Chapter 6: B-Lymphocytes Development
Development of B Cells
The development of B cells in the bone marrow
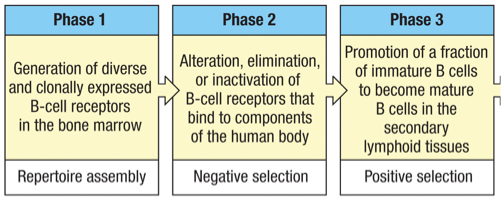
-1st of the 6 “broad stages” in the life cycle of B cells
Selection and further development of the B-cell repertoire in secondary lymphoid tissues (blood, lymph nodes, spleen, peyer’s patches)
-2nd, 3rd, 4th, 5th and 6th stages in the life cycle of B cells
1st stage = maturation in the bone marrow (primary lymphoid tissue)
Acquires functional B-cell receptors -Ig gene rearrangements
2nd stage = testing of Ig (B-cell receptor) to normal constituents of the body (self-reactive)
Potential for auto reactivity and autoimmune disease
3rd stage = a small fraction of immature B-cells become mature cells in secondary lymphoid tissues
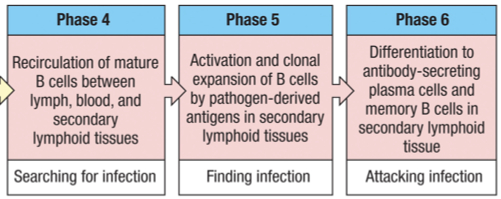
4th stage = recirculation of mature B-cells between the blood, lymph and secondary lymphoid tissues.
5th stage = antigen contact B cell progeny (clonal expansion)
6th stage = differentiate into plasma cells and long-lived memory B cells
Key Processes in B Cell Maturation
Generation (Somatic Recombination)
Occurs in the bone marrow.
Each B cell undergoes genetic rearrangements to produce a unique antibody.
Negative Selection
Removal of B or T cells that recognize self-antigens, establishing tolerance.
Critical for preventing autoimmune responses.
Positive Selection
Encouragement of cells that can recognize foreign antigens.
Final maturation signals received in secondary lymphoid tissue.
Stages of B Cell Development
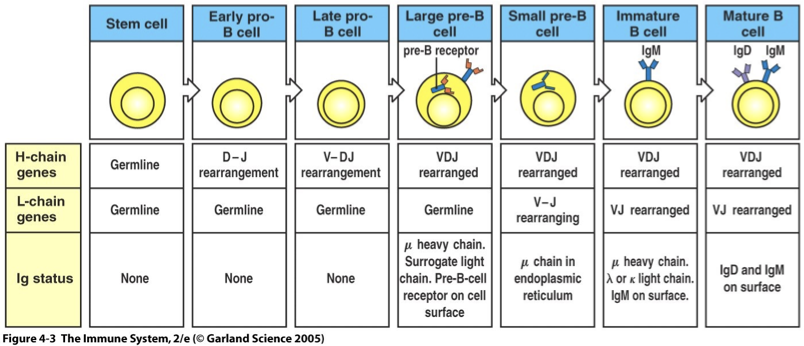
Stem cell - Ig genes are in the germline configuration
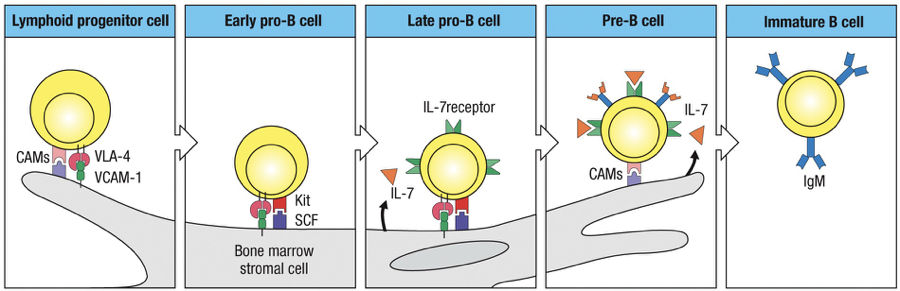
Early Pro-B Cell: Identification begins with D-J rearrangements of heavy chain
Late Pro-B Cell: V to D-J rearrangement of heavy chain occurs
Large Pre-B Cell: Identification of the heavy chain and formation of pre-B cell receptors (pre-BCR).
