(FC) Regulation of Glycolysis to Shuttle Mechanism
Regulation of Glycolysis
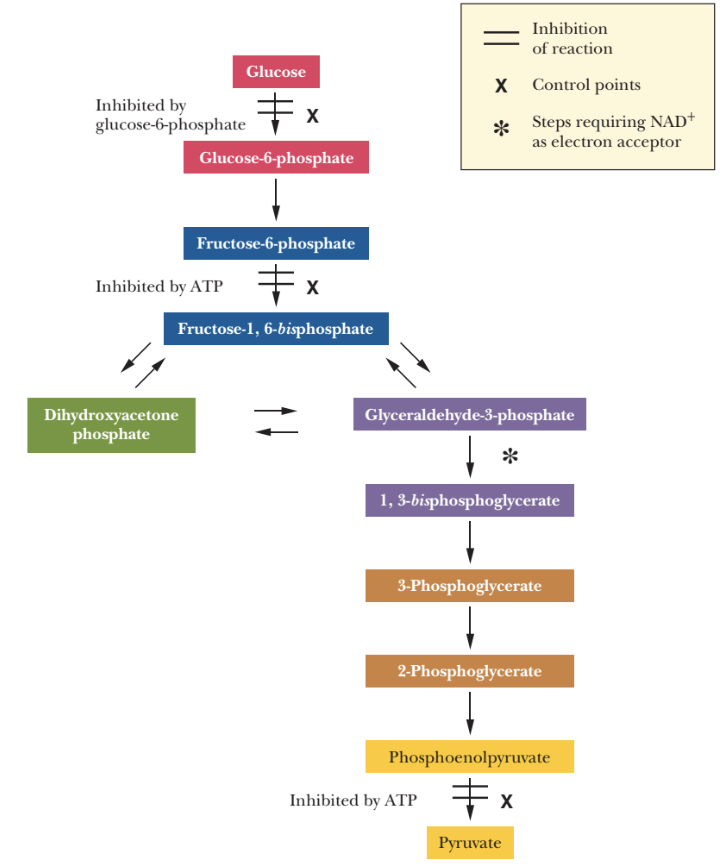
Hexokinase (Step 1)
Phosphofructokinase (Step 3)
Pyruvate kinase (Step 10)
these enzymes catalyze the irreversible reactions
irreversible kinase reactions that primarily drive glycolysis forward
reactions exhibit large decrease in ΔG
Hexokinase (Step 1)
glucose to glucose-6-phosphate
inhibited by the product, glucose-6-phosphate (G6P)
simple inhibition
it would result in slowing down glycolysis if there is an abundant production of G6P
Substrate: can accept any hexose (glucose, galactose and fructose)
exception: in liver, it is specific for glucose (thats why its called glucokinase)
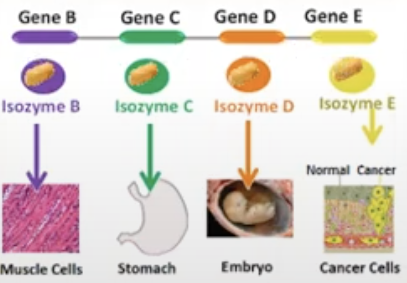
exist as multiple isozymes: I-V
Isozyme
referring to the same enzyme so they catalyze by the same enzyme but is found in different tissues in the body, they are formed from different genes
Phosphofructokinase (PFK)
key regulatory enzyme of glycolysis
rate limiting step
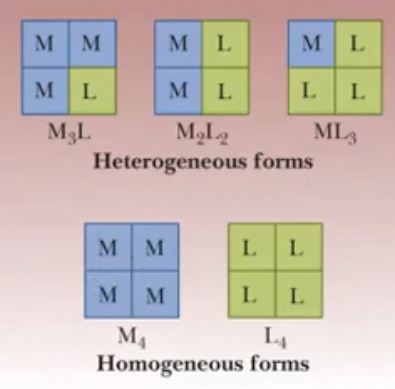
Tetrameric (4 polypeptide chains) isozyme: L & M Units
muscles are rich in M4
liver rich in L4
red blood cells are rich in the combination of 4 units
Allosteric enzyme
it has inhibitors and activatiors
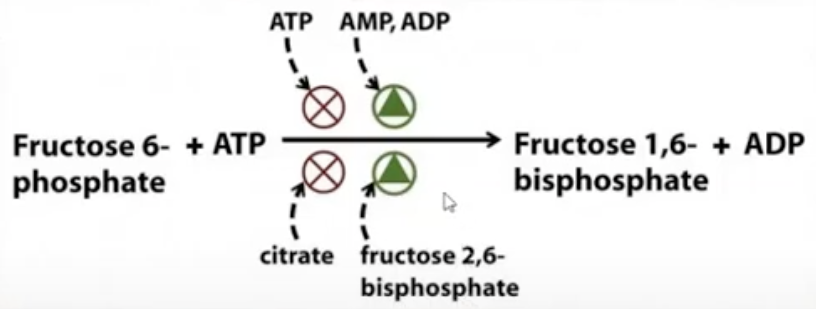
(for the pic)
if we still have a lot of energy present (ATP) then some of the ATP will inhibit PFK
if we lack energy, if ADP and AMP are abundant (low energy), it will act as activators, it will tell PFK to make the reaction faster
Fructose-1,6-bisphosphate will also act as an inhibitor to PFK (if we formed a lot of the fructose-1,6-bisphosphate)
Pyruvate kinase (PK)
Step 10 in glycolysis
Allosteric
inhibitors: ATP and Ala
ATP: If you have a lot of energy, no need to do glycolysis
Ala: can easily be converted to pyruvate
Activators: FBP ("feed forward”)
if we have a lot of FBP, it will tell PK to do the reactions faster, so that FBP will not accumulate in Step 3
If it activates PK, it will inhibit phosphofructosekinase
if we have a lot of FBP, it will tell PK to do the reactions faster but it will tell PFK to stop
activator for PK but an inhibitor for PFK
Tetrameric isozyme: M, L and A units
muscles rich in M
liver rich in L
other tissues rich in A or a combination of the 3
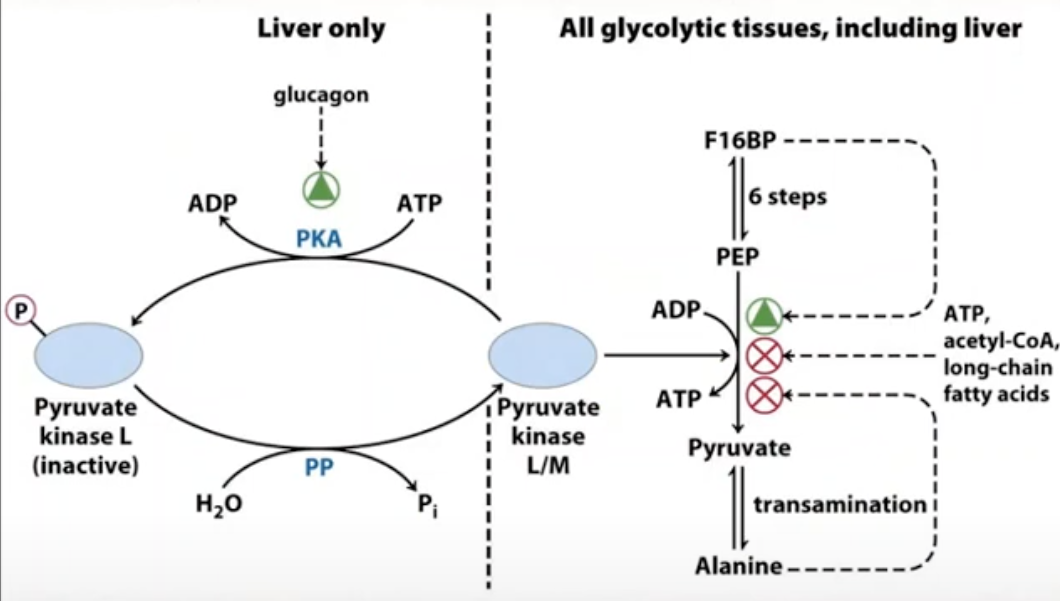
