Exam 2024
Describe the three carbon pumps in the ocean. Which of these are most important for the global carbon balance? explain why biocalcification by coccolithophores, pteropods, etc. increases CO2 exports from the ocean to the atmosphere
Answer: the three carbon pumps in the ocean are the biological pump, the microbial pump and the carbonate pump.
The biological carbon pump is driven by photosyntehsis in the surface of the ocean. Phytoplankton in the euphotic zone take up dissolved CO2 and convert it into organic matter. A part of this organic carbon is trasnferred through out the food web, but a significant amount sinks to the bottom as particulate matter, well known as marine snow, and comprises of dead organisms, fecal pellets, and other aggregates. This is how carbon sinks are formed at the bottom of the ocean, going from the atmosphere and ocean to the depths.
The solubility pump is where CO2 dissolved in the seawater at the ocean’s surface, with the speed of this process depending on the temperature. COlder waters can hold more dissolved CO2 than warmer waters. In high latitude regions, surface waters become more dense due to cooling and increased salinity and sink to the deep ocean, carrying the dissolved inorganic carbon with them. This circulation driven trasnport removes carbon from the atmosphere and takes place across a timescale of hundreds to thousands of years. This pump plays a crucial role in distributing the dissolved carbon within the ocean.
the carbonate pump involves the production of calcium carbonate shells and skeletons by marine organisms such as coccolithophores, foraminifera, and pteropods. These organisms combine calcium ions with carbonate ions in the sea water to form solid CaCO3 strucutres. When these organisms dies, their shells sink to the bottom and contribute to the calcerous sediments on the seafloor. This process looks like it is another pathway for carbon sequestration.
In terms of the global carbon balance, the biological and solubility pumps are the most significant. The biological pump is the dominant mechanism for long term removal of carbon from the atmosphere — ocean system, while the solubility pump governs the larger scale exchanges between the surface and deep ocean. In contrast the carbonate pump does not fucntion as an efficient sink for atmospheric CO2 and instead acts primarily as a modifier of the oceans carbon chemistry, often in a way that enhances CO2 release.
The reason biocalcification tends to increase the export of CO2 from the ocean to the atmosphere lies in the chemistry of the carbonate system. When calcium carbonate is formed, carbonate ions are removed from the seawater. This removal disturbs the equilbrium among the dissolved carbon species. The reduction in the carbonate concentration leads to a shift in chemical equilibria that increases the concentration hydrogen ions and drives the system towards higher concentrations of dissolved CO2. The increase in dissolved CO2 in the surface waters raises the partial pressure of CO2, making it more susceptible to trasnferring back into teh atmosphere. Although calcifying organisms incorporate carbon into their solid strucutres, the net effect of this process is to reduce the ocean’s capacity to store CO2 and to enhance outgassing. This illustrates a key paradox in ocean biogeochemistry: biological processes can simultaneously contribute to both teh removal and release of carbon dioxide, depending on the pathways involved.Beer-Lambert’s law describes the attenuation of monochromatic light in a homogeneous medium. Write down the law in mathematical notation and explain the different terms and their units. What are the major sources of light attenuation in open water environments? How do you expect the color of down-welling light to change with depth a coastal environment like the Oslo Fjord?
Answer: The mathematical equation for beer-lambert’s law is as follows:
I(d) = I0e-kd, where I(d) represents the light intensity at a specific depth, I0 is the light attenuation at the surface, e is the natural logarithm, k is the attenuation coefficient with the units of inverse length (m-1), and d is the depth. The light attenuation decreases exponentially as you go deeper.
light attenuation in the open ocean environments comes from several sources. The first is the absorption of the photons by teh water molecules, which is strongly dependent on the wavelength. Biologicla components such as phytoplankton also playa role in attenuation, where their pigment absorb specific wavelengths of light for photosynthesis. Dissolved organic matter (DOM) also contributes to absorption, specifically in the blue and ultraviolet parts of the spectrum, while suspended particles can both absorb and scatter light. In clear open ocean water, attenuation is low and is dominated by the optical properties of pure water and phytoplankton, whereas in coastal environments teh presence of suspended sediments and dissolved organic materials significatly increase attenuation.
A key feature of light attenuation in seawater is its dependence on wavelength. Red light is absorbed more rapidly, while blue green wavelengths peentrate the deepest. As a result, teh spectral composition of light changes with depth. within the first few meters, red wavelengths are rapidly removed and the remaining light becomes dominated by blue and green wavelengths. At greater depths, blue light also gets attenuated, leaving a narrower band of blue green light. In coastal environments such as the oslo fjord, these changes are more pronounced due to higher concetrations of suspended particles and dissolved organic matter. Near the surface, the light field initally resembles that of the open ocean, but red wavelengths are quickly lost within a short depth range. As depth increases, teh enhanced turbidity leads to stronger attenuation of blue light as well, o that the dominant colour shifts towards green rather than blue. With further depth, the overall intensity decreases rapidly, and the remaining light becomes light becomes dim and spectrally restricted. Consequently, compared to the open ocean, coastal waters exhibit a shallower penetration of light and a shift in dominant wavelengths towards the green and even yellow part of the spectrum. This environment has important ecological implications, a it constraints the depth distribution of photosynthesis and influences the types of pigments used by marine organisms.What is Ekman transport and how does it affect the direction of the major ocean gyres?