Small Pre-B Cell: V-J Rearrangement of light chain occurs
Immature B Cell: Successful light chain gene rearrangement and expression of IgM on the cell surface
Mature B Cell: Use of alternative splicing of heavy-chain mRNA to place IgD on the cell surface with IgM, ready for final maturation signals.
T CD4s Involved in B Cell Activation
Types of T Helper Cells
CD4T helper type 2 (Th2) usually activates B cell growth.
CD4 T-Follicular Helper (TFH) activates B cells in the follicles of Lymph nodes where the B cells congregate with Follicular Dendritic Cells (FDCs)
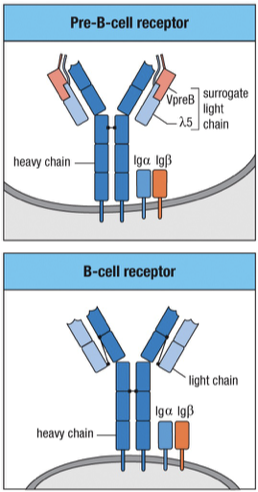
Surrogate Light Chain and Pre-B Cell Receptors
Surrogate Light Chains Components
Variable Pre-B (VpreB): Surrogate for the variable domain.
Lambda 5 (): Surrogate for the constant region, known as lambda, important for identification in exams.
Developmental Stages
Pre-B cell: Large pre-B cell stage: Cells undergo tests to ensure a proper signal Transduction process.
Following a successful rearrangement check, the cells proliferate into small pre-B cells.
Signals and Outcomes
Signals from the pre-BCR inform on the successful binding of the surrogate light chain, and outcomes include:
Stopping production of RAG1/2 somatic recombination
Tagging of the RAG2 enzyme for destruction, leading towards allele exclusion.
Heavy chain becomes less accessible (closing of DNA)
Proliferation of the B cell (rounds of division)
Large pre-B cells → proliferates → producing many small pre-B cells
Small pre-B-cells
No longer have pre-B-cell receptor
Ig heavy chains are restricted to the cytoplasm
Rearrangement of the Ig light-chain loci begins
Central Tolerance/Negative Selection
Definition
Central tolerance is achieved through negative selection, where self-reactive B cells are eliminated in the bone marrow.
Similar processes occur for T cells in the thymus
Implications of Neglecting Tolerance
Without proper tolerance, there’s a higher risk of autoimmune diseases.
Immunocompromised State: Factors such as aging affect overall immune function, leading to increased susceptibility to diseases.
B Cell Maturation Factors
B cells in bone marrow maturation interact with stromal cells which support maturation through growth factor signals like KIT, SCF and IL-7.
Kit on the B cell binds to → stem-cell factor (SCF) on the stromal cell
Activation of Kit causes the B cell to proliferate
B cells at later stage of maturation require interleukin-7 (IL-7) to stimulate their growth and proliferation
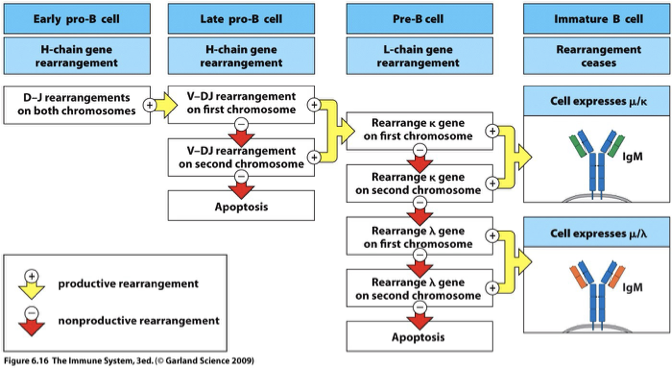
Productive vs. Nonproductive Rearrangements
Productive Rearrangement
Successful gene segments lead to functional BCRs, contributing to survival and normal function.
