PS219 TOTAL NOTE
L1 - The Neuron
1. Overall Objective
To understand human behaviour as resulting from an intricate system of chemical checks & balances
as opposed to: ‘understand it in categorical – or even worse: dichotomous – terms’
example (non-)dichotomies:
starving - full to burst
deliriously happy - desperately sad
healthy - unhealthy (diseased)
normal - abnormal
homosexual - heterosexual
male - female
Example health/disease:
If you experience the following symptoms:
Auditory & visual hallucinations
Delusions
Disorganised thought
Emotional dysfunction
Then in what condition are you in?
2. Objective of today’s lecture:
To understand neural signalling as a system of chemical checks & balances
Why bother…?
Part 2 – Neurons & Glia
1. Neurons are special!
Function:
integrate signals from many other cells to generate electrical impulses to send rapidly & over great distances to specific target cells. The target cells then do the same.
The input from each signalling cell modifies the activity of target cells
Each part of the network controls other parts of the network
The network controls itself
In ongoing, structured communication
“A system ‘designed’ to implement careful checks & balances”
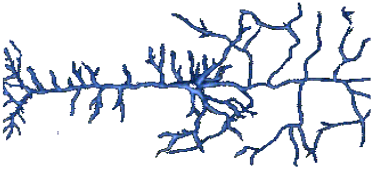
Form & Size:
Most body cells round(ish) ‘blobs’
Neuron:
Special requirements:
Virtually no possibility to store energy, but are very energy demanding: Glucose [sugar] & oxygen must be constantly supplied
Without supply, neurons:
stop working within seconds
die within minutes
Life span:
Neurons do not divide (develop once from neural stem cells)
Neurogenesis (generating new neurons) is complete 5 months after conception (before birth) - with some exceptions in some parts of the brain. This is debated.
Neuron death is part of normal brain development: 20-80% of all neurons die during maturation!
Dependent on area of the brain being looked at.
2. Glia cells
Provide a ‘protected environment’ for neurons to survive
Develop – like neurons – from neural stem cells
Approximately as any glia cells as neurons
Types (examples only):
Astrocytes: they form part of the blood-brain barrier (things cannot easily reach neurons. To get to neurons, have to go through astrocytes). Involved in physical support (keep neuron in place) and signalling
Microglia: mobile task force. Involved in defence and repair and digestion.
Oligodendroglia: Produces the myelin sheath to wrap around axons
Part 3 – Signal Transmission Within a Neuron
1. Structures of a Neuron:
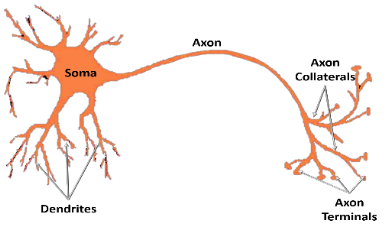
The main input area is the dendrites and soma.
The signals received from the main input areas converge at the Axon-Hillock. The Axon hillock integrates inputs. If the sum of inputs exceeds a certain threshold, it generates and releases AP along the axon to collaterals and terminals, which then release a chemical output.
There is ‘passive electrotonic transmission’ at the main input areas.
Then there is a generator potential which generates (or fails to generate) AP’s at the Axon-Hillock
Then there is ‘active AP transmission’ in the axon.
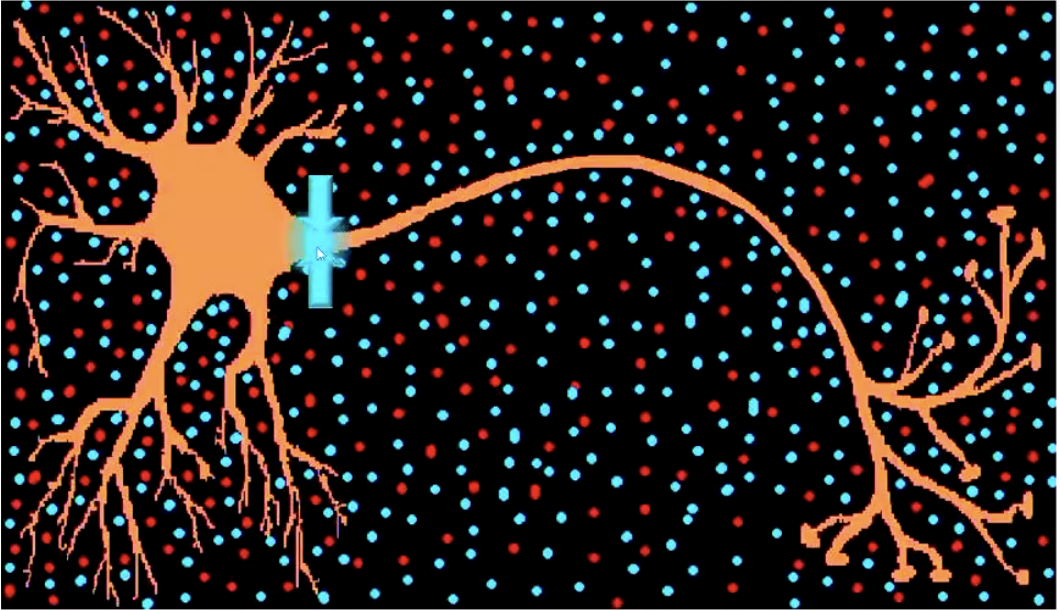
There is electrical transmission all within the neuron, and then chemical transmission between neurons.
Two types of (electrical) signal transmission:
Passive Electrotonic Transmission:
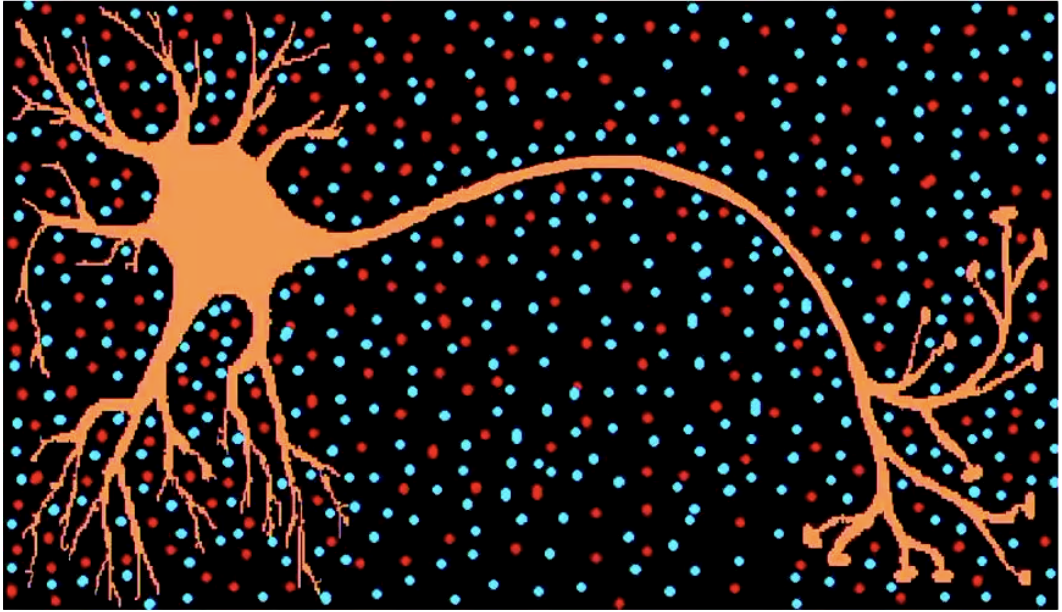
Requires electrically charged particles (ions) to move.
Passive ion transport inside the cell (dendrites & soma). Some positive ions enter, some negative ions enter and travel to the Axon-Hillock.
Decays as ions ‘get lost’ on the way
Forms generator potential - determines whether or not we get an AP.
Action potential:
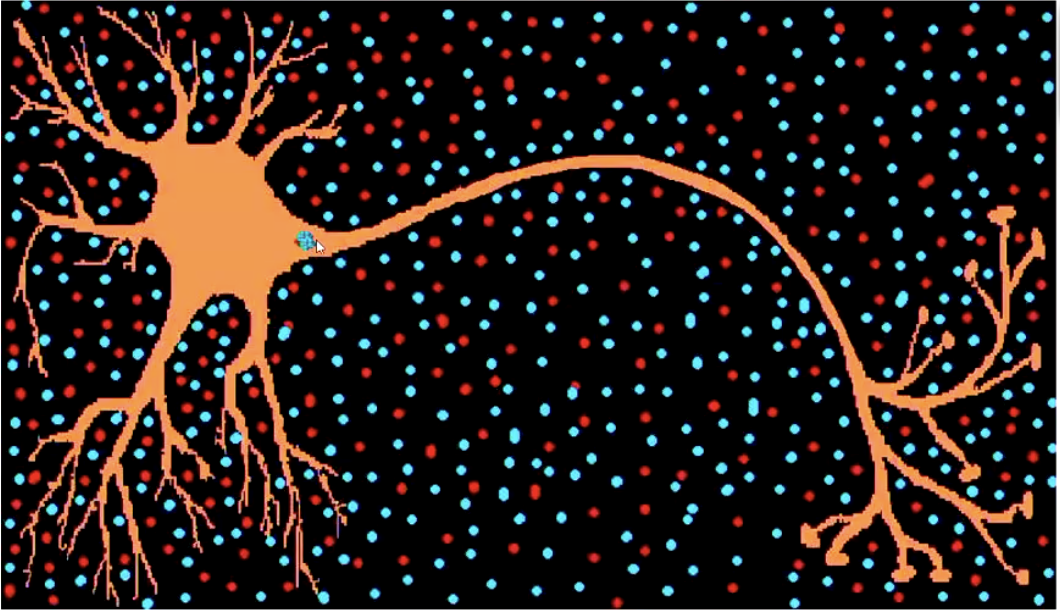
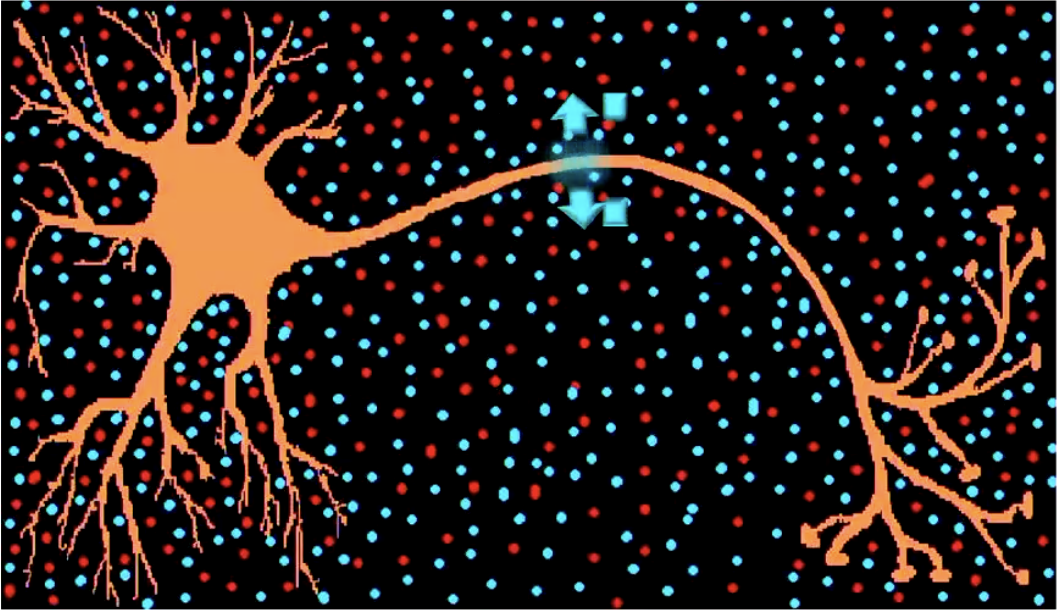
Active, self-replicating wave of depolarisation from the axon-hillock to terminal buttons. Positive ions get ‘sucked into’ the membrane, and then are ‘spat out’ again
No decay (always the same size)!
Positive ions are sucked in and kicked out again.
When the positive ions are spat out, that burst of positivity opens the gates in the adjacent area of the axon, which then pulls the positive ions in the environment back in again. This continues to happen all the way down the Axon.
3. Cell Membrane & Resting Potential
At rest (no AP fired), inside of cell membrane is more negative than outside
(typically around -60 to -70 mV)
Difference in electrical charge = electric potential (aka voltage) = this is the potential of the cell to create electrical current (just moving positive ions)
4. Electrical current = moving ions
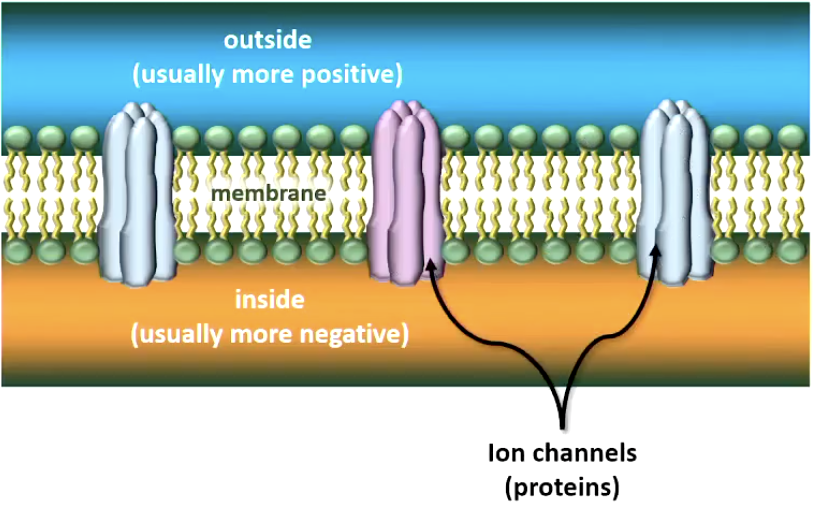
Ions can pass through the membrane via ion-specific protein channels
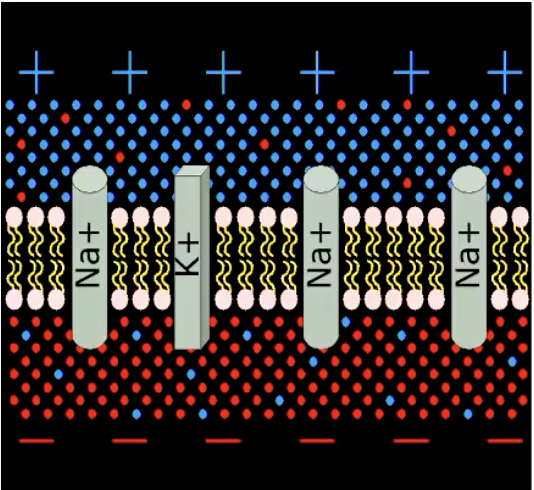
Some are always open (of no interest to us)
Some need energy to operate: “Na/K pump” pumps 3 Na+ ions out for each 2 K+ it lets in => maintains resting potential
Resting potential is energy demanding. Main reason why brain needs constant supply of oxygen and glucose.
Some channels are opened by chemical or electrical stimulation
Allowing specific ions to enter (or leave) the cell
If positive ions enter (or negative ions leave), the inside of the membrane becomes less negative than before (depolarised)
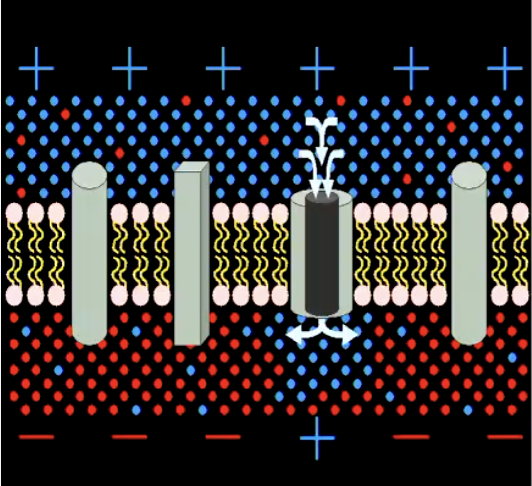
If positive ions leave (or negative ions enter), inside of membrane becomes more negative than before (hyperpolarised)
Action Potential
Getting started:
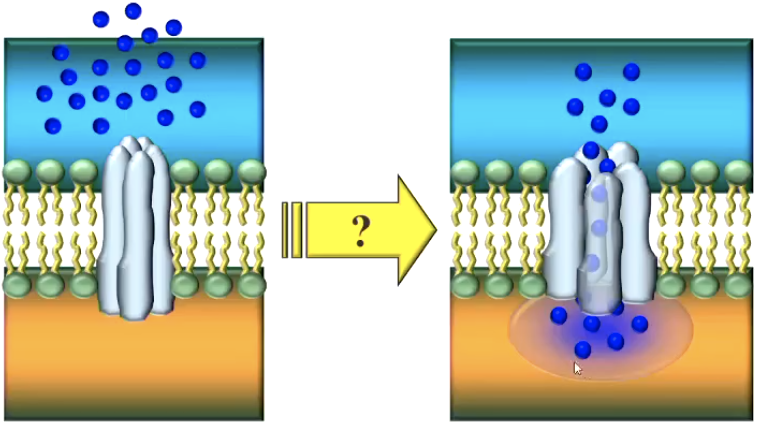
There is first an electrical charge (bunch of positive ions) on inside of membrane and arriving at ion-specific protein channels
Voltage-gated Na+ channels open and positive sodium ions flow through
Na+ ions enter the cell => Membrane depolarises
Reaching Peak Level
More Na+ channels open => If certain peak level is reached, more depolarisation via Hodgkin-Huxley cycle (other ion specific protein channels open)
At threshold (approx. -50 mV), all Na+ channels in the area open: => briefly, the membrane reverses polarity (more positive inside than outside)
This is an instance of “checks and balances”: threshold may be adjust → certain chemicals may change threshold, temperature can change threshold of neurons.