Answer: Ekman transport refers to the net movement of the surface of the ocean water driven by wind stress and modified by the coriolis effect. When the wind blows across the ocean surface, it exerts a force that moves the upper layer of the water. Due to the Earth’s rotation, this motion is deflected to the right of the wind direction in the northern hemisphere and to the left in the southern hemisphere. As the movement is transferred further downward through the water column via turbulent mixing, each succesive layer is defelcted further creating a spiral strucutre of current directions known as the Ekman spiral. Although individual layers move at different angles, the net trasnport of water in teh ekman layer is 90 dgrees to the wind direction. This porcecss causes the formatin of th emajor ocean gyres. In subtropical oceans, the wind systems consist primarily of trade winds, which blow westward near the equator, and the westerlies which blow eastwards at mid latitudes. Ekman transport causes casues the surface waters under the trade winds to be deflected pole ward, while waters under the westerlies are deflected equatorward. As a result, the water convergres in the central regions of the ocean basins. This convergrence leads to a buildup of water, creating a slight elevation of the sea surface in the gyre center. The direction of this circulation is deterined by the interplay between pressure gradient and the coriolis effect. In the northern hemisphere, teh flow around teh center is clockwise and in the southern hemisphere it is counterclockwise.assume a pelagic food chain with an even or odd number of trophic levels. Which process will control phytoplankton biomass when the number of trophic levels is even? What happens if we add a top predator level to a food chain?
Answer: in pelagic ecosystems, the control of phytoplankton biomass depends critically on the number of trophic levels and the interactions between them. The key distinction lies betweenn bottom up control, where nutrient availability regulates primary production, and top down control, where grazing pressure by higher trophic levels determines phytoplankton abundance.
when a pelagic food chain contains an even number of trophic levels, phytoplankton biomass is primarily controlled by bottom up processes. In such systems, the top predator supresses the population of herbivores indirectly through trophic interactions. For example, in a four level food chain consisting phytoplankton, herbivorous zooplankton, small predators, and top predators, the top predators reduces the abundance of the intermediate predator. This allows herbivore populations to increase, leading to stornger grazing pressure on phytoplankton. However, because the herbivores themselves are not strongly suppressed, the system ultimately reflects teh availability of nutrients that support phytoplankton growth. Thus phytoplankton biomass remains relatively low and closely tied to nutrient supply, indicating bottom up control.
In contrast, when the number of trophic levels is odd, phytoplankton biomass tends to be controlled by top down processes. In a three level system, for instance, predators reduce herbivore populations, therby releasing phytoplakton from grazing pressure and allowing biomass to accumulate. This alteration between top down and bottom up control depending on trophic level parity is a classic example of a trophic cascade.
Adding a top predator to a food chain alters this balance by increasing the number of trophic levels by one, therby switiching from even to odd and vice versa. When a top predator is added to an even level system, it introduces a new level of predation that supressses teh previous top predators, a decrease in herbivores, and ultimately an increase in phytoplnkton biomass. In other words, adding a top predator can reverse the direction of control and shift the system toward top down regulation.you have 0.2 L of water sample on a glass fiber filter and extracted it in 2 mL ethanol. A flourometric analysis reveals that this extract contains 0.2 µg / mL of chlorophyll a. How many µg / L of chlorophyll a were there in the original sample? Justify your answer.
Answer: the amount of chlorophyll in the 2 mL ethanol sample = 0.2 µg/mL x 2 mL = 0.4 µg chlorophyll
So we had 0.4 µg of chlorophyll a in 0.2L of water sample.
Therefore in 1L, we have 0.4/0.2 (µg/L) = 2 µg/L of chlorophyll awhat is the difference between net and gross primary production? how does net primary production relate to the compensation depth? what is the difference between critical depth and compensation depth?
Answer: net primary production is how much carbon is left after respiration. gross primary production is the total amount of carbon produced through out photosynthesis.
compensation depth is where the rate of primary production and respiration is equal, so that there is no net loss or gain. Above the compensation depth, the NPP is positive, and below the compensation depth, the NPP is negative
critical depth is for the whole water column and compensation depth is for a specific individualwhat is the basin wide tilt and what is a walker cell? what happens to the basin wide tilt during El Nino events?
Answer:What is recruitment overfishing? Mention at least three mechanisms that can be used to reduce the risk of overfishing.
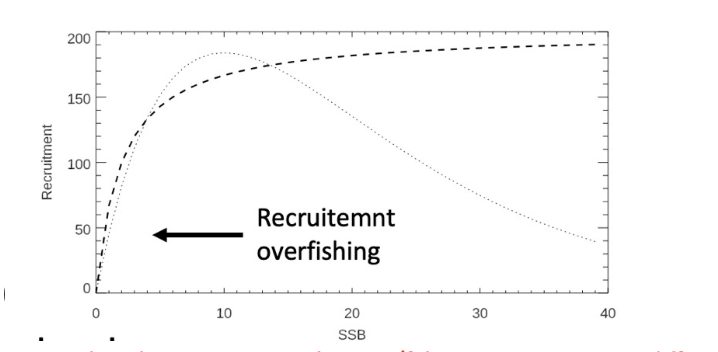
Answer: