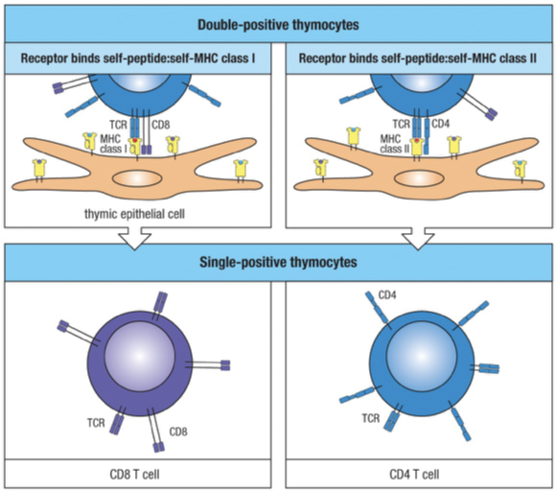
Positive selection also determines whether a double positive thymocyte matures into a CD8 or CD4 T cell, known as “single-positive” thymocytes.
CD4 T cells only interact with MHC class II molecules and CD8 T cells interact with MHC class I molecules.
During positive selection…
When a CD4 CD8 double-positive thymocyte interacts through its TCR with a class I MHC molecule, CD8 is recruited and CD4 is excluded.
When a CD4 CD8 double-positive thymocyte interacts through its TCR with a class II MHC molecule, CD4 is recruited and CD8 is excluded.
Rearrangement of the -chain locus continues throughout the 3-4 days of positive selection – hence a T cell can change the specificity of the TCR it expresses.
Once a T cell is positively selected, rearrangement of the -chain stops.
Note: Some double positive thymocytes can express two -chains (one from maternal allele and one from the paternal allele) and thus two types of TCR and undergo positive selection by engagement of one of these receptors.
The number of positively selected cells is very small; therefore, it is rare that one cell will have two selected TCRs (one receptor is usually nonfunctional).
Negative Selection
Single positive thymocyte after positive selection moves to medulla and becomes mature Thymocyte
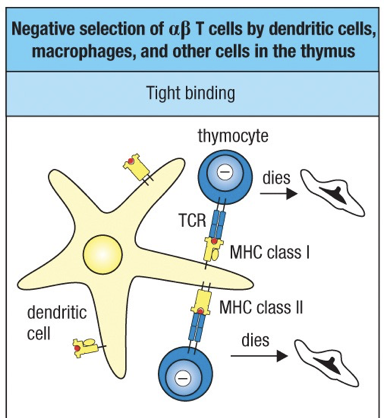
Negative selection serves to delete T cells whose antigen receptors bind too strongly to the complexes of self-peptides AND self-MHC molecules presented by thymic cells.
Central Tolerance – Tolerance to self gained in the primary lymphoid tissue
Negative selection is mediated by several cell types, most important of which are the bone marrow-derived dendritic cells and macrophages.
Engagement of the MHC molecule of one of these specialized thymic antigen-presenting cells by the TCR of a thymocyte causes that cell to undergo apoptosis and phagocytosis by macrophages.
Strong Binding = apoptosis
Moderate/Weak Binding = live
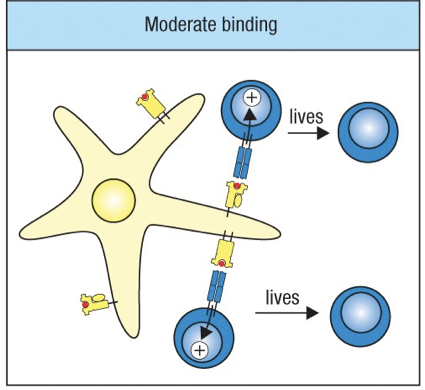
Negative selection cannot eliminate T cells bearing TCRs that can bind to self-peptides not present in the thymus (the ones in the periphery of body already)
AIRE autoimmune regulator – used by medullary thymic epithellial cells to create self peptides that are normally only found in the periphery.
Such cells enter the periphery but are rendered anergic or inactivated.
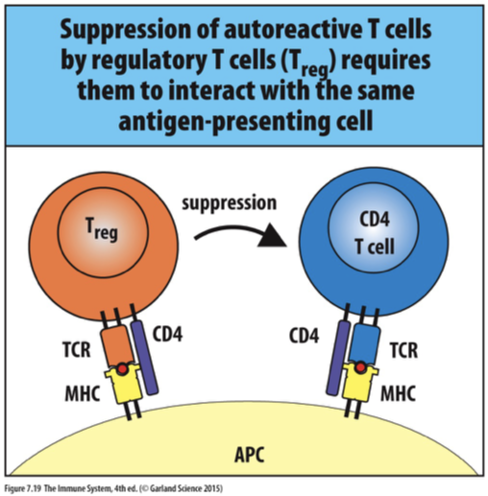
Regulatory T cells
CD4 T-cells express (Distinct from naïve T-cells)
CD25
FoxP3, a transcriptional repressor, is used by the regulatory T-cells (unique to regulatory T-cells)
Contact with MHC II – self-antigen can suppress proliferation of naïve T-cells responding to self-antigens
Suppressive effects require contact between the two T-cells and secretion of non-inflammatory cytokines released by T
IL-10: Interleukin 10
TGF-: Transforming growth factor beta
T-Cell further differentiation
The T cells that survive the selection processes in the thymus become mature, naïve T cells that recirculate through blood into the secondary lymphoid organs.
Mature T cells are longer-lived than B cells and in the absence of specific antigen stimulation will continue to circulate in the body for many years.
The T cell zones of lymphoid organs are sites where naïve T cells are activated by antigen – which provokes the final phase of T-cell development and differentiation.
Mature T cells become effector cells that can stay in lymphoid tissues or migrate to sites of infection.
There are several different types of effector T cells.
CD8 T cells become activated cytotoxic T cells.
CD4 T cells differentiate under the influence of cytokines into TH1 or TH2 helper T cells.
Which type of CD4 T cell predominates depends on the nature of the pathogen and immune response needed.
In a healthy individual there are about twice the number of CD4 T cells to CD8 T cells.
In patients with AIDS this proportion changes because the AIDS virus infects and kills CD4 T cells.