Closing Down
Na+ channels close & K+ channels open => K+ ions leave the cell: membrane repolarises. Sodium/Potassium pump pushes out positive sodium ions
K+ channels close at resting potential => briefly, more K+ ions outside than inside the cell => membrane hyperpolarised (inside even more negative than usual).
Even more difficult to trigger an AP, meaning there would need to be a stronger signal to trigger an AP
Part 4. Signal Transmission Between Neurons
1. Synapse - Structure & Processes
Neurons ‘communicate’ at synapses:
axon terminals of the pre-synaptic neuron (‘sender’) come close to dendritic spines of the post-synaptic neuron (‘receiver’)
Electrical synapse: almost no distance between axon terminal and dendritic spine
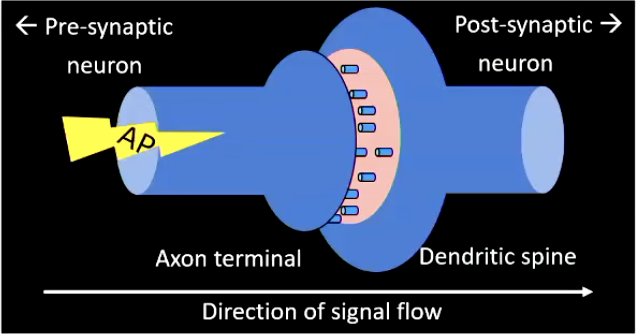
AP pass through and directly influences post-synaptic membrane potential
This is fast, but a disadvantage is it is inflexible. very little opportunity to implement checks and balances. This is good for fast reflexes.
As opposed to chemical synapse...
Chemical synapse (get screenshot from lecture capture):
axon terminal and dendritic spine separated by a small gap (synaptic cleft)
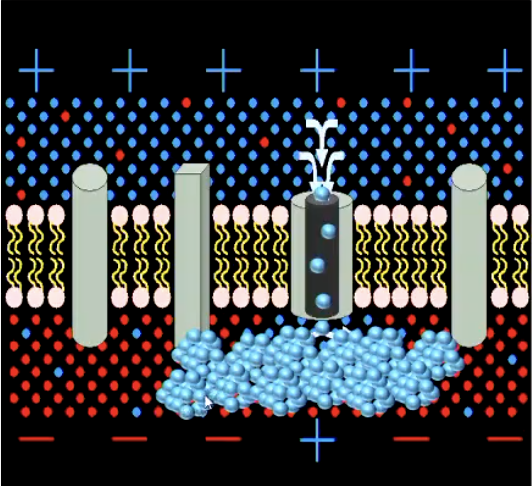
AP triggers opening of Ca++ channels -> Ca++ions enter and bind with vesicles -> cause neurotransmitter (NT) vesicles to fuse with cell membrane -> NTs released into synaptic cleft
Checks and Balances: Something only happens when there are enough Ca++ ions in the environment. Nothing occurs otherwise.
NT bind to receptor molecules of ion channels in the post-synaptic membrane -> channels open
Checks and Balances: It is not sufficient that the vesicles release NTs for the NTs to bind to receptor sites on the post-synaptic ion-channels, there has to be ‘enough’ NTs for the likelihood of binding to increase. Neurons regulate how many NTs are produced.
Ion-flow alters potential of post-synaptic membrane:
If positive ions enter, membrane depolarises (excitatory post-synaptic potential) - easier for AP to generate
If negative ions enter / positive ions leave, membrane hyperpolarises (inhibitory post-synaptic potential) - difficult for AP to generate
Astrocytes can affect neuron signal transmission. They can influence the balance of calcium in the environment surrounding the axon terminal and dendritic spine.
2. Ion Channels
Voltage-gated channels:
respond to changes in electrical charge (e.g., Na+ - and K+-channels in the axon hillock and the axon, Ca++ - channels in the axon terminal)
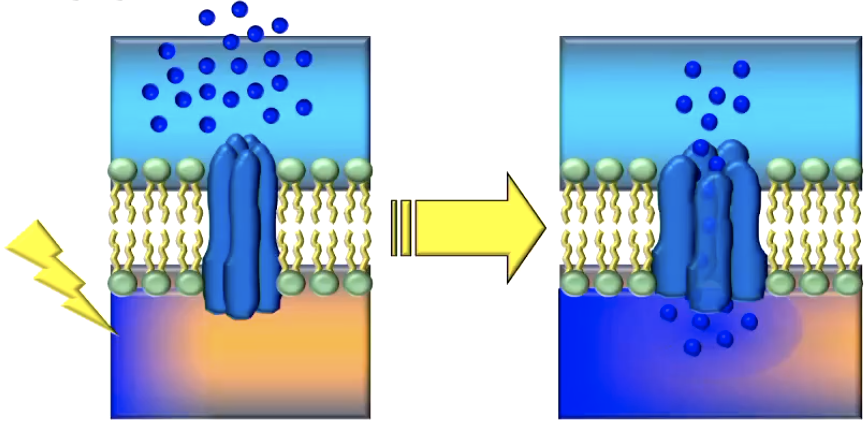
The electrical charge passing the voltage-gated ion channel forces the protein channel to ‘unscrew”, and open. Examples of voltage-gated channels include Na+ and K+ channels in the Axon Hillock and the axon. Na+ channels respon to little electrical charge but close when it becomes too positive. K+ channels don’t respond to little electrical charge, but open when the charge is very strong.
Ca++ are another example that respond when the AP arrives at the Axon terminal
Transmitter-gated channels:
are not (or not only) affected by voltage changes
all respond to chemicals (neurotransmitter) instead
Two types of transmitter-gated channel receptors:
Ionotropic receptors open their channel directly (Simple)
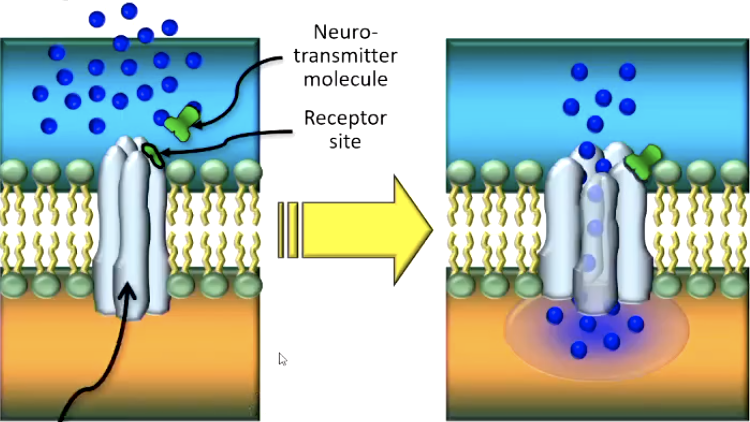
Metabotropic receptors open it indirectly (through g-proteins)
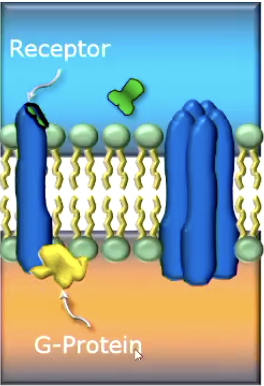
Receptor site is isolated from protein channel. NT binds with receptor site. Signals G-protein attached inside the receptor to send ‘secondary messengers’ to open the channel.
Checks and Balances: NT may bind correctly with the right receptor, but the chemicial environment within the cell may have too few Second Messengers released, meaning channel still won’t open.
Channel With Ionotropic Receptor: Channel With Metabotropic Receptor:
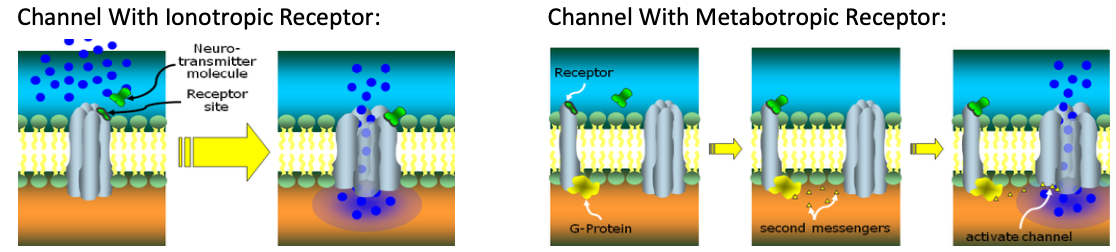
Neurtransmitter fits onto a ‘docking site’ on the protein of the gate, which forces a structural change to the channel, allowing it to open.
Checks and Balances: The correct NT is needed at the correct receptor site, wrong NT cannot bind there and channel won’t open.
3. NT Receptors & Key-Lock Principle
All ion channels in the post-synaptic membrane are transmitter-gated and of different types. Different channels respond to different NTs.
But different channels respond to different neurotransmitters
(because they have differently-shaped receptor sites)
Key-Lock Principle:
Type of NT determines which type of channel opens
(because each ‘gate’ has a ‘lock’ that requires a specific ‘key’)
But type of channel determines which type of ion can enter/leave the cell
And type of ion (pos/neg) determines how the cell behaves next => Therefore, NTs and receptors determine how the cell behaves next
Part 5. Information Processing
1. Excitation & Inhibition:
At an excitatory synapse,
positive ions enter (depolarisation; EPSP),
making it more likely that an AP will be triggered at the axon hillock
Key term - “more likely”. Every individual excitatory PSP is tiny, and one alone is never enough to push the generator potential past its threshold to generate an AP. If enough EPSP occurs, then the threshold can be crossed.
At an inhibitory synapse,
negative ions enter (hyperpolarisation; IPSP),
making it less likely that an AP will be triggered at the axon hillock
IPSP’s are also tiny, and only push the potential away from threshold a tiny bit and can be overridden by arriving at the same time as a large enough positive potential.
Whether a synapse is excitatory or inhibitory depends on the type of
neurotransmitter released by the axon terminal
ion channel & receptor present in the post-synaptic membrane
Remember: For an AP to be triggered, membrane potential at the axon hillock must depolarise beyond threshold (~ -50 mV)
2. Post-Synaptic Generator Potential (Post-synaptic summation)
Single AP causes tiny electrochemical changes in post-synaptic neuron
Insufficient to generate new AP!
But each neurons receive input from many other neurons in an ongoing pattern
All these small individual changes become integrated (‘blend together’) in post-synaptic neuron’s axon hillock
Slow build-up of generator potential
Across time: GP combines PSPs occurring in rapid succession (temporal summation);
Across space: GP combines PSPs from different synapses of one post-synaptic neuron (spatial summation)
Therefore, GP (unlike AP!) comes in different sizes
(‘graded’) - The bigger (more positive) the GP, the more likely it becomes that the post-synaptic neuron itself ‘fires’ an AP
Only if the input pattern is just right can the GP become large (positive) enough to trigger an AP.
“This is ‘Information Processing’ in the NS. These integration & transformation processes are the basis for ALL behaviour!!
3. Four (plus one) principles of signal transmission
1. Neural impulses serve different purposes:
Perception: Transmission of information from receptors into the brain (sensory neurons)
Control: Transmission of neural commands from the brain to muscles and glands (via motor neurons)
Coordination: Transmission of signals between neurons (interneurons)
2. Signal transmission is electro-chemical
Within a neuron, signals are transmitted electrically
Between neurons, signals are transmitted chemically
3. One type of neural impulse:
Action potentials cannot be modified (!)
All types of neurons transmit the same electrical signal (!)
4. Two types of information coding:
Quantitative aspect (‘how strong’): represented by a neuron’s firing rate: The stronger the input, the quicker AP’s follow each other
Qualitative interpretation (‘what’): determined by the location in the brain where the signal is received
Fifths principle of signal transmission...
Signal transmission is determined by the structures of the nervous system
Cognition is determined by the structures of the nervous system (as are perception, action, emotion…)
L2 - The Nervous System
Part 1 – Functional Architecture 1 – Types of Structures
Information Processing & Behaviour
What is a Nervous System good for? Or: Which problem is the NS trying to solve?
To interact flexibly with the environment:
To flexibly link unlimited input options with unlimited response options
Principles of signal transmission I - ( see Lecture 1. )
Recall: Qualitative interpretation (‘what’) of neural signals determined by the location in the brain where the signal is received.
Therefore: Structured information processing (perceiving, feeling, remembering, behaving...) is determined by a structured nervous system
Part 2 – Functional Architecture 2 – Types of Processes
Signal transmission
Synapses can be excitatory (making it more likely that a new action potential will be generated in the post-synaptic neuron)
OR
Inhibitory (making it less likely that a new action potential will be generated in the post-synaptic neuron)
Synaptically linked neurons from coherent (functional)
structures
Signal transmission can be
bottom-up (from ‘lower’ or more basic processing to ‘higher’ or more elaborate processing)
top-down (the other way round)
and also:
Feed-forward (to areas that have not previously been activated in this particular signal transmission) or
Feedback (to areas that have already participated in this particular signal transmission, to modify or adjust their processing)
Feedback Loops
Feed-forward & feed-back input both use excitatory and inhibitory connections, combining to form complex feedback loops:
Without feedback:
With feedback:
Positive: increases
probability of
subsequent signal output
Negative: maintains or decreases the probability of a subsequent signal output
Structured (i.e., functional) signal transmission through a combination of all these options, thus:
Structured transmission of signals needs a structured organisation of nerve cells!
Part 3 – Anatomical Structures 1 – Nervous System
Overall Structure – CNS & PNS
Overall:
80 – 100 billion neurons;
100 – 10,000 connections per neuron;
tendency to form densely packed assemblies
Two Types of Nerve Cell Assemblies:
Structure layers (CNS: cortex, pl: cortices; PNS: none)
Clusters (CNS: nucleus; pl. nuclei; PNS: ganglion, pl. ganglia)
Centralised Nervous System:
Central nervous system (CNS):
Brain & spinal cord – largest, most densely packed accumulation of neural assemblies;
Highly interconnected & protected by bones (skull & vertebrae)
Peripheral nervous system (PNS):
Other neurons & neural assemblies, scattered throughout the body & connected to the CNS
The ‘classic’ distinction: CNS / PNS clearly separated
Problem: CNS and PNS anatomically interlinked…
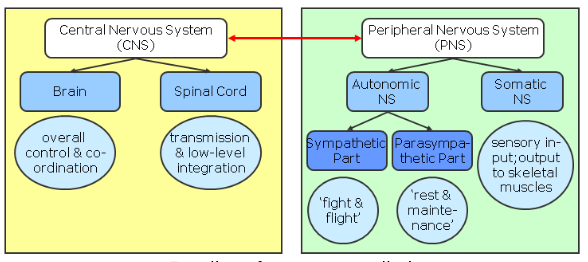
Division of the Peripheral Nervous System
PNS – somatic division:
Afferent (sending sensory signals into the CNS): cell bodies outside the CNS (“dorsal root ganglia”)
Efferent (sending motor signals out to
skeletal muscles): cell bodies within the
spinal cord (CNS)!
PNS – autonomic division:
Afferent: cell bodies are also in dorsal root ganglia
Efferent: (sending signals to cardiac muscles, smooth muscle, muscles & glands):two-neuron chain:
pre-ganglionic neurons: within the spinal cord (!)
postganglionic neurons: outside the spinal cord
CNS and PNS are functionally interlinked
No difference between CNS and PNS signalling
Constant signal exchange between both
How are CNS and PNS different?
CNS is protected by:
Bone
Meninges (‘membranes’)
Cerebrospinal fluid
Blood-brain barrier
Are CNS neurons more ‘fragile’?
PNS neurons can regenerate damaged axons
CNS neurons (generally) can not
From PNS into CNS: Spinal Cord
Reflexes: Some simple behaviours generated by the spinal cord alone:
Monosynaptic reflex:
sensory & motor neurons make direct contact (e.g., knee- jerk reflex)
Polysynaptic reflex:
sensory & motor neurons connect via one or more inter- neurons (e.g., withdrawal reflex)
Note: not all reflexes are mediated by the spinal cord!
Note: “Reflex” in this technical sense is a ‘hard-wired’ response, not any spontaneous behaviour! (not “I punched him in the face reflexively” – just: “I punched him in the face unthinkingly”)
Part 4 – Anatomical Structures 2 – CNS / Brain
Overall Structure of the Brain
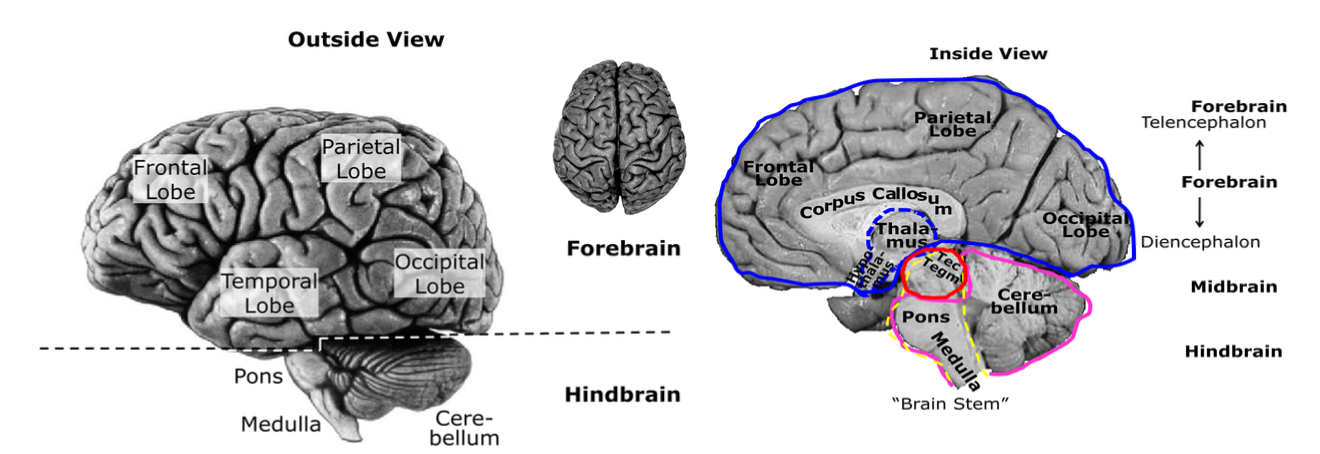
Diencephalon
Thalamus
A group of relatively big nuclei, on top of the midbrain, deep in the centre of the brain
Main relay station for most sensory signals (bottom-up)
Top-down input from cortex (modulating sensory signals)
Hypothalamus
Group of small nuclei
In front of & below the thalamus
Connected to and controlling
Pituitary gland
‘master gland’ of the ES
controls the activity of all other glands
Telencephalon
Three main structures: Basal ganglia, limbic system, cerebral cortex
(1) Basal Ganglia
A group of large nuclei around the thalamus
Important in motor control
Consist of:
Globus pallidus
Putamen
Caudate
Amygdala (‘almond’):
Anatomically: Part of the basal ganglia
Functionally: Part of the limbic system:
(2) Limbic System
Functional (not anatomical) structure: Closely connected cortical & sub-cortical areas crucial for memory and emotion
Cortical: Cingulate gyrus (evolutionary old cortical area!)
Subcortical:
Mammillary bodies, connected via the fornix to
Hippocampus, connected to
Amygdala
Also: olfactory bulb anatomically & functionally closely connected
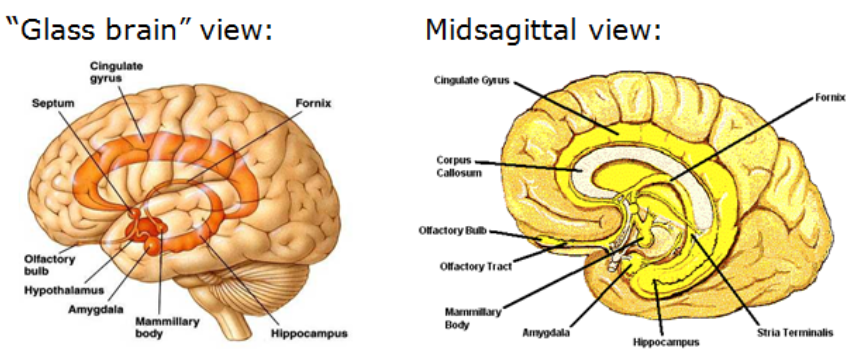
(3) Cerebral Cortex
Anatomical structure:
Highly folded, multi-layered sheet of neurons
completely covering each hemisphere
forming gyri (s. gyrus, outward folded areas) & sulci (sulcus, inward folded areas)
Large sulci used as ‘landmarks’
(e.g., longitudinal fissure separates left/right hemi-sphere
central fissure separates frontal/parietal lobes)
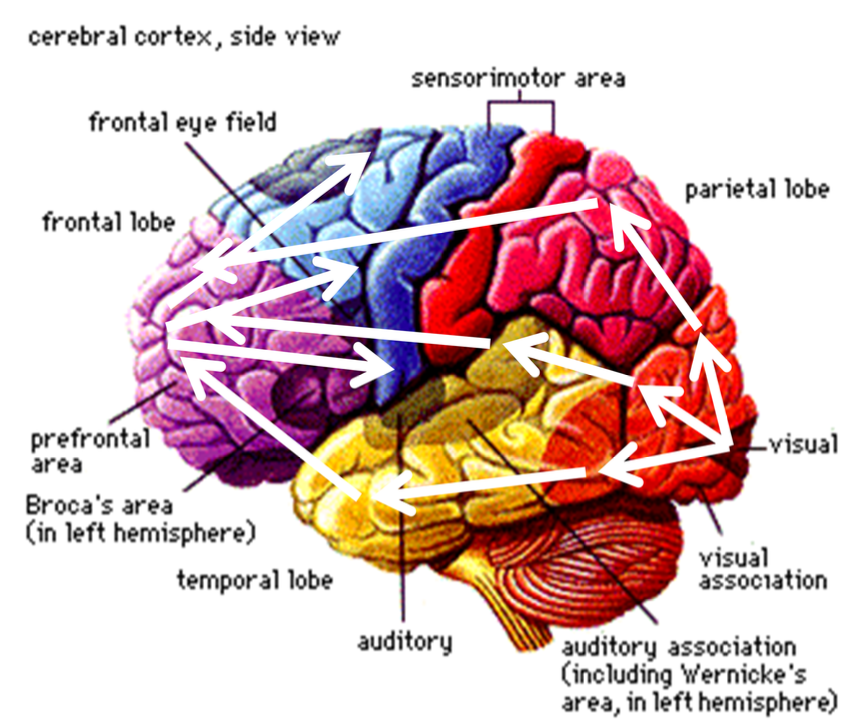
Functional structure:
Sensory input from thalamus send to primary sensory areas:
Vision: occipital cortex
Audition: temporal cortex
Somatosensory perception: parietal cortex
Signals transmitted on to secondary sensory areas
on to ‘higher’ association areas
on to frontal lobes (action planning)
motor cortex (movement execution)
back to subcortical structures -- up again to cortical areas -- etc etc etc
“Information Processing” in the NS consists of reverberating activity in a highly organized structure"
Summary of Structures & Functions
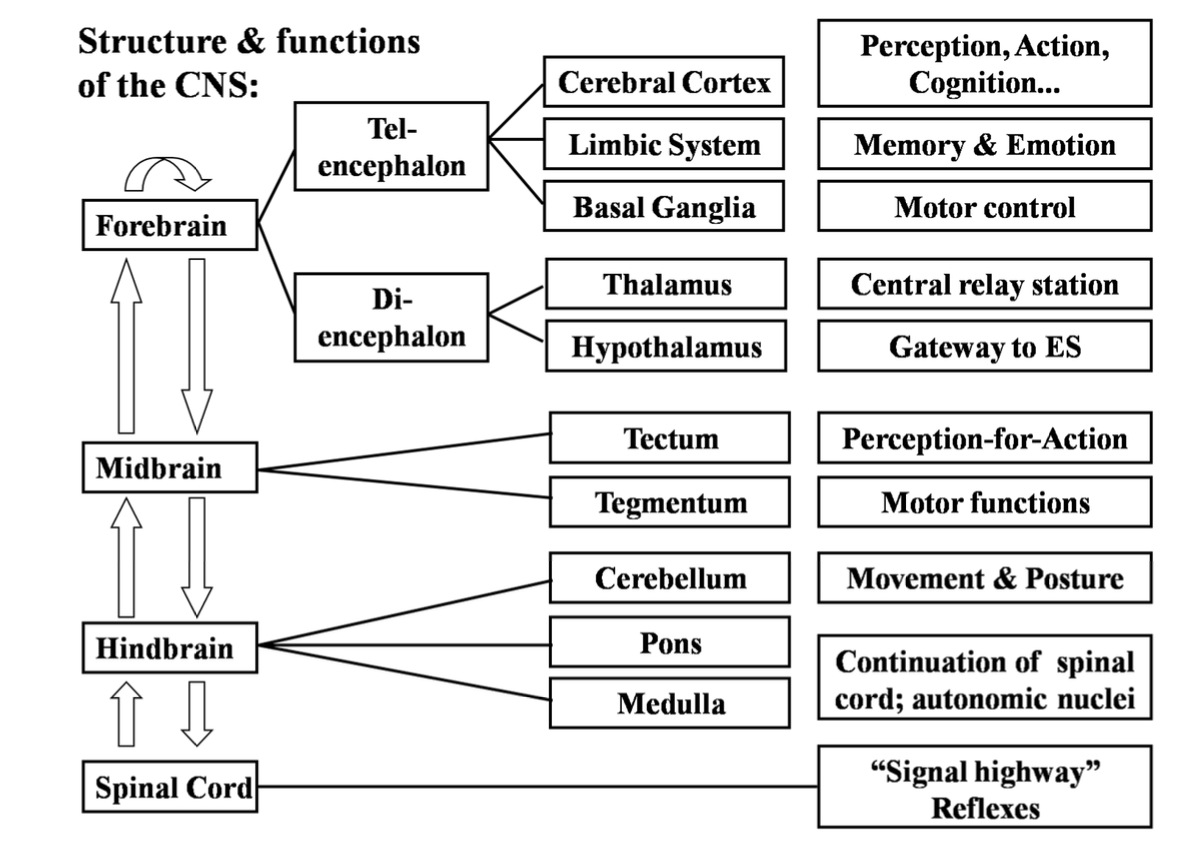
L3 - Genes and Hormones
Part 1 – Genes
Genes & Chromosomes
Genes:
Structural: Sections of DNA
Evolutionary: Substrate of heredity
Functionally: Blueprints for protein synthesis
Passive stage: Chromatin strands are densely packed
Active stage: DNA in loose chromatin strands
Recall: Cells are chemical factories
Consists of a thick chemical soup inside a cell membrane
Exist in a chemical soup outside the membrane (environment)
Live by exchanging chemicals through the cell membrane
All these processes involve proteins
Genes & Proteins
Proteins:
Complex molecules (very long, highly folded amino-acid chains)
Function:
Building blocks of cell structures (e.g., protein channels)
Active ingredient that ‘works’ on inert biological molecules (e.g., enzymes)
Produced in response to (internal & external) chemical environment
Function: to deal with the chemical environment
Genes ‘code’ proteins
Contain information from which proteins are built
Each gene codes for a specific protein
‘Activating’ a gene = causing production of this protein
Interlude: Gene-Environment Interaction
A very simple set of rules can produce a large variety of outcomes:
depending only on
the exact environment, and
the exact starting point!
Environment can change the rules! (can change which rules are active)
A cell’s genetic code is a ‘rule book’ rather than a ‘blueprint’:
Does not contain instructions on how to build/operate a cell,
but instructions on how to respond to the environment
Some responses alter a cell’s form or function,
Others alter which genes (rules) are active,
thereby changing a cell’s responses to the environment.
Genes & Proteins & Steroid Hormones
Produced with the help of steroidogenic enzyme
Characteristics: Fat-like substances (i.e., can easily pass through cell membrane)
Sources:
Gonads (ovaries & testes)
Adrenal glands
Types:
(I) Sex hormones:
Estrogens & progesterone: Promote development of female sex characteristics
Androgens: Promote development of male sex characteristics
(II) Corticosteroids:
Control stress response & sugar metabolism
Function:
Modify gene expression (i.e., activate genes)
Bind to specific receptor molecules inside the cell
Receptor molecules bind to specific gene & activates it
Result: Proteins will be produced
Part 2. Sex Determination in Mammals
Human body cells: contain 46 chromosomes
forming 23 matched pairs:
22 pairs of autosomes
1 pair of sex chromosomes (XX or XY)
Human gametes (egg & sperm cells) contain
only 23 chromosomes
22 single autosomes
1 single sex chromosome (X or Y)
Fertilisation:
Combination of maternal &
paternal chromosome half-sets
Chromosomal Sex:
In mammals, sex is determined by a combination of sex chromosomes
XX: chromosomal female
XY: chromosomal male
Male development depends on the presence of the Y chromosome:
An embryo with Y chromosome (XY) will typically develop as a male
An embryo without Y chromosome (XX) will typically develop as a female
What’s special about the Y chromosome?
Sometimes, chromosomal sex and phenotypical sex don’t match:
XX males: when a particular section of the Y chromosome is translocated to another chromosome
XY females: when that particular section of the Y chromosome is missing
Conclusion: this region on the Y chromosome determines male development (SRY – sex- determining region of the Y chromosome)
SRY carries the TDF gene
Codes TDF (testis-determining factor) protein
Which turns developing gonads into testes
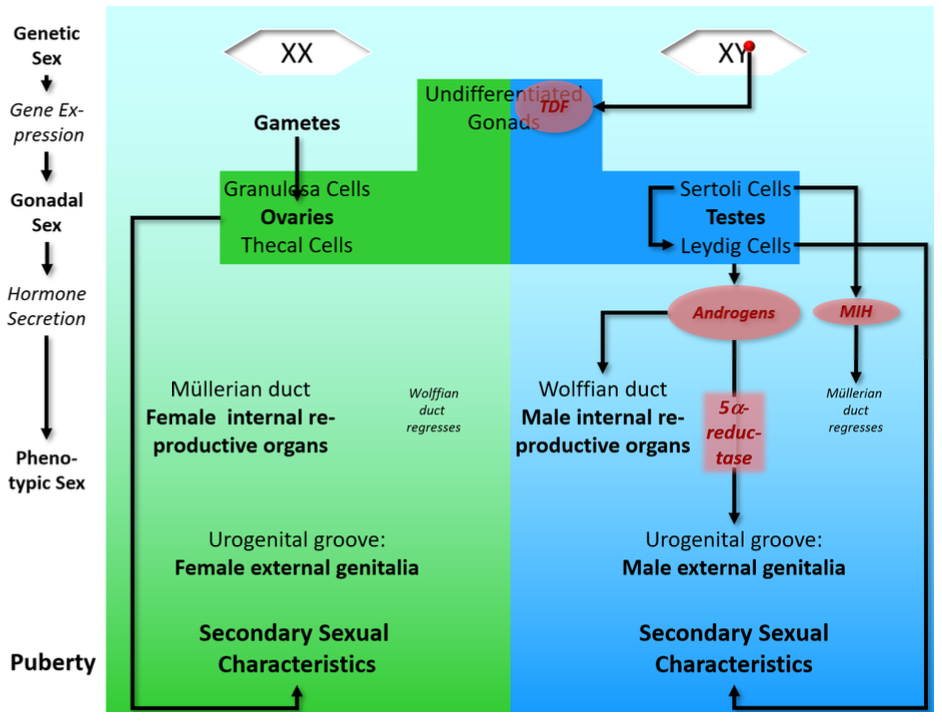
Part 3. Sexual Development
Embryonic Development
Early stages of sexual development:
Gonads develop in the first month;
Initially undifferentiated (can become either testes or ovaries)
Chemicals from undifferentiated gonads flood surrounding tissue
Can ‘switch on’ TDF gene
Initial MALE development (in normal XY male)
TDF gene present in all cells (on Y chromosome)
Gonadal signals trigger TDF protein production
immature gonads respond to TDF -> Turn into testes
Sertoli cells:
Secrete MIH (‘Müllerian inhibiting hormone’) -> Müllerian duct degenerates
Trigger development of Leydig cells
Leydig cells:
secrete androgens (testosterone)
FEMALE development starts by default:
Gonads develop into ovaries:
Granulosa (supporting) cells
Thecal cells
Begin to secrete sex hormones only in puberty (if gametes present)
N.b.: gamete-producing cells originate somewhere else (in the yolk sac) and migrate to the gonads (through the gut)
Subsequent MALE development follows hormonal ‘instructions’:
Androgens masculinise all body tissues (except brain)
Müllerian duct: already gone
Wolffian duct -> male internal sexual organs
Urogenital groove -> male external genitals
needs 5α-reductase (an enzyme in urogenital tissue, converts testosterone into DHT)!
Androgens make a body ‘male shaped’
FEMALE development follows default path:
All body tissues develop according to their intrinsic program
(intrinsic = the original, not chemically altered path)
Müllerian duct -> female internal sexual organs
Wolffian duct: degenerates
Urogenital groove -> female external genitals
A female body happens in the absence of chemical “shaping”
Summary flow chart:
Developmental Disorders
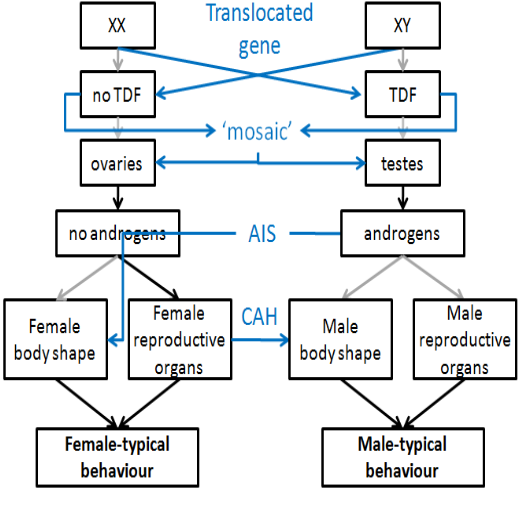
2.1. Chromosomal
Only one sex chromosome:
No X: not able to develop
X0 females: Turner Syndrome
No gametes => No functional ovaries => No (or little) hormone production during puberty => No sexual maturation
More than two sex chromosomes
Multiple Y
No X (YY0, YYY0…): not able to develop (see above)
One X (XYY, XYYY…): Male phenotype
Multiple X
No Y (XXX, XXXX…): Female phenotype
One Y (XXY, XXXY…): Male phenotype (Klinefelter Syndrome)
Generally: the more extra chromosomes, the higher the likelihood of physical & cognitive defects
2.2. Hormonal
Insufficient MIH (Persistent Müllerian Duct Syndrome):
XY males with dysfunctional Sertoli cells:
No MIH: Development of Müllerian duct into female internal reproductive organs not suppressed
‘Intersex’ appearance of internal reproductive organs – both male and female internal reproductive organs (neither able to develop fully)
External phenotype: not affected
Androgen overproductions (Congenital Adrenal Hyperplasia; CAH)
Fetus with overactive adrenal gland
In XX females: Possible ‘intersex’ appearance of external genitals (intermediate between male and female)
But: Internal reproductive organs usually not affected (adrenal gland develops too late – produces androgens after regression of Wolffian duct)
External phenotype: ranging from unaffected (female) to severely affected (pseudo-male)
Note: CAH affects males as well!
2.3. Receptors & Enzymes
Androgen Insensitivity Syndrome:
XY males with non-functioning androgen receptors
Body develops according to intrinsic (female) program;
But normal hormone (MIH & androgen) production:
Development of internal female reproductive system suppressed
External phenotype (depending on severity): Female
5alpha-reductase Deficiency Syndrome:
XY males without 5α-reductase
Testosterone not converted to DHT:
Female (or intersex) development of external genitals
But: Increased testosterone production at puberty may cause development of male external genitals
External phenotype: Female (at birth) to male (at puberty)
Take-home message: even at this most basic level, there many “shades of grey” between “fully female” and “fully male” – it can only get worse when we move on to the (far more complex!) brain…
Part 4. Brain Development
Hormonal influence (in rodents):
Brain requires hormones to develop in male-typical form
Relevant hormones are estrogens (the female sex hormone)
Evidence:
Experimental (behavioural): Treating new-born female rats with estrogens will cause them to show male-typical sexual behaviour as adults
Neuroanatomical: SDN-POA (sexually dimorphic nucleus of the preoptic area of the hypothalamus):
Problem:
All (rat) embryos are exposed to maternal estrogens
Then why do not all rats show male- typical sexual behaviour?
Why do brains develop in different (sexually dimorphic) forms?
A possible solution? Aromatisation hypothesis:
The mothers’ estrogen is inactivated (by α-fetoprotein);
But androgens – produced by the newly developed testes of male rat embryos – can enter the brain
Inside the brain, aromatase converts androgens into estrogens
Human brain development:
Male and female brains assumed to develop differently (do they really?)
No influence of maternal estrogens (cannot cross placenta) →Aromatisation?
Evidence supporting aromatisation hypothesis:
Synthetic estrogens can cross the placenta
Daughters might show more male-specific behaviour?
Evidence against aromatisation hypothesis: Androgen-insensitive XY-individuals
normal levels of androgens
normal levels or aromatase
normal estrogen receptors in the brain
Should develop ‘male-typical’ brains – but show ‘female-typical’ behavioural patterns instead…
Current assumption: androgen receptors play a role in human male brain development
Problem: not at all clear what a “male human brain” even is…
L4 - Hormones and Learning
Part 1. Sex & Gender
Some Definitions:
Sex:
Category of ‘female’ and ‘male’
Specifically with respect to molecular (e.g., genotype) and anatomical (e.g. reproductive organs) features
Recall L3: even at this level, a clear categorical distinction is not possible!
Sexual behaviours
Specifically, behaviours associated with sexual reproduction (to ‘have sex’; to ‘engage in sexual intercourse’). Key word - ‘sexual’ reproduction. It is possible for other species to reproduce asexually.
Gender:
Collection of psychological traits that differ between the sexes (cognitive, behavioural, personality... do they really differ??)
Definition of ‘gender’ based on our understanding of ‘sexes’!
and on our understanding of gender...
But how to define someone’s sex?? (shown below). However, there is still no guarantee, as there are possibilities that may mean while an individual may have androgens (for example), their receptors may not be receptive/responsive to this, they may still end up with a female body shape.
Sex & Sexual Behaviour
Problem: What is sex-typical behaviour??
A simplistic answer:
Things that women do, and men don’t do
Things that men do, and women don’t do
This is too simplistic as pregnancy and childbirth are the only 2 behaviours that women do, and men don’t do.
A simple answer:
Sex-typical behaviours & postures during sexual reproduction (e.g., ‘lordosis’ & ‘mounting’ posture of copulating rodents)
Rats can only copulate in this position
Reproductive success depends co-ordinated sex-typical behaviour
Advantage of genetic / hormonal control of such behaviours
Organisational vs activational effects of sex hormones
Organisational effect of sex hormones:
Normal XX mammal will have:
Low levels of androgens
♀ (female) typical body shape
♀ (female) typical brain structures
Normal XY mammal will have:
High levels of androgens
♂Typical body shape
♂Typical brain structures
Activational effect of sex hormones:
(normal xx mamal) More ♀ sex hormones (only after onset of puberty)
[Normal XX mammal](Species-specific) ♀-typical sexual behaviour repertoire
(Normal XY mammal) More ♂ sex hormones (especially after onsent of puberty)
[Normal XY mammal] More ♂-typical sexual behavioural repertoire
Reproduce
Part 2. Environmental Effects on Sexual Behaviour
Learning
Even instinctive behaviour already influenced by learning
Experimental evidence: Mixed-sex groups (when a male is place in an isolated environment with a receptive female, the two will reproduce) vs. iso-sex groups (when a receptive female is place with a male from thus group, the males response is different. The male will be much less responsive to the femlaes advances, and teh chances of reproduction are reduced) vs. isolation rearing of rats (when a male is place with a receptive female, the response is different. The male is not receptive to the females advances at all. Low chances of reproduction):
No adults present during rearing: no imitation learning!
Pre-puberty sexual learning depends on social interaction with members of the opposite sex!
HOMEWORK: look up “WESTERMARCK EFFECT”
Effects Without Learning
Environmental influence on behaviour begins even earlier:
Post-natal: Rat mothers show ‘Gender/sex-specific maternal behaviour’
Rat mothers lick the genital region of their newborn pups (to keep them clean & stimulate urination)
Male pups get licked more than female pups (triggered by their specific smell)
Male pups that are only licked as much as females show reduced male-typical sexual behaviour as adults
Pre-natal: Maternal stress effect
‘Stressing’ pregnant rats during a certain ‘critical period’ causes their male offspring to show reduced male-typical behaviour as adults
Pre-natal: Uterine contiguity effect
Female rats that have developed in the uterus next to male siblings show increased male-typical behaviour
How can early environmental influence adult behaviour?
Post-natal: Gender/sex-specific maternal behaviour
Gender-specific licking of genital region increases androgen levels in newborn male rats
Androgen levels increase → promotes higher male-typical brain development (recall: aromatisation & SDN-POA) => larger SDN-POA
Less licking, less androgen → smaller SDN-POA)
Male pups that are only licked as much as females show reduced male-typical sexual behaviour as adults
Pre-natal: Maternal stress effect
Maternal stress (during a critical period0 disrupts timing of peak androgen levels in male embryos:
Levels peak before critical brain development => smaller SDN-POA
Pre-natal: Uterine contiguity effect
Bloodstream carries androgens from male siblings to ‘neighbouring’ female embryos
Affecting their brain development => larger SDN-POA (females usually have smaller SDN-POA, than males, so androgens increase this size)
“Environment affects hormone levels Hormone levels affect brain structures brain structures affect behaviour”
Role of SDN-POA: inhibit female-typical sexual behaviour (in men)? (this is pure speculation!)
Female sexual behaviour considered “default”
Small SDN-POA: default behaviour not inhibited
Large SDN-POA: can inhibit default behaviour and/or trigger alternative behaviour
What about the effect of mixed-sex rearing?
Social learning has probably no direct effect on sex hormone levels
More likely: animals practice and re-enforce instinctive behaviours during play
In other words:
Genes & environment determine pre- and post-natal sex hormone levels (molecular effect)
Sex hormone levels modulate sex-specific brain development (anatomical effects)
Having particular sex-specific brain structures makes animals more likely to exhibit sex-specific behaviour (behavioural effect)
But this still does not invariably trigger (force) that behaviour → Sex-specific behaviours must be practiced during adolescences to be performed properly in adulthood
Part 3. Biological Basis of Learning
(How can practice affect instinctive behaviour?)
1a. Structures - General
How can practice affect instinctive behaviours?
Brain structures not strictly ‘hard-wired’
Experience changes brain structures at the cellular level
Neuro-plasticity - the brain can be formed and shaped in different ways depending on learning through the environment
Some definitions:
Instinct = evolved, species-specific adaptation of a species’ behaviour towards the environment
Learning = rapid, intra-individual adaptation of one’s behaviour towards the environment
Memory = Lasting effects of learning
Role of the cortex in memory (an old-fashioned view):
The human and rat brains are somewhat similar
Karl Lashley’s (1940s) experiments on rats:
Trained rats to navigate a maze
Remove part of the cortex
Test memory for maze
Results:
The more cortex removed, the worse the performance
Irrespective of which part of cortex had been removed
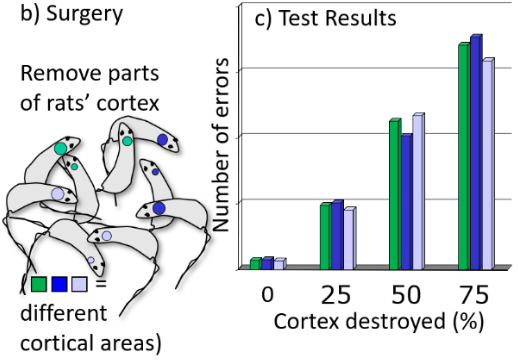
Lashley’s Laws:
Mass Action: Learning & memory are a function of intact cortex mass – more tissue, more learning & memory
Equipotentiality: Each part of the cortex is equally involved in learning & memory
Why Lashley was (somewhat) wrong:
There are brain structures that are specifically involved in learning and memory, but they are (mostly) sub-cortical
Information mostly from lesion studies
Deliberate lesions in animal studies
Patients with brain lesions (not deliberately induced!)
Information therefore incomplete, but suggests specific brain structures involved in learning
Also, information gained from animal studies can also only be applied to humans to a limited extent. In this case, anima;s can only learn from trial and error. They cannot be instructed in how to do something. Where as humans can learn through instruction from another human and thus possibly get the task right on the first time.
1b. Structures - Specific
Specifically relevant structures:
Factual & relational learning: Limbic structures:
Hippocampus
Amygdala
Mammillary bodies
Motor skill learning:
Cerebellum
Basal Ganglia
Hippocampus:
Receives Input (via entorhinal cortex) from :
Subcortical areas:
Amygdala
Septal nuclei
Cortical areas
Limbic cortex (aka cingulate cortex)
All association areas (high-level cognitive processing: inter sensory integration, thought, reasoning, planning etc.)
Hippocampus sends it output to:
Subcortical:
Amygdala
Septal nuclei
Thalamus (via mammillary bodies)
Cortical areas:
Limbic system
All association areas
Lesions of the hippocampus result in
Impaired spatial / navigation skills (animal studies)
Anterograde amnesia (recall patient H.M.)
Amygdala
Directly connected to lateral hypothalamus ->ES
Involved in learning biologically significant information
Electrical stimulation: fearful & aggressive behaviour
Lesion: failure to learn ‘conditioned fear response’ (animals can’t learn that a stimulus signals danger)
Septal nuclei
Involved in processing of (or generating) reward:
Electrical stimulation: pleasurable – animals continually self-stimulate
Lesion: over-activity, failure to calm down
Processes
Possible mechanisms for Hebbian leaning:
Mediated by NMDA receptors
Specific neurotransmitter receptor protein
To open ion channel, both pre-synaptic and post- synaptic cell must be simultaneously active
Pre-synaptic cell: release of glutamate
Post-synaptic cell: depolarised
Recall PS111:
Cells that fire together, wire together
Out of sync, lose their link
Learning changes neural signalling:
Long-Term Potentiation (LTP):
Post-synaptic cell responds more strongly to input (> PSPs)
after a period of persistent strong (i.e., high-frequency) input
Long-Term Depression (LTD):
Post-synaptic cell responds less strongly to input (< PSPs)
after a period of persistent weak (i.e., low-frequency) input
Learning changes structure of the brain:
Short-term molecular changes after increased co- ordinated activity:
Pre-synaptic: more neurotransmitter
Post-synaptic: more ion channels
Short-term structural changes: larger pre- & post-synaptic areas
Long-term structural changes result from short-term molecular changes:
Formation of new synapses
Shift in synaptic input
Why are Septal nuclei, amygdala & hippocampus so important for learning?
Particularly rich in NMDA receptors - therefore particularly suited to support ‘Hebbian Learning’
Septal nuclei: fast association of stimuli or events with positive feelings (learning what’s good)
Amygdala: fast association of stimuli or events with negative feelings (learning what hurts)
Hippocampus: fast association of stimuli or events with each other (factual and relational learning)
Hippocampus: A ‘gateway’ for establishing new connections between cell assemblies?
Animal evidence: Gene-manipulated mice with increased number of NMDA receptors show increased spatial abilities
A memory pill?
Would a treatment that increases the number of NMDA receptors in the hippocampus improve learning in humans?
Yes, but:
Hippocampus is an extremely sensitive and ‘unstable’ structure – susceptible to seizure activity and anoxia (lack of oxygen)
Probable cause is great number of NMDA receptors and corresponding high metabolic demands
Part 4. Human Sexual Behaviour Determined by Learning?
Interim Summary (some fundamental postulates)
All behaviour ultimately results from electro-chemical neural processes in the brain
These processes are determined by
Short-term molecular changes due to increased (co-ordinated) activity
Long-term structural changes following molecular changes
Structures are determined
genetically at the macroscopic level: adaptation through evolution
by experience at the microscopic (cellular) level: adaptation through learning
Humans are exceptionally good learners!
Some Basic Problems
Problem 1:
What exactly is sexual behaviour in humans?
only behaviour that directly results in reproduction?
all behaviours that indirectly increase the likelihood of repro-duction?
all behaviours that are perceived as sexual, whether or not they affect reproduction?
How (sex- or gender-)specific is sexual behaviour in humans?
Problem 2:
Memory & learning research investigates how clearly defined information or skills are learned
Sexual learning, in contrast, involves learning a broad, ill-defined (or not defined) set of information and behaviours
Problem 3:
No large-scale, systematic studies in humans exist – or are even possible!
Some evidence against learning in human sexual behaviour (and some limitations thereof):
5α-reductase syndrome
‘girls turning into boys’ at puberty
apparently no problems adapting ‘male-typical sexual behaviour’
(but perhaps parents and peers already expected the change?)
CAH syndrome
girls with pathologically increased androgen levels
might display strong ‘male-typical’ playing behaviour (despite being discouraged to do so)
(but might not be more likely to become lesbians?)
Children raised by homosexual couples
are not more likely to become homosexual than children raised by heterosexual couples
(but perhaps in humans, too, sexual learning depends on peer interactions rather than explicit teaching / role models?)
Take-home message: Lots of data, but nothing conclusive
(i.e., nobody knows)
L5 - Hormonal Control and Sexual Behaviour
Part 1. General Background: Learning & Physiological Arousal
Memory & Arousal Experiment (‘Picture & Story Paradigm’)
Method:
Participants see pictures while hearing either
an emotionally neutral story (group N), or
an emotionally arousing story (group A)
In each group, participants are injected with either
a chemically inactive substance (placebo; ‘Pl’), or
an adrenalin antagonist (‘Ant’; prevents effects of adrenalin)
Memory for pictures is measured some time later
Interpretation:
Effect of emotional arousal on learning mediated by adrenaline
Those in the N group who had the placebo or not had baseline memory. Those who were in the A group and had the placebo had increased memory in the critical test condition. Those in the A group that were injected with adrenaline did not have the same memory effect and returned to baseline.
Extension:
Similar experiments, testing other stress hormones:
Adrenalin
Noradrenalin
Corticosteroids
All show similar results
Conclusion:
Stress hormones facilitate learning (up to a certain extent)
Part 2. Hormones & Social Learning
Stress Hormones
Sources of stress hormones:
Adrenal cortex:
Controlled by hormones from pituitary gland (hypothalamus!)
Secretes corticosteroids
Adrenal medulla:
Controlled by neural signals from autonomic NS (hypothalamus!)
Secretes adrenalin
Brainstem
Locus coeruleus & LTA
Secrete noradrenalin
Stress hormones affect:
Diencephalon
Thalamus (sensory input)
Hypothalamus (ES)
Stress hormones act as neuro-modulators🚩
Motor Systems
Striatum of BG
Cerebellum
Limbic System (memory & emotion)
Amygdala
Hippocampus
Cingulate Cortex
Neocortex (thought)
Stress hormones are involved in sympathetic arousal:
Increased alertness
“Fight-or-flight response” (actually: “fight-flight-or-mate response”)
Neocortex Interprets Arousal (‘Scary Bridge’ Study; Dutton and Aron, 1974)
Scary Bridge Experiment Method:
Participants: Men who have just crossed either:
shaky suspension bridge (experimental group) or
stable stone bridge (control group)
Female experimenter
asks men to answer a questionnaire, and
to call her later if they have questions about the study
Result:
More phone calls from men who just crossed the shaky bridge
Interpretation:
Crossing the shaky bridge is scary – body releases stress hormones. However, these ‘stress hormones’ do not come with a specific meaning.
Given the chance, participants interpret (threat-induced) adrenalin rush as sexual arousal:
Conclude that they found the woman attractive
→Physiological arousal can have different cognitive-emotional ‘meanings’!
Corollary:
Both negative and positive emotions might facilitate learning!
Hormones & Social Learning: A (Functional) Model
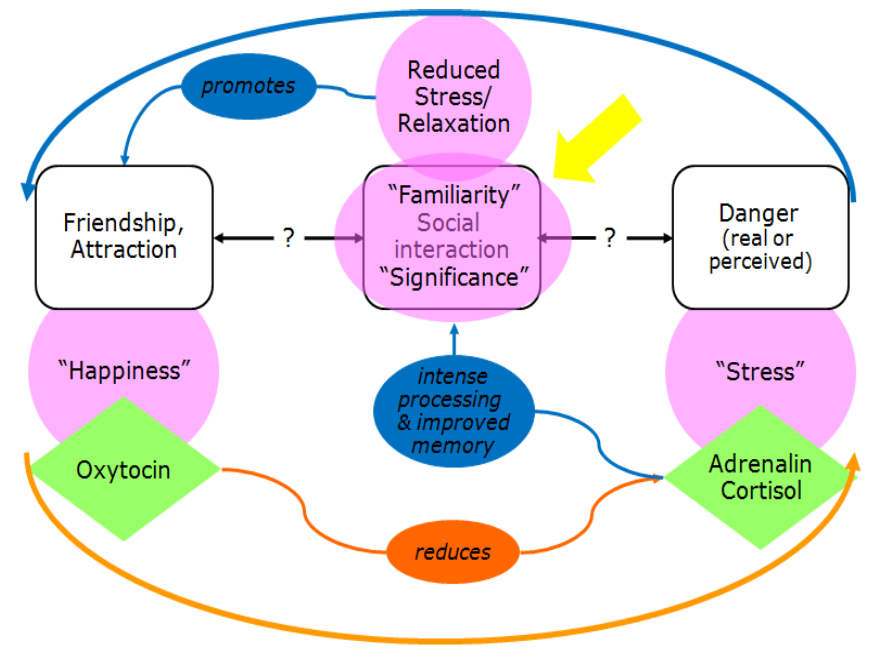
(n.b. consider what would happen with a social interaction that FAILS to reduce stress)
INTERLUDE: The Hypothalamus
Collection of various nuclei
Diverse input:
Neural
Hormonal
Direct (e.g., temperature, blood sugar level)
Part of / connected with
CNS
Autonomic PNS -> adrenal medulla
ES (via pituitary gland) -> adrenal cortex (‘HPA axis’)
Output:
Neural
Hormonal
Controls:
Growth & Homeostasis (-> Builds & maintains the body)
Stress response (=> Keeps it alive)
Reproductive behaviour (=> Ensures survival of the species)
Hormones & Social Learning: A (Functional) Model – Now With Brain
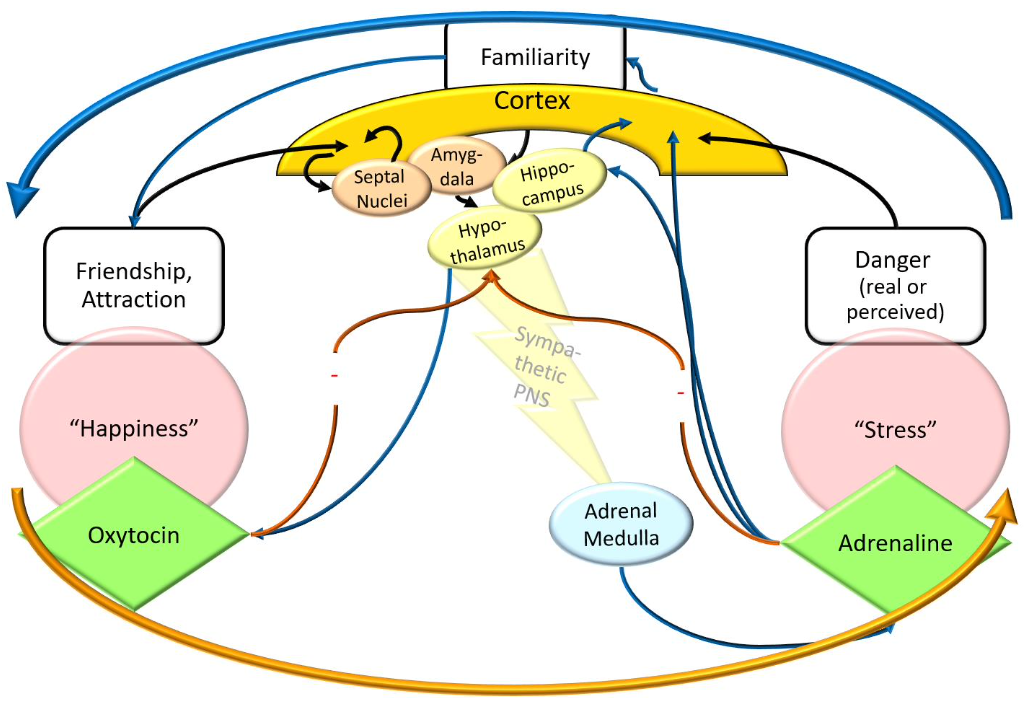
Hormones & Social Learning: Summary (Hormonal influence on social bonding)
Adrenalin promotes social bonding
Improves learning of / memory for salient environmental stimuli
e.g., people
Increases sense of familiarity / closeness with these people
Also, might indirectly increase oxytocin levels
Oxytocin: hormone & neuro-modulator
Produced by hypothalamus & pituitary gland
Role in social bonding:
Associated with feelings of well-being
Higher levels in socially or romantically attached people*
Levels increase dramatically during puberty
Released particularly during orgasm, breastfeeding, and other reproduction-related activities
* Gordon, I., Zagoory-Sharon, O., Schneiderman, I., Leckman. J. F., Weller, A., & Feldman, R. (2008). Oxytocin and cortisol in romantically unattached young adults: Associations with bonding and psychological distress. Psychophysiology, 45, 349-352.
Part 3. Hormones & Sexual Behaviours
1. Physiological Arousal & Puberty
Hormonal influences become fully effective only after puberty
Hypothesis: insufficient levels of oxytocin before?
Childhood
Sex hormone levels remain at +/- constant low level
No fertility
No secondary sexual characteristics
No display of complete sexual behaviour sequence
Sex-differences in non-sexual behaviour (?)
Puberty:
Increase in hormone levels
Initiated & controlled by hypothalamus
Resulting in
Maturation of reproductive system, gamete production
Development of secondary sexual characteristics
Ability to display complete sexual behaviour sequence
Not a single event – occurs over a period of time
Trigger: as yet unknown
(one possible factor – at least for girls – is body mass)
Sexual & Sex-Specific Behaviour
In most mammals, mature reproductive behaviour is:
Sexually dimorphic (i.e., sex specific)
Instinctive
Given a specific state, a specific behaviour will almost certainly occur
Controlled by hormones:
Organisational level (influencing the development of structures)
Activational level (influencing the display of specific behaviours; nb: “complete sexual behaviour sequence” even in rats is pretty complex!)
Reflecting hormonal differences between males and females:
Hormones & Behaviour: Activational Level
Sexual behaviour of females: Controlled by hormonal cycle:
Most female mammals sexually active only during fertile phase
Sexual behaviour of males: Depends on presence of androgens, not on their exact level
Castration at birth (no androgen): No sexual behaviour
Castration in adulthood: Decline of sexual behaviour
Testosterone therapy: Restores sex drive to pre-surgery levels
Part 4. Hormones & Socio-Sexual Behaviours
1. Flexibility of Sexual Behaviours – In Animals
Sexual behaviour is instinctive:
Under ‘normal’ conditions, specific male-typical or female typical patterns will develop
BUT: Surprisingly large variations in sexual behaviour:
In social animals, social status can determine sexual behaviour
Some possible ‘models’:
Mainly females with offspring, few adult males
young males leave (e.g., lions, elephants)
young females leave (e.g., bonobos, gorillas)
Similar numbers of males & females
both young males & females leave (e.g., humans)
Alpha pair
one pair reproduces, all others help raising the offspring (e.g., marmoset)
• Social suppression of reproductive behaviour:
In humans?
PROBLEM: Dissociation of sexual behaviour and reproduction
In animals: generalisation, but little dissociation
(exception: bonobos!)
In humans: almost total dissociation
Little sex-specific behaviour
(Almost) no sex-exclusive behaviour
Sex-typical behaviour: Unclear status
High variability
Social / cultural variations
Inter-individual variations
Intra-individual (e.g., contextual) variations
Some relationship between hormone level and sexual behaviour:
No sexual activity before puberty (similar to other animals)
Hormones and sexual behaviour in humans
General pattern of hormone levels similar to other animals: Testosterone levels in men: Estrogen levels in women:
Different from other animals:
Sexual behaviour of female humans (and bonobos, possibly all primates?) ‘emancipated’ from menstrual cycle
but perhaps higher sex drive around ovulation?
Female humans (possible all primates?) are sexually responsive to androgens
To conclude:
Hormone levels influence sexual behaviour (Del Giudice, M. (2009). Sex, attachment, and the development of reproductive strategies. Behavioral and Brain Sciences, 32, 1-21.)
Hormone levels are influenced by environmental factors
specifically: by social factors!
L6 - Brain Structure: “Female” and “Male” brain.
Part 1. Female and Male Behaviour
Brain & Behaviour
Some basic assumptions:
All complex behaviour depends on processes in the brain - Anything mnore complex than a simple reflex arc requires a ‘brain’.
Which processes a brain can perform depends on its structure (the braijn structures mediate the processes that occur within them):
different Systematically different structures give rise to
systematically different processes, resulting in
systematic behaviours - if there are systematic differences in the structures, this causes systematic differences in the processes that occur within them, then causing systematic differences in the behaviours exhibited.
Some further conclusions:
If two groups differ systematically in their behaviour, then this suggests corresponding differences in their brain structures
Systematic differences in male & female (sexual) behaviour suggest systematic differences between males’ & females’ brains (specifically: in those parts controlling sexual behaviour)
(BUT REMEMBER: ENVIRONMENT SHAPES BRAINS!) - example in rats, an isolated rat and a rat that had experience living with other rats within the same environment as them. The rat in the isolated environment had a cortex half the size of that of the rat that had experienced living with other rats.
(systematic differences refer to differences in behaviour that are observable across cultures and across time. E.g., applying this to humans, female behaviour in the UK now would not be the same as female behaviour 2000 years ago in Greece.
Systematic differences in male and female sexual behaviour??
Common Assumptions (examples only):
“Men are more attracted by physical signs of fertility than woman“
“Men have their first sexual experience at a younger age than women”
“Men have more sex than women”
Are any of these valid?
All of these studies are self-report-based
Social pressures can lead to biased values in the studies looking into the sex drives of both men and women.
The vast majority are Anglo-American
Recall: large inter-cultural differences in sexual behaviour!
Study on Swedish college students: results opposite to several of the above assumptions (Weinberg, Lottes & Shaver (1995), Archives of Sexual Behaviour, 24, 409-437)
This means that the common assumtions do not really line up with the truth about the sexual behaviours of humans.
Look at animal evidence first…
Part 2. Structural Differences – Animal Evidence
Mating behaviour: SDN-POA in rats (e.g., Paredes, 2003)
Behavioural: Sex-specific movements and postures during mating
Anatomical:
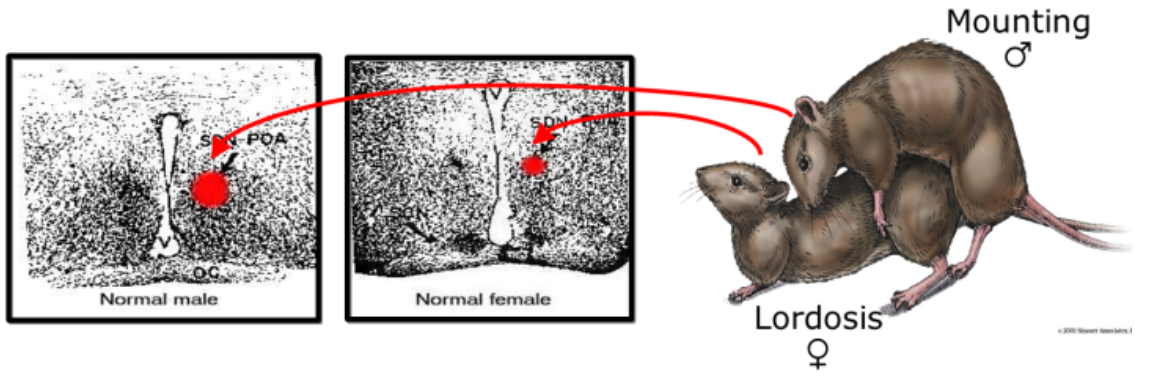
SDN-POA larger in males than in females
controls male-typical sexual behaviour
Empirical Evidence:
Measuring the electrical activity of this area - Male sexual activity increases the firing rate of SDN-POA neurons
Electrical stimulation of this area triggers male sexual behaviour
Volume of SDN-POA correlates directly with the level of sexual activity
Treat female rat embryos with androgens:
SDN-POA develops to male size (organisational effect)
male-typical sex. behaviour as adults (mounting behaviour) (only after androgen therapy -> activational effect)!
Courtship behaviour: HVC in songbirds
Behavioural: Singing is sex-specific (male-typical) behaviour
Anatomical:
In the songbird’s brain, some areas much larger in males than females
These are precisely the areas controlling song production (HVC)
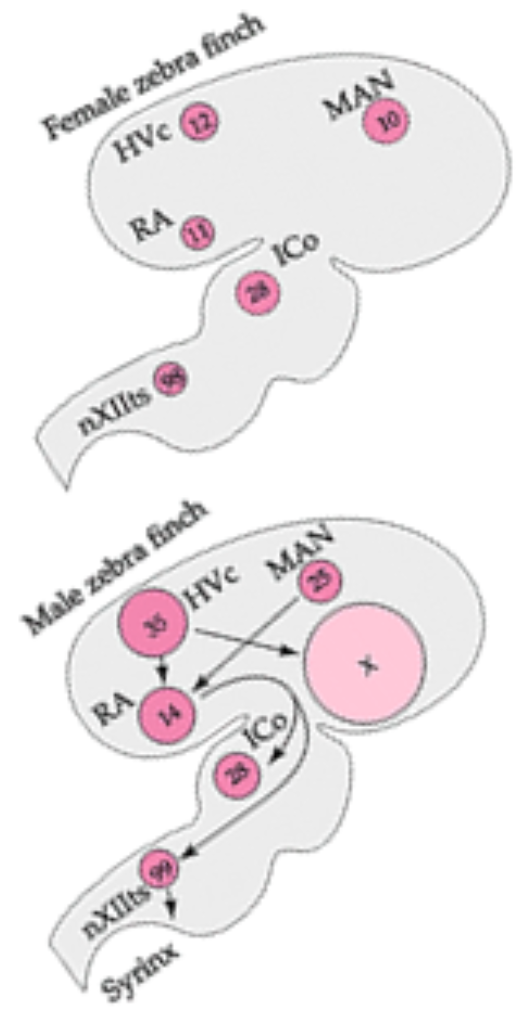
Experimental evidence:
Treat female hatchlings with sex hormones (androgens into the bloodstream, or estrogens directly into the brain – cf aromatisation hypothesis, lecture 3):
HVC develop to male-typical size (organisational effect)
This results in singing behaviour just like males
but only after androgen therapy (activational effect)
Sex Differences – Animals v Humans
In most animals, sexual behaviour is automatic:
Triggered by specific signals (whenever conditions are right)
These are generally controlled by ‘lower’ (non-cortical) brain areas (brainstem, midbrain, hypothalamic nuclei)
(empirical evidence: electrical stimulation of these areas and you will find the behaviour being displayed by the animal)
These behaviours are (relatively) sex-specific
(Relatively) uniform within each sex - the behaviour doesn’t change within the species
BUT: evidence for social learning of sexual behaviour even in rodents (recall lecture 4!)