L isozymes in liver
the PK in liver is rich in isozyme
role of liver in PK: regulating blood glucose level at a particular range
low blood glucose triggers production of protein kinase (not PK)
protein kinase is produced if we have low blood glucose
it will deactivate pyruvate kinase
glycolysis stops in liver if blood glucose is low so that the blood glucose will increase
Why is it necessary to maintain blood glucose at a certain level
brain does not have energy storage mechanism of its own so it needs a regular supply of glucose
so if our brain needs to produce energy, it will get glucose from the blood and then do glycolysis
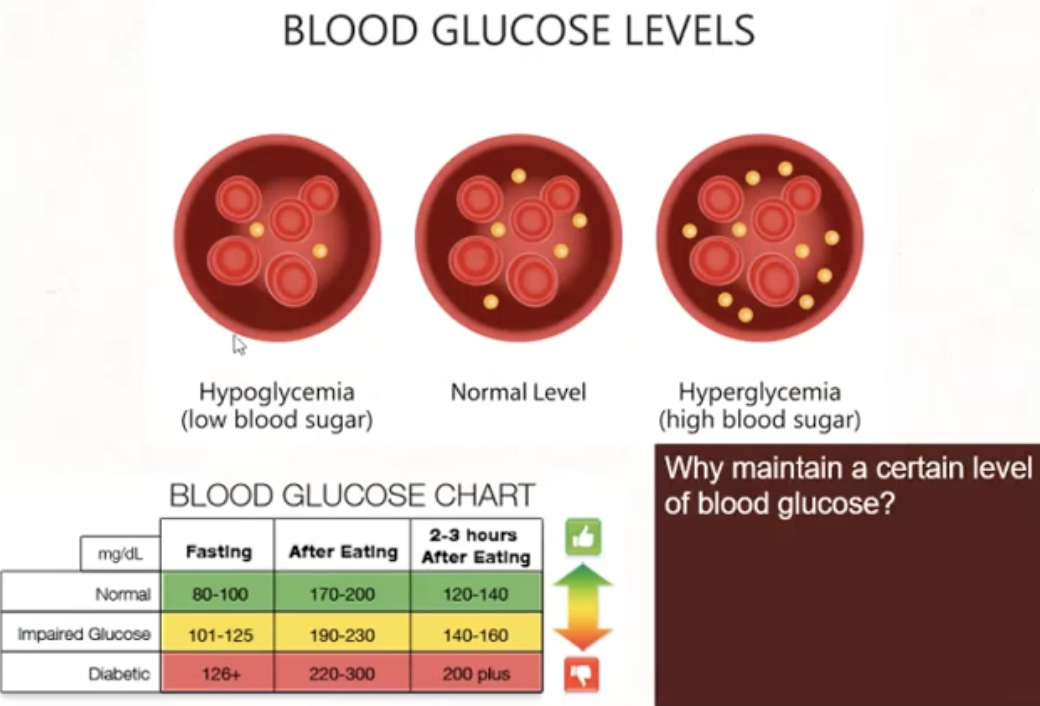
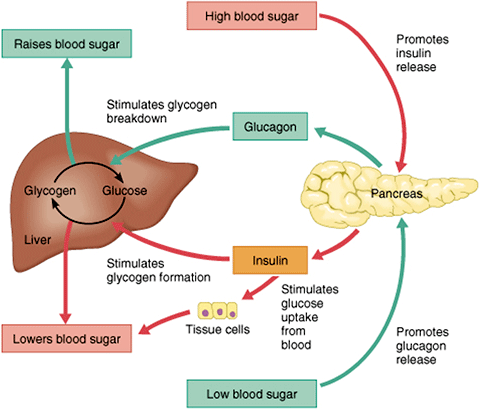
Hormones
to lower blood glucose
Insulin
Somatostatin
to increase blood glucose
Glucagon
Epinephrine
Cortisol
ACTH (adrenocorticotropic hormone)
Growth hormone
Thyroxine
Step 6 of glycolysis produces 2NADH
1 mole glucose = 2 moles NADH
NADH
is a source of the electrons
each mole of NADH, 2.5 ATP is produced
a big molecule so it cannot freely enter or exit the mitochondria
How will the cytosolic NADH transfer it electrons from the cytosol to the mitochondria?