Nonproductive Rearrangement
Unsuccessful gene segments that cannot lead to BCR
The two copies are on homologous chromosomes; one inherited from mother and one from father
Gene rearrangements can be made on both homologous chromosomes
Unproductive rearrangement on one chromosome leads to rearrangement at the locus on the other chromosome
Productive rearrangements → proceed to the next stage of development
If all rearrangements are unproductive → B cell does not produce Ig and dies in the bone marrow
Apoptosis
ONLY Final nonproductive rearrangements lead to cell apoptosis
Heavy chain rearranged (Late pro-B cell) → large Pre-B cell → Light chain rearrangement → Light chain rearrangemet → Large Pre-B cell
2 copies of each (homologous chromosomes) x 2 light chain types ( and ) = 4 MINIMUM potential light chain combinations
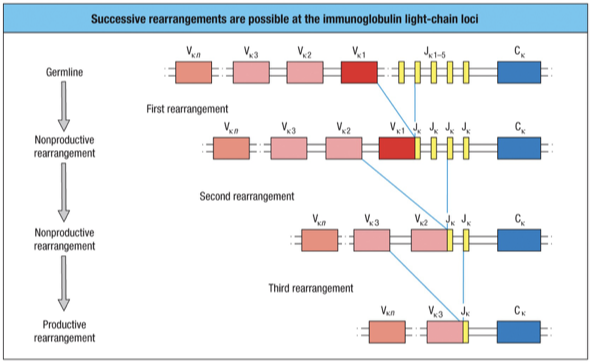
Further rearrangement within L-chain Loci
After an unproductive rearrangement of V to a J, a second rearrangement can be made by any other V that is on the 5’ (left) side of the first joint with a J that is on the 3’ (right) side of the first joint
When the second joint is made, the intervening DNA containing the first joint will be excised
There are 5 J gene segments and many more V gene segments
five successive attempts at productive rearrangement of the L-chain gene on a single chromosome
Success → Stop Mechanisms
Successful gene rearrangement is signaled by the appearance of the protein product of the gene at the cell surface by Ig and Ig
A signal is sent back to the cell interior to shut down the processes of DNA recombination and repair needed for gene arrangement
RAG genes are turned off and no further rearrangement is possible
Once heavy-chain gene is successfully rearranged also further rearrangement of heavy-chain genes is shut down
Leads to allelic exclusion = same (homogenous) strong product BCR = high avidity
Same process happens with light-chain
Possible outcome if RAG-1 and 2 aren’t shut down after a productive rearrangement in a B-cell → no allelic exclusion giving heterogenous BCRs with low avidity binding
In Late pro-B cells, (IgM gene) heavy chains assemble into dimers in the ER
Since this is before light chains, heavy-chains assembled into a complex with the 5 and the VpreB polypeptides to form a surrogate L-chain along with Ig and Ig and is called a pre-B cell
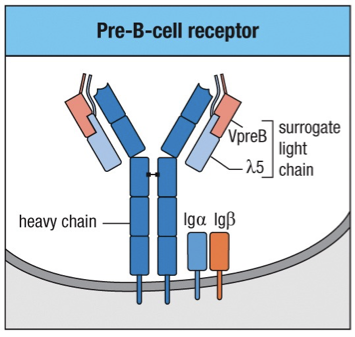
Checkpoints in Development
Major checkpoints exist during B cell maturation:
Checkpoint 1: Tests heavy chain rearrangement at Large Pre-B Cell
The Pre-BCR’s form dimers and oligomers that provide signals to allow maturation to continue.
Turns expression of RAG’s off, tags RAG2 for destruction and closes the DNA down = Allelic exclusion at the heavy chain locus
Leads to a positive signal that prevents apoptosis and allows B cell to start dividing.
After checkpoint, pre-BCR is no longer made, heavy chain, Ig and Ig continue to be made in ER
Result: RAG genes (turned off in dividing Large pre-B cells) are turned on → light chain rearrangement begins
Checkpoint 2: Tests light chain rearrangement at Small Pre-B Cell; can lead to cell death if there is non-rearrangement failure.