In higher primates, sexual behaviour is even less automatic:
Less sex-specific - less clear to say that one behaviour is sex-specific to male or female monkeys
Less uniform within each sex - males and females have a variety of different sexual behaviours to exhibit
More under the control of ‘higher’ (i.e. cortical) brain areas (or so we like to think…) - less controlled by nuclei in the lower areas, e.g. brainstem
Human sex behaviour is most varied. → so most difficult to study.
Are there any other (more easily studied) differences between men and women?
Part 3. Non-Sexual Sex Differences
Potential areas of human cognitive sex differences:
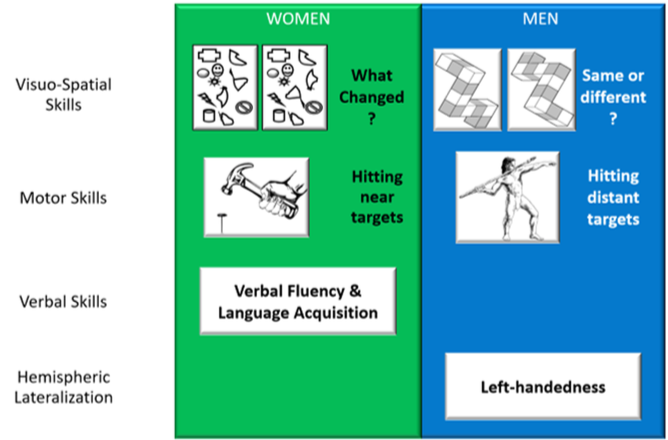
Are these differences ‘real’?
Very small!
Large role of culture, socialisation, learning:
Differences have become even smaller in recent years (e.g., Estes & Felker, 2011)
These differences are often context/experience dependent - females tend to do better in these tasks when the tasks are administered by a female experimenter than a male experimenter
Potentially instruction dependent - Wording of instruction affect the performance of the task.
At least for handedness and language development (and possibly some visuo-spatial tasks?), differences seem stable & similar across different cultures.
If these differences are therefore not as pronounced as they appear to be, then what exactly remains that is concrete?
It is found that only a few of these differences that are lasting and seemingly concrete are that left handness tend to be more present in men than in women, and that hitting distance targets is also more present in men than in women (but this is not really cognitive and is more physiological due to strength differences).
Part 4. Hemispheric Specialization
Different functional specialisation of the cerebral hemispheres - either side of the brain specialises in some tasks(“functional asymmetry”):
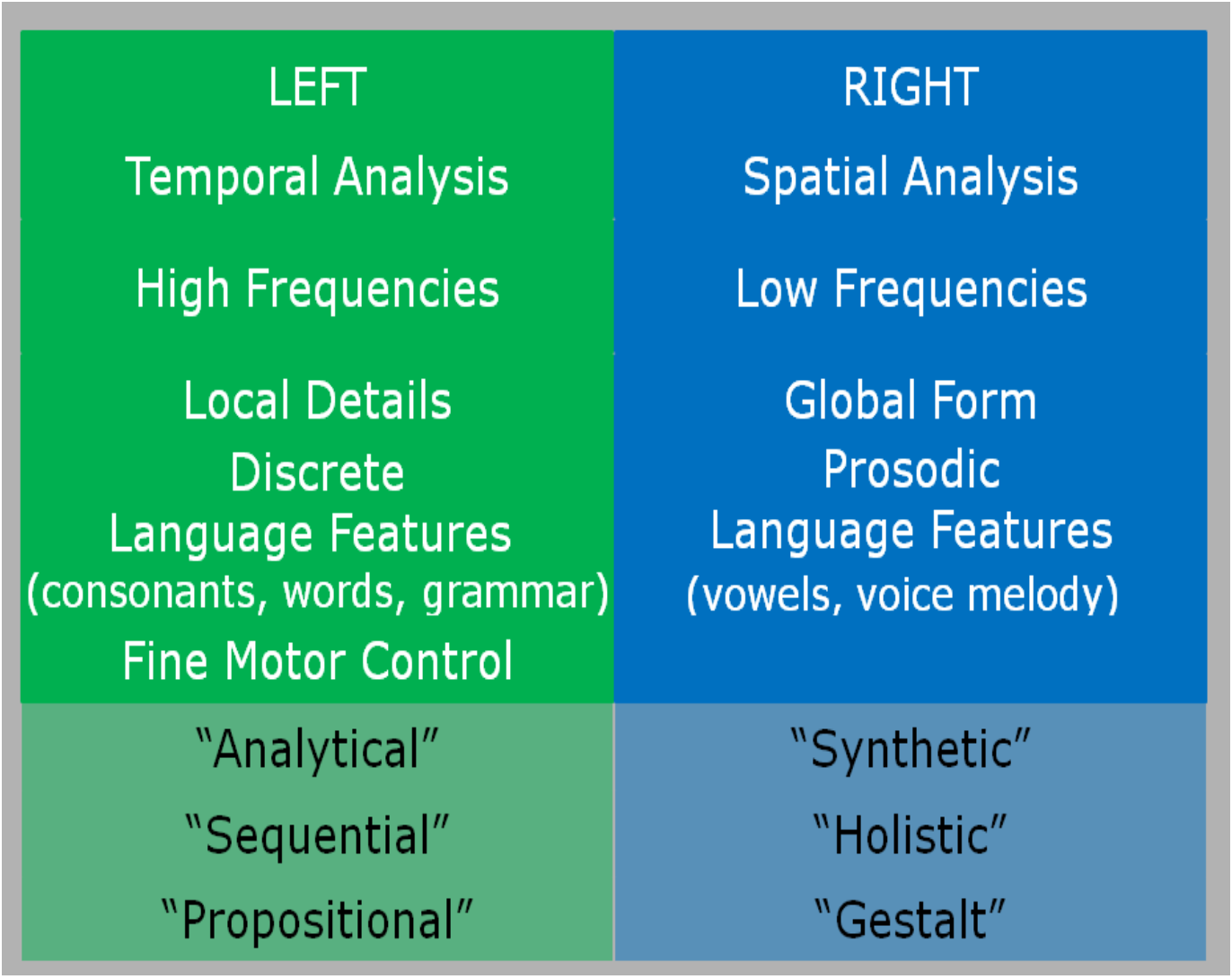
In the spatial domain, high frequency means rapid chang contrasts (rapidly changing from black to white to black to white and so on), whereas the low frequencies refer to slowly changing contrasts (an extended period of black, then an extended period of white, and then so on).
Discrete language features refer to consonants, whereas prosodic refers to vowels.
Empirical Evidence I: Invasive Procedures in Humans (not experimental - and is in the context of brain surgery where the brain surgery is required).
Wada test
Inject an anaesthetic into left or right internal carotid artery (to ‘knock it out’)
Assess each hemisphere’s language & memory functions with behavioural tests (e.g., picture naming)
Result (among others):
In most people, language functions almost exclusively in the left hemisphere
Split-brain surgery
Cutting the corpus callosum disconnects the hemispheres
Behavioural effects (examples):
patient cannot name objects presented in the left visual field, but can name objects presented in the right visual field
cannot name objects (without looking) by touching them with the left hand, but can name them when touching with the right (largely word processing and discrete language functions are done in the left hemisphere, and due to the brain and body operating contra-laterally, this means that when the two hemispheres are split, the right side of the brain does not have access to the language functions that the left does, resulting in the left hand not helping in naming objects because the right hemisphere of the brain just doesn’t have access to the words).
Empirical Evidence – Stimulation & Lesion Studies (experimentally induced only in animals!):
Electrical stimulation
Conducted during brain surgery
Stimulating particular areas interferes with particular tasks
For example, getting patients to talk while their brain has been exposed, and the professional will poke their brain in a specific area. When poking the brain in Broca’s area, the patient will stop talking. This is done to locate where this particular area of the brain is not in the exact same place for all patients, and the professional needs to know where it is for this particular patient in order for them not to damage key areas of the brain.
Lesion studies
Removal (partial or total) of one hemisphere
Behavioural tests to assess remaining functions
Patient studies (i.e., lesion studies)
Behavioural tests
Brain-imaging methods
Methods suitable for testing healthy participants:
Visual hemifield presentation (e.g., in a lexical decision task)
Dichotic listening (e.g., in the shadowing task) - participants wearing headphones hear nonsense syllables, and are then asked to repeat what they hear back. If the word enters the right hemisphere, reaction time will be slow, and if it enters the left hemisphere, reaction time will be fast. This is because of the contralateral wiring of the body. Right ear, left hemisphere, left ear, right hemisphere. Signals entering the right ear will enter the left hemisphere. This means that the word centres in the left side of the brain will be quicker in responding, as this is where the word centres are. The words entering the left ear will go to the left hemisphere, and will then have to communicate with the language centres in the left hemisphere of the brain, meaning the process is longer and thus reaction time is slower.
Part 5. Cognitive Sex Differences & Hemispheric Specialisation
Empirical Evidence – Sex-Specific Results:
Behavioural studies: 🚩🚩 (rewatch)
Women often show less behavioural asymmetry than men:
Especially at the end of the menstrual cycle (high levels of female sex hormones (estradiol & progesterone)
Brain-imaging studies:
Women ‘use’ both hemispheres in tasks where men ‘use’ mainly one hemisphere
(n.b.: ‘use’ = to show particularly highly correlated activity – obviously, the whole brain is used in each task!)
Clinical Evidence:
After a stroke, women recover language skills more quickly than men
Hypothesis: Women’s brains are less functionally lateralized:
More language functions in the right hemisphere (RH) than in men:
1. Women’s RH contains more language functions than men’s
Language processing of rVF words in LH, but language processing of lVF words (at least partially) in RH.
After a stroke, women can use remaining language functions in the undamaged hemisphere to ‘bootstrap’ speech
Right hemisphere less specialised for visuo-spatial task
Men outperform women in mental rotation tasks etc.
Possible reasons:
More equal development of both hemispheres in women? (go to 2)
Better interconnection of both hemispheres in women? (go to Part 6, 1.)
Galaburda-Geschwind Model🚩🚩
(Note: discussed here mainly because it’s historically interesting and shows what neuroscientists are looking for, not because it’s still believed to be correct!):
"Grand Theory“, integrating handedness - sex hormones - cerebral lateralization - cognitive skills and deficits - disorders of the immune system
Originally proposed in the early 1980s by Geschwind and Behan (Geschwind, N., & Behan, P. (1982). Left- handedness: Association with immune disease, migraine, and developmental learning disorder. Proceedings of the National Academy of Science, 79, 5097-5100)
Developed throughout the 80s by Geschwind and Galaburda (culminating in Geschwind, N., & Galaburda, A. S. (1987). Cerebral Lateralization. Cambridge, MA: MIT Press)
Today no longer discussed as a major model of cognitive differences
Hypothesis:
Pre-natal testosterone slows development of left hemisphere
Newborns’ brains differ due to pre-natal testosterone levels:
High levels (mostly boys): ‘asymmetrical hemispheres’ (LH less mature than RH)
Low levels (mostly girls): ‘symmetrical hemispheres’ (both hemispheres mature equally)
Evaluation:🚩🚩
Sex hormones influence brain development (at least in other animals)
But cognitive functions might not be as strongly localised! (this will make any model difficult to evaluate)
The model can account for
Larger number of male left-handedness
Superior visuo-spatial skills in men (e.g., mental rotation)
Faster language acquisition in girls
It cannot (directly) account for
Superior visuo-spatial skills in women (e.g., spatial memory)
Some special relationships between, e.g., visuo-spatial giftedness and reduced lateralisation
Part 6. Structural Sex Differences
Main candidates:
Overall brain size
Corpus Callosum🚩🚩
Corpus Callosum as a whole larger in women?
Splenium more bulbous in women?
Isthmus relatively larger in women?
Probably most frequently cited structural gender difference!
Mixed evidence – possibly only artefact? (e.g., Ardekani Figarsky, & Ssidtis, 2013, vs. Luder, Toga, & Thompson,2014)
Other candidate structures
Cortical:
Posterior temporal cortex: higher neuron density in women?
Temporal plane: larger size asymmetry in men?
Evidence is not yet totally convincing...
Sub-cortical candidate structures
INAH-3 of the hypothalamus larger in men?
Summary:
As yet, no clear evidence for gender differences in the anatomy of the forebrain
BUT: Anatomical differences might be too subtle to be detected easily & reliably!
Perhaps differences in local patterns of connectivity?
Or differences in the relative density of different neuron types in some brain areas?
Or differences at the level of neurotransmitter and receptor molecules?
Or we might be hunting for something that doesn’t exist…
The brain mosaic
Proposed by Daphna Joel and coworkers in 2015 (Joel et al. (2015). Sex beyond the genitalia: The human brain mosaic. PNAS, 112, 15468–15473)
Main findings of their study:
Several brain structures do show sex differences on average
But these differences are not distributed in an internally consistent way (i.e., in an individual’s brain, some of these structures might be female-typical, some male-typical, and some in-between):
L7 - Homeostasis
Part 1. General Background
Homeostasis
The human body consists of cells (Humans consist of 30 trillion cells. At least 80% of these cells are red blood cells. These cells are tiny and so barely make up a significant amount of volume within the human body). There are also 38 trillion bacteria. These are even smaller than red blood cells, and so make up even less volume.
Cells are living things – need specific conditions to survive & function:
e.g., temperature, level of acidity, salt & water, nutrients & energy, etc.
Suggests that multicellular organisms evolved from single cells living & moving in a saltwater environment
These cells also require a stable environment.
They can only survive small deviations from these stable environments.
Large deviations: cell functioning is disrupted. This leads to cell death.
In a multicellular organism:
Cells can’t move to a suitable environment
Cells on the body’s inside, cut off from ‘natural’ environment (recall lectures 1 and 2)
BUT: Environmental conditions change constantly:
Internally generated:
Nutrients used up
Waste products accumulate
Growth & reproduction …
Externally generated:
Temperature, humidity, etc.
Light & dark
Availability of nutrients…
Unlike other multicellular organisms, the vast majority of humans live in an environment where their source of nutrients is easy to locate and obtain.
In the real world, there are seasonal changes, which mean that proper nutrition to keep multicellular organisms healthy is not always available
For single-cell organisms, if the environment does not provide the nutrients that it needs, it can simply move to an environment where there are adequate nutrients.
In a multicellular organism, it is not possible for these cells to move to another environment to gain better nutrients. This means that cells must work & cooperate to create & maintain a suitable environment. This is where homeostasis comes in.
Homeostasis: To actively keep an organism’s internal states within a critical range
All of the processes that keep the organism’s internal state within the critical range of temperature, acidity, nutrients, etc.
Homeostatic Systems
are controlled by negative feedback loops (e.g., room temperature control)
Say there is a set point where we want our room temperature to be at precisely (e.g. 21 degrees Celsius). The actual level is not currently meeting the set point. What we want is a system that compares these two, and for this, we need a set of detectors that respond to the room temperature. The room temperature is the signal that is picked up by the detectors. This signal is fed to the control centre, where it is compared against the set point. If the actual level is on the cold side, it is below the set point, the control centre will take action by switching on the central system, and the room temperature will start to rise. At some point, the room temperature will rise so high to the point that it will exceed the set point. The central system will identify this and shut off the heat. This will then cause the heat levels to eventually begin to fall once more. This will then cause the central heating to switch the heat on again, and the cycle continues.
produce oscillating behaviour around the set point:
Is there a homeostatic control centre for eating behaviour?
Eating as a Homeostatic Process
Why do we need to eat
Nutrition (building & maintaining the body):
Essential amino acids (only 9 out of the 20 amino acids – building blocks of proteins – produced by the human body!)
Essential fatty acids (building blocks of fat (e.g., cell membranes))
Minerals (elements like iron, sodium, calcium: parts of all body structures (e.g., iron in red blood cells, calcium in bones, etc.))
Vitamins (no common chemical structure; organic nutrients needed in small amounts (e.g., chemical partners for enzymes, etc.))
Energy generation (powering the cells) :
Carbohydrates
Fat
Proteins (to a much lesser extent)
Control of eating = nutrient & energy regulation
Short-term control:
When to start a meal
When to end a meal
Long-term control: Food not constantly available => evolution of mechanisms to store energy
Release stored energy
Anticipate the need for energy & nutrient
Short-term & long-term control interact:
Meal size & frequency determines long-term body weight
so we’ll mainly look at short-term control…
Short-Term Eating Control
How are meal size & frequency controlled?
“We eat when we are feeling hungry – we stop eating when we have had enough”
Plausible, but:
We usually start eating before we feel hungry
We usually stop eating before the brain can receive satiety signals
Control of eating is related to ‘feeling hungry’, but that’s not the whole story...
Homeostatic factors
Biochemical signals indicating the state of the energy stores
Systems or structures to detect & interpret these signals
Non-Homeostatic factors
Learning: adapting the system to its specific environment
Mood as a non-adaptive factor
Part 2. Generating, Storing, & Utilising Energy
Energy
We eat to build up energy
Required by all chemical processes that make a body live
Energy generation (simplified!)
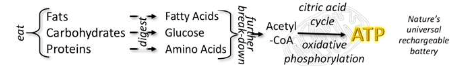
Available energy used for:

Over time, basal metabolism adjusts to caloric intake:
Less intake => less spending
Return to normal intake: reserves build up before basal metabolism increases
Reduced basal metabolism might increase life expectancy?
Energy Storage & Conversion
Excess energy can be stored in two ways:
Short-term: Liver & muscles store glucose as glycogen
Long-term: Fat cells store fat
Stored energy must be converted into usable energy (cells cannot utilise glycogen or fat):
Neurons: utilise almost exclusively glucose
All other body cells (except red blood cells): utilise glucose & fatty acids
Protein hormones (produced by the pancreas) convert energy:
Insulin: Converts glucose into glycogen
Glucagon: Converts glycogen back into glucose
Cells might need help in utilising energy:
Neurons: utilise glucose directly
Other body cells:
utilise fatty acids directly
utilise glucose with insulin
Part 3. Signals Involved in Eating Control
Insulin is important in regulating metabolic processes:
Produced by the pancreas
Released insulin allows the body to
make direct use of some of the glucose provided by a meal
store some of the glucose (in the form of glycogen) for later use
Insulin release is controlled by multiple systems
Cephalic phase (before a meal): signals from the brain
Digestive phase (during a meal): signals from gut hormones
Absorptive phase (after a meal): signals from the liver
How can we know this?
Disrupt the signal pathway and see what happens…
If insulin is that carefully controlled…
Are blood levels of insulin the crucial signal for eating control?
Hypothesis:
Insulin levels low => start meal
Insulin levels high => end meal
Experimental evidence:
Low blood insulin levels: the animal keeps eating
Inject some insulin => higher insulin levels: animal eats less
Problem: Injecting more insulin: the animal eats more!
Possible explanation:
High insulin levels convert all glucose to glycogen
Now glucose levels are low
→Signalling ‘hunger’
Are blood levels of glucose the crucial signal for eating control?
Hypothesis:
Glucose levels low => start meal
Glucose levels high => end of meal
Experimental evidence:
Low blood glucose levels: the animal keeps eating
Inject glucose => higher glucose levels: animal eats less
Supporting evidence: Glucose receptors in the hypo-thalamus (VMH)
Problems:
Glucose levels don’t vary much during the day
Diabetics: highly increased glucose levels, but often feel constant hunger
Injecting glucose into VMH does not make the animal eat less
Integrated view: Is the utilisation of glucose the crucial signal for eating control?
Hypothesis:
Start a meal when glucose levels in the liver (storage organ) are low
End the meal when the liver gets lots of glucose
Experimental evidence:
Functional anatomy: Liver sends signals to the brain via vagus nerve (autonomic NS)
Interfering with this signal
Either: cutting the vagus nerve (signal not transmitted)
Or: providing the liver (but not the rest of the body) with glucose (liver fails to signal low blood glucose levels)
Both reduce eating in hungry animals
Conclusion: when the liver does not signal “low glucose levels” (or when the brain does not receive the signal), the animal does not act (feel?) hungry
Problem: Reduced eating after cutting the vagus nerve is only temporary.
And so on…
Other possible hunger/satiety signals:
Blood levels of free fatty acids
Blood levels of CKK (a hormone released by the intestines in the presence of fat)
Gut distension
→ All showing basically the same problems as the possible signals discussed before…
Conclusion: probably...
no single signal is under all conditions necessary & sufficient for controlling meal size & frequency
several signals integrated & utilised in varying combinations…
Most likely site for such integration and control: hypothalamus
Part 4. Structures Involved in Eating Control
Hypothalamus: Input & Output
Central Eating Control
The hypothalamus contains two centres for eating control:
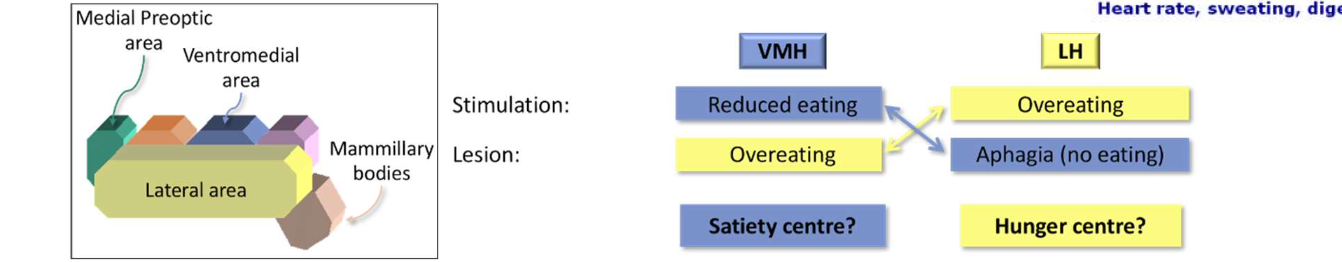
Evidence against ‘dual centre’ (two centres for eating control) hypothesis:
VMH-lesioned rats are less likely to eat bitter food than normal controls
Lesioned rats defend their new (pathological) body weight just as well as unlesioned rats
Probably ‘higher’ brain areas involved in eating control (particularly the limbic system)
Conclusion:
Not one single signal, system, or structure alone controls eating behaviour
All signals & structures discussed here may play a role
Eating (probably) regulated by a hierarchical network:
peripheral structures (liver, intestines)
brainstem centres (not discussed here)
hypothalamic nuclei (LH & VMH)
‘higher’ brain areas (particularly the limbic system)
Multiple systems & signals => redundancy
If one component no longer operates properly, other components can (at least partly) compensate for the loss
System is (relatively) failsafe!
Part 5. Non-Homeostatic Factors: Learning & Mood
The role of learning - mealtimes:
For humans, the time of day is the single most important factor determining when to start a meal (people start to feel hungry around learned mealtimes)
The same can be shown in animals:
Rats given 1 meal a day, at a fixed time
Begin to show anticipatory behaviour prior to learned meal time
Even when they had just eaten!
The role of learning - cues:
Stimuli not directly related to, but associated with, food can trigger a desire to eat (e.g., at a restaurant table…)
Again, a similar effect can be shown in animals:
Rats trained to associate a neutral stimulus with being fed
Once they have learned, food is made continuously available
But whenever the trained stimulus is presented, the rat will eat
Emotional learning – the role of pleasure
To some extent, taste preferences are ‘hard-wired’
Sweetness indicates that a fruit is ripe
Toxic substances often taste bitter
Fine-tuning of food preferences is learned
Some people find tarantulas delicious, some find pork disgusting…
Anticipation of very pleasurable food can stimulate hunger even if we have just eaten a big meal
A very monotonous diet reduces appetite (“sensory-specific satiety”)
Mood - Emotional Influences on Eating Control
Do people eat more when they are feeling ‘down’?
Experimental evidence (Meyer & Wallace, 1999a, b):
Neutral, appetitive, or emotional words presented subliminally (supposed to induce corresponding feeling)
Task: try to identify the word (participants do not succeed)
Afterwards, participants are offered food
Measure of interest: how much does each participant eat?
Result: participants primed with abandonment-related emotional words eat more
Part 6. Long(er)-Term Control
Fat tissue is subject to long-term regulation:
Some animals show seasonal variation in body fat (‘fattening up’ during spring, ‘slimming down’ during winter)
Surgical removal of fat tissue has only a transient effect on weight:
If fat tissue is removed, it either regenerates
or other fat tissues increase
Fat tissue plays an active role in eating control:
Fat cells produce a hormone (leptin) involved in eating behaviour
Control Function of Leptin – Empirical Evidence:
Genetically modified mice with two copies of the obese gene (‘ob/ob mice’)
2-3 times heavier than normal mice
have more & larger fat cells than normal mice
but cannot produce leptin
Brain monitors leptin levels
Leptin receptors in several cortical & hypothalamic areas
Leptin acts as a satiety signal
Injecting ob/ob mice with leptin (especially directly into the hypothalamus) results in drastic weight loss
Animals that have defective leptin receptors also become obese
Fat Tissue and Eating Control – A Model
Does leptin play a similar role in humans?
No evidence of defective leptin production in most obese people (obesity is rather associated with high leptin production)
Perhaps leptin insensitivity might cause obesity in humans? (no clear evidence yet)
Likely to be multiple, interacting reasons for obesity (Physiological, Emotional, Habit, etc...)
L8 - Biological Rhythms: Sleep
Part 1. Introduction to Biological Rhythms & A Historical Conundrum
Biological Rhythms: Regular changes in behaviour & internal states of living organisms
Ultradian: faster than a day
Sleep stages (different stages when we are asleep, e.g. light sleep and REM sleep)
Eating behaviour (we eat more than once a day, typically, they are at fairly regular intervals)
Circadian Rhythm: takes about a day
Sleep-wake cycle
Hormone production (for some hormones, not all)
Infradian: slower than a day
Hibernation cycle (in bears, for example, they hibernate for long periods of time)
Menstrual cycle (4 days for rats, 4 weeks for human women)
2. Sleep and Wakefulness
Traditional view: sleep as the passive default state
Hypothesis: The brain needs stimulation from the environment to wake up (without stimulation, it remains asleep)
Experimental evidence: Brèmer (1930s) transected cats’ brains by
Cutting between medulla & brainstem (this creates the encéphale isolé – “isolated brain”):
normal sleep-wake cycle
Cutting at midbrain level (cerveau isolé – “isolated forebrain”):
constant deep sleep
Reticulo-thalamic system wakes up the brain: 🚩
But something doesn’t add up: 🚩
Part 2. The Sleep-Wake Cycle
General
Several biological systems co-vary with the sleep-wake
cycle (see figure):
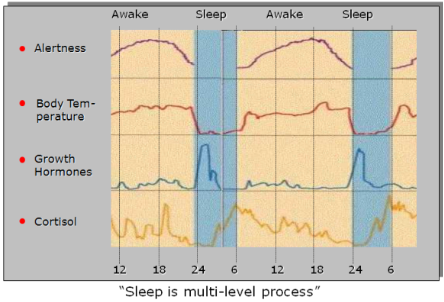
(of particular interest to) Cortisol:
Cortisole is released when: hypothalamus releases CRH → the pituitary then secretes ACTH → which causes the adrenal gland to secrete cortisol
In response to stress or low blood sugar
Function:
Gluconeogenesis (‘sugar-new-making’, i.e., sugar ‘from scratch’, not from glycogen)
Immune suppression
Further effects:
Increases blood pressure
Inhibits CHR (negative feedback loop!) 🚩
Cycles controlled by two factors:
Internally generated (‘endogenous’) by (one or more) ‘biological clocks’
External cues (‘zeitgeber’) reset biological clocks
2. Supra-Chiasmatic Nucleus
SCN is the ‘master pacemaker
Lesions to the supra-chiasmatic nucleus (SCN) (or age-related neuron loss in this area) disrupt circadian rhythms
Nucleus of the anterior region of the hypothalamus (Situated directly above the optic chiasm)
Input:
Connected to retinae (retino-hypothalamic tract)
Sensitive to light
Output:
Signals to pineal gland
SCN generates circadian rhythms:
SCN cells show circadian firing patterns
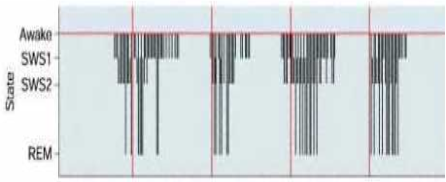
🚩
Activity stimulates pineal gland -> Pineal gland produces melatonin → Melatonin promotes sleep
Firing patterns of SNC neurons remain circadian (but not entrained!)
even without zeitgebers
even in a preparation outside the body!
Retinal input signals light versus dark:
Light/dark cycle ‘reset’ the biological clock (‘zeitgeber’)
Blind people without light perception show abnormal circadian rhythms
Treatment of blind (light-insensitive) people with oral administration of melatonin
Part 3. What Distinguishes Sleep From Wakefulness?
Hypothesis: the sleeping brain is less active than the wakeful brain
Test: measure brain activity during sleep and wakefulness
INTERLUDE: How to Measure Brain Activity
I: Measuring metabolic activity
Basic logic:
More active neurons need more oxygen & glucose
Oxygen or glucose levels in the brain = index of how much activity is going on
Localize where the oxygen and glucose goes precisely = know where higher or lower activity is going on
Functional Magnetic Resonance Imaging (fMRI): measures oxygen levels through the different magnetic properties of oxygen-rich versus oxygen-poor blood
Positron Emission Tomography (PET): measures distribution of a radioactive substance (typically glucose)
II: Measuring electrical activity
Basic logic:
For neurons, being active = exchange of electrical signals
A single neuron’s signal too small to measure at a distance
Combined signals of many neurons strong enough if the following conditions are fulfilled:
The group consists of sufficiently many neurons
They are aligned in parallel (so that their electrical currents all move in the same direction)
Their activity is synchronized (i.e., all fire at the same time)
Electroencephalogram (EEG): 🚩
Electrodes on the scalp (i.e., some distance from neurons)
EEG measures synchronous firing of groups of neurons
Small, rapidly changing groups: Desynchronized EEG activity – highly active brain
Large, persistent groups: Synchronized EEG activity – less active brain
(cont.) Hypothesis: the sleeping brain is less active than the wakeful brain
Reduced metabolic activity during sleep
Reduced electrical activity during sleep
Part 4. Sleep: A Multi-Stage Process 🚩
… sleeping brain is less active - except in REM sleep!
Electrical & metabolic brain activity almost like waking state
Most vivid dreaming
Sleep stages are characterised by specific bodily states:
Sleep stages are characterised by specific
electrical brain activity:
… in combination with specific muscle activity:
2. REM Sleep
Rapid-Eye-Movement (REM) sleep is ‘paradoxical’:
Body held in paralysis, while
Brain activity resembles waking state
Sleep stage with the most vivid dreams
FOOD FOR THOUGHT:
Dream content often negative: helplessness, falling, running without moving forward, threat ...
Psychoanalytical interpretation: Subconscious mind as a ‘battleground’: We want something forbidden(most likely sex with our parent), but we know it’s forbidden, so we hide it from ourselves, and punish ourselves for wanting it.
Psychobiological interpretation: Subconscious mind perceives the body (we are, in fact, immobilized & helpless!)
3. Sleep stages follow systematic progression
During sleep, REM and non-REM phases alternate
Time spent in each phase changes systematically:
More time spent in deep sleep (stages 3 & 4) in the 1st half of the night
More time spent in REM & light sleep in the 2nd half of the night
Part 5. Multiple System Regulate Sleep & Wakefulness
Network involved in sleep & wakefulness – overall:
Basal forebrain - Thalamic nuclei - Hypothalamic nuclei - Brainstem nuclei
Wakefulness:
ARAS
Basal forebrain
Locus coeruleus
Promotes wakefulness
Produces noradrenalin (Recall: stress response)
Virtually no noradrenalin release during sleep
Non-REM sleep:
Basal forebrain:
Most neurons involved in increasing alertness
Some involved in triggering SWS:
Stimulation: can induce SWS
Lesion: can prevent SWS
Raphe nuclei:
Produce serotonin
Promote non-REM sleep
(Remember: non-REM sleep not a passive default state in the absence of environmental stimulation!)
REM sleep:
Pontine nuclei:
Control REM sleep:
Stimulation: triggers REM sleep
Lesion: prevent REM sleep
Note: Lesion of nearby nuclei prevents REM paralysis!
Produce acetylcholine
Enhanced levels of ACh during REM
ACh “activates” the brain (recall role of ACh in learning)
FOOD FOR THOUGHT:
The dreaming brain (figure on the right):
Similarities of REM sleep and schizophrenia - Is schizophrenia a form of ‘waking dream state’?
Part 6. Beyond the Day-to-Day: Life-Span Changes & Evolutionary Perspective
Life-Span Changes of Sleep Patterns
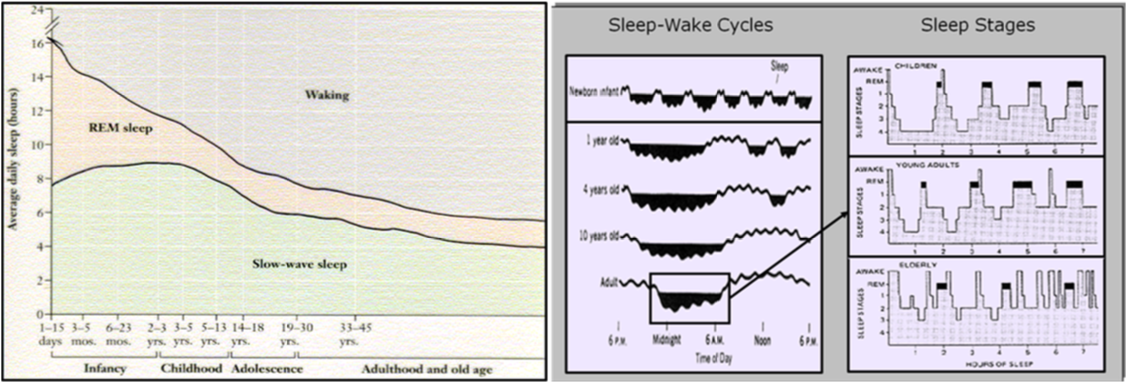
How Important is Sleep?