uses shuttle mechanism
Shuttle Mechanism
transport metabolites (i.e. NADH) between mitochondria (ETC and ATP synthesis) and cytosol (glycolysis)
Glycerol-phosphate shuttle
Malate-aspartate shuttle
Glycerol-phosphate shuttle
a mechanism for transferring e- from NADH in the cytosol to FADH2 in the mitochondria
NADH (from glycolysis in cytosol) cant cross the mitochondrial membrane but dihydroxyacetone phosphate and glycerol phosphate can
from the glycerol-phosphate shuttle the NADH was able to transfer its 2 electrons to FAD
per mole of cytosolic NADH only 1.5 ATP is produced
because inside the mitochondria, FAD is the one accepting the electrons
extensively studies in insect flight muscles
also observed in mammalian muscles and the brain
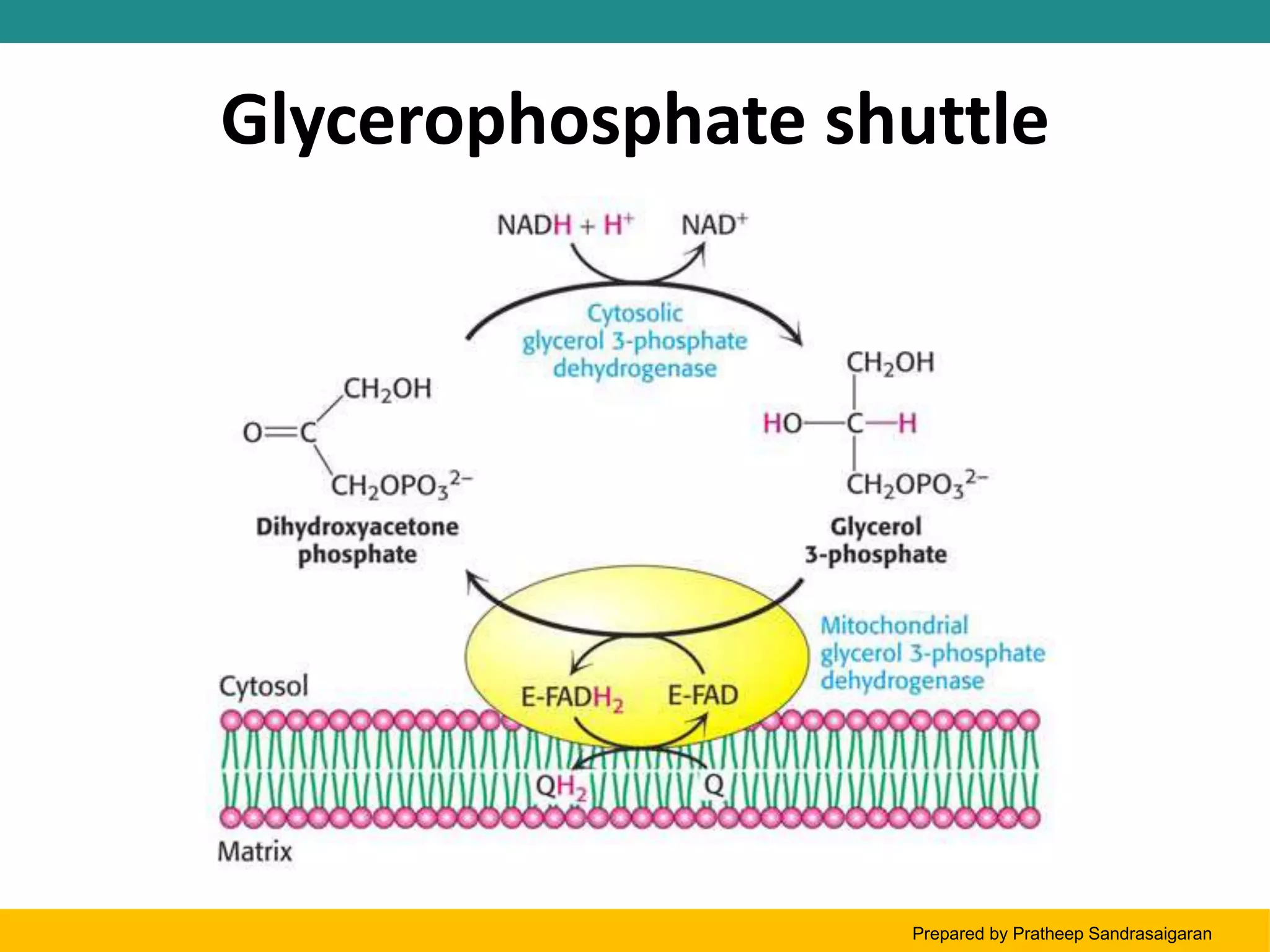
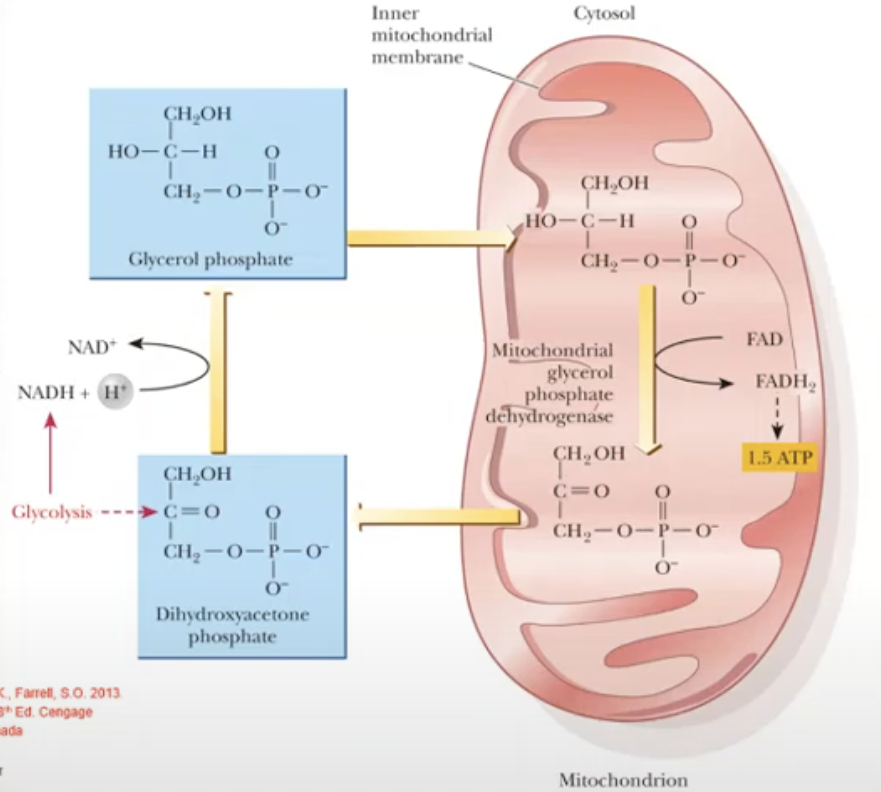
NADH (product from step 6 in glycolysis) will give its electrons to dihydroxyacetone phosphate (DHAP, cytosolic) in the process, DHAP is reduced to produce glycerol phosphate (cytosolic)
Glycerol phosphate (cytosolic) is small so it can easily enter the mitochondrial membrane
Once inside the mitochondria, the glycerol phosphate will give its electrons to FAD and is converted to FADH2, and in the process the glycerol phosphate is converted back to DHAP (mitochondrial)
FADH2 enters the ETC (via complex II) to produce 1.5 ATP
DHAP (mitochondria) it will exit the mitochondria and go to the cytosol to repeat the process
Malate-Aspartate shuttle
a mechanism for transferring e- from NADH in the cytosol to NADH in the mitochondria
oxaloacetate cant ross the mitochondrial membrane but malate can
is more complex but more efficient
for each mole of cytosolic NADH, 2.5 ATP is produced
observed in mammalian kidney, liver and heart
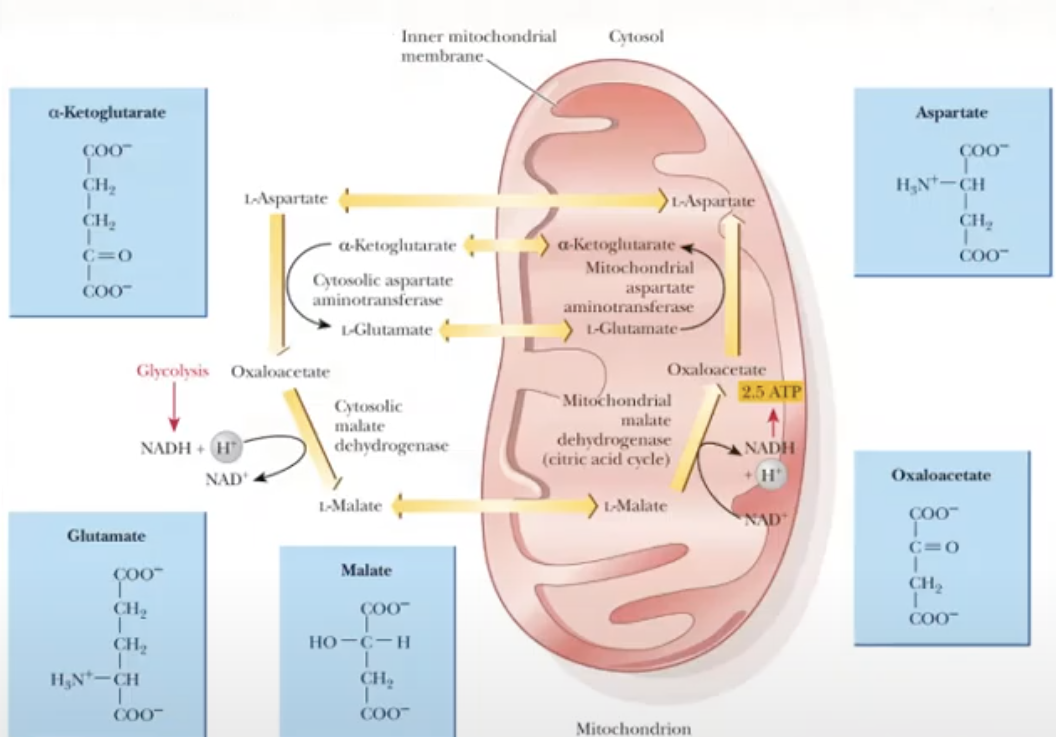
1st step: NADH will give its electrons to oxaloacetate (cytosolic) and will be reduced to malate
malate is a small molecule (4-C) so it can easily cross the mitochondrial membrane
once inside the mitochondrial membrane, malate will give the electrons to NAD+ and will be converted to NADH
L-Malate will be converted to oxaloacetate
NADH will give the electron to ETC via complex I to produce 2.5 ATP
oxaloacetate (mitochondrial) is converted to aspartate (mitochondrial)
aspartate (mitochondrial) can easily exit the mitochondria and will go back to the cytosol
in the cytosol, the aspartate (cytosolic) is converted back to oxaloacetate (cytosolic) to repeat the cycle
2 electrons from the cytosolic NADH was eventually transferred to the mitochondrial NADH