On completion of a productive light chain gene rearrangement a light chain protein is made and assembles with heavy chain to form IgM
IgM (BCR) associates with Ig and Ig and is transported to the cell surface
Result: Presence of the BCR with Ig and Ig tells the cell to halt further light-chain gene rearrangements
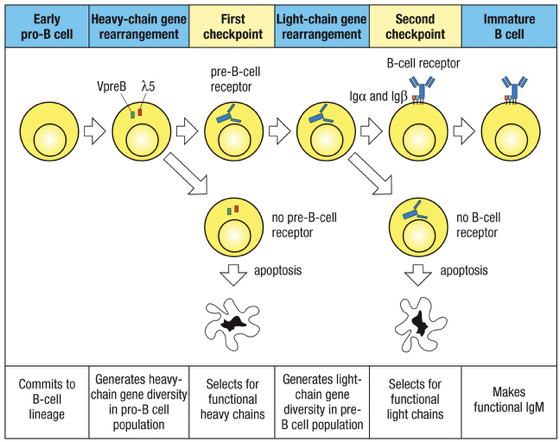
Changes in Key Protein Expression
The rearrangement of Ig genes and the expression of pre-BCR and IgM on the cell surface requires several categories of specialized proteins at different times during B-cell development
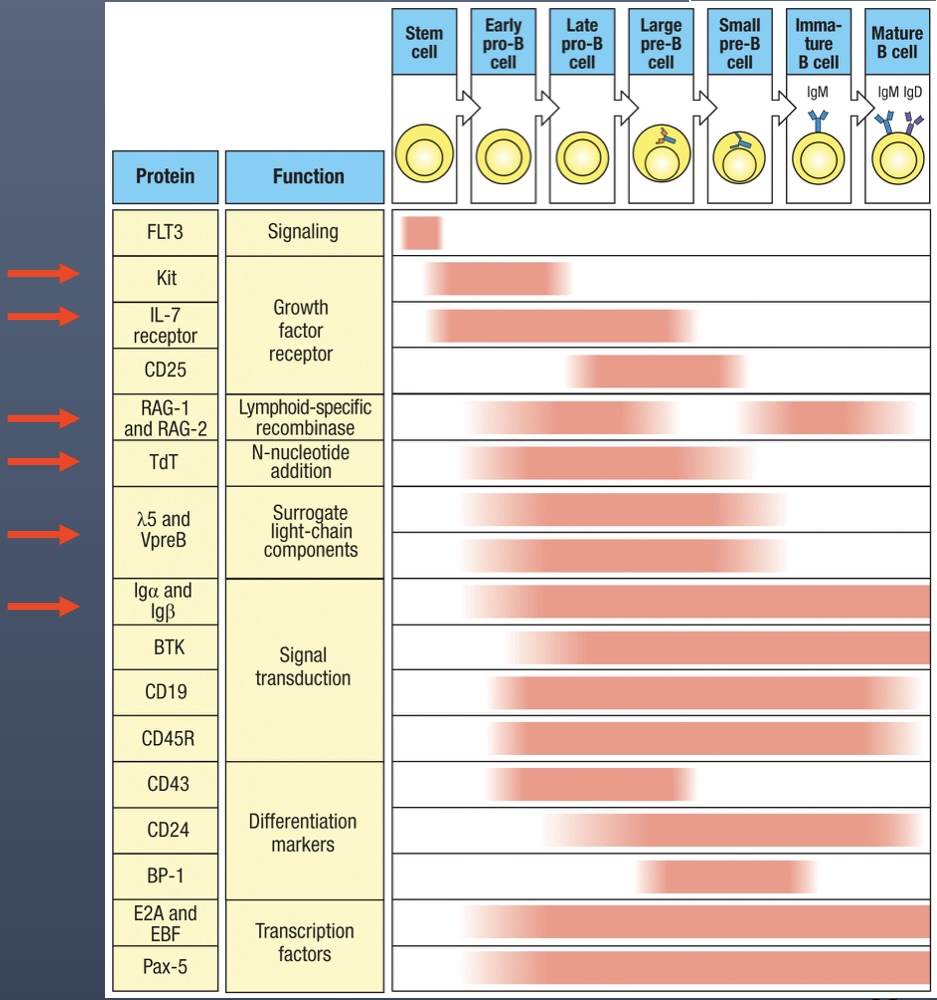
KIT - present only in stem cell development in bone marrow
IL7 Receptors - present in stem cell development and in Early pro-B Cell
RAG-1/RAG-2 - present in Early pro-B cell and Late pro-B cell (for VDJ recombination of the heavy chain), then turned off, and then turned back on in Small pre-B cell
TdT - Stays on from Early pro-B cell (beginning of heavy chain rearrangement) through Small pre-B cell (rearrangement of light chain)
5 and VpreB - Only active in the Large pre-B cell with the pre-B cell receptor and surrogate chain
Ig and Ig - present on B cell surfaces after Late pro-B cell (heavy chain complete)
Translocations and Cancer
Translocations occur only during the first attempt to rearrange a heavy-chain gene
This would have counted as an unproductive rearrangement and the other gene would then be rearranged
In cases where the 2nd rearrangement is also unproductive, the cell dies and thus cannot give rise to a tumor
Mechanism of Translocations
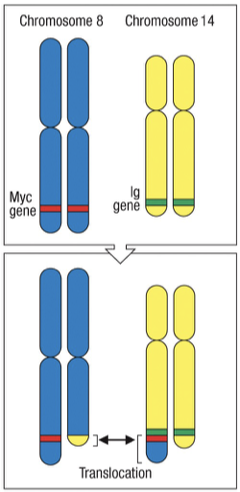
Translocations during somatic recombination can lead to oncogenes being misregulated (e.g., MYC gene involved in Burkitt's Lymphoma).
Genes from different chromosomes can affect cell cycles, leading to cancerous growth.
Key Proto-Oncogenes: Include BCL2 (protective against premature apoptosis in B cells) and MYC
Chromosomal Rearrangements In Burkitt’s Lymphoma
MYC is normally involved in regulating cell division. Abnormal expression as a result of translocation causes increased growth.
MYC Gene Location: Chromosome 8.
IG heavy Gene Location: Chromosome 14
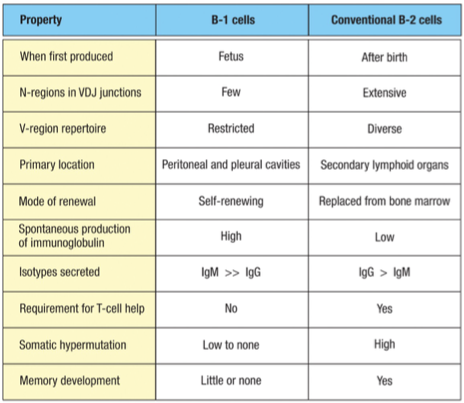
B1 vs B2 Cell Types
B1 Cells: Innate
Primarily secretes IgM > IgG without T-cell help.
Have a role in the initial immune response, acting via innate mechanisms.
Produce antibodies quickly, pre-birth and early after birth, but lack memory.
Also known as CD5 B cells
Pool of self renewing B-1 Cells established (not required to be in bone marrow)
5% of B cells in body
B2 Cells: Adaptive (regular)
Primarily secretes IgG > IgM
Require T-cell activation to produce antibodies and are involved in adaptive immunity.
Able to engage in somatic hypermutation and class switch recombination for diverse antibody production.
Have memory and are located in secondary lymphoid tissues but replaced from bone marrow
95% of B cells in body
Negative Selection for Self-Reacting B Cells
Quality control mechanisms prevent the maturation of B cells whose receptors bind normal components of the human body – called self-antigens (protein, carbohydrates, lipids found on the surface of human cells.
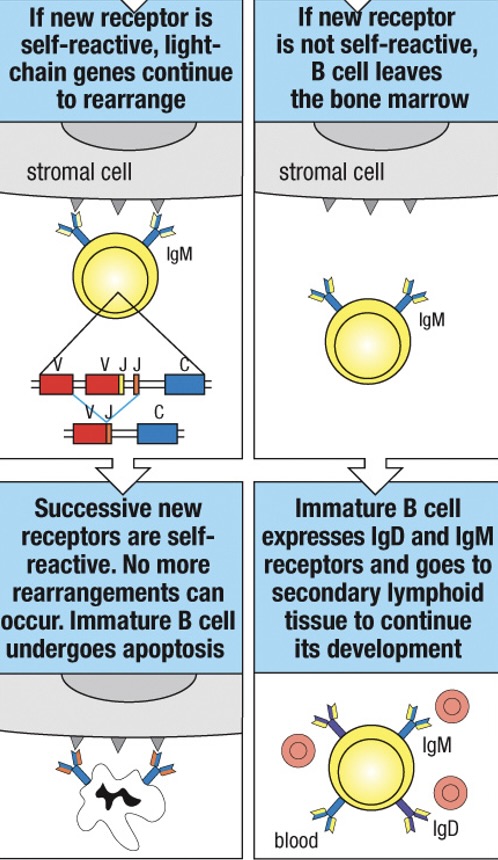
Immature B cells that do not encounter a self-antigen leave the bone marrow and enter the peripheral circulation expressing both IgM and IgD on their surface
Immature B cells that bind soluble self-antigens (monovalent self-antigens) are rendered unresponsive or anergic (or unresponsive) to the antigen
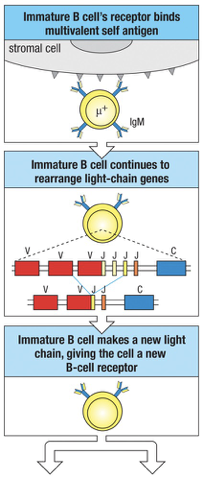
When a developing B cell produces surface IgM that are strongly cross-link by multivalent self-antigens (MHC complex molecule on cell surfaces) the B cell undergoes receptor editing
The amount of IgM on the surface is reduced and the RAG genes are not turned off
Continued synthesis of RAG proteins allow the cell to continue L-chain gene rearrangement
Usually leads to a new productive rearrangement and expression of a new L-chain which combines with the previous H-chain to form a new receptor (receptor editing)
Receptor Editing in B Cells
B cells can undergo receptor editing if they recognize self-antigens (multivalent) during maturation:
Receptor editing allows changes to the light chain of the BCR to avoid autoimmunity.
If this new receptor is not self-reactive the cell is “rescued” and continues normal development much like a cell that had never reacted with self
If the cell remains self-reactive, it may be rescued by another cycle of rearrangement but if it continues to react strongly with self it will undergo apoptosis and be deleted from the repertoire
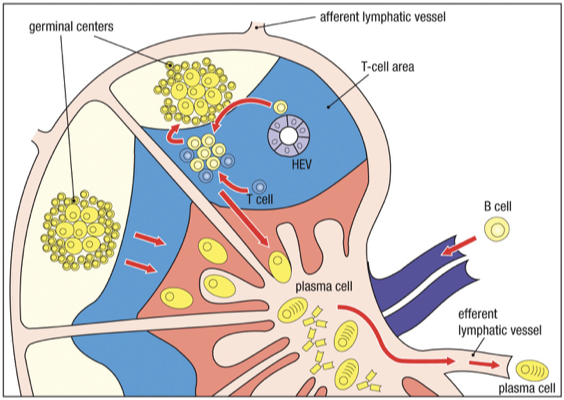
Lymph Node Circulation/Activation
In a lymph node, there are discrete sites for B cells and T cells.
Expansion of effector B cell region (plasma cells that secrete antibodies) occurs in lymphoid follicles.
As lymphocyte development proceeds, follicle shape changes - germinal center
From the bone marrow, an immature B cell (high IgM, no IgD) will migrate via the blood to the secondary lymphoid organs, directed by cytokines
CCL19 and CCL21: Guide B cells into lymph node, bind to CCR7
B cells enter the cortex of the lymph node through the wall of specialized high endothelial venules (HEV), also directed by cytokines
CXCL13: Draws B cells into follicular areas during migration, binds to CCXR5
Late immature or transitional (IgD increasing close to maturity) B cell then interacts with Follicular dendritic cells to receive their final signal to fully mature and survive.
BAFF: B Cell Activating Factor released by Follicular Dendritic Cell
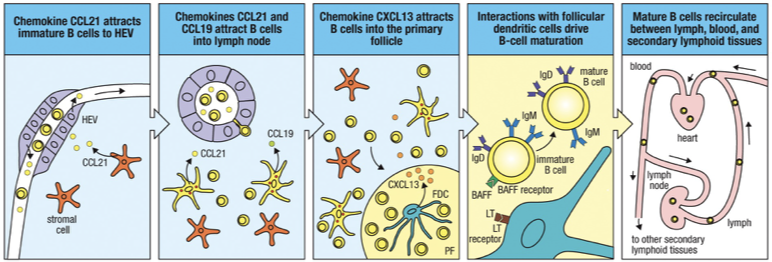
Result: Now called naïve B-cells (mature). Immature B-cells that fail to enter the follicle will die.
Secondary lymphoid tissue follicles are the sites where mature, naïve B cells encounter specific antigen (held on Follicular Dendritic Cells)
Antigen-specific B cells stay in the T-cell areas, and are activated by antigen-specific, CD4 helper T cells
The B cell binds the antigen on the FDC.
The B cell internalizes the antigen.
It processes it and presents peptide on MHC II.
The B cell then moves to the T-B border to get help from CD4 T cells.
CD4 T cells (TFH and TH2) provide signals that activate the B cells to proliferate and differentiate
Both Th2 and T-Follicular Helper (TFH) cells secrete IL-4, which drives isotype switching to IgE
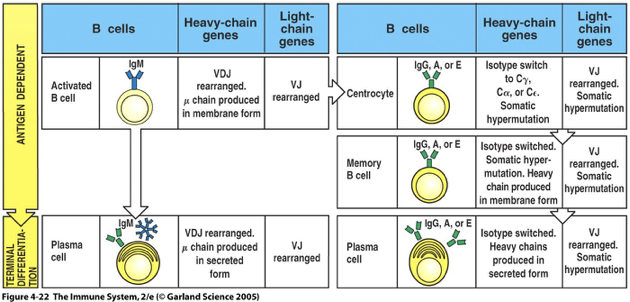
Somatic Hypermutation and isotype switching happen after B cells receive help from CD4 T cells
In lymph nodes and spleen some of the activated B cells immediately proliferate and differentiate into plasma B cells and secrete antibody (lower affinity)
Other activated B cells migrate to a primary follicle that matures into a secondary follicle containing a germinal center
Primary Follicle (Before Infection)
When a B cell recognizes antigen and receives help from CD4 T cells, the follicle becomes activated Secondary Follicle Forms (with germinal center) and undergoes proliferation
Germinal Center
Dark Zone(dark stains): B cells = centroblasts
proliferation,
somatic hypermutation (higher/lower/same binding site affinity)
isotype switching
Light Zone (light stains): B cells = Centrocytes and also containes Follicular Dendritic Cells
B cells that mature into non-dividing centrocytes that have undergone isotype switching and somatic hypermutation
Retesting BCR
Centrocytes compete for original antigen on FDCs to test which one is best
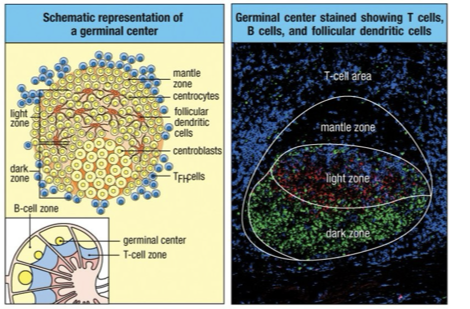
Selection After Affinity Maturation
Cells that survive the selection process after affinity maturation undergo further proliferation and migrate from the germinal center to other sites in the secondary lymphoid tissues or bone marrow
Selected B cells differentiate into plasma cells secreting high affinity, isotype-switched antibody
As immune response subsides, germinal center B cells develop into memory B cells capable of making high affinity antibody when re-exposed to the same antigen (basis of secondary immune response)
At end of infection, less antigen is available. Memory B cells have the strongest affinity Igs on their surface
B Cell Population Dynamics
No follicle visit die (typically less than a week)
Follicle visit unless stimulated by specific-Ag die (mature B cell can live for weeks)
B Cell Half Life: 3-8 weeks for B cell
B cell has to constantly get BAFF signal in order to survive and maintain its population within the immune system.
This critical signal ensures that B cells continue to proliferate and differentiate, allowing for a robust and responsive immune response during infection
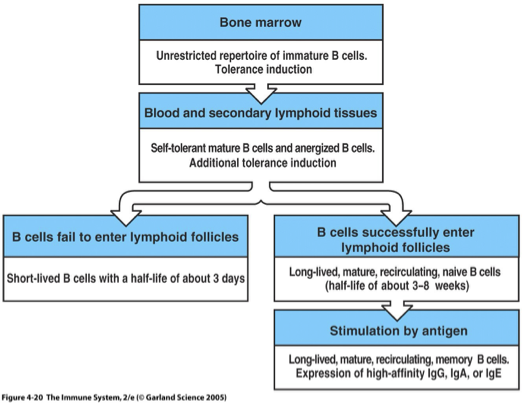
As infection ends
Most B cells die of apoptosis as amount of antigen lowers
Some remain as memory cells, providing long-lasting immunity by quickly responding to future infections with the same antigen
Adaptive Immune Responses of Mature B Cells
3 ways to differentiate into Plasma B cells (antibody secreting)
Primary Adaptive Immune Response
Activated B cells differentiate into plasma cells which produce only IgM antibodies specific to the encountered antigen, thereby initiating the body’s defense mechanism.
Centrocytes (post somatic hypermutation and isotype switching) also differentiate into plasma cells
Secondary Adaptive Response
Memory B cells differentiate into plasma cells which produce antibodies specific to the remembered antigen