Sleep deprivation – Animal studies: Prolonged sleep deprivation is fatal:
Initially decrease in
Subsequently:
Body temperature
Failure of immune system
Metabolism
Loss of resistance to infection
Appetite
Death
‘saving energy’?
Caveat: This might be due to the stress of being kept awake, not to lack of sleep as such!
Sleep deprivation in humans:
Prolonged sleep deprivation results in impaired physical & cognitive function
Impaired speech
Impaired memory
Hallucinations etc.
Obviously, one cannot test whether this would eventually result in death!
After sleep deprivation, the body will try to ‘catch up’ on missed sleep
Can be selective: if you miss out on REM sleep, structure of next night’s sleep will change to include more REM sleep (“REM rebound”)
Large inter-individual variety:
Most people need approx. 8 hours of sleep
But some function on an average of 4 hours or less
Evolutionary perspective: The paradox of sleep
Sleep is dangerous (being unable to move & to detect dangers)
BUT: All animals with brains show some form of sleep (Even insects have periods of rest)
In higher animals, sleep is controlled by a complicated system of interacting neuro-endocrine functions
Some animals have even developed a method to stay awake while sleeping!
Therefore, there has to be some evolutionary advantage in sleep!
We just haven’t fully worked out what it is yet… (but DNA repair might play a role)
L9 - Evolution
Slide 2
Outline - (What you should understand from this lecture)
A functional definition of evolution and some evidence supporting it.
How do we apply evolution to human behaviour?
An explanation of how trade-offs influence adaptation and how these shape the human experience, from death to thought.
Slide 3
Recap
Part 1: What is Evolution
Part 2: The evidence of evolution
Part 3: What evolution does
Part 4: Evolution and Psychology
Slide 4 - Part 1: What is Evolution
Slide 5
What’s the question that evolutionary theory aims to explain?
Why do we see the kinds of living things and behaviours that we do in the world?
Charles Darwin, 1809–1882
Slide 6
Darwin’s Finches
The classic example:
Finches on the same island shared features of other finches on that island, but also showed adaptations (beak shape) that were more similar to finches on different islands. The beak shapes were adapted to eat different things.
Darwin proposed:
Adaptive Radiation (i.e., Evolution)
Slide 7
Tinbergen’s four questions:

Mechanism, adaptive value, ontogeny, and phylogeny
Question 1 - How now? (Proximate Synchronic)
Mechanism
Physiology, molecular mechanism, external stimuli, etc.
It serves a purpose; it is functional
Question 2 - Diachronic proximate (How historical?)
Ontogeny
How does the behaviour develop over the lifetime of the organism, how much of the behaviour is learned/innate, and what factors influence development
What is the history of the mechanism, and what is the history of what is currently happening? How did it develop? How did it come to be?
Question 3 - Synchronic Ultimate (Why now)?
Adaptive value
How does this contribute to the organism’s fitness (survival and reproduction)
Why is this behaviour occurring now, and what are the implications of this behaviour on the survival and continuation of the organism/species?
Example: competition in males has evolutionarily been proven to be good for reproduction. Males that do not compete evolutionarily do not reproduce.
Question 4 = Diachronic Ultimate (Why Historical)?
Phylogeny
What is the ancestral trait? What were the selective pressures that led to this trait’s evolution?
Why does a species exhibit a certain behaviour? Looking at this from a historical context. For example, most mammals have exhibited a certain behaviour (e.g. breastfeeding) for the past 30/40/50 etc million years, which is why the current iteration of the species exhibits this behaviour. The fact that the past iterations of this species used breastfeeding (and they have survived) means that there has to be an evolutionary advantage to this behaviour.
Slide 8
Evolution: a functional definition
Three requirements
Reproduction
Variation
Differential Survival (differential selection)
Any system that has these three things evolves.
Slide 9
Evolutionary Algorithms: Genetic Algorithms
start with random designs
Then begin the following:
evaluate them based on some fitness function
let individuals reproduce in proportion to their fitness
add some mutation
repeat
Evolutionary algorithms are used to design things.
The algorithm (a set of rules) is used to generate and select among various alternative solutions.
The fitness function is used to ‘select’ the best performing solutions for reproduction.
These are then reproduced with a little bit of ‘variation’.
Usually, the very best solutions are preserved also to continue competing.
Slide 11 - Part 2: The evidence for evolution
Slide 12
Some empirical evidence for Evolution
The fossil record (species that don’t exist currently, gradual change over species over time and space)
Vestigial organs — organs that no longer perform the function for which they evolved
(Ostrich wings, eyes in blind mole rats, the human tail, the whale pelvis)“Why Evolution is True” by Jerry Coyne is a good place to look for more evidence.
Slide 13
Pseudogenes (dead genes, e.g., broken Vitamin C gene in humans (GLO)) - this is to do with the environment of which humans once lived and the food sources available at the time have different levels of vitamin c in the food source.
Unintelligent Design:
The flounder eye
Child mortality due to small female pelvis: 1 in 10 maternal deaths in the 1700s (Chamberlain 2006); 100 infant deaths per 1000 births in 1900 (ABS).
Three-foot laryngeal nerve from the brain to the larynx!
“Why Evolution is True” by Jerry Coyne is a good place to look for more evidence.
Slide 14
Some (more) empirical evidence for Evolution
Convergent evolution (placental mammals and marsupials, independent eye evolution in over 40 groups of animals): Different but similar solutions to the same problem
Islands have missing species found on continents (the same missing species; no freshwater fish in Hawaii — animals that don’t disperse over oceans are not found on islands)
Evolution in the laboratory, artificial selection
No other theory offers the same predictive and explanatory power for why we see what we do in the world. It doesn’t mean it’s completely true (what is?), it just means that it’s an incredibly good predictive model much like Newtonian physics.
“Why Evolution is True” by Jerry Coyne is a good place to look for more evidence.
Slide 15
How do we know genes influence behaviour?
For biological evolution, it’s 99.99% about the genes
Is this a protein for anger?
Almost all psychological traits are polygenic — influenced by multiple genes.
Slide 17
Most mental disorders are polygenic
(not single-gene Mendelian traits)
Slide 18
The Watershed Model of Mental Disorders
Each human carries about 500 to 2000 deleterious point mutations inherited from ancestors — with about half of those expressed in the brain. This means that there are many different ways to change the genetic information within the genes. The different ways to change it are the point mutations. The individual base pairs get “changed”. This will therefore change the protein slightly. It can still work, but is a slightly different gene now.
Each will have 500 - 2000 of these deleterious mutations. Deleterious means bad. However, because humans have two parents, there are two genes, one from each parent. This means that the mutations may not be expressed, as maybe the other gene does not have the same deleterious mutation, or maybe the mutation is in a different area.
For males, this can be different. Males will have more mental disorders because they only get one X and one Y gene, whereas women will have 2 X genes, thus meaning that if one X gene is bad in women, there is a chance that it just will not be expressed and will only express the non-deleterious gene. But if the male has a deleterious mutation in his X or Y gene, it is more likely to be expressed. There is also more variation that is possible in males.
Because of this, males are more likely to have more variation in their expressed genes because they only have one of the two gene copies to express.
55% of coding DNA is expressed in the neurons of the human brain - this means that more than half of all DNA can contribute towards behaviour.
Keller & Miller, 2006, Resolving the paradox of common, harmful, heritable mental disorders. BBS
A simplified way to explain the watershed model is that all the genetic information is tributaries that flow into one larger river. This larger river is the behaviour expressed from the build up of genetic information.
Slide 19
Part 3: What evolution does
Slide 20
Adaptation requires trade-offs
Otherwise, we would all evolve to live forever (survival) and have an infinite number of offspring per second (reproduction).
Trade-offs are the rule of life: we can do one thing only if we give up another.
Common economic trade-offs:
speed-accuracy trade-off
exploration-exploitation trade-off
size-number trade-off (bigger means fewer)
growth-reproduction trade-off
short-long term trade-off (Inter-temporal discounting, r (rate of growth) vs. K (long term survival) selected species)
Slide 21
Why do we die? - (Death is an inter-temporal trade-off)
Henrietta Lacks
In one sense, we don’t die: our germ line is immortal. Only the soma dies (senescence). Henrietta Lacks’ germ cells make up the HeLa cell line, cells from a cancerous tumour (20 tons now exist and are responsible for numerous medical breakthroughs, The Immortal Life of Henrietta Lacks). Some of her cells don’t die — but “she” died of cancer in 1951.
Death is basically a life-history trade-off (early v. late; Kirkwood & Austad, 2000, Nature). There are several evolutionary theories (they are not mutually exclusive and may all explain death).
Disposable soma theory: extrinsic mortality weakens the forces of selection in late life.(If most of a species die young, then selection can’t act to counteract late-acting deleterious mutations. If 90% of wild mice die in the first year (from cold), then selection for long-life genes only benefits 10% of the population. If selection for long life requires resources that could keep them warmer, then it’s not going to happen. Fruit fly’s selected for long life in the lab (by artificially delaying reproduction). Long-lived flies reproduce less, supporting disposable soma and pleiotropy theories.)
Antagonistic pleiotropy: “good early” genes can be selected for even if they have “bad later” effects. Risk-taking in males, using your resources to assist your offspring (instead of your own longevity: semelparous species reproduce only once and often go out with a massive expenditure of resources into that one reproduction, like salmon, wheat, and many “annual” spiders).
Mutation-accumulation: This one is less well supported. All cells age, but mutations may not be the primary reason.
Hydra does not age or die of old age
Read: Why do we die, Hills, Psychology Today, Statistical Life
Slide 22
A core trade-off in learning? - Less is sometimes more
A neural network designed to learn aspects of language performs better when the attention span (memory span) of the network starts off small and grows gradually.
Elman. 1991. Incremental learning, or the importance of starting small.
In: Proc. of the 13th Annual Conf. of the Cognitive Science Society, pp. 443–448. Hillsdale, NJ: Erlbaum
Slide 23
Why aren’t we smarter already?
more memory, better focus, more willpower
Think about the evolution of attention —
How focused do you want to be?
Slide 24
What should we pay attention to? - (Attention is an explore-exploit trade-off)
Attentional focus is a trade-off between too much and too little.
Too much, and you can never change your mind, never attend to new alternatives, never move on (OCD)
Too little and you don’t attend long enough, you never get started, if the payoff isn’t fast enough, you don’t get it.
Inverted-u shaped function
Hills & Hertwig, “Why aren’t we smarter already,” Current Directions in Cognitive Psychology, 2011
Slide 25
Part 4: Evolution and Psychology
Slide 26
Vertical Transmission
Where do different behaviours come from?
What are some possible sources?
Culture (Horizontal transmission) — things like beliefs, social norms, etc
Learning from the environment — Behaviourism/Skinner
Evolution (vertical transmission)
Slide 27
The application of Evolution to Psychology involves understanding how our past influences our future
Partly, this is about our entire evolutionary history — from bacteria to Bach.
“Evolutionary Psychology” is often based on the idea that the time that defined human evolution was about 50 to 200 thousand years ago, in the Environment of Evolutionary Adaptation (for humans): Humans lived in small polygamous groups (one husband, many wives), they used tools, they hunted and collected food, and they fought with one another.
Evolutionary Psychology suggests that our behaviour/mind is “adapted” to the EEA, like the finch’s beak is adapted to eating certain foods. Though in many instances, we can imagine this adaptation is much older.
Slide 28
Parental investment theory
Parental investment theory (Trivers, 1972; Williams, 1975) - “Individuals of the sex investing less will compete among themselves to breed with members of the sex investing more, since an individual of the former can increase its reproductive success by investing successively in the offspring of several members of the limiting sex” (p. 141, Trivers, 1972).
According to parental investment theory...
What do women want in a man?
The best possible mate — one who will invest in their offspring; he has resources.
What do men want in a woman?
Men want reproductively viable mates (young women)
Slide 30
Grandparental investment
Mothers have maternal certainty — they know they are the mother. Fathers have paternal uncertainty — it’s hard to be sure.
Thus, as grandparents go, mother’s-mother should invest more than father’s-father.
Bishop et al., 2009. Differential investment behavior between grandparents and grandchildren: the role of paternal uncertainty. Evolutionary Psychology.
In many surveys of grandparental investment, mother’s mothers are notoriously more involved in raising, gift-giving, emotional investment, etc, than other grandparents. And the relative amount of each grandparents contribution follows evolutionary theory.
Slide 31
The Cinderella Effect
The increased incidence of stepchildren being abused/murdered by stepparents as opposed to genetic parents.
Daly & Wilson (2001) analysed a Canadian data set found 74 fatalities among 28.3 million genetic fathers, and 55 among 0.17 million stepfathers. That is, 2.6 per million vs. 321.6 per million, it represents a 100-fold increase in death rate for stepfathers killing their stepchildren. This has been found repeatedly in other populations.
Slide 32
Who should take more risks?
If women are basically guaranteed a sexual partner(s), while men must compete for women (in a polygamous society as likely in our evolutionary past), then... men must compete for mates...
Parental investment theory says that males should compete for mates and that females should select them.
Slide 33
Young male syndrome: Males take more risks when other males are around
In our long evolutionary history (not just human history) males don’t just win battles, they win mates.
Slide 34
Men take more risks than women
Found many times over...
Wilson & Daly, 1985
There is a “pacifying effect” on males who establish a relationship with a female partner. Marriage reduces male risk-taking.
Slide 35
Men drive fast and die young
Slide 36
Balloon analogue risk task
must pump up a balloon to get money, but risk it popping... who will pump more, men or women?
Slide 37
Men risk more
From Fischer & Hills, Young Male Syndrome and the Baby Effect (2012).
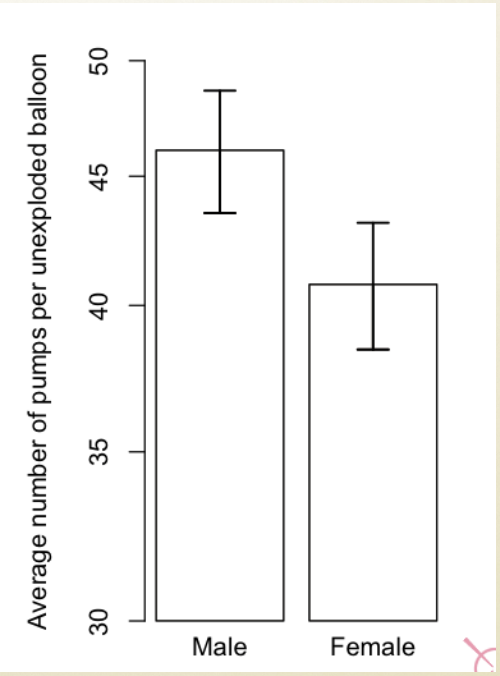
Slide 38
Paired with different social partners...
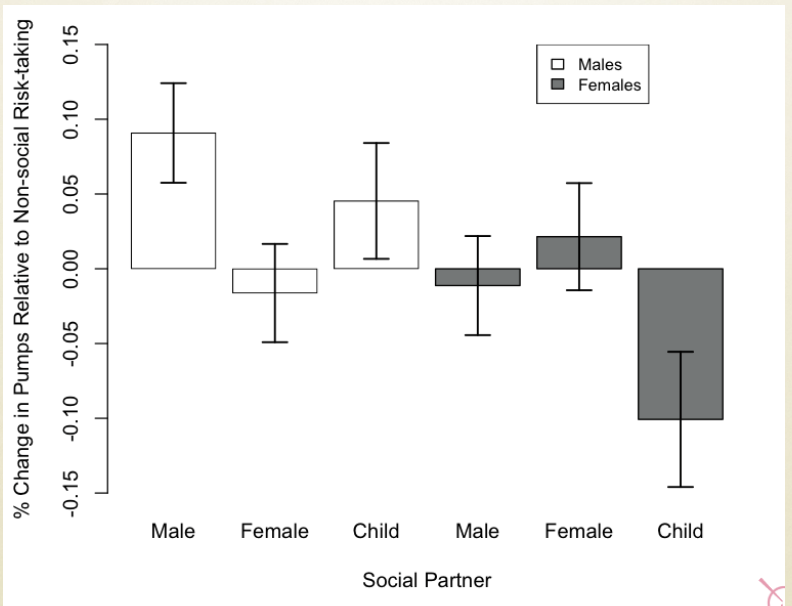
Slide 39
Violence associated with suicide should be especially common among men with no mate

Pedahzur et al., 2010. Altruism and fatalism: the characteristics of Palestinian suicide terrorists. Deviant Behavior.
Slide 40
Conclusions
Evolution and evolutionary psychology provide a theoretical ground for explaining much of “innate” human behavioural drives.
These include our altruistic behaviour, our aggressive behaviour, our reproductive behaviour, and so on.
However, evolution and evolutionary psychology are but one of many potential alternative explanations — and there are even competing ideas even within these approaches. Many of these alternatives have yet to be empirically